
Evolutionary Analysis and Functional Identification of Ancient Brassinosteroid Receptors in Ceratopteris richardii

 , , and
, , and 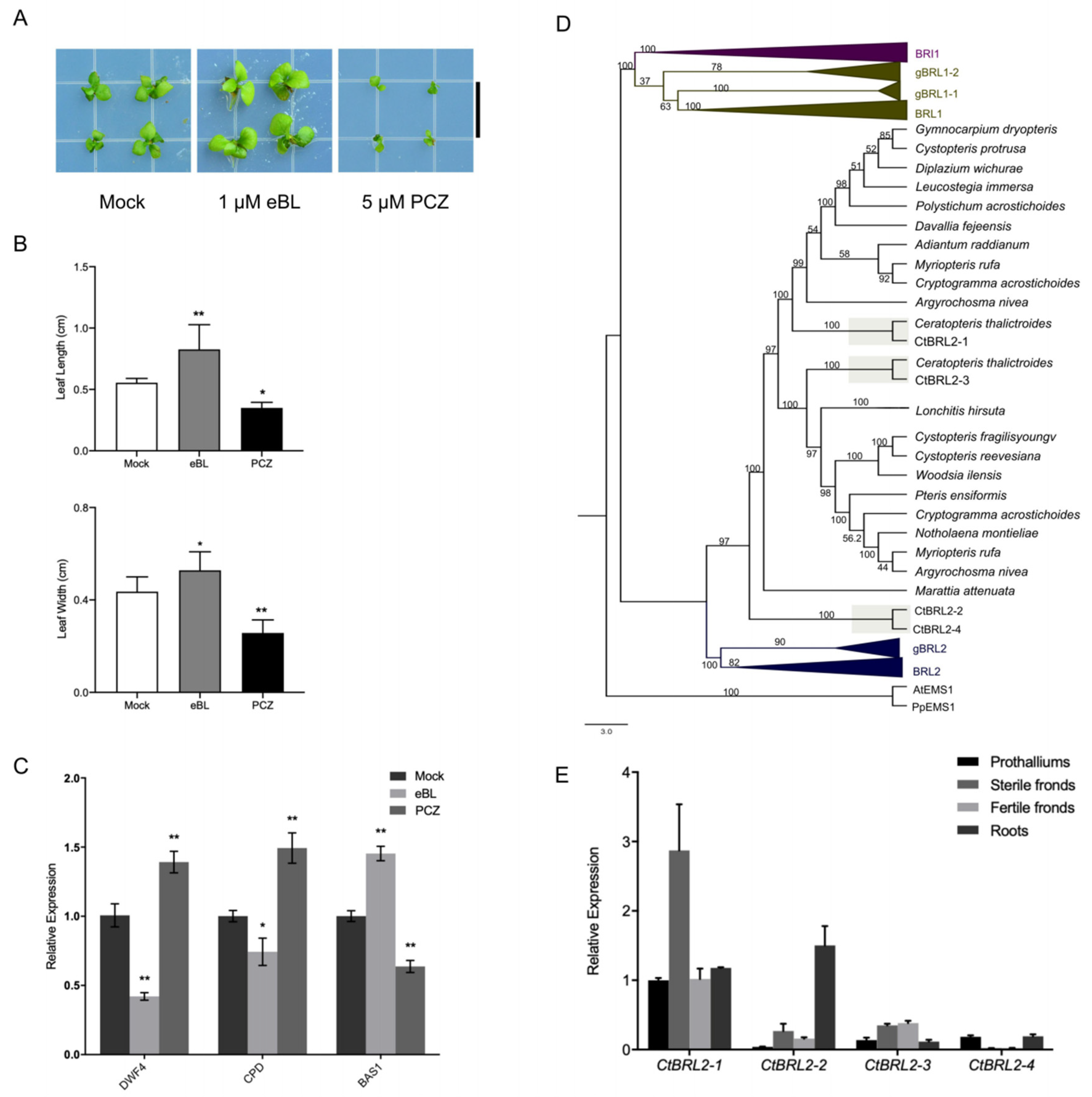{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
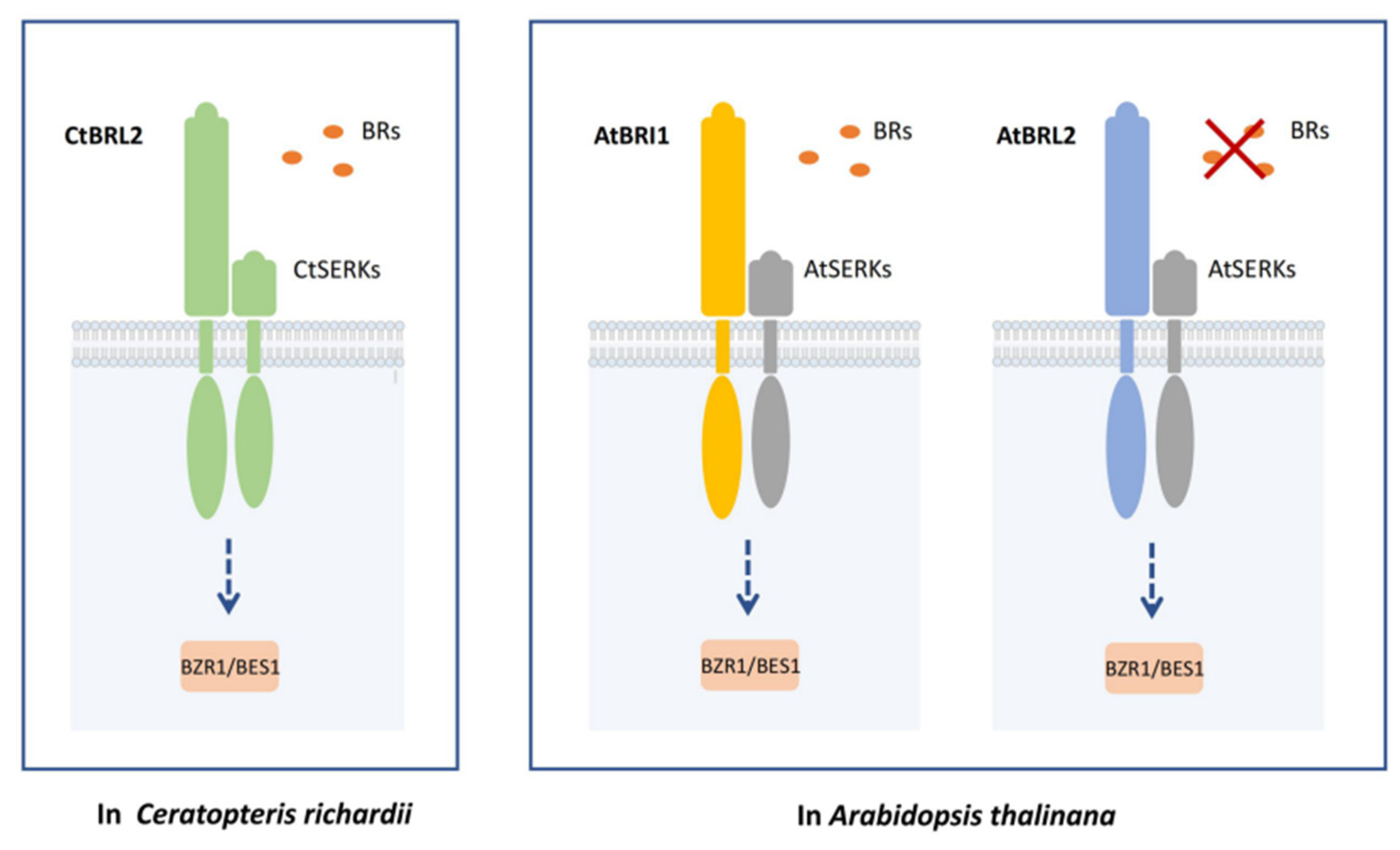{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Brassinolide Promotes Growth, Whereas Propiconazole Suppresses Growth in C-Fern
2.2. Presence of BRI1-like Receptors in C-Fern
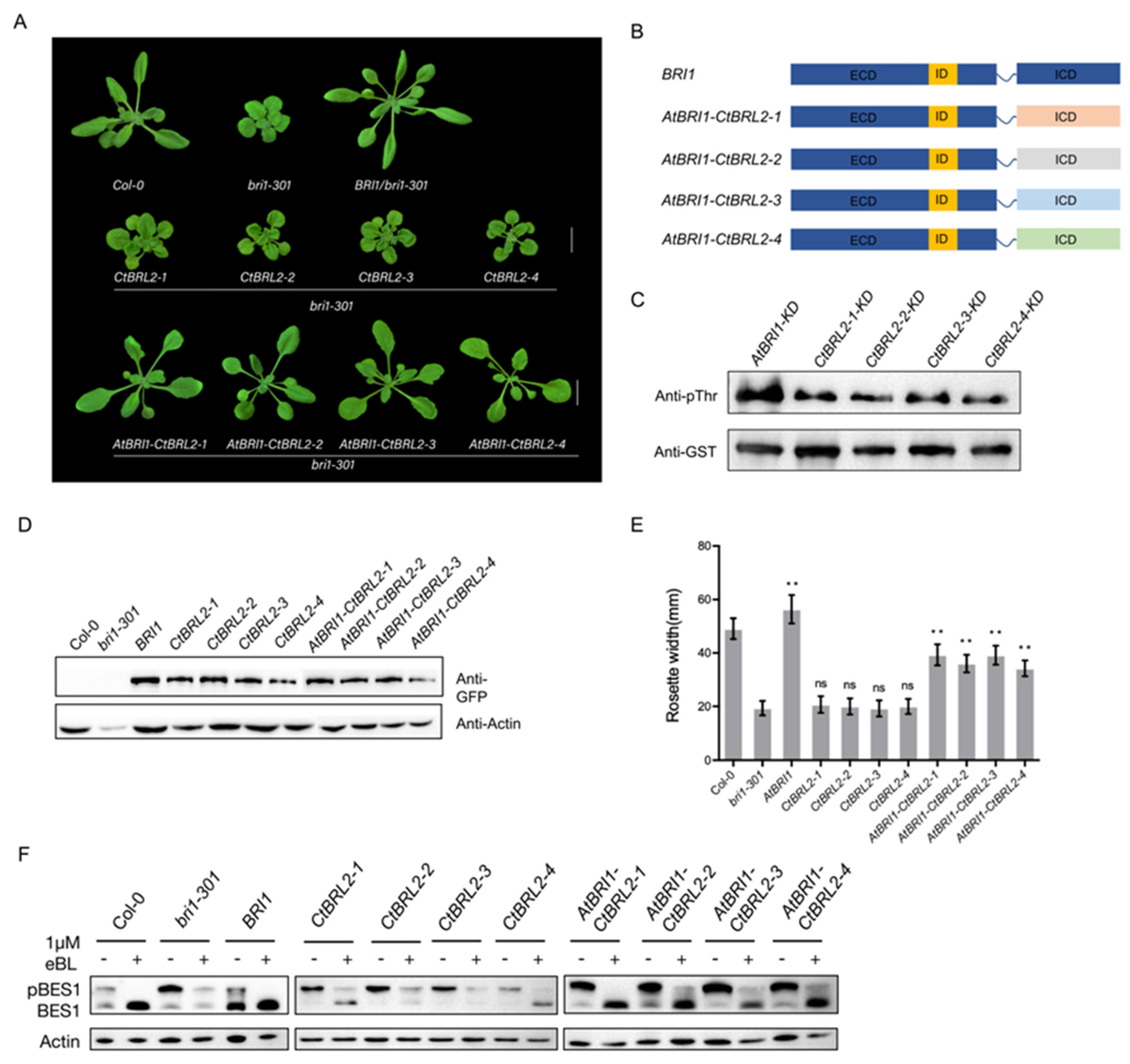
2.3. The Intracellular Domain of CtBRL2 Could Trigger the BR Signal
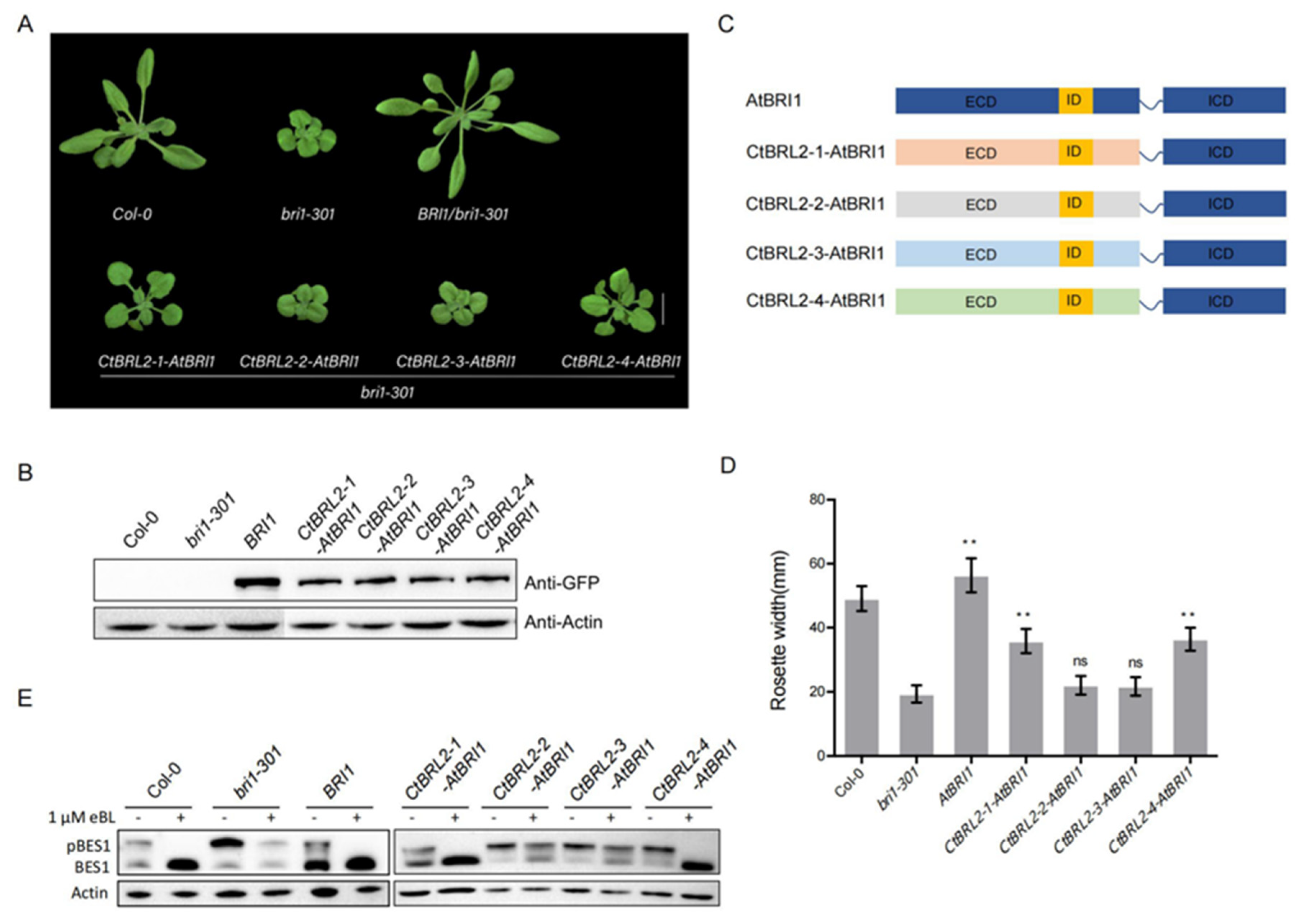
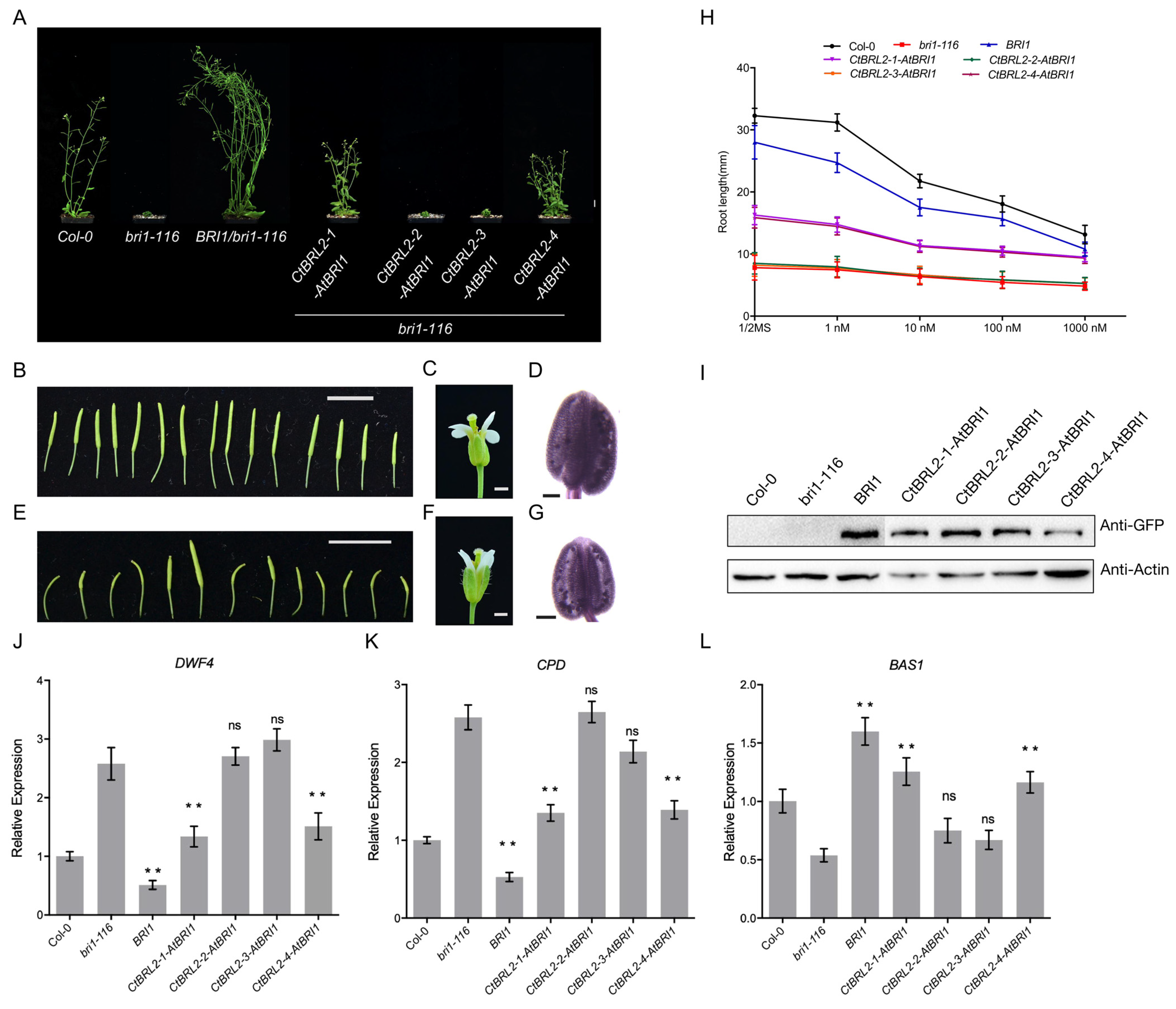
2.4. The Extracellular Domains of CtBRL2-1 and CtBRL2-4 Could Replace the Corresponding BRI1 Domains
2.5. The CDs of CtSERK1 and CtBRL2s Could Transphosphorylate Each Other In Vitro
3. Discussion
4. Materials and Methods
4.1. Phylogenetic Analysis
4.2. Plant Materials and Growth Conditions
4.3. Plant Material and Treatments for Ceratopteris richardii
4.4. Generation of Constructs and Transgenic Plants
4.5. RT-PCR and qRT-PCR Analysis
4.6. In Vitro Kinase Assay
4.7. Pull-Down Assay
4.8. Protein Extraction and Western Blot Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chow, B.; McCourt, P. Plant hormone receptors: Perception is everything. Genes Dev. 2006, 20, 1998–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, P. The Plant Hormones: Their Nature, Occurrence, and Functions. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2010; Volume 5, pp. 1–15. [Google Scholar]
