
Effects of Melatonin on the Transcriptome of Human Granulosa Cells, Fertilization and Blastocyst Formation

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
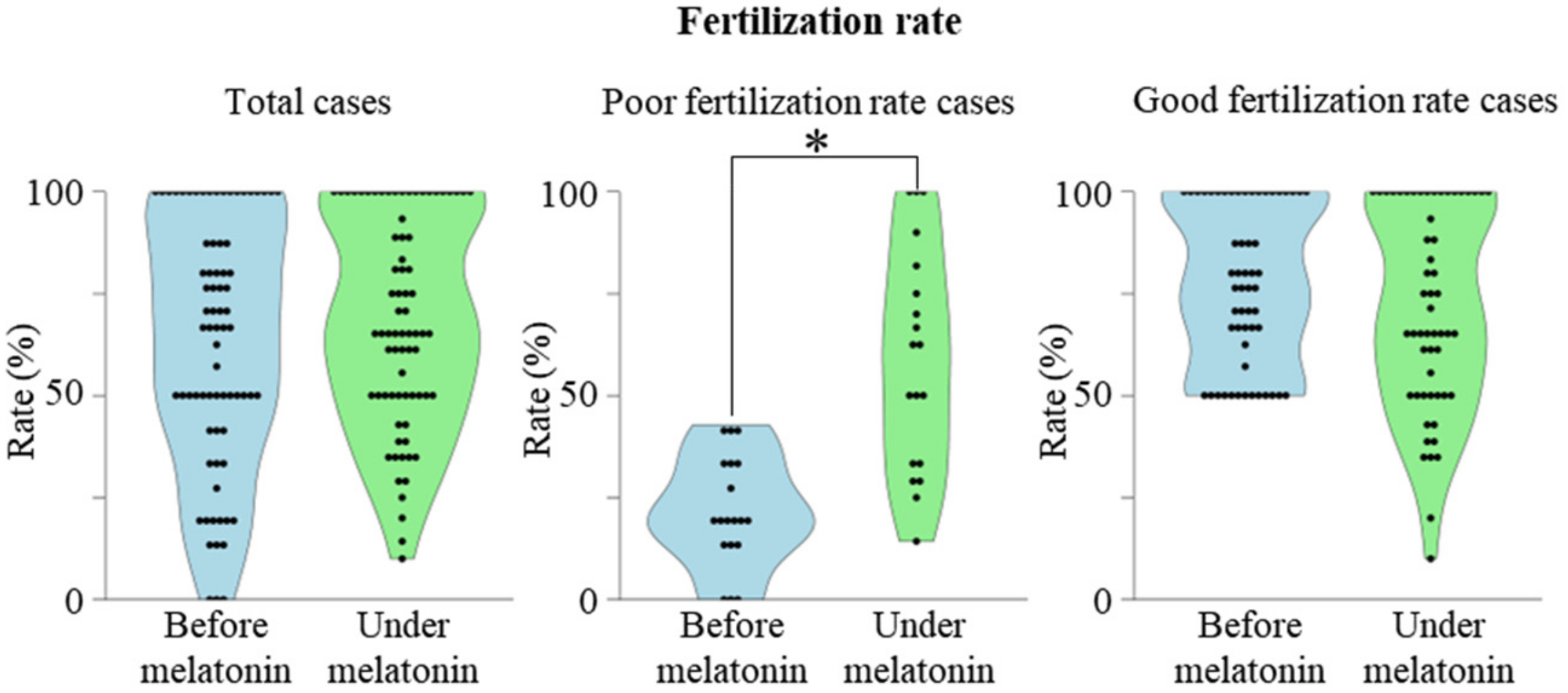
2.1. Association between Previous Fertilization Rate and the Effect of Melatonin
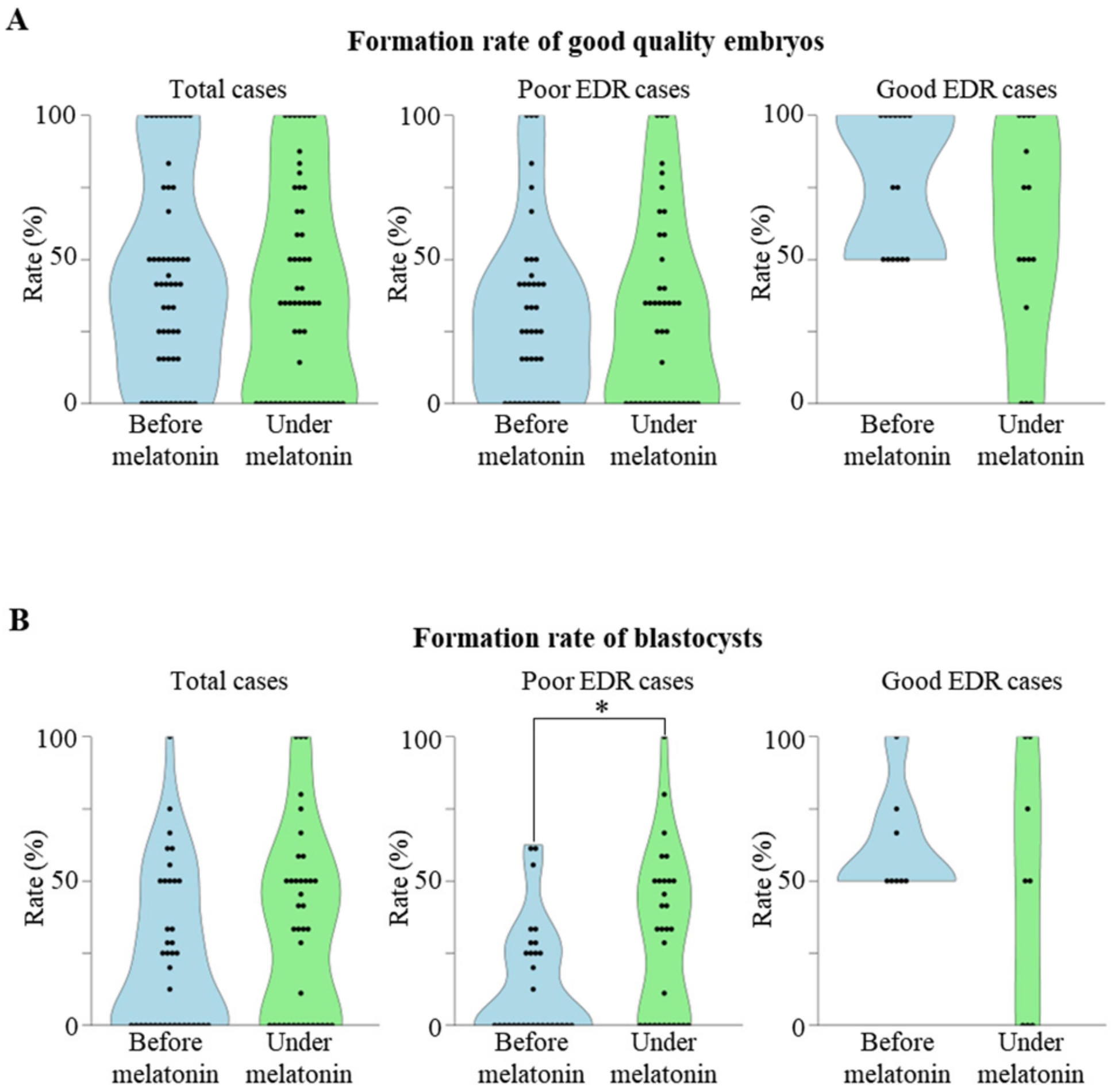
2.2. Association between Previous Embryo Development Rate (EDR) and the Effect of Melatonin
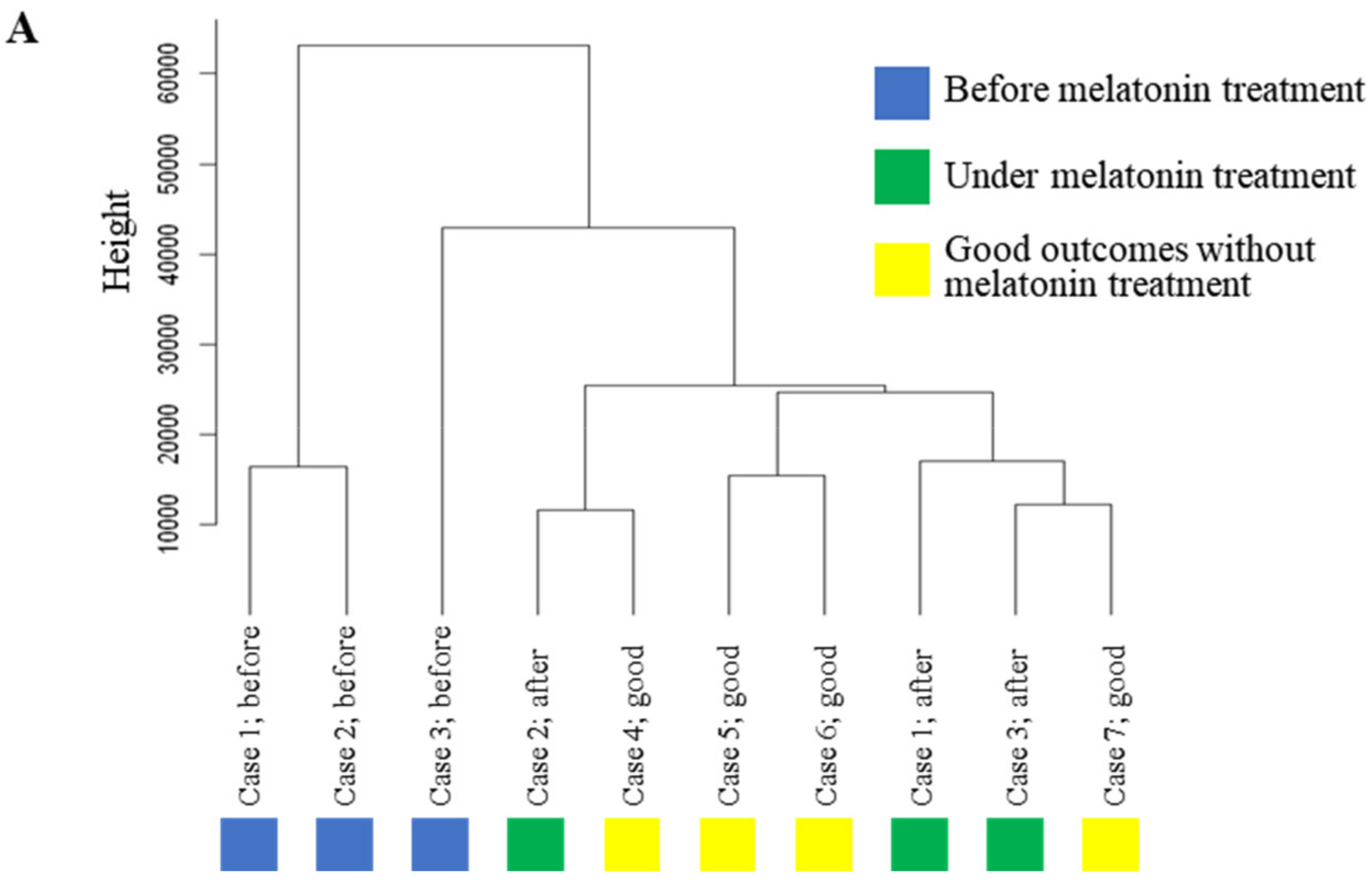
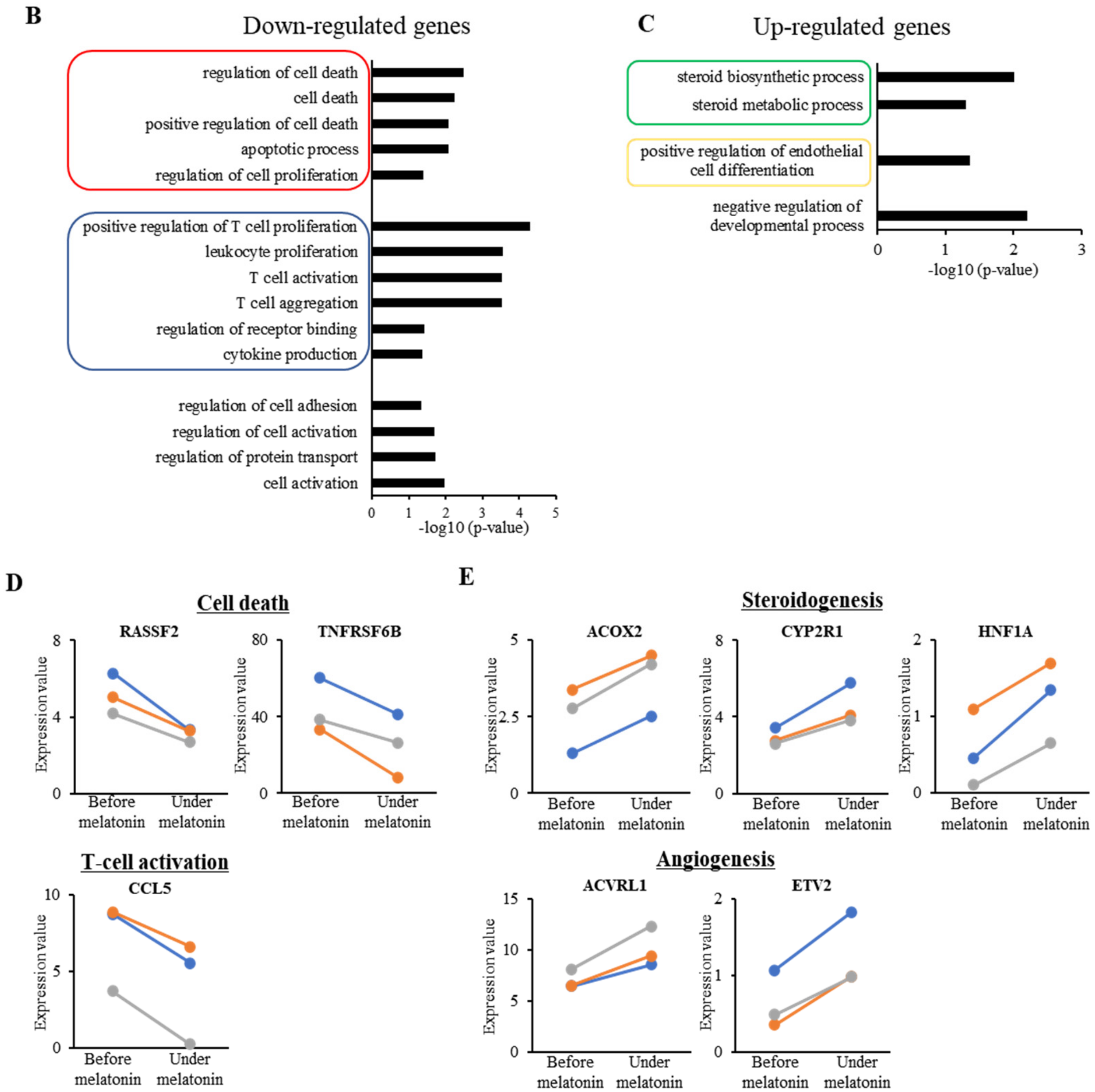
2.3. Effect of Melatonin Treatment on Transcriptome Profile in GCs
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. ART Procedure
4.3. Melatonin Treatment
4.4. Outcomes
4.5. Transcriptome Analysis of Human GCs
4.6. Real-Time RT-PCR
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Jozaki, M.; Tanabe, M.; Shirafuta, Y.; Mihara, Y.; Shinagawa, M.; Tamura, I.; Maekawa, R.; Sato, S.; Taketani, T.; et al. Importance of Melatonin in Assisted Reproductive Technology and Ovarian Aging. Int. J. Mol. Sci. 2020, 21, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Qian, Y.; Chen, X.; Ji, X.; Ou, S.; Li, R.; Yang, D.; Li, Y. Effect of early rescue ICSI and split IVF-ICSI in preventing low fertilization rate during the first ART cycle: A real-world retrospective cohort study. Reprod. Med. Biol. 2022, 21, e12420. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, O.; Jwa, S.C.; Kuwahara, A.; Katagiri, Y.; Kuwabara, Y.; Hamatani, T.; Harada, M.; Ichikawa, T. Assisted reproductive technology in Japan: A summary report for 2017 by the Ethics Committee of the Japan Society of Obstetrics and Gynecology. Reprod. Med. Biol. 2020, 19, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Lee, J. Is age-related increase of chromosome segregation errors in mammalian oocytes caused by cohesin deterioration? Reprod. Med. Biol. 2020, 19, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y.; et al. Melatonin as a free radical scavenger in the ovarian follicle. Endocr. J. 2013, 60, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Seino, T.; Saito, H.; Kaneko, T.; Takahashi, T.; Kawachiya, S.; Kurachi, H. Eight-hydroxy-2′-deoxyguanosine in granulosa cells is correlated with the quality of oocytes and embryos in an in vitro fertilization-embryo transfer program. Fertil. Steril. 2002, 77, 1184–1190. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Kizuka, F.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; et al. The role of melatonin as an antioxidant in the follicle. J. Ovarian Res. 2012, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Tamura, H.; Takasaki, A.; Taketani, T.; Tanabe, M.; Lee, L.; Tamura, I.; Maekawa, R.; Aasada, H.; Yamagata, Y.; Sugino, N. Melatonin and female reproduction. J. Obstet. Gynaecol. Res. 2014, 40, 1–11. [Google Scholar] [CrossRef]
