
Inhibition of Recruitment and Activation of Neutrophils by Pyridazinone-Scaffold-Based Compounds

 , , , , , , , and
, , , , , , , and 
Abstract
:1. Introduction
2. Results
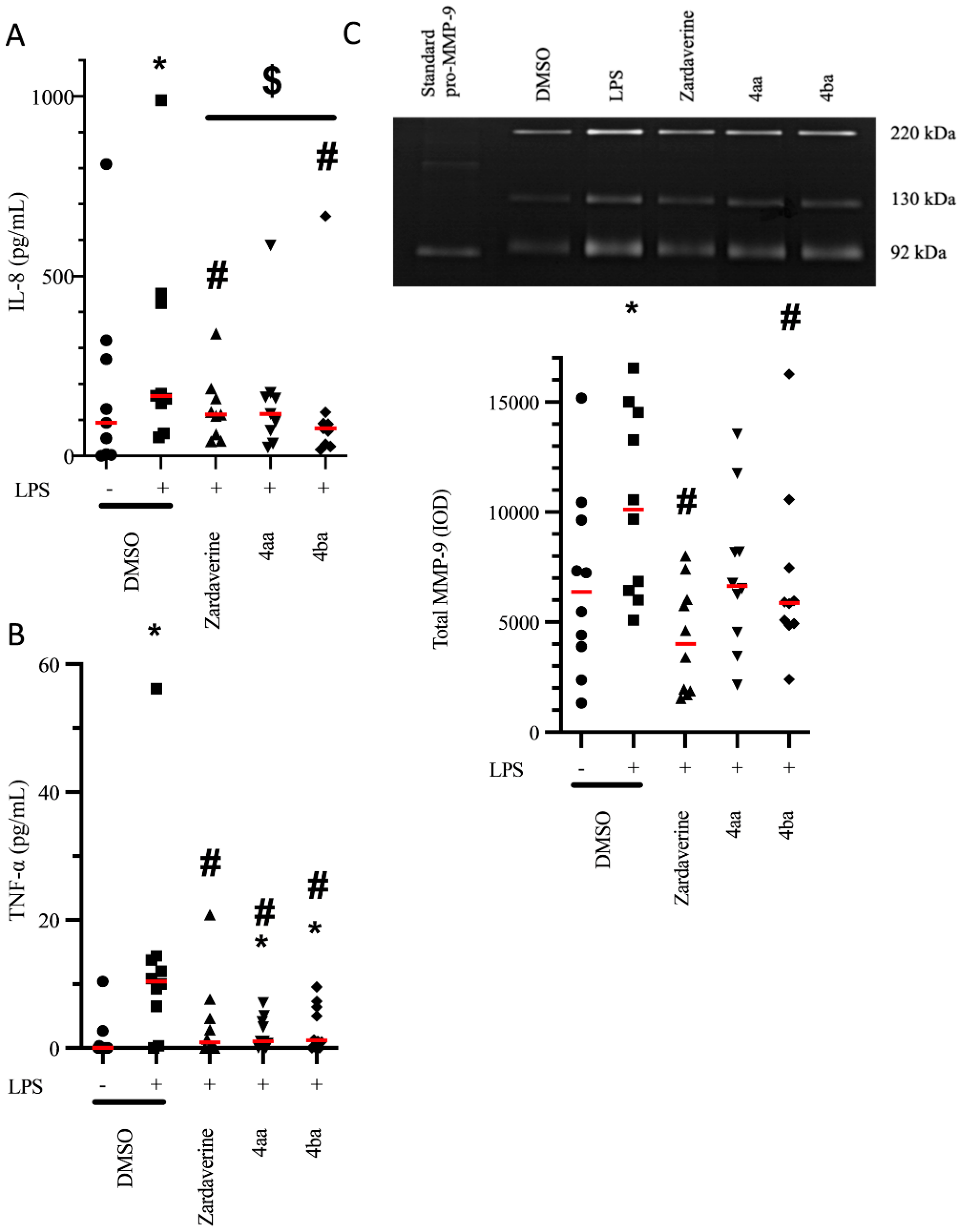
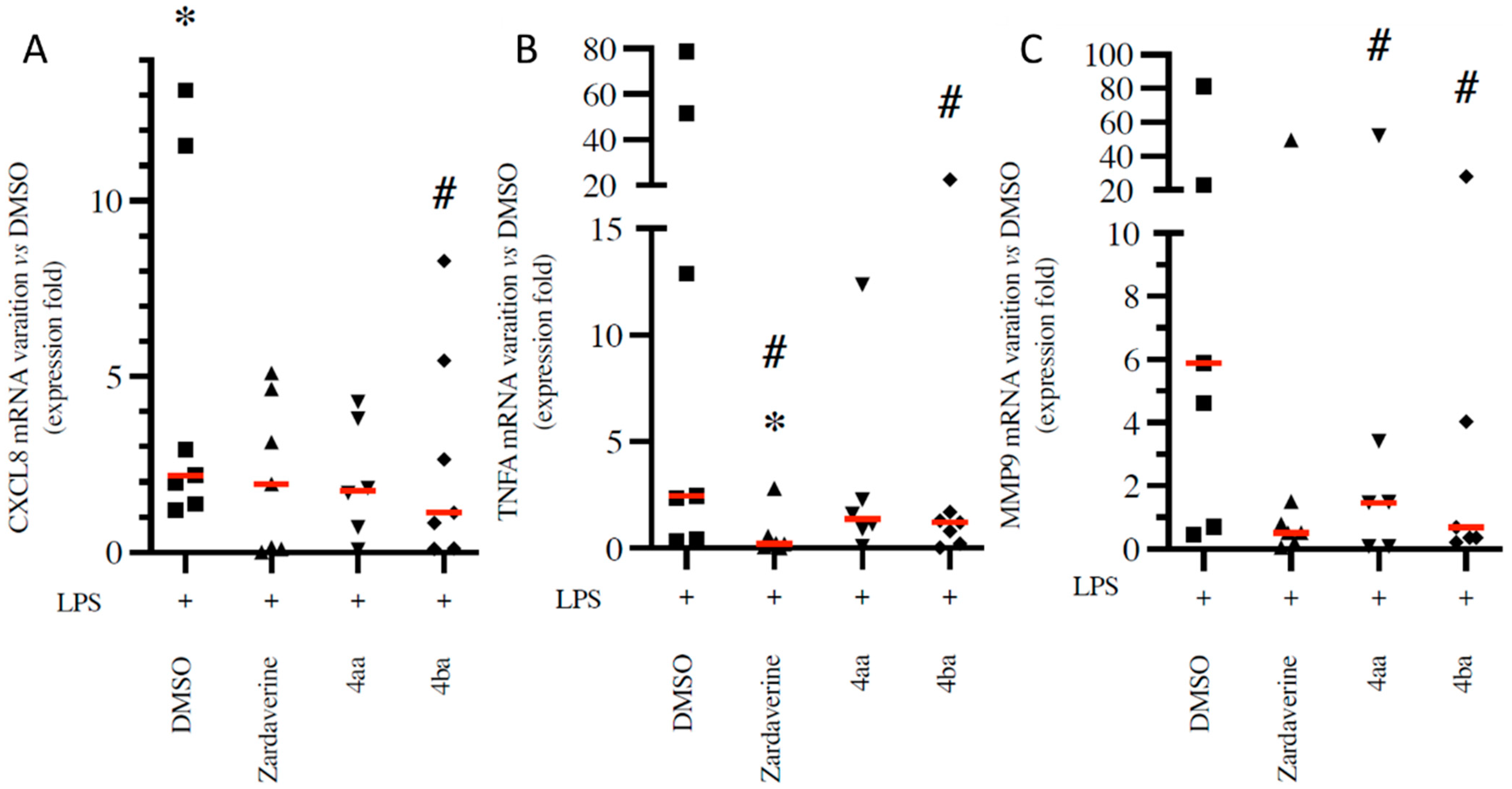
2.1. Pyridazinone-Scaffold-Based Compounds Decrease the Release of Pro-Inflammatory Mediators by Activated PMNs
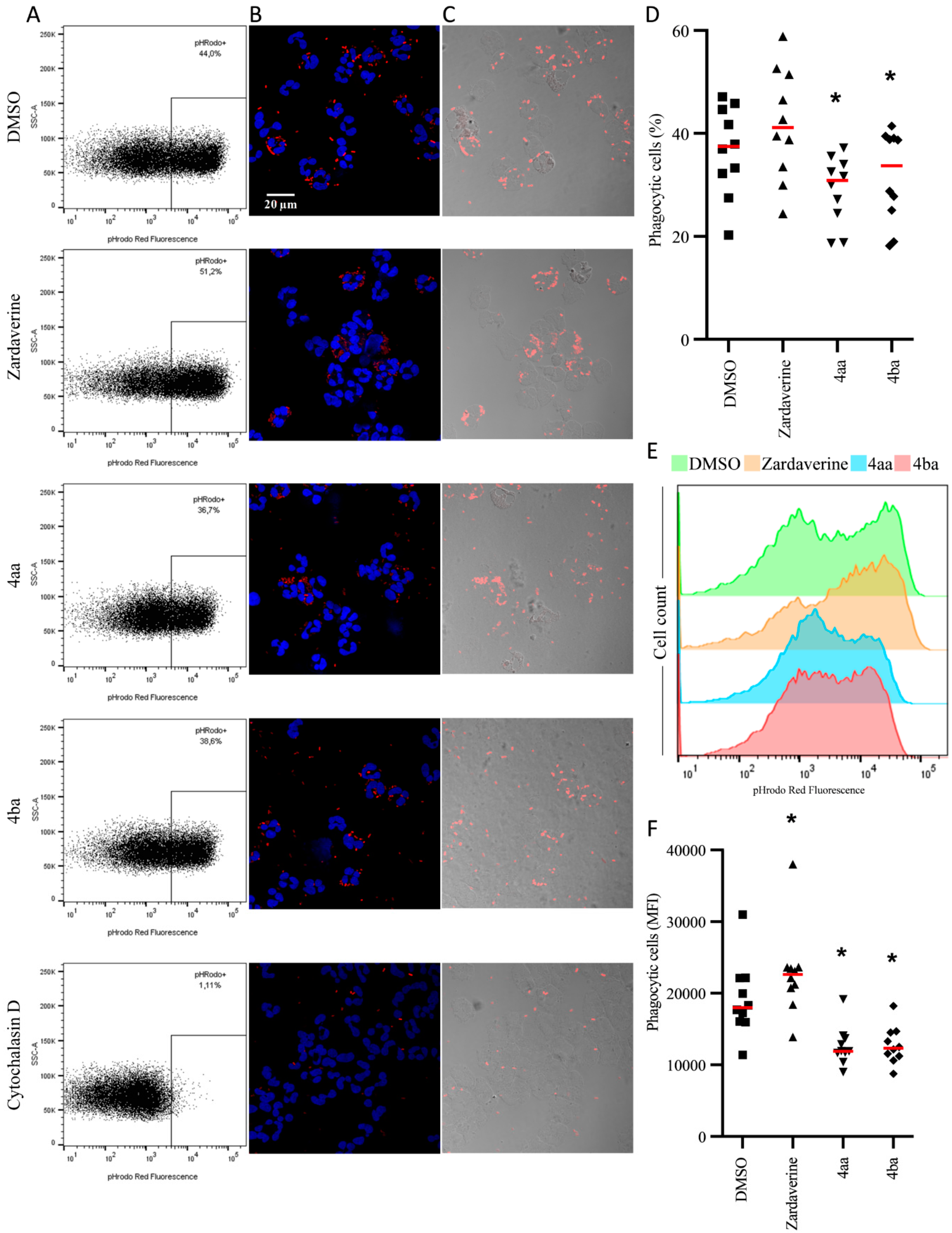
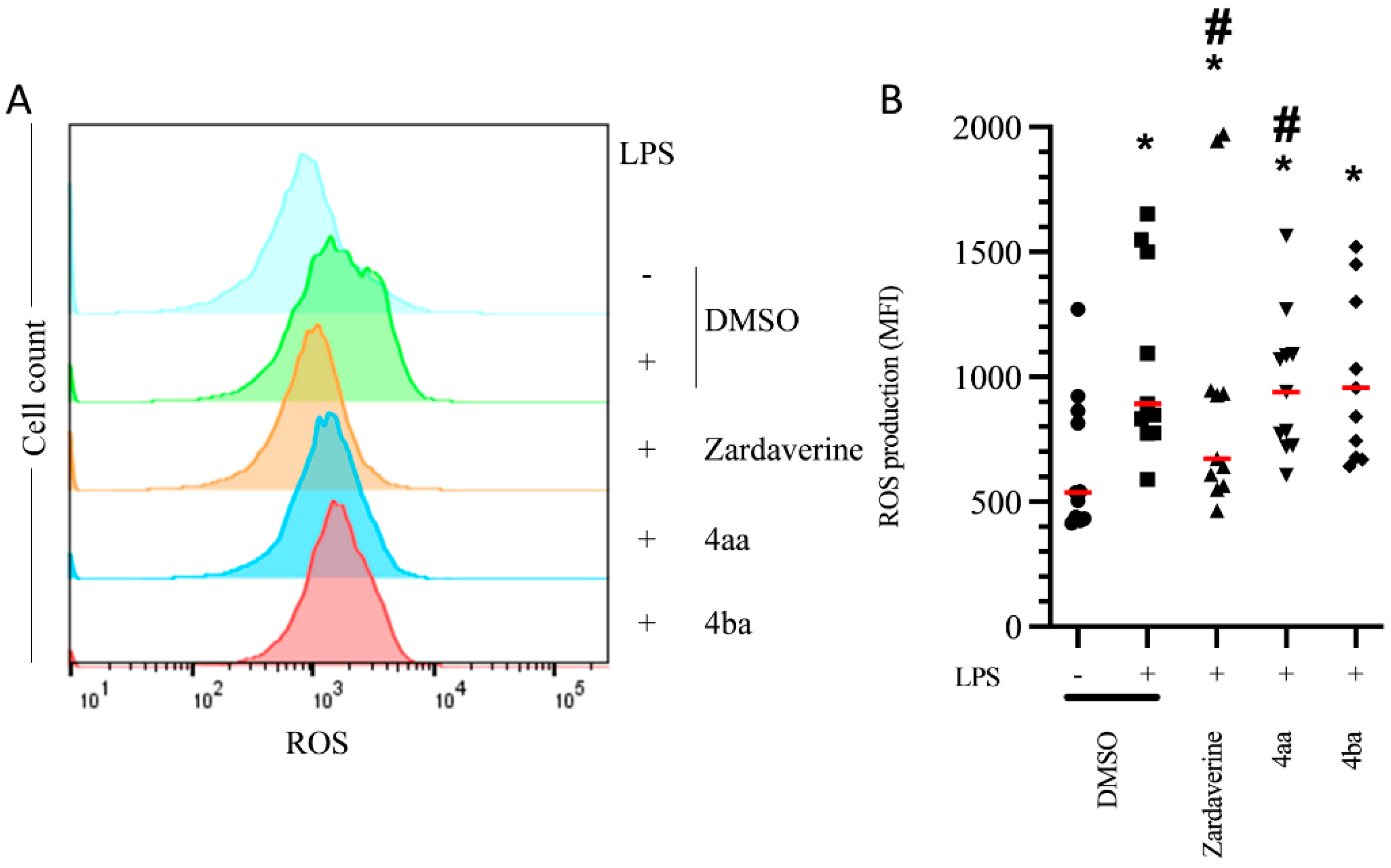
2.2. Pyridazinone-Scaffold-Based Compounds Do Not Impact Beneficial Functions in Activated PMNs
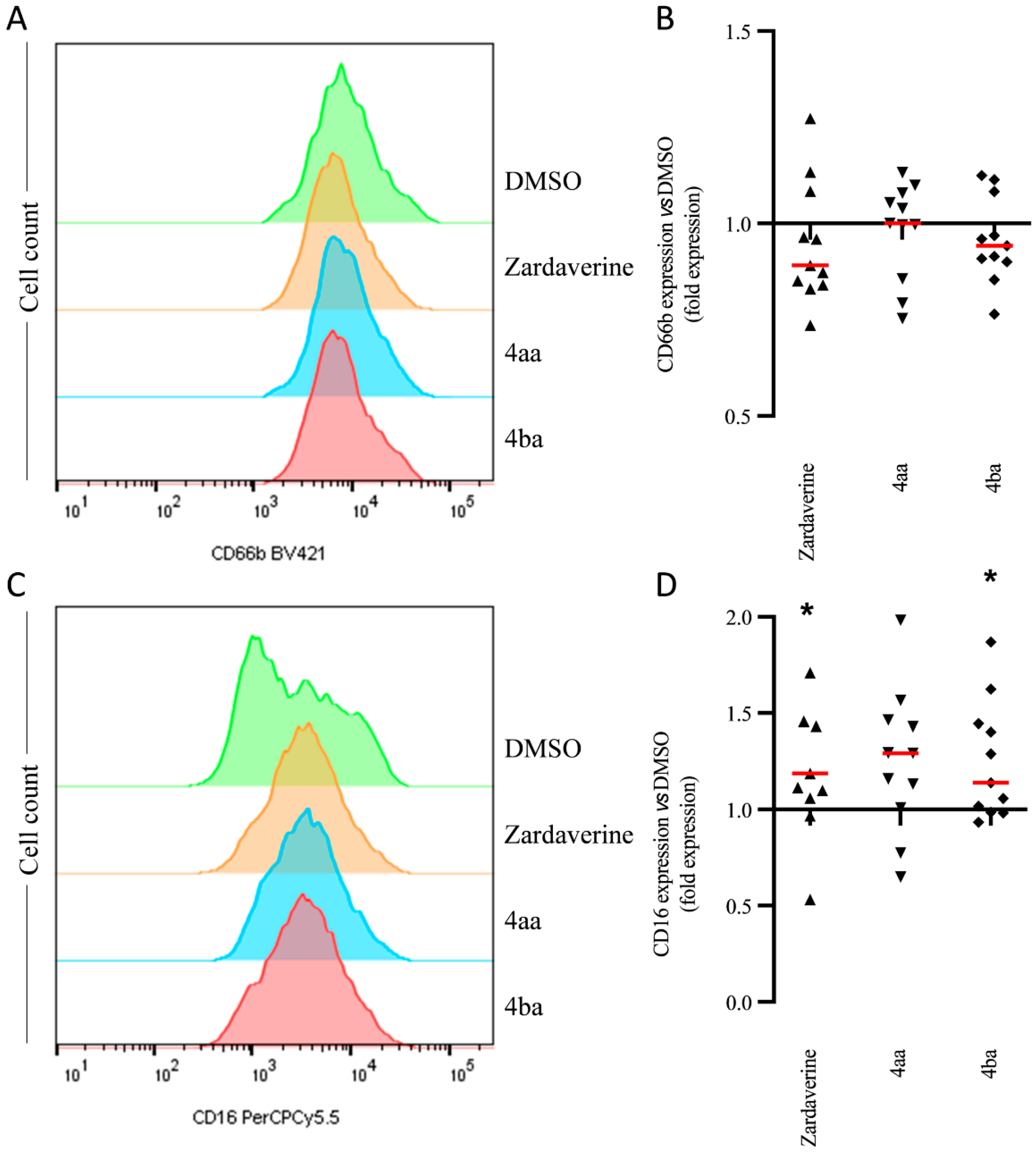
2.3. Pyridazinone-Scaffold-Based Compounds Modulate PMN Phenotype after Transepithelial Migration
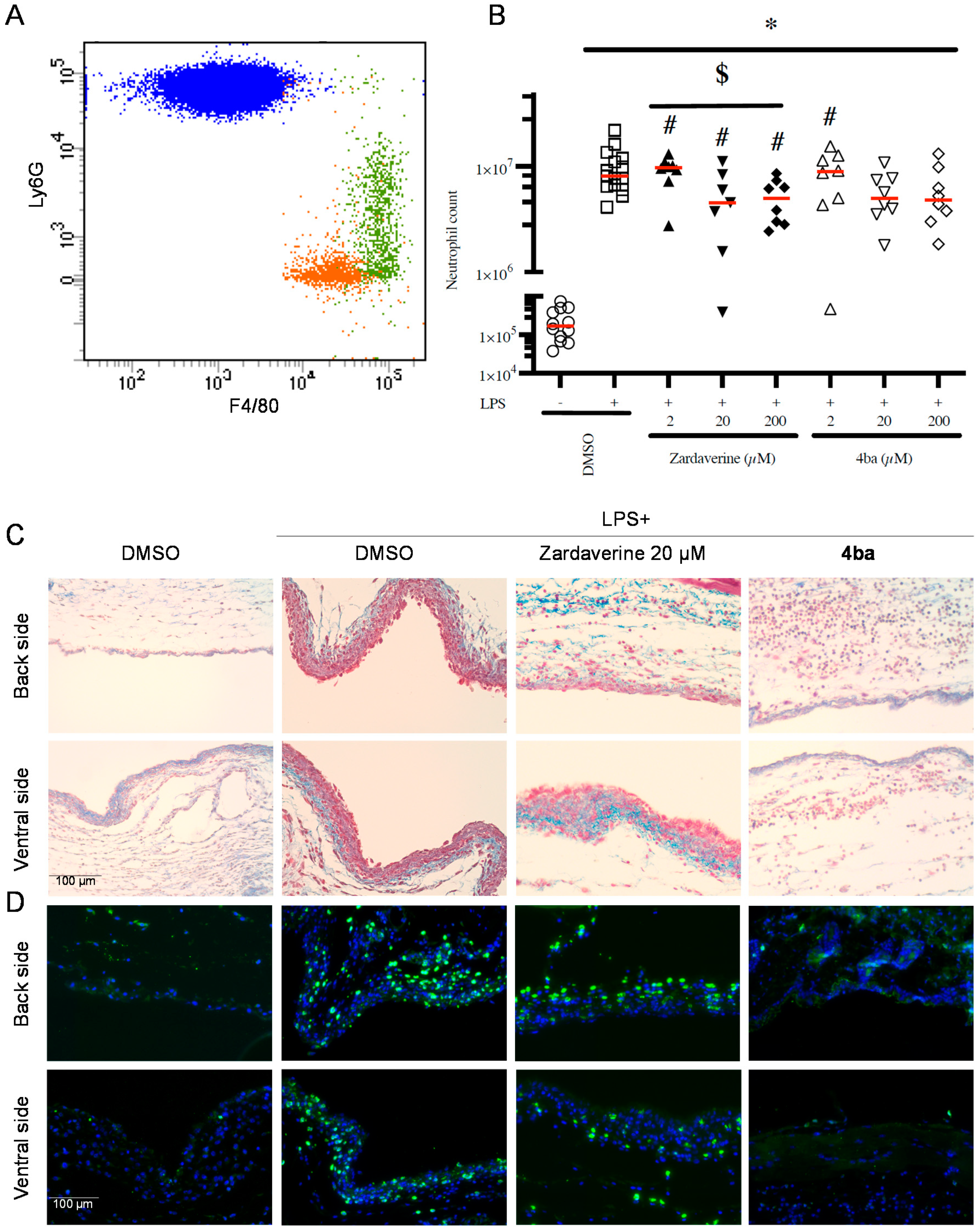
2.4. Pyridazinone-Scaffold-Based Compounds Decrease PMNs Recruitment In Vivo
3. Discussion
4. Materials and Methods
4.1. Media and Reagents
4.2. Compound Preparation
4.3. Collection of Blood Samples, Cell Isolation and Culture
4.4. ELISA
4.5. Gelatin Zymography
4.6. RNA Purification and Reverse-Transcription
4.7. Phagocytosis
4.8. Oxidative Stress Quantification
4.9. Transepithelial Migration
4.10. Air Pouch Model of PMN-Driven Tissue Inflammation
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faurschou, M.; Borregaard, N. Neutrophil granules and secretory vesicles in inflammation. Microbes Infect. 2003, 5, 1317–1327. [Google Scholar] [CrossRef]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The Multifaceted Functions of Neutrophils. Annu. Rev. Pathol. 2014, 9, 181–218. [Google Scholar] [CrossRef] [Green Version]
- Tirouvanziam, R.; Gernez, Y.; Conrad, C.K.; Moss, R.B.; Schrijver, I.; Dunn, C.E.; Davies, Z.A.; Herzenberg, L.A.; Herzenberg, L.A. Profound functional and signaling changes in viable inflammatory neutrophils homing to cystic fibrosis airways. Proc. Natl. Acad. Sci. USA 2008, 105, 4335–4339. [Google Scholar] [CrossRef] [Green Version]
- Witko-Sarsat, V.; Rieu, P.; Descamps-Latscha, B.; Lesavre, P.; Halbwachs-Mecarelli, L. Neutrophils: Molecules, functions and pathophysiological aspects. Lab. Investig. 2000, 80, 617–653. [Google Scholar] [CrossRef] [Green Version]