- Wickett, N.J.; Mirarab, S.; Nguyen, N.; Warnow, T.; Carpenter, E.; Matasci, N.; Ayyampalayam, S.; Barker, M.S.; Burleigh, J.G.; Gitzendanner, M.A.; et al. Phylotranscriptomic analysis of the origin and early diversification of land plants. Proc. Natl. Acad. Sci. USA 2014, 111, E4859–E4868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donoghue, P.C.; Harrison, C.J.; Paps, J.; Schneider, H. The evolutionary emergence of land plants. Curr. Biol. 2021, 31, R1281–R1298. [Google Scholar] [CrossRef]
- Clouse, S.D. Brassinosteroids. Arabidopsis Book 2011, 9, e0151. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-Y.; Bai, M.-Y.; Oh, E.; Zhu, J.-Y. Brassinosteroid Signaling Network and Regulation of Photomorphogenesis. Annu. Rev. Genet. 2012, 46, 701–724. [Google Scholar] [CrossRef]
- Mao, J.; Li, J. Regulation of Three Key Kinases of Brassinosteroid Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 4340. [Google Scholar] [CrossRef]
- Szekeres, M.; Németh, K.; Koncz-Kálmán, Z.; Mathur, J.; Kauschmann, A.; Altmann, T.; Rédei, G.P.; Nagy, F.; Schell, J.; Koncz, C. Brassinosteroids Rescue the Deficiency of CYP90, a Cytochrome P450, Controlling Cell Elongation and De-etiolation in Arabidopsis. Cell 1996, 85, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chory, J. A Putative Leucine-Rich Repeat Receptor Kinase Involved in Brassinosteroid Signal Transduction. Cell 1997, 90, 929–938. [Google Scholar] [CrossRef] [Green Version]
- Nomura, T.; Kushiro, T.; Yokota, T.; Kamiya, Y.; Bishop, G.J.; Yamaguchi, S. The Last Reaction Producing Brassinolide Is Catalyzed by Cytochrome P-450s, CYP85A3 in Tomato and CYP85A2 in Arabidopsis. J. Biol. Chem. 2005, 280, 17873–17879. [Google Scholar] [CrossRef] [Green Version]