- Tamura, H.; Nakamura, Y.; Korkmaz, A.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and the ovary: Physiological and pathophysiological implications. Fertil. Steril. 2009, 92, 328–343. [Google Scholar] [CrossRef]
- Tamura, H.; Takasaki, A.; Miwa, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Taketani, T.; Matsuoka, A.; Yamagata, Y.; Shimamura, K.; et al. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J. Pineal Res. 2008, 44, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Batioglu, A.S.; Sahin, U.; Gurlek, B.; Ozturk, N.; Unsal, E. The efficacy of melatonin administration on oocyte quality. Gynecol. Endocrinol. 2012, 28, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Eryilmaz, O.G.; Devran, A.; Sarikaya, E.; Aksakal, F.N.; Mollamahmutoglu, L.; Cicek, N. Melatonin improves the oocyte and the embryo in IVF patients with sleep disturbances, but does not improve the sleeping problems. J. Assist. Reprod. Genet. 2011, 28, 815–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahromi, B.N.; Sadeghi, S.; Alipour, S.; Parsanezhad, M.E.; Alamdarloo, S.M. Effect of Melatonin on the Outcome of Assisted Reproductive Technique Cycles in Women with Diminished Ovarian Reserve: A Double-Blinded Randomized Clinical Trial. Iran J. Med. Sci. 2017, 42, 73–78. [Google Scholar]
- Nishihara, T.; Hashimoto, S.; Ito, K.; Nakaoka, Y.; Matsumoto, K.; Hosoi, Y.; Morimoto, Y. Oral melatonin supplementation improves oocyte and embryo quality in women undergoing in vitro fertilization-embryo transfer. Gynecol. Endocrinol. 2014, 30, 359–362. [Google Scholar] [CrossRef]
- Espino, J.; Macedo, M.; Lozano, G.; Ortiz, A.; Rodriguez, C.; Rodriguez, A.B.; Bejarano, I. Impact of Melatonin Supplementation in Women with Unexplained Infertility Undergoing Fertility Treatment. Antioxidants 2019, 8, 338. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.H.; Miyano, T. Interaction between growing oocytes and granulosa cells in vitro. Reprod. Med. Biol. 2020, 19, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Regan, S.L.P.; Knight, P.G.; Yovich, J.L.; Leung, Y.; Arfuso, F.; Dharmarajan, A. Granulosa Cell Apoptosis in the Ovarian Follicle—A Changing View. Front. Endocrinol. 2018, 9, 61. [Google Scholar] [CrossRef]
- Fang, L.; Li, Y.; Wang, S.; Yu, Y.; Li, Y.; Guo, Y.; Yan, Y.; Sun, Y.P. Melatonin induces progesterone production in human granulosa-lutein cells through upregulation of StAR expression. Aging 2019, 11, 9013–9024. [Google Scholar] [CrossRef]