- Dransfield, I.; Buckle, A.M.; Savill, J.S.; McDowall, A.; Haslett, C.; Hogg, N. Neutrophil apoptosis is associated with a reduction in CD16 (Fc gamma RIII) expression. J. Immunol. 1994, 153, 1254–1263. [Google Scholar]
- Lee, W.L.; Harrison, R.E.; Grinstein, S. Phagocytosis by neutrophils. Microbes Infect. 2003, 5, 1299–1306. [Google Scholar] [CrossRef]
- Ingersoll, S.A.; Laval, J.; Forrest, O.A.; Preininger, M.; Brown, M.R.; Arafat, D.; Gibson, G.; Tangpricha, V.; Tirouvanziam, R. Mature cystic fibrosis airway neutrophils suppress T cell function: Evidence for a role of arginase 1 but not programmed death-ligand 1. J. Immunol. 2015, 194, 5520–5528. [Google Scholar] [CrossRef]
- Lacy, P. Mechanisms of degranulation in neutrophils. Allergy Asthma Clin. Immunol. 2006, 2, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Mallia, P.; Message, S.D.; Contoli, M.; Gray, K.K.; Telcian, A.; Laza-Stanca, V.; Papi, A.; Stanciu, L.A.; Elkin, S.; Kon, O.M.; et al. Neutrophil adhesion molecules in experimental rhinovirus infection in COPD. Respir. Res. 2013, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Baggiolini, M.; Clark-Lewis, I. Interleukin-8, a chemotactic and inflammatory cytokine. FEBS Lett. 1992, 307, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.C.; Garcia, C.C.; Teixeira, M.M.; Amaral, F.A. The CXCL8/IL-8 chemokine family and its receptors in inflammatory diseases. Expert Rev. Clin. Immunol. 2014, 10, 593–619. [Google Scholar] [CrossRef] [Green Version]
- Beyrau, M.; Bodkin, J.V.; Nourshargh, S. Neutrophil heterogeneity in health and disease: A revitalized avenue in inflammation and immunity. Open Biol. 2012, 2, 120134. [Google Scholar] [CrossRef] [Green Version]
- Liles, W.C.; Dale, D.C.; Klebanoff, S.J. Glucocorticoids inhibit apoptosis of human neutrophils. Blood 1995, 86, 3181–3188. [Google Scholar] [CrossRef] [Green Version]
- Ronchetti, S.; Ricci, E.; Migliorati, G.; Gentili, M.; Riccardi, C. How Glucocorticoids Affect the Neutrophil Life. Int. J. Mol. Sci. 2018, 19, 4090. [Google Scholar] [CrossRef] [Green Version]