- Caño-Delgado, A.; Yin, Y.; Yu, C.; Vafeados, D.; Mora-García, S.; Cheng, J.-C.; Nam, K.H.; Li, J.; Chory, J. BRL1 and BRL3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development 2004, 131, 5341–5351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, A.; Wang, H.C.; Walker, J.C.; Li, J. BRL1, a leucine-rich repeat receptor-like protein kinase, is functionally redundant with BRI1 in regulating Arabidopsis brassinosteroid signaling. Plant J. 2004, 40, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Guerra, M.; Marquès-Bueno, M.; Mora-García, S.; Caño-Delgado, A.I. Delving into the evolutionary origin of steroid sensing in plants. Curr. Opin. Plant Biol. 2020, 57, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Bishop, G.J.; Kaneta, T.; Reid, J.B.; Chory, J.; Yokota, T. The LKA gene is a BRASSINOSTEROID INSENSITIVE 1 homolog of pea. Plant J. 2003, 36, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiu, S.-H.; Bleecker, A.B. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. USA 2001, 98, 10763–10768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaillais, Y.; Belkhadir, Y.; Balsemão-Pires, E.; Dangl, J.L.; Chory, J. Extracellular leucine-rich repeats as a platform for receptor/coreceptor complex formation. Proc. Natl. Acad. Sci. USA 2011, 108, 8503–8507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Nagpal, P.; Vitart, V.; McMorris, T.C.; Chory, J. A Role for Brassinosteroids in Light-Dependent Development of Arabidopsis. Science 1996, 272, 398–401. [Google Scholar] [CrossRef]