- Li, Y.; Fang, L.; Zhang, R.; Wang, S.; Li, Y.; Yan, Y.; Yu, Y.; Cheng, J.C.; Sun, Y.P. Melatonin stimulates VEGF expression in human granulosa-lutein cells: A potential mechanism for the pathogenesis of ovarian hyperstimulation syndrome. Mol. Cell Endocrinol. 2020, 518, 110981. [Google Scholar] [CrossRef]
- Cheng, J.C.; Fang, L.; Li, Y.; Wang, S.; Li, Y.; Yan, Y.; Jia, Q.; Wu, Z.; Wang, Z.; Han, X.; et al. Melatonin stimulates aromatase expression and estradiol production in human granulosa-lutein cells: Relevance for high serum estradiol levels in patients with ovarian hyperstimulation syndrome. Exp. Mol. Med. 2020, 52, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Schoolcraft, W.B.; Wagley, L.; Schlenker, T.; Stevens, J.; Hesla, J. A prospective randomized trial of blastocyst culture and transfer in in-vitro fertilization. Hum. Reprod. 1998, 13, 3434–3440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaban, B.; Urman, B.; Sertac, A.; Alatas, C.; Aksoy, S.; Mercan, R. Blastocyst quality affects the success of blastocyst-stage embryo transfer. Fertil. Steril. 2000, 74, 282–287. [Google Scholar] [CrossRef]
- Dumoulin, J.C.; Coonen, E.; Bras, M.; van Wissen, L.C.; Ignoul-Vanvuchelen, R.; Bergers-Jansen, J.M.; Derhaag, J.G.; Geraedts, J.P.; Evers, J.L. Comparison of in-vitro development of embryos originating from either conventional in-vitro fertilization or intracytoplasmic sperm injection. Hum. Reprod. 2000, 15, 402–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Wang, J.; Zhang, Z.; Yang, M.; Li, Y.; Tian, X.; Ma, T.; Tao, J.; Zhu, K.; Song, Y.; et al. Mitochondria Synthesize Melatonin to Ameliorate Its Function and Improve Mice Oocyte’s Quality under in Vitro Conditions. Int. J. Mol. Sci. 2016, 17, 939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, M.; Tamura, H.; Taketani, T.; Okada, M.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y.; Sugino, N. Melatonin protects the integrity of granulosa cells by reducing oxidative stress in nuclei, mitochondria, and plasma membranes in mice. J. Reprod. Dev. 2015, 61, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Wang, R.X.; Li, M.H.; Sun, T.C.; Zhou, Y.W.; Li, Y.Y.; Sun, L.H.; Zhang, B.L.; Lian, Z.X.; Xue, S.G.; et al. Melatonin Reduces Androgen Production and Upregulates Heme Oxygenase-1 Expression in Granulosa Cells from PCOS Patients with Hypoestrogenia and Hyperandrogenia. Oxid. Med. Cell Longev. 2019, 2019, 8218650. [Google Scholar] [CrossRef]