- Schudt, C.; Winder, S.; Forderkunz, S.; Hatzelmann, A.; Ullrich, V. Influence of selective phosphodiesterase inhibitors on human neutrophil functions and levels of cAMP and Cai. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1991, 344, 682–690. [Google Scholar] [CrossRef]
- Ariga, M.; Neitzert, B.; Nakae, S.; Mottin, G.; Bertrand, C.; Pruniaux, M.P.; Jin, S.L.C.; Conti, M. Non redundant function of phosphodiesterases 4D and 4B in neutrophil recruitment to the site of inflammation. J. Immunol. 2004, 173, 7531–7538. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.L.; Ding, S.L.; Lin, S.C. Phosphodiesterase 4 and its inhibitors in inflammatory diseases. Chang Gung Med. J. 2012, 35, 197–210. [Google Scholar] [CrossRef]
- Wedzicha, J.A.; Calverley, P.M.; Rabe, K.F. Roflumilast: A review of its use in the treatment of COPD. Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.F.; Lv, X.D.; Chen, W.Y.; Yang, Q.; Fang, Z.X.; Lu, W.F. Effect of roflumilast on chronic obstructive pulmonary disease: A systematic review and meta-analysis. Ir. J. Med. Sci. 2018, 187, 731–738. [Google Scholar] [CrossRef]
- Gupta, S. Side-effects of roflumilast. Lancet 2012, 379, 710–711. [Google Scholar] [CrossRef]
- Barberot, C.; Moniot, A.; Allart-Simon, I.; Malleret, L.; Yegorova, T.; Laronze-Cochard, M.; Bentaher, A.; Médebielle, M.; Bouillon, J.P.; Hénon, E.; et al. Synthesis and biological evaluation of pyridazinone derivatives as potential anti-inflammatory agents. Eur. J. Med. Chem. 2018, 146, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Allart-Simon, I.; Moniot, A.; Bisi, N.; Ponce-Vargas, M.; Audonnet, S.; Laronze-Cochard, M.; Sapi, J.; Hénon, E.; Velard, F.; Gérard, S. Pyridazinone derivatives as Potential Anti-inflammatory Agents: Synthesis and Biological Evaluation as PDE4 inhibitors. RSC Med. Chem. 2021, 12, 584. [Google Scholar] [CrossRef] [PubMed]
- Forrest, O.A.; Ingersoll, S.A.; Preininger, M.K.; Laval, J.; Limoli, D.H.; Brown, M.R.; Lee, F.E.; Bedi, B.; Sadikot, R.T.; Goldberg, J.B.; et al. Frontline Science: Pathological conditioning of human neutrophils recruited to the airway milieu in cystic fibrosis. J. Leukoc. Biol. 2018, 104, 665–675. [Google Scholar] [CrossRef]