- Noguchi, T.; Fujioka, S.; Takatsuto, S.; Sakurai, A.; Yoshida, S.; Li, J.; Chory, J. Arabidopsis det2 Is Defective in the Conversion of (24R)-24-Methylcholest-4-En-3-One to (24R)-24-Methyl-5α-Cholestan-3-One in Brassinosteroid Biosynthesis1. Plant Physiol. 1999, 120, 833–840. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chory, J.; Yi, P.; Han, Z.; Li, X.; Olson, E.N. Brassinosteroids Regulate Dissociation of BKI1, a Negative Regulator of BRI1 Signaling, from the Plasma Membrane. Science 2006, 313, 1118–1122. [Google Scholar] [CrossRef]
- Hothorn, M.; Belkhadir, Y.; Dreux, M.; Dabi, T.; Noel, J.P.; Wilson, I.A.; Chory, J. Structural basis of steroid hormone perception by the receptor kinase BRI1. Nature 2011, 474, 467–471. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Russinova, E. Brassinosteroid signalling. Curr. Biol. 2020, 30, R294–R298. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A New Class of Transcription Factors Mediates Brassinosteroid-Regulated Gene Expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Bai, Q.; Wu, L.; Liu, H.; Liu, Y.; Xu, W.; Li, G.; Ren, H.; She, X.; Wu, G. EMS1 and BRI1 control separate biological processes via extracellular domain diversity and intracellular domain conservation. Nat. Commun. 2019, 10, 4165. [Google Scholar] [CrossRef] [Green Version]
- Bajguz, A. Effect of brassinosteroids on nucleic acids and protein content in cultured cells of Chlorella vulgaris. Plant Physiol. Biochem. 2000, 38, 209–215. [Google Scholar] [CrossRef]
- Cheon, J.; Fujioka, S.; Dilkes, B.P.; Choe, S. Brassinosteroids Regulate Plant Growth through Distinct Signaling Pathways in Selaginella and Arabidopsis. PLoS ONE 2013, 8, e81938. [Google Scholar] [CrossRef] [Green Version]
