
Exogenous Melatonin Enhances Photosynthetic Capacity and Related Gene Expression in A Dose-Dependent Manner in the Tea Plant (Camellia sinensis (L.) Kuntze)

 and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of Exogenous MT Application on the Contents of Chl in Tea Leaves
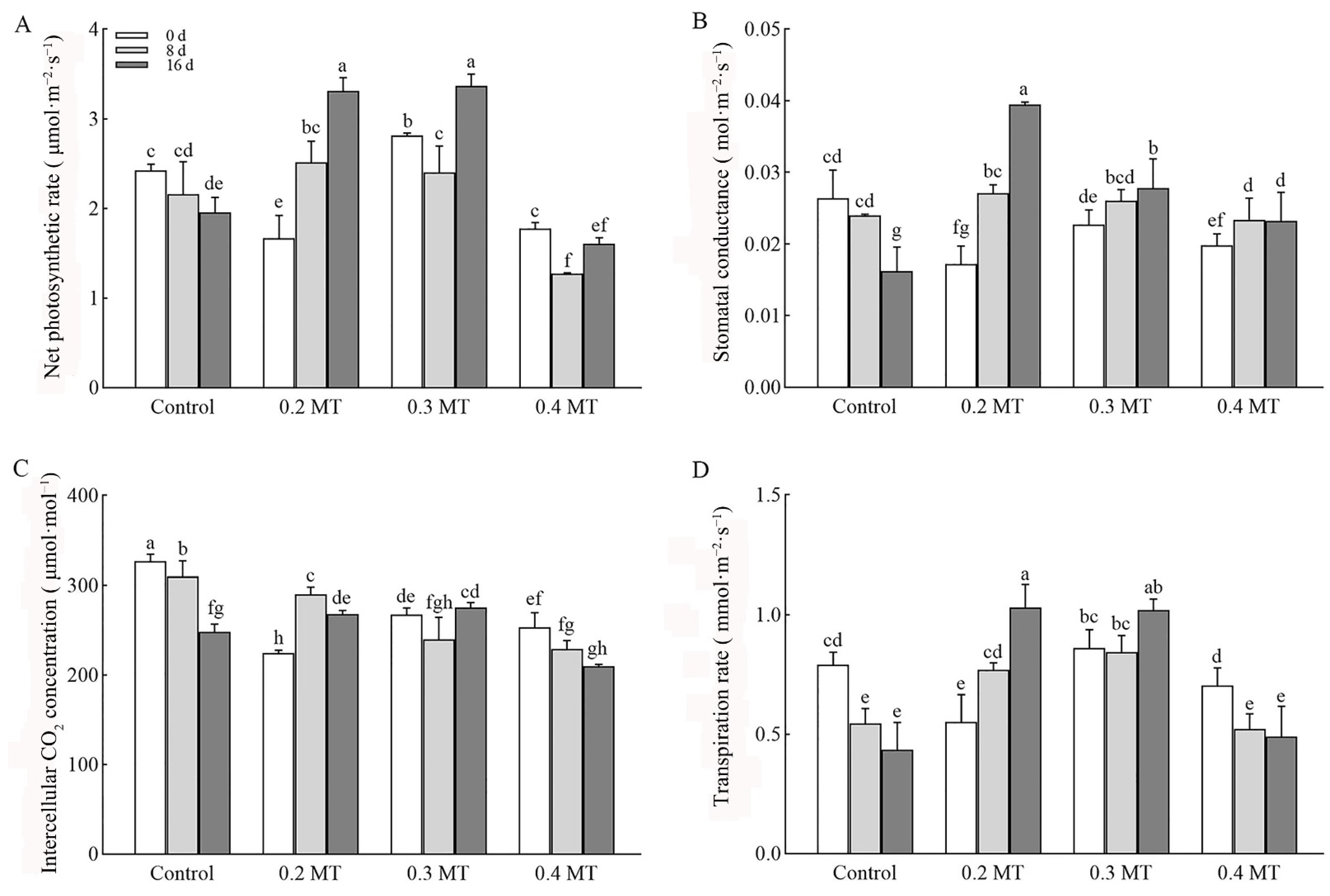
2.2. Effects of Exogenous MT Application on Photosynthetic Parameters in Tea Plants
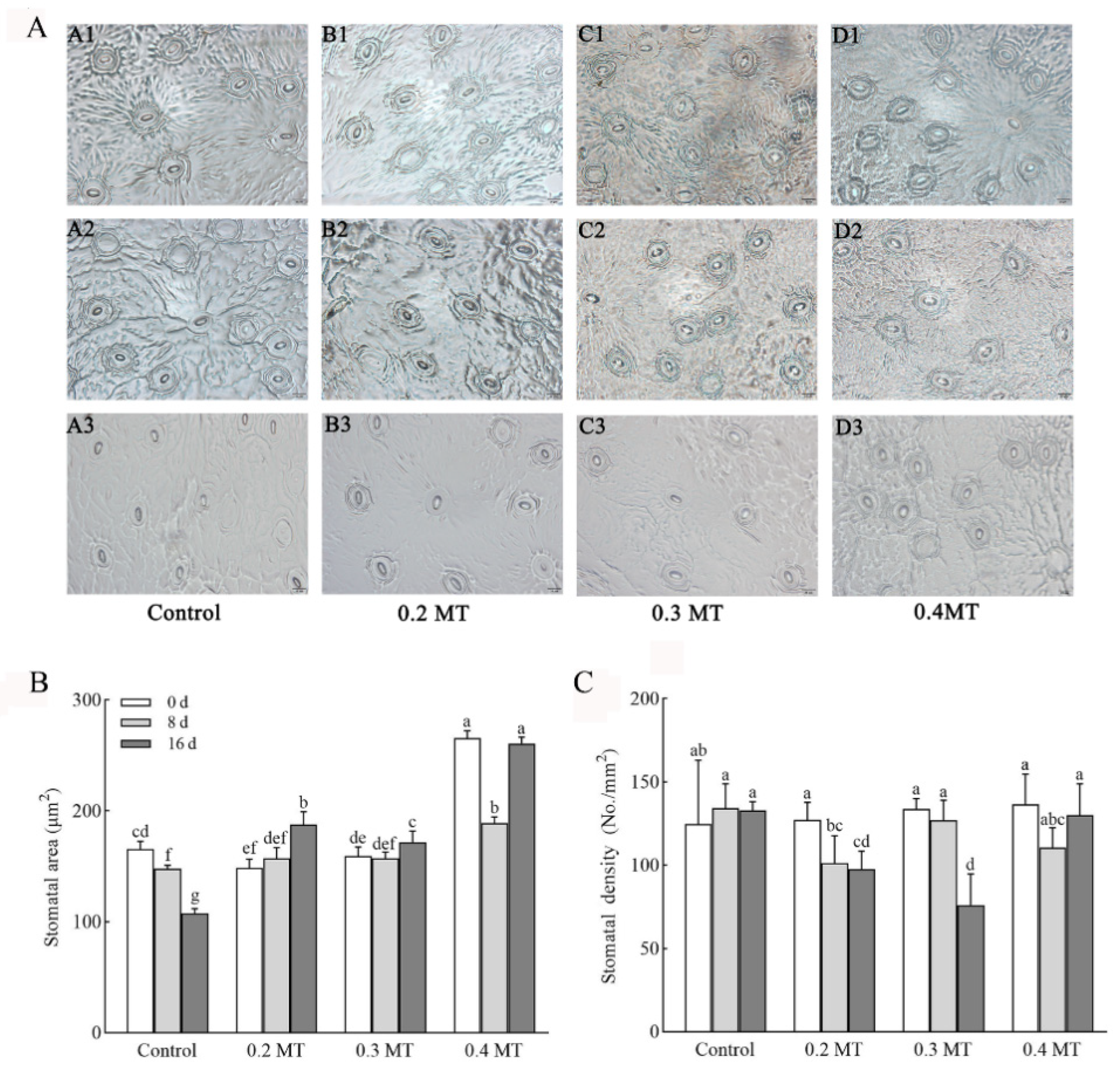
2.3. Effects of Exogenous MT Application on Stomatal Characteristics in Tea Leaves
2.4. Effects of Exogenous MT Application on Chl Fluorescence Parameters in Tea Plants
2.4.1. Effects of Exogenous MT Application on Fv/Fm in Tea Plants