- Shen, M.; Cao, Y.; Jiang, Y.; Wei, Y.; Liu, H. Melatonin protects mouse granulosa cells against oxidative damage by inhibiting FOXO1-mediated autophagy: Implication of an antioxidation-independent mechanism. Redox. Biol. 2018, 18, 138–157. [Google Scholar] [CrossRef]
- Zhang, H.; Li, C.; Wen, D.; Li, R.; Lu, S.; Xu, R.; Tang, Y.; Sun, Y.; Zhao, X.; Pan, M. Melatonin improves the quality of maternally aged oocytes by maintaining intercellular communication and antioxidant metabolite supply. Redox. Biol. 2021, 49, 102215. [Google Scholar] [CrossRef]
- Vos, M.D.; Ellis, C.A.; Elam, C.; Ulku, A.S.; Taylor, B.J.; Clark, G.J. RASSF2 is a novel K-Ras-specific effector and potential tumor suppressor. J. Biol. Chem. 2003, 278, 28045–28051. [Google Scholar] [CrossRef] [Green Version]
- Hudlikar, R.R.; Sargsyan, D.; Wu, R.; Su, S.; Zheng, M.; Kong, A.N. Triterpenoid corosolic acid modulates global CpG methylation and transcriptome of tumor promotor TPA induced mouse epidermal JB6 P+ cells. Chem. Biol. Interact. 2020, 321, 109025. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.L.; Lin, W.W. Decoy receptor 3: An endogenous immunomodulator in cancer growth and inflammatory reactions. J. Biomed. Sci. 2017, 24, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, O.; Burke, J.D.; Gao, D.F.; Fish, E.N. The chemokine CCL5 regulates glucose uptake and AMP kinase signaling in activated T cells to facilitate chemotaxis. J. Biol. Chem. 2012, 287, 29406–29416. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.M.; Kubota, H.; Okita, M.; Maeda, T. The anti-inflammatory and antioxidant effects of melatonin on LPS-stimulated bovine mammary epithelial cells. PLoS ONE 2017, 12, e0178525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kepka, M.; Szwejser, E.; Pijanowski, L.; Verburg-van Kemenade, B.M.; Chadzinska, M. A role for melatonin in maintaining the pro- and anti-inflammatory balance by influencing leukocyte migration and apoptosis in carp. Dev. Comp. Immunol. 2015, 53, 179–190. [Google Scholar] [CrossRef]
- Zhang, N.; Hartig, H.; Dzhagalov, I.; Draper, D.; He, Y.W. The role of apoptosis in the development and function of T lymphocytes. Cell Res. 2005, 15, 749–769. [Google Scholar] [CrossRef]
- Bender, H.R.; Trau, H.A.; Duffy, D.M. Placental Growth Factor Is Required for Ovulation, Luteinization, and Angiogenesis in Primate Ovulatory Follicles. Endocrinology 2018, 159, 710–722. [Google Scholar] [CrossRef]
- Kizuka, F.; Tokuda, N.; Takagi, K.; Adachi, Y.; Lee, L.; Tamura, I.; Maekawa, R.; Taketani, T.; Tamura, H.; Suzuki, T.; et al. Involvement of bone marrow-derived vascular progenitor cells in neovascularization during formation of the corpus luteum in mice. Biol. Reprod. 2012, 87, 55. [Google Scholar] [CrossRef]