- Cassatella, M.A. The production of cytokines by polymorphonuclear neutrophils. Immunol. Today 1995, 16, 21–26. [Google Scholar] [CrossRef]
- Hilda, J.N.; Das, S.D. TLR stimulation of human neutrophils lead to increased release of MCP-1, MIP-1α, IL-1β, IL-8 and TNF during tuberculosis. Hum. Immunol. 2016, 77, 63–67. [Google Scholar] [CrossRef]
- Huber, A.R.; Kunkel, S.L.; Todd, R.F.; Weiss, S.J. Regulation of transendothelial neutrophil migration by endogenous interleukin-8. Science 1991, 254, 99–102. [Google Scholar] [CrossRef]
- Henkels, K.M.; Frondorf, K.; Gonzalez-Mejia, M.E.; Doseff, A.L.; Gomez-Cambronero, J. IL-8-induced neutrophil chemotaxis is mediated by Janus kinase 3 (JAK3). FEBS Lett. 2011, 585, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.C.; Cheng, W.J.; Korinek, M.; Lin, C.Y.; Hwang, T.L. Neutrophils in Psoriasis. Front. Immunol. 2019, 10, 2376. [Google Scholar] [CrossRef]
- Cecchi, I.; Arias de la Rosa, I.; Menegatti, E.; Roccatello, D.; Collantes-Estevez, E.; Lopez-Pedrera, C.; Barbarroja, N. Neutrophils: Novel key players in Rheumatoid Arthritis. Current and future therapeutic targets. Autoimmun. Rev. 2018, 17, 1138–1149. [Google Scholar] [CrossRef]
- Gao, H.; Ying, S.; Dai, Y. Pathological Roles of Neutrophil-Mediated Inflammation in Asthma and Its Potential for Therapy as a Target. J. Immunol. Res. 2017, 2017, 3743048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasper, A.E.; McIver, W.J.; Sapey, E.; Walton, G.M. Understanding the role of neutrophils in chronic inflammatory airway disease. F1000Res 2019, 26, 8. [Google Scholar] [CrossRef]
- Milara, J.; Lluch, J.; Almudever, P.; Freire, J.; Xiaozhong, Q.; Cortijo, J. Roflumilast N-oxide reverses corticosteroid resistance in neutrophils from patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2014, 134, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, D.W.; Lawrence, T.; Perretti, M.; Rossi, A.G. Inflammatory resolution: New opportunities for drug discovery. Nat. Rev. Drug Discov. 2004, 3, 401–416. [Google Scholar] [CrossRef] [Green Version]