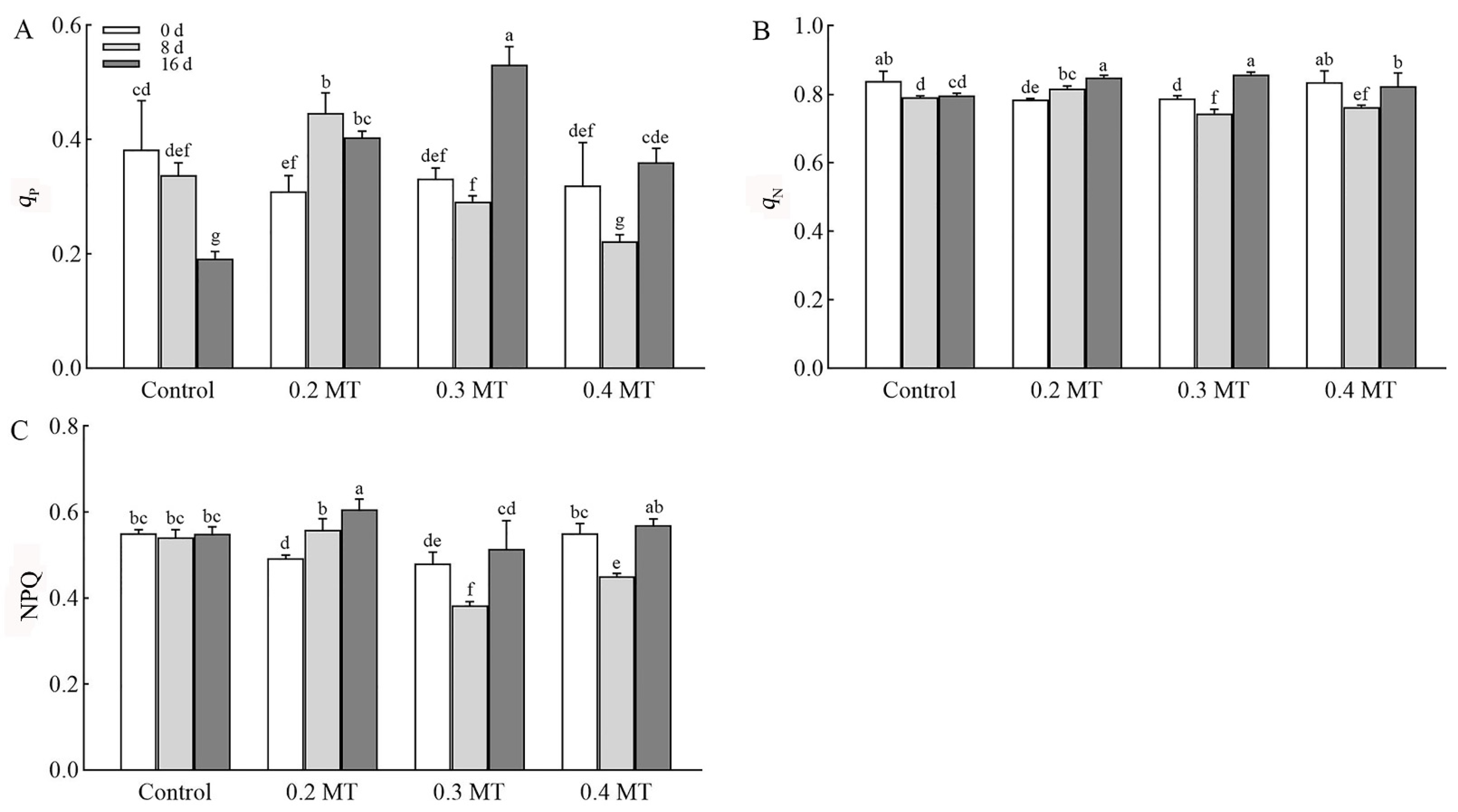
2.4.2. Effects of Exogenous MT Application on Chl Fluorescence Quenching Coefficients
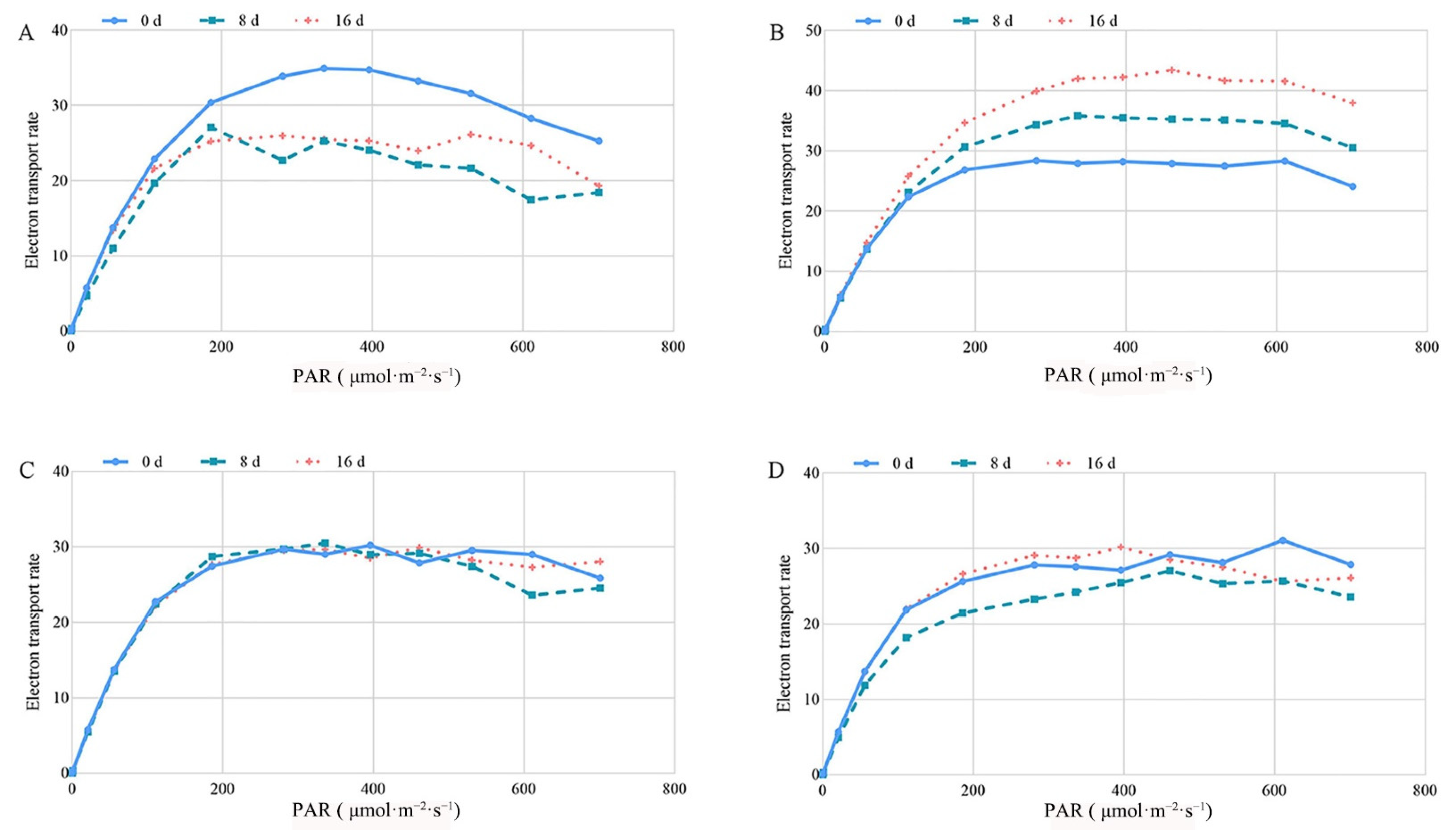
2.4.3. Effects of Exogenous MT Application on the Light Response Curve in Tea Plants
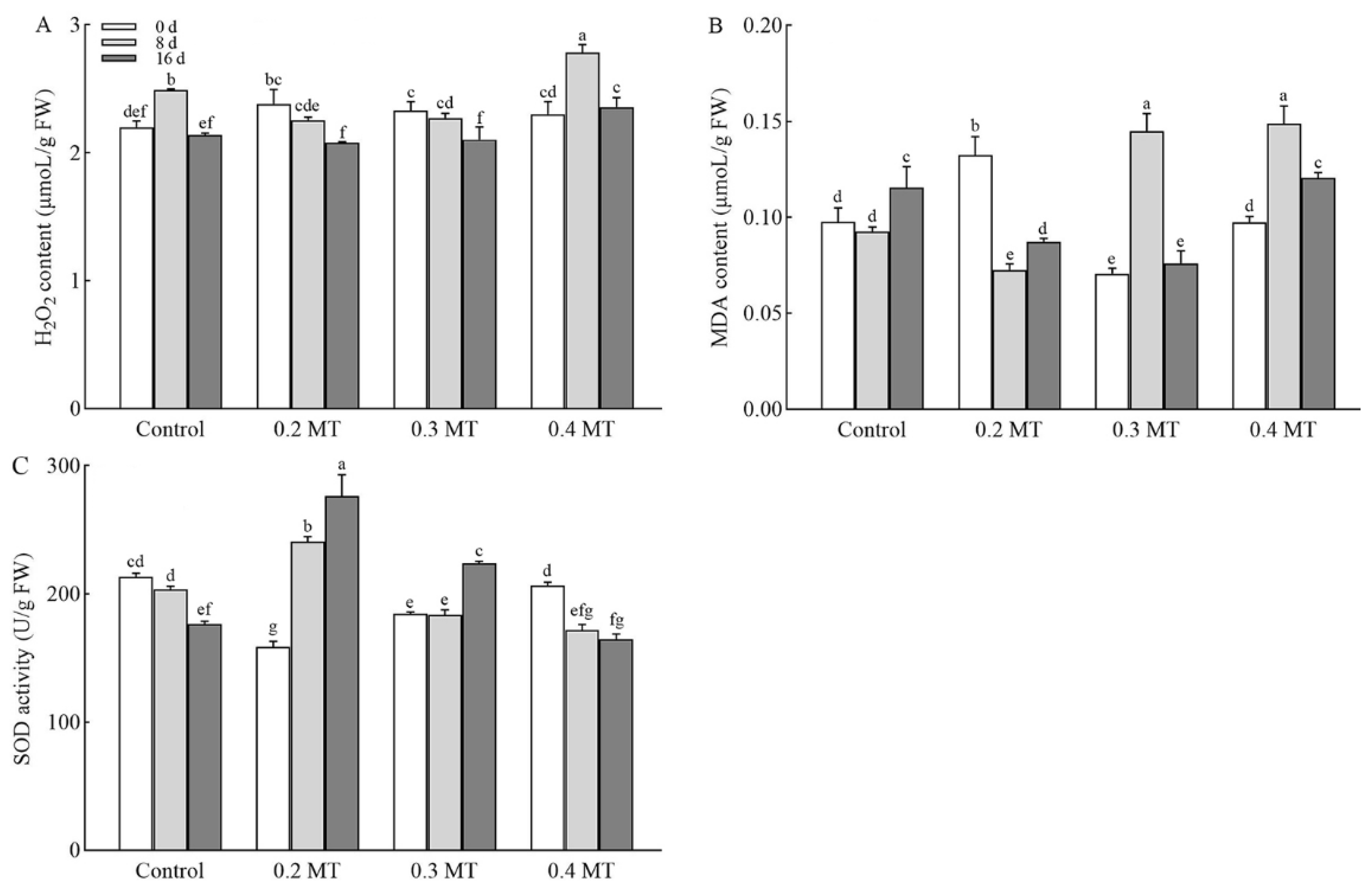
2.5. Effects of Exogenous MT Application on H2O2 and MDA Contents in Tea Plants
2.6. SOD Activity as Influenced by Exogenous MT Application in Tea Leaves
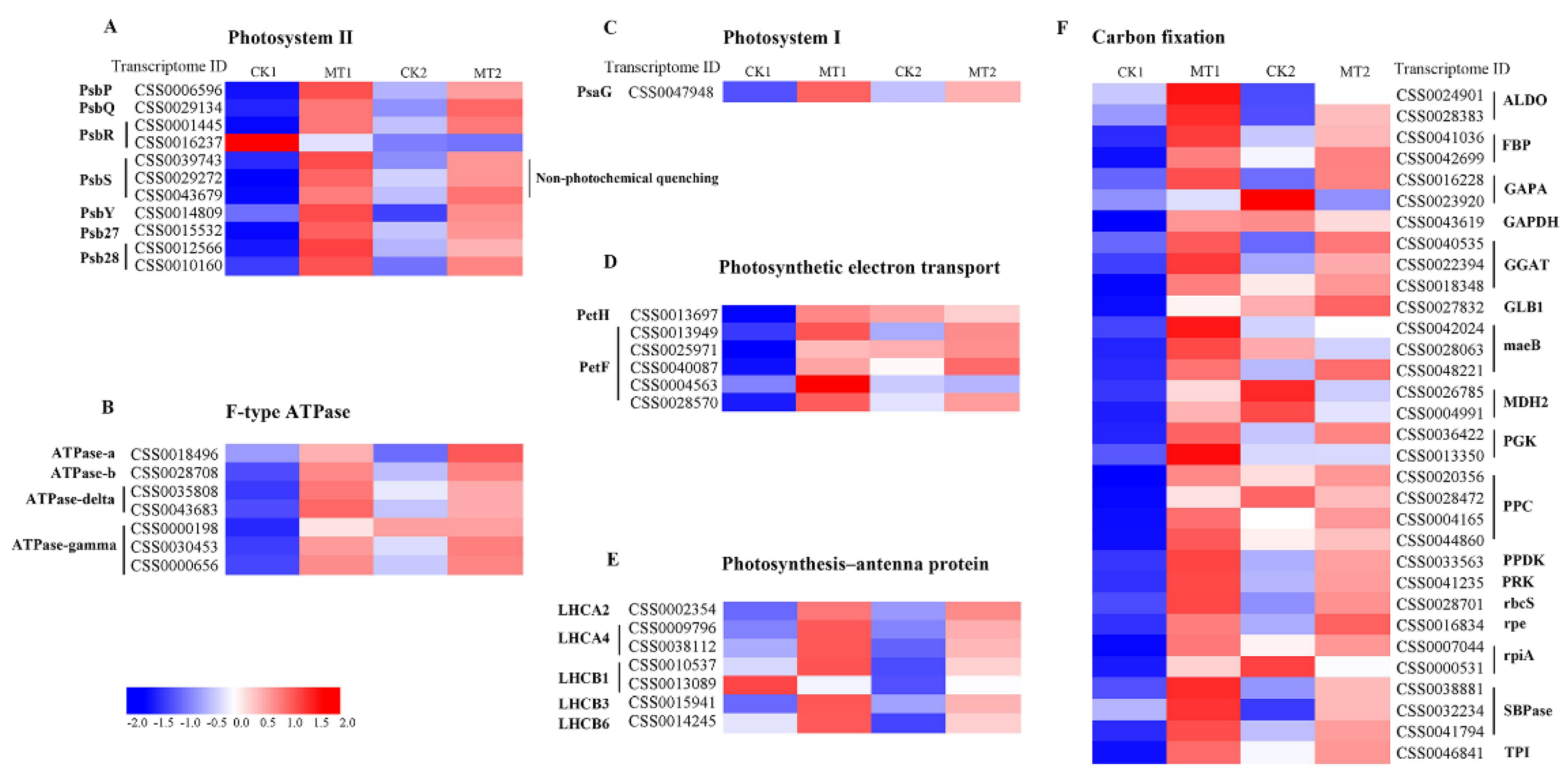
2.7. Transcriptome-Based Identification of DEGs Involved in Photosynthesis under MT Treatments
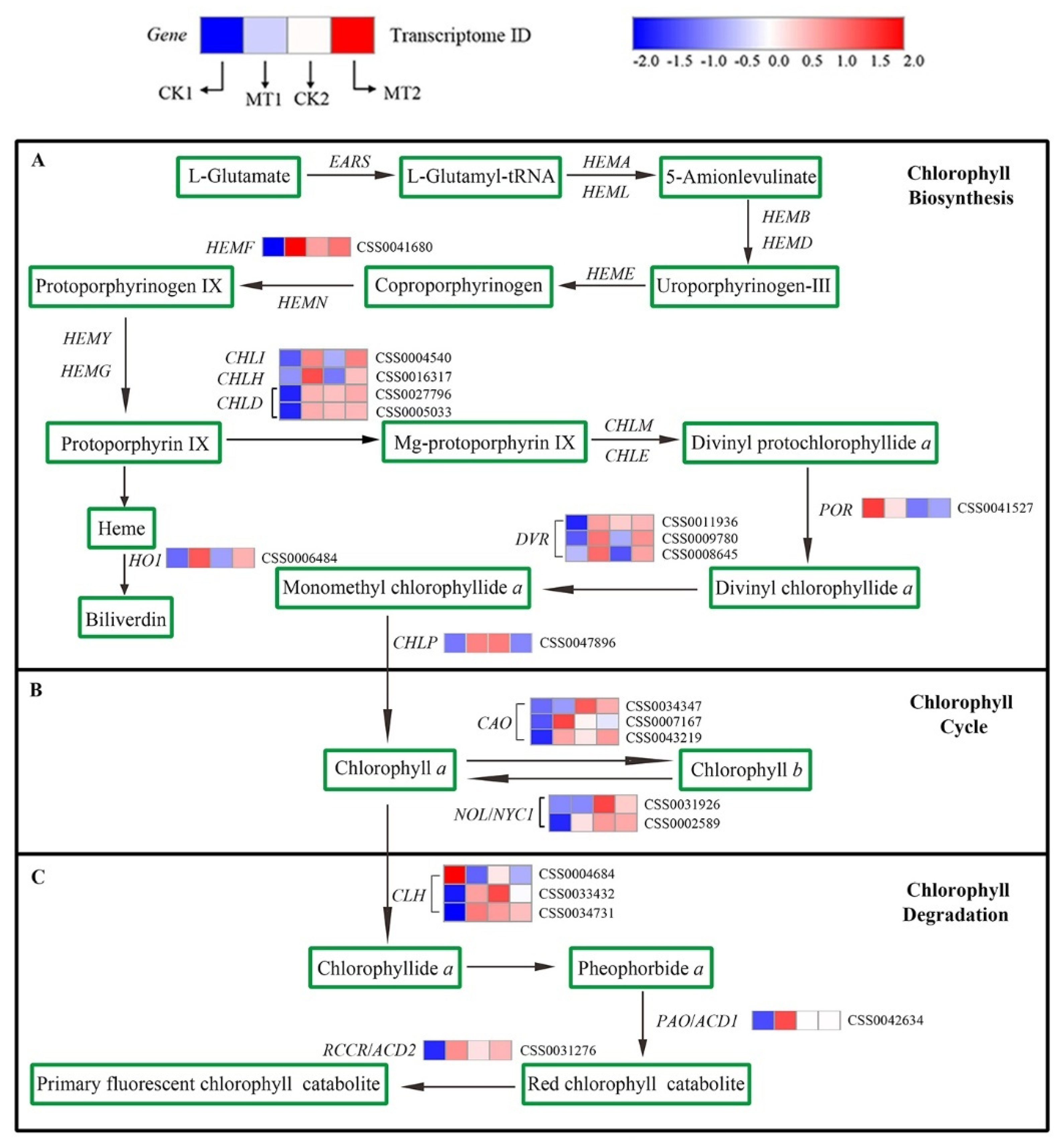