- Shinagawa, M.; Tamura, I.; Maekawa, R.; Sato, S.; Shirafuta, Y.; Mihara, Y.; Okada-Matsumoto, M.; Taketani, T.; Asada, H.; Tamura, H.; et al. C/EBPbeta regulates Vegf gene expression in granulosa cells undergoing luteinization during ovulation in female rats. Sci. Rep. 2019, 9, 714. [Google Scholar] [CrossRef] [Green Version]
- Van Blerkom, J.; Antczak, M.; Schrader, R. The developmental potential of the human oocyte is related to the dissolved oxygen content of follicular fluid: Association with vascular endothelial growth factor levels and perifollicular blood flow characteristics. Hum. Reprod. 1997, 12, 1047–1055. [Google Scholar] [CrossRef]
- Van Blerkom, J. Intrafollicular influences on human oocyte developmental competence: Perifollicular vascularity, oocyte metabolism and mitochondrial function. Hum. Reprod. 2000, 15 (Suppl. 2), 173–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokal, E.V.; Vrtovec, H.M.; Virant Klun, I.; Verdenik, I. Prolonged HCG action affects angiogenic substances and improves follicular maturation, oocyte quality and fertilization competence in patients with polycystic ovarian syndrome. Hum. Reprod. 2005, 20, 1562–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malamitsi-Puchner, A.; Sarandakou, A.; Baka, S.G.; Tziotis, J.; Rizos, D.; Hassiakos, D.; Creatsas, G. Concentrations of angiogenic factors in follicular fluid and oocyte-cumulus complex culture medium from women undergoing in vitro fertilization: Association with oocyte maturity and fertilization. Fertil. Steril. 2001, 76, 98–101. [Google Scholar] [CrossRef]
- Naredi, N.; Singh, S.K.; Sharma, R. Does Perifollicular Vascularity on the Day of Oocyte Retrieval Affect Pregnancy Outcome in an In Vitro Fertilization Cycle? J. Hum. Reprod. Sci. 2017, 10, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Bhal, P.S.; Pugh, N.D.; Chui, D.K.; Gregory, L.; Walker, S.M.; Shaw, R.W. The use of transvaginal power Doppler ultrasonography to evaluate the relationship between perifollicular vascularity and outcome in in-vitro fertilization treatment cycles. Hum. Reprod. 1999, 14, 939–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taketani, T.; Tamura, H.; Takasaki, A.; Lee, L.; Kizuka, F.; Tamura, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Shimamura, K.; et al. Protective role of melatonin in progesterone production by human luteal cells. J. Pineal Res. 2011, 51, 207–213. [Google Scholar] [CrossRef]