- Fox, S.; Leitch, A.E.; Duffin, R.; Haslett, C.; Rossi, A.G. Neutrophil apoptosis: Relevance to the innate immune response and inflammatory disease. J. Innate Immun. 2010, 2, 216–227. [Google Scholar] [CrossRef] [Green Version]
- Zelová, H.; Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef]
- Houghton, A.M. Matrix metalloproteinases in destructive lung disease. Matrix Biol. 2015, 44, 167–174. [Google Scholar] [CrossRef]
- Genschmer, K.R.; Russell, D.W.; Lal, C.; Szul, T.; Bratcher, P.E.; Noerager, B.D.; Roda, M.A.; Xu, X.; Rezonzew, R.; Viera, L.; et al. Activated PMN Exosomes: Pathogenic Entities Causing Matrix Destruction and Disease in the Lung. Cell 2019, 176, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Delclaux, C.; Delacourt, C.; D’Ortho, M.P.; Boyer, V.; Lafuma, C.; Harf, A. Role of gelatinase B and elastase in human polymorphonuclear neutrophil migration across basement membrane. Am. J. Respir. Cell Mol. Biol. 1996, 14, 288–295. [Google Scholar] [CrossRef]
- Bradley, L.M.; Douglass, M.F.; Chatterjee, D.; Akira, S.; Baaten, B.J. Matrix metalloprotease 9 mediates neutrophil migration into the airways in response to influenza virus-induced toll-like receptor signaling. PLoS Pathog. 2012, 8, e1002641. [Google Scholar] [CrossRef] [Green Version]
- Geurts, N.; Martens, E.; Van Aelst, I.; Proost, P.; Opdenakker, G.; Van den Steen, P.E. Beta-hematin interaction with the hemopexin domain of gelatinase B/MMP-9 provokes autocatalytic processing of the propeptide, thereby priming activation by MMP-3. Biochemistry 2008, 47, 2689–2699. [Google Scholar] [CrossRef]
- Zalavary, S.; Bengtsson, T. Adenosine inhibits actin dynamics in human neutrophils: Evidence for the involvement of cAMP. Eur. J. Cell Biol. 1998, 7, 128–139. [Google Scholar] [CrossRef]
- Soares, A.C.; Souza, D.G.; Pinho, V.; Vieira, A.T.; Barsante, M.M.; Nicoli, J.R.; Teixeira, M. Impaired host defense to Klebsiella pneumoniae infection in mice treated with the PDE4 inhibitor rolipram. Br. J. Pharmacol. 2003, 140, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Dupré-Crochet, S.; Erard, M.; Nüβe, O. ROS production in phagocytes: Why, when, and where? J. Leukoc. Biol. 2013, 94, 657–670. [Google Scholar] [CrossRef]