- Yokota, T.; Ohnishi, T.; Shibata, K.; Asahina, M.; Nomura, T.; Fujita, T.; Ishizaki, K.; Kohchi, T. Occurrence of brassinosteroids in non-flowering land plants, liverwort, moss, lycophyte and fern. Phytochemistry 2017, 136, 46–55. [Google Scholar] [CrossRef]
- Gómez-Garay, A.; y Galán, J.M.G.; Cabezuelo, A.; Pintos, B.; Prada, C.; Martín, L. Ecological Significance of Brassinosteroids in Three Temperate Ferns. In Current Advances in Fern Research; Fernández, H., Ed.; Springer: Cham, Switzerland, 2018; pp. 453–466. [Google Scholar]
- Nakazato, T.; Barker, M.S.; Rieseberg, L.H.; Gastony, G.J. Evolution of the nuclear genome of ferns and lycophytes. In Biology and Evolution of Ferns and Lycophytes; Haufler, C.H., Ranker, T.A., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 175–198. [Google Scholar]
- Sessa, E.B.; Der, J.P. Evolutionary Genomics of Ferns and Lycophytes. Adv. Bot. Res. 2016, 78, 215–254. [Google Scholar] [CrossRef]
- Li, F.-W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; de Vries, J.; Delaux, P.-M.; Eily, A.; Koppers, N.; Kuo, L.-Y.; Li, Z.; et al. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460–472. [Google Scholar] [CrossRef] [Green Version]
- Winter, K.-U.; Saedler, H.; Theißen, G. On the origin of class B floral homeotic genes: Functional substitution and dominant inhibition in Arabidopsis by expression of an orthologue from the gymnosperm Gnetum. Plant J. 2002, 31, 457–475. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Shen, Y.; Pu, R.; Hou, M.; Wei, Q.; Zhang, X.; Li, G.; Ren, H.; Wu, G. Brassinosteroids synthesised by CYP85A/A1 but not CYP85A2 function via a BRI1-like receptor but not via BRI1 in Picea abies. J. Exp. Bot. 2021, 72, 1748–1763. [Google Scholar] [CrossRef]