- Miller, W.L. Genetic disorders of Vitamin D biosynthesis and degradation. J. Steroid. Biochem. Mol. Biol. 2017, 165, 101–108. [Google Scholar] [CrossRef]
- Nenicu, A.; Luers, G.H.; Kovacs, W.; David, M.; Zimmer, A.; Bergmann, M.; Baumgart-Vogt, E. Peroxisomes in human and mouse testis: Differential expression of peroxisomal proteins in germ cells and distinct somatic cell types of the testis. Biol. Reprod. 2007, 77, 1060–1072. [Google Scholar] [CrossRef] [Green Version]
- Shih, D.Q.; Bussen, M.; Sehayek, E.; Ananthanarayanan, M.; Shneider, B.L.; Suchy, F.J.; Shefer, S.; Bollileni, J.S.; Gonzalez, F.J.; Breslow, J.L.; et al. Hepatocyte nuclear factor-1alpha is an essential regulator of bile acid and plasma cholesterol metabolism. Nat. Genet. 2001, 27, 375–382. [Google Scholar] [CrossRef]
- Salehnia, M.; Zavareh, S. The effects of progesterone on oocyte maturation and embryo development. Int. J. Fertil. Steril. 2013, 7, 74–81. [Google Scholar]
- Pang, Y.; Zhao, S.; Sun, Y.; Jiang, X.; Hao, H.; Du, W.; Zhu, H. Protective effects of melatonin on the in vitro developmental competence of bovine oocytes. Anim. Sci. J. 2018, 89, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, J.; Li, Y.; Guo, X.; Li, J.; Zhong, R.; Zhang, X. Melatonin-induced demethylation of antioxidant genes increases antioxidant capacity through RORalpha in cumulus cells of prepubertal lambs. Free Radic. Biol. Med. 2019, 131, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Tamura, I.; Kawamoto-Jozaki, M.; Fujimura, T.; Doi-Tanaka, Y.; Takagi, H.; Shirafuta, Y.; Mihara, Y.; Taketani, T.; Tamura, H.; Sugino, N. Relationship between follicular size and developmental capacity of oocytes under controlled ovarian hyperstimulation in assisted reproductive technologies. Reprod. Med. Biol. 2021, 20, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Iwase, A.; Furui, K.; Kitagawa, T.; Matsui, Y.; Yoshikawa, M.; Kikkawa, F. Preferable correlation to blastocyst development and pregnancy rates with a new embryo grading system specific for day 3 embryos. J. Assist. Reprod. Genet. 2007, 24, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomsu, M.; Sharma, V.; Miller, D. Embryo quality and IVF treatment outcomes may correlate with different sperm comet assay parameters. Hum. Reprod. 2002, 17, 1856–1862. [Google Scholar] [CrossRef] [Green Version]
- Tamura, I.; Maekawa, R.; Jozaki, K.; Ohkawa, Y.; Takagi, H.; Doi-Tanaka, Y.; Shirafuta, Y.; Mihara, Y.; Taketani, T.; Sato, S.; et al. Transcription factor C/EBPbeta induces genome-wide H3K27ac and upregulates gene expression during decidualization of human endometrial stromal cells. Mol. Cell Endocrinol. 2021, 520, 111085. [Google Scholar] [CrossRef]