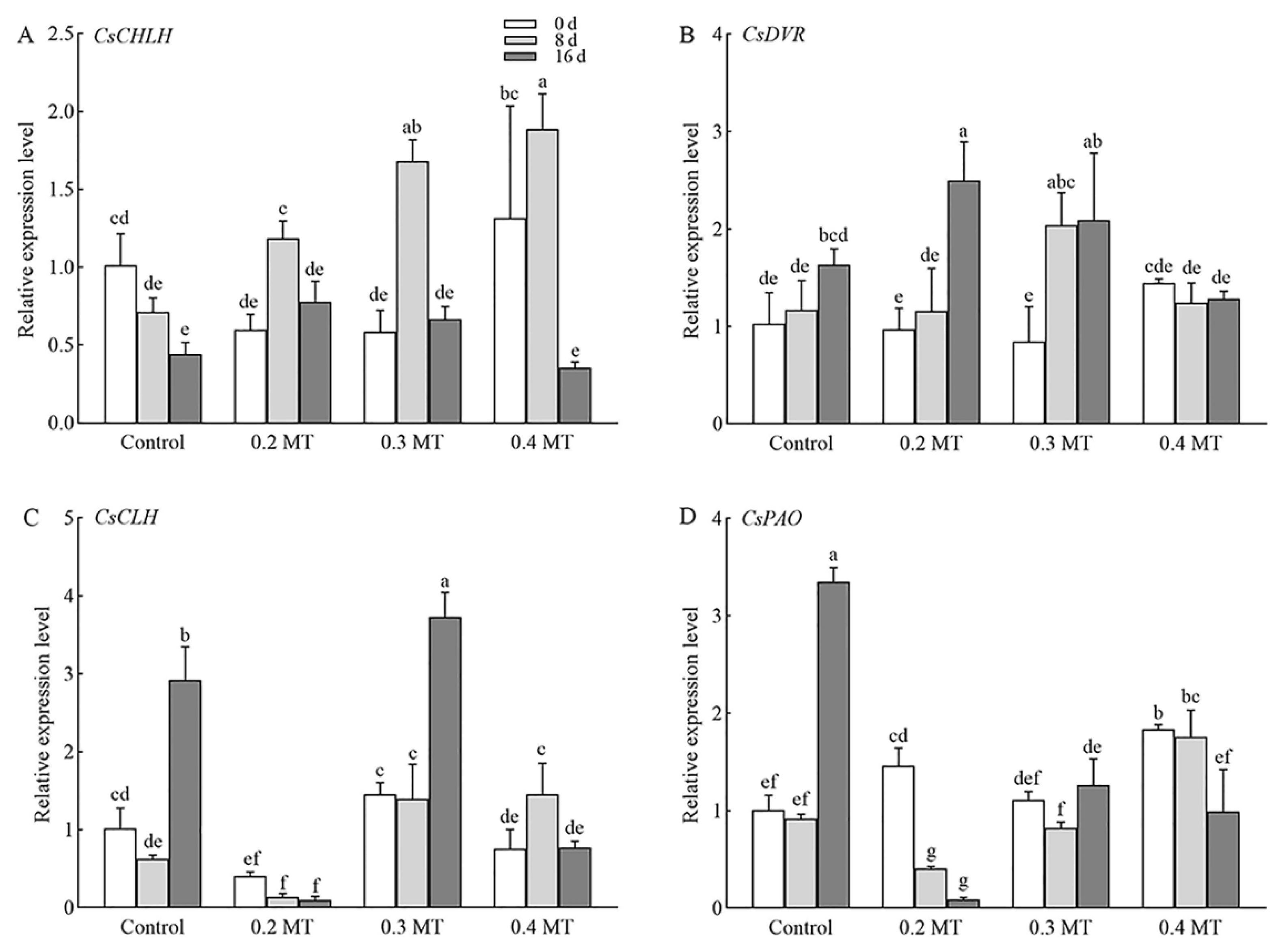
2.8. Transcriptome-Based Identification of DEGs Involved in Chl Metabolism under MT Treatments
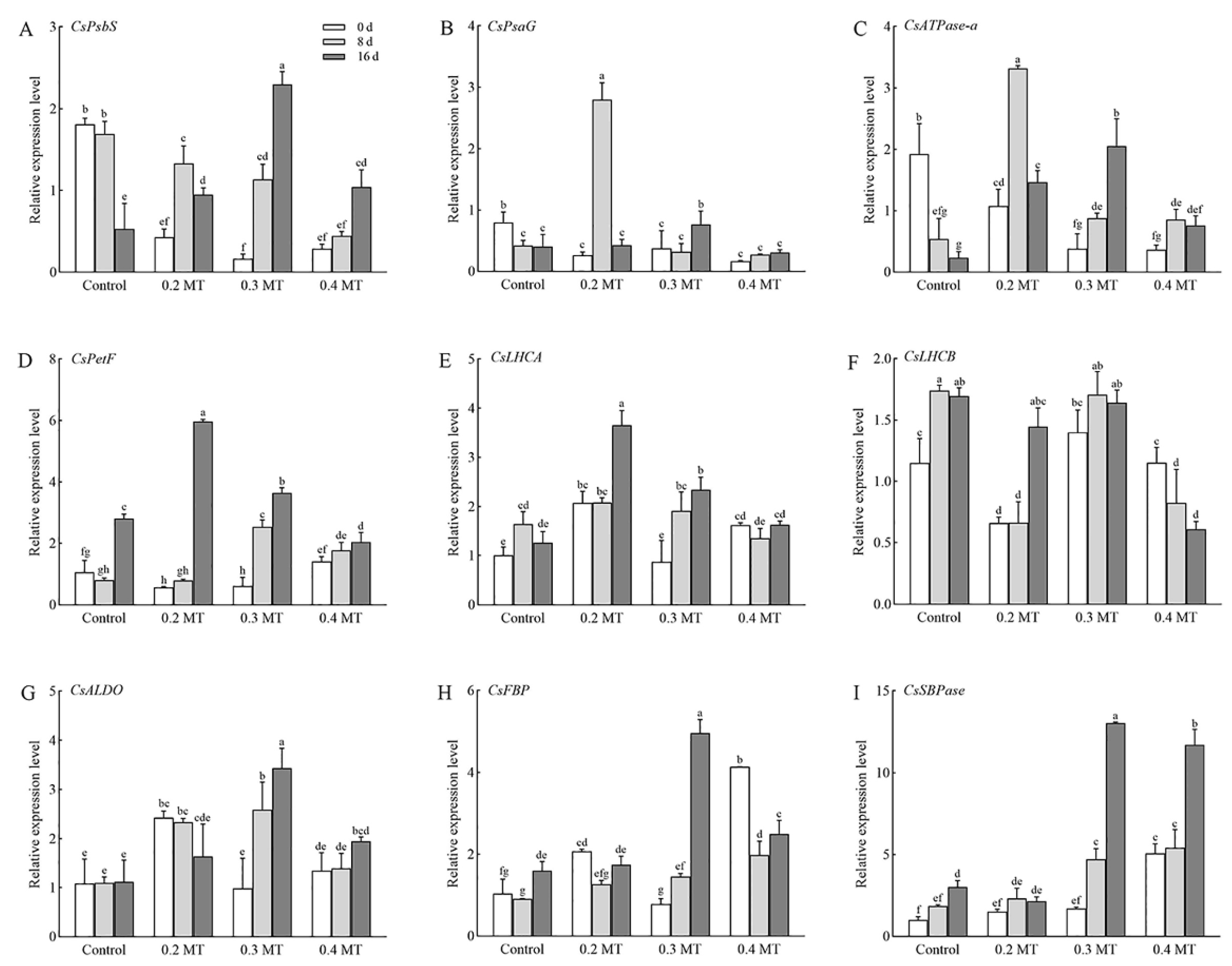
2.9. Expression Profiles of Genes Involved in Photosynthesis and Chl Metabolism in Tea Leaves under MT Treatments
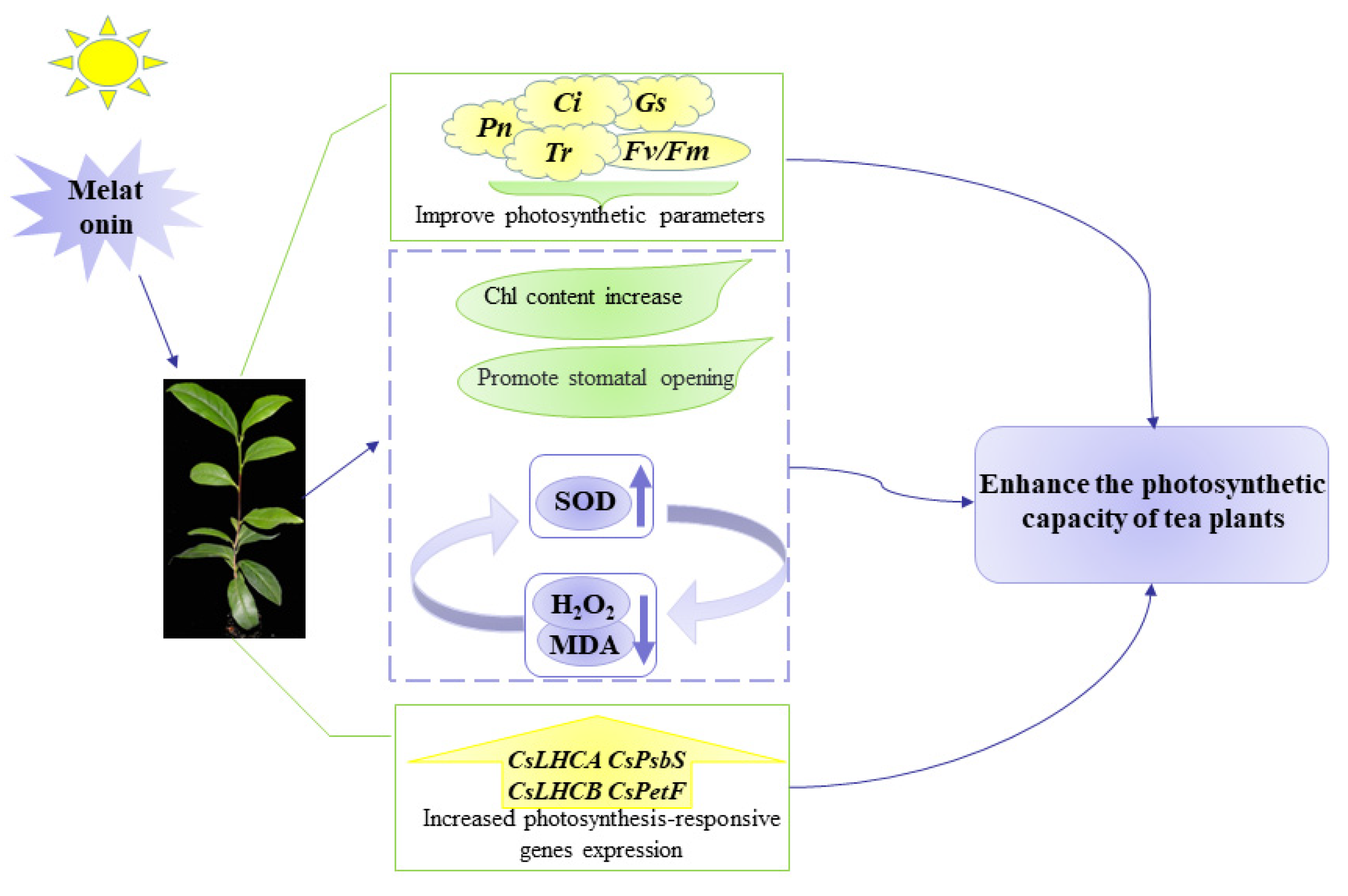
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Exogenous Melatonin Treatments
4.2. Determination of Chl a, b and Total Chl Contents
4.3. Determination of Pn, Gs, Ci, and Tr
4.4. Stoma Observation
4.5. Measurement of Chl Fluorescence Parameters
4.6. Determination of H2O2 and MDA Contents
4.7. Determination of Antioxidant Enzyme (SOD) Activity