- Fernández, H.; Grossmann, J.; Gagliardini, V.; Feito, I.; Rivera, A.; Rodríguez, L.; Quintanilla, L.G.; Quesada, V.; Cañal, M.J.; Grossniklaus, U. Sexual and Apogamous Species of Woodferns Show Different Protein and Phytohormone Profiles. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, U.; Santiago, J.; Nicolet, J.; Olsson, V.; Spiga, F.M.; Hothorn, L.A.; Butenko, M.A.; Hothorn, M. Mechanistic basis for the activation of plant membrane receptor kinases by SERK-family coreceptors. Proc. Natl. Acad. Sci. USA 2018, 115, 3488–3493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Wang, Z.-Y.; Li, J.; Zhu, Q.; Lamb, C.; Ronald, P.; Chory, J. Perception of Brassinosteroids by the Extracellular Domain of the Receptor Kinase BRI1. Science 2000, 288, 2360–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannell, N.; Emms, D.M.; Hetherington, A.J.; MacKay, J.; Kelly, S.; Dolan, L.; Sweetlove, L.J. Multiple Metabolic Innovations and Losses Are Associated with Major Transitions in Land Plant Evolution. Curr. Biol. 2020, 30, 1783–1800.e11. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-Y.; Nakano, T.; Gendron, J.; He, J.; Chen, M.; Vafeados, D.; Yang, Y.; Fujioka, S.; Yoshida, S.; Asami, T.; et al. Nuclear-Localized BZR1 Mediates Brassinosteroid-Induced Growth and Feedback Suppression of Brassinosteroid Biosynthesis. Dev. Cell 2002, 2, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gou, X. Receptor-Like Protein Kinases Function Upstream of MAPKs in Regulating Plant Development. Int. J. Mol. Sci. 2020, 21, 7638. [Google Scholar] [CrossRef]