- Tamura, I.; Ohkawa, Y.; Sato, T.; Suyama, M.; Jozaki, K.; Okada, M.; Lee, L.; Maekawa, R.; Asada, H.; Sato, S.; et al. Genome-wide analysis of histone modifications in human endometrial stromal cells. Mol. Endocrinol. 2014, 28, 1656–1669. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Yamamoto, S.; Sakaguti, S.; Isayama, K.; Oka, M.; Nagano, H.; Mizukami, Y. A novel somatic mutation of SIN3A detected in breast cancer by whole-exome sequencing enhances cell proliferation through ERalpha expression. Sci. Rep. 2018, 8, 16000. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Supek, F.; Bosnjak, M.; Skunca, N.; Smuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar]
- Shirafuta, Y.; Tamura, I.; Ohkawa, Y.; Maekawa, R.; Doi-Tanaka, Y.; Takagi, H.; Mihara, Y.; Shinagawa, M.; Taketani, T.; Sato, S.; et al. Integrated Analysis of Transcriptome and Histone Modifications in Granulosa Cells During Ovulation in Female Mice. Endocrinology 2021, 162, bqab128. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Poor Fertilization Rate Cases | Good Fertilization Rate Cases | Poor EDR Cases | Good EDR Cases | |
|---|---|---|---|---|
| No. of patients | 19 | 56 | 41 | 15 |
| Patient age (mean ± SD) | 35.4± 3.7 | 37.3 ± 4.3 | 36.9 ± 4.2 | 38.3 ± 4.1 |
| Patient’s complications | ||||
| Myoma/adenomyosis (%) | 15.8 | 16.1 | 26.7 | 12.2 |
| Endometriosis (%) | 26.3 | 16.1 | 13.3 | 17.1 |
| PCOS (%) | 5.3 | 5.4 | 0.0 | 7.3 |
| Tubal factor (%) | 21.1 | 21.4 | 13.3 | 24.4 |
| Unilateral obstruction (%) | 10.5 | 12.5 | 13.3 | 12.2 |
| Bilateral obstruction (%) | 10.5 | 8.9 | 0.0 | 12.2 |
| Male factor (%) | 15.8 | 16.1 | 26.7 | 12.2 |
| Asthenozoospermia (%) | 5.3 | 8.9 | 13.3 | 7.3 |
| Oligospermia (%) | 10.5 | 7.1 | 13.3 | 4.9 |
| EDR, embryo development rate |
| Blastocyst Formation Rate (%) | GnRH Agonist Protocol | Fertilization | |||||
|---|---|---|---|---|---|---|---|
| Case No. | Age | Before Melatonin Treatment | Under Melatonin Treatment | Before Melatonin Treatment | Under Melatonin Treatment | Before Melatonin Treatment | Under Melatonin Treatment |
| 1 | 32 | 0 | 40.0 | long protocol | long protocol | ICSI | ICSI |
| 2 | 33 | 0 | 28.9 | long protocol | long protocol | cIVF | cIVF |
| 3 | 36 | 0 | 100.0 | long protocol | long protocol | ICSI | ICSI |
| 4 | 35 | 61.5 | long protocol | cIVF | |||
| 5 | 36 | 50.0 | long protocol | cIVF | |||
| 6 | 34 | 50.0 | short protocol | cIVF | |||
| 7 | 37 | 100.0 | long protocol | cIVF | |||