- Koga, H.; Recke, A.; Vidarsson, G.; Pas, H.H.; Jonkman, M.F.; Hashimoto, T.; Kasprick, A.; Ghorbanalipoor, S.; Tenor, H.; Zillikens, D.; et al. PDE4 Inhibition as Potential Treatment of Epidermolysis Bullosa Acquisita. J. Investig. Dermatol. 2016, 136, 2211–2220. [Google Scholar] [CrossRef] [Green Version]
- Butcher, S.K.; Chahal, H.; Nayak, L.; Sinclair, A.; Henriquez, N.V.; Sapey, E.; O’Mahony, D.; Lord, J.M. Senescence in innate immune responses: Reduced neutrophil phagocytic capacity and CD16 expression in elderly humans. J. Leukoc. Biol. 2001, 70, 881–886. [Google Scholar]
- Kim, S.; Nadel, J.A. Role of neutrophils in mucus hypersecretion in COPD and implications for therapy. Treat. Respir. Med. 2004, 3, 147–159. [Google Scholar] [CrossRef]
- Chmiel, J.F.; Aksamit, T.R.; Chotirmall, S.H.; Dasenbrook, E.C.; Elborn, J.S.; LiPuma, J.J.; Ranganathan, S.C.; Waters, V.J.; Ratjen, F.A. Antibiotic management of lung infections in cystic fibrosis. I. The microbiome, methicillin-resistant Staphylococcus aureus, gram-negative bacteria, and multiple infections. Ann. Am. Thorac. Soc. 2014, 11, 1120–1129. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.W.; Stevens, A.J.; Brennan, B.S.; Davies, D.; Rowland, M.; Houston, J.B. The six-day-old rat air pouch model of inflammation: Characterization of the inflammatory response to carrageenan. J. Pharmacol. Toxicol. Methods 1994, 32, 139–147. [Google Scholar] [CrossRef]
- Duarte, D.B.; Vasko, M.R.; Fehrenbacher, J.C. Models of Inflammation: Carrageenan Air Pouch. Curr. Protoc. Pharmacol. 2016, 72, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Hsu, C.Y.; Elzoghby, A.O.; Alalaiwe, A.; Hwang, T.L.; Fang, J.Y. Oleic acid as the active agent and lipid matrix in cilomilast-loaded nanocarriers to assist PDE4 inhibition of activated neutrophils for mitigating psoriasis-like lesions. Acta Biomater. 2019, 90, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Kosutova, P.; Mikolka, P.; Kolomaznik, M.; Balentova, S.; Adamkov, M.; Calkovska, A.; Mokra, D. Reduction of lung inflammation, oxidative stress and apoptosis by the PDE4 inhibitor roflumilast in experimental model of acute lung injury. Physiol. Res. 2018, 67, S645–S654. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Sato, S.; Yamamoto, T.; Ishikawa, S.; Onizuka, M.; Sakakibara, Y. Phosphodiesterase type 4 inhibitor reduces the retention of polymorphonuclear leukocytes in the lung. Am. J. Physiol. Lung. Cell Mol. Physiol. 2002, 282, L1376–L1381. [Google Scholar] [CrossRef] [Green Version]
- Konrad, F.; Bury, A.; Schick, M.A.; Ngamsri, K.C.; Reutershan, J. The unrecognized effects of phosphodiesterase 4 on epithelial cells in pulmonary inflammation. PLoS ONE 2015, 10, e0121725. [Google Scholar] [CrossRef] [Green Version]
- Dunne, A.E.; Kawamatawong, T.; Fenwick, P.S.; Davies, C.M.; Tullett, H.; Barnes, P.J.; Donnelly, L.E. Direct Inhibitory Effect of the PDE4 Inhibitor Roflumilast on Neutrophil Migration in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Cell Mol. Biol. 2019, 60, 445–453. [Google Scholar] [CrossRef]
- Totani, L.; Piccoli, A.; Dell’Elba, G.; Concetta, A.; Di Santo, A.; Martelli, N.; Federico, L.; Pamuklar, Z.; Smyth, S.S.; Evangelista, V. Phosphodiesterase type 4 blockade prevents platelet-mediated neutrophil recruitment at the site of vascular injury. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1689–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gérard, S.; Raoul, M.; Sapi, J. A New Approach to the Synthesis of N-Alkylated 2-Substituted Azetidin-3-ones. Eur. J. Org. Chem. 2006, 10, 2440–2445. [Google Scholar] [CrossRef]
- Bartoli, G.; Bosco, M.; Carlone, A.; Pesciaioli, F.; Sambri, L.; Melchiorre, P. Organocatalytic asymmetric Friedel-Crafts alkylation of indoles with simple alpha,beta-unsaturated ketones. Org. Lett. 2007, 9, 1403–1405. [Google Scholar] [CrossRef]
- Hong, L.; Sun, W.; Liu, C.; Wang, L.; Wong, K.; Wang, R. Enantioselective Friedel–Crafts Alkylation of 4,7-Dihydroindoles with Enones Catalyzed by Primary–Secondary Diamines. Chem. Eur. J. 2009, 15, 11105–11108. [Google Scholar] [CrossRef]