- Bai, Q.; Li, C.; Wu, L.; Liu, H.; Ren, H.; Li, G.; Wang, Q.; Wu, G.; Zheng, B. Engineering Chimeras by Fusing Plant Receptor-like Kinase EMS1 and BRI1 Reveals the Two Receptors’ Structural Specificity and Molecular Mechanisms. Int. J. Mol. Sci. 2022, 23, 2155. [Google Scholar] [CrossRef]
- Wang, X.; Kota, U.; He, K.; Blackburn, K.; Li, J.; Goshe, M.B.; Huber, S.C.; Clouse, S.D. Sequential transphosphorylation of the BRI1/BAK1 receptor kinase complex impacts early events in brassinosteroid signaling. Dev. Cell 2008, 15, 220–235. [Google Scholar] [CrossRef] [Green Version]
- Hecht, V.; Vielle-Calzada, J.P.; Hartog, M.V.; Schmidt, E.D.; Boutilier, K.; Grossniklaus, U.; De Vries, S.C. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol. 2001, 127, 803–816. [Google Scholar] [CrossRef]
- Gou, X.; Yin, H.; He, K.; Du, J.; Yi, J.; Xu, S.; Lin, H.; Clouse, S.D.; Li, J. Genetic Evidence for an Indispensable Role of Somatic Embryogenesis Receptor Kinases in Brassinosteroid Signaling. PLoS Genet. 2012, 8, e1002452. [Google Scholar] [CrossRef] [Green Version]
- Hirano, K.; Nakajima, M.; Asano, K.; Nishiyama, T.; Sakakibara, H.; Kojima, M.; Katoh, E.; Xiang, H.; Tanahashi, T.; Hasebe, M.; et al. The GID1-Mediated Gibberellin Perception Mechanism Is Conserved in the Lycophyte Selaginella moellendorffii but Not in the Bryophyte Physcomitrella patens. Plant Cell 2007, 19, 3058–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasumura, Y.; Crumpton-Taylor, M.; Fuentes, S.; Harberd, N.P. Step-by-Step Acquisition of the Gibberellin-DELLA Growth-Regulatory Mechanism during Land-Plant Evolution. Curr. Biol. 2007, 17, 1225–1230. [Google Scholar] [CrossRef] [Green Version]