| long protocol, GnRHa (900 μg/day) was given from the mid-luteal phase in the previous cycle until two days before oocyte retrieval. | |||||||
| cIVF, conventional in vitro fertilization | ICSI, intracytoplasmic sperm injection | ||||||
| Down-Regulated Genes | Up-Regulated Genes | ||
|---|---|---|---|
| Gene | p Value | Gene | p Value |
| NUP62CL | 1.08 × 10−03 | RP11-211N8.3 | 3.88 × 10−04 |
| TRAF6 | 1.40 × 10−03 | SNRPGP2 | 9.07 × 10−04 |
| ZNF501 | 1.98 × 10−03 | RPS17P5 | 9.07 × 10−04 |
| RP11-355I22.7 | 2.26 × 10−03 | RPS11P7 | 9.07 × 10−04 |
| RP11-3N2.13 | 3.29 × 10−03 | CTD-2013M15.1 | 9.94 × 10−04 |
| ZDHHC15 | 5.43 × 10−03 | COX7B | 1.06 × 10−03 |
| CENPV | 5.82 × 10−03 | RP4-738P15.1 | 1.36 × 10−03 |
| RP11-564C4.6 | 6.78 × 10−03 | RP11-787I22.3 | 1.38 × 10−03 |
| CD244 | 6.83 × 10−03 | RP11-874J12.4 | 1.39 × 10−03 |
| AC078883.3 | 7.24 × 10−03 | CTC-507E2.1 | 1.68 × 10−03 |
| ZNF835 | 7.36 × 10−03 | SMIM11 | 2.09 × 10−03 |
| RP11-63E5.6 | 7.54 × 10−03 | GJB7 | 3.60 × 10−03 |
| MMP9 | 7.63 × 10−03 | PTTG1 | 4.90 × 10−03 |
| RP11-768F21.1 | 8.44 × 10−03 | CXCL6 | 5.12 × 10−03 |
| AC093668.2 | 8.57 × 10−03 | RP11-315I20.1 | 5.28 × 10−03 |
| RP11-73M18.7 | 1.09 × 10−02 | ACOX2 | 5.66 × 10−03 |
| AC061975.7 | 1.14 × 10−02 | MAGOH2 | 6.78 × 10−03 |
| AC090673.2 | 1.14 × 10−02 | RP11-440L14.1 | 6.78 × 10−03 |
| CTD-2514K5.4 | 1.14 × 10−02 | ENPP7P10 | 7.43 × 10−03 |
| MRPS21P3 | 1.14 × 10−02 | RPL7AP30 | 7.66 × 10−03 |
| RN7SL474P | 1.14 × 10−02 | RP11-514P8.7 | 7.91 × 10−03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamura, I.; Tamura, H.; Kawamoto-Jozaki, M.; Shirafuta, Y.; Fujimura, T.; Doi-Tanaka, Y.; Mihara, Y.; Taketani, T.; Sugino, N. Effects of Melatonin on the Transcriptome of Human Granulosa Cells, Fertilization and Blastocyst Formation. Int. J. Mol. Sci. 2022, 23, 6731. https://doi.org/10.3390/ijms23126731
Tamura I, Tamura H, Kawamoto-Jozaki M, Shirafuta Y, Fujimura T, Doi-Tanaka Y, Mihara Y, Taketani T, Sugino N. Effects of Melatonin on the Transcriptome of Human Granulosa Cells, Fertilization and Blastocyst Formation. International Journal of Molecular Sciences. 2022; 23(12):6731. https://doi.org/10.3390/ijms23126731
Chicago/Turabian StyleTamura, Isao, Hiroshi Tamura, Mai Kawamoto-Jozaki, Yuichiro Shirafuta, Taishi Fujimura, Yumiko Doi-Tanaka, Yumiko Mihara, Toshiaki Taketani, and Norihiro Sugino. 2022. "Effects of Melatonin on the Transcriptome of Human Granulosa Cells, Fertilization and Blastocyst Formation" International Journal of Molecular Sciences 23, no. 12: 6731. https://doi.org/10.3390/ijms23126731