- Adachi, S.; Tanaka, F.; Watanabe, K.; Harada, T. Oxazaborolidinone-Catalyzed Enantioselective Friedel−Crafts Alkylation of Furans and Indoles with α,β-Unsaturated Ketones. Org. Lett. 2009, 11, 5206–5209. [Google Scholar] [CrossRef]
- Schwartz, J.D.; Monea, S.; Marcus, S.G.; Patel, S.; Eng, K.; Galloway, A.C.; Mignatti, P.; Shamamian, P. Soluble factor(s) released from neutrophils activates endothelial cell matrix metalloproteinase-2. J. Surg. Res. 1998, 76, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Ardi, V.C.; Kupriyanova, T.A.; Deryugina, E.I.; Quigley, J.P. Human neutrophils uniquely release TIMP-free MMP-9 to provide a potent catalytic stimulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 20262–20267. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Sequences | Efficiency | |
|---|---|---|---|
| Forward Primer (5′⟶3′) | Reverse Primer (5′⟶3′) | ||
| MMP9 | GAACCAATCTCACCGACAGG | GCCACCCGAGTGTAACCATA | 1.94 |
| CXCL8 | AGACAGCAGAGCACACAAGC | CTCCTTGGCAAAACTGCAC | 1.94 |
| TNF | CAGCCTCTTCTCCTTCCTGAT | GCCAGAGGGCTGATTAGAGA | 1.95 |
| RPS18 | TGCGAGTACTCAACACCAACA | GCATATCTTCGGCCCACA | 1.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moniot, A.; Braux, J.; Siboni, R.; Guillaume, C.; Audonnet, S.; Allart-Simon, I.; Sapi, J.; Tirouvanziam, R.; Gérard, S.; Gangloff, S.C.; et al. Inhibition of Recruitment and Activation of Neutrophils by Pyridazinone-Scaffold-Based Compounds. Int. J. Mol. Sci. 2022, 23, 7226. https://doi.org/10.3390/ijms23137226
Moniot A, Braux J, Siboni R, Guillaume C, Audonnet S, Allart-Simon I, Sapi J, Tirouvanziam R, Gérard S, Gangloff SC, et al. Inhibition of Recruitment and Activation of Neutrophils by Pyridazinone-Scaffold-Based Compounds. International Journal of Molecular Sciences. 2022; 23(13):7226. https://doi.org/10.3390/ijms23137226
Chicago/Turabian StyleMoniot, Aurélie, Julien Braux, Renaud Siboni, Christine Guillaume, Sandra Audonnet, Ingrid Allart-Simon, Janos Sapi, Rabindra Tirouvanziam, Stéphane Gérard, Sophie C. Gangloff, and et al. 2022. "Inhibition of Recruitment and Activation of Neutrophils by Pyridazinone-Scaffold-Based Compounds" International Journal of Molecular Sciences 23, no. 13: 7226. https://doi.org/10.3390/ijms23137226