4.8. Transcriptome-Based Identification of Chl Biosynthesis and Photosynthesis-Related Differentially Expressed Genes (DEGs)
4.9. RNA Isolation and Gene Expression Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xia, E.H.; Zhang, H.B.; Sheng, J.; Li, K.; Zhang, Q.J.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Li, W.; et al. The tea tree genome provides insights into tea flavor and independent evolution of caffeine biosynthesis. Mol. Plant 2017, 10, 866–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Liu, H.; Wang, Y.; Teng, R.M.; Liu, J.X.; Lin, S.J.; Zhuang, J. Cytosolic ascorbate peroxidase 1 modulates ascorbic acid metabolism through cooperating with nitrogen regulatory protein P-II in tea plant under nitrogen deficiency stress. Genomics 2020, 112, 3497–3503. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, Z.W.; Wu, Z.J.; Wang, X.Y.; Teng, R.M.; Zhuang, J. Differentially expressed protein and gene analysis revealed the effects of temperature on changes in ascorbic acid metabolism in harvested tea leaves. Hortic. Res. 2018, 5, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.W.; Li, H.; Liu, J.X.; Wang, Y.; Zhuang, J. Integrative transcriptome, proteome, and microRNA analysis reveals the effects of nitrogen sufficiency and deficiency conditions