- Furumizu, C.; Sawa, S. Insight into early diversification of leucine-rich repeat receptor-like kinases provided by the sequenced moss and hornwort genomes. Plant Mol. Biol. 2021, 107, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Holton, N.; Caño-Delgado, A.; Harrison, K.; Montoya, T.; Chory, J.; Bishop, G.J. Tomato BRASSINOSTEROID INSENSITIVE1 Is Required for Systemin-Induced Root Elongation in Solanum pimpinellifolium but Is Not Essential for Wound Signaling. Plant Cell 2007, 19, 1709–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecchia, M.A.; García-Hourquet, M.; Lozano-Elena, F.; Planas-Riverola, A.; Blasco-Escamez, D.; Marquès-Bueno, M.; Mora-García, S.; Caño-Delgado, A.I. The BES1/BZR1-family transcription factor MpBES1 regulates cell division and differentiation in Marchantia polymorpha. Curr. Biol. 2021, 31, 4860–4869.e4868. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, B.; Xing, K.; Zhang, J.; Liu, H.; Ali, K.; Li, W.; Bai, Q.; Ren, H. Evolutionary Analysis and Functional Identification of Ancient Brassinosteroid Receptors in Ceratopteris richardii. Int. J. Mol. Sci. 2022, 23, 6795. https://doi.org/10.3390/ijms23126795
Zheng B, Xing K, Zhang J, Liu H, Ali K, Li W, Bai Q, Ren H. Evolutionary Analysis and Functional Identification of Ancient Brassinosteroid Receptors in Ceratopteris richardii. International Journal of Molecular Sciences. 2022; 23(12):6795. https://doi.org/10.3390/ijms23126795
Chicago/Turabian StyleZheng, Bowen, Kaixin Xing, Jiaojiao Zhang, Hui Liu, Khawar Ali, Wenjuan Li, Qunwei Bai, and Hongyan Ren. 2022. "Evolutionary Analysis and Functional Identification of Ancient Brassinosteroid Receptors in Ceratopteris richardii" International Journal of Molecular Sciences 23, no. 12: 6795. https://doi.org/10.3390/ijms23126795