on theanine metabolism in the tea plant (Camellia sinensis). Hortic. Res. 2020, 7, 65. [Google Scholar] [CrossRef]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aroma. 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Hu, Q.; Liu, Y.N.; Cheng, P.L.; Cheng, H.; Liu, W.X.; Xing, X.J.; Guan, Z.Y.; Fang, W.M.; Chen, S.M.; et al. Strigolactone represses the synthesis of melatonin, thereby inducing floral transition in Arabidopsis thaliana in an FLC-dependent manner. J. Pineal Res. 2019, 67, e12582. [Google Scholar] [CrossRef]
- Posch, B.C.; Kariyawasam, B.C.; Helen, B.; Onoriode, C.; Richards, R.A.; Reynolds, M.P.; Richard, T.; Atkin, O.K. Exploring high temperature responses of photosynthesis and respiration to improve heat tolerance in wheat. J. Exp. Bot. 2019, 70, 5051–5069. [Google Scholar] [CrossRef]
- Heyneke, E.; Fernie, A.R. Metabolic regulation of photosynthesis. Biochem. Soc. Trans. 2018, 46, 321–328. [Google Scholar] [CrossRef]
- Parry, M.A.J.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef]
- Dubbels, R.; Reiter, R.J.; Klenke, E.; Goebel, A.; Schnakenberg, E.; Ehlers, C.; Schiwara, H.W.; Schloot, W. Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J. Pineal Res. 1995, 18, 28–31. [Google Scholar] [CrossRef]
- Kolář, J.; Johnson, C.H.; Macháčková, I. Exogenously applied melatonin (N-acetyl-5-methoxytryptamine) affects flowering of the short-day plant Chenopodium rubrum. Acta Physiol. Plant. 2003, 118, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Li, Q.T.; Chu, Y.N.; Reiter, R.J.; Yu, X.M.; Zhu, D.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, J.S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef] [Green Version]
- Debnath, B.; Islam, W.; Li, M.; Sun, Y.T.; Lu, X.C.; Mitra, S.; Hussain, M.; Liu, S.; Qiu, D.L. Melatonin mediates enhancement of stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 1040. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Lee, D.E.; Jang, H.; Byeon, Y.; Kim, Y.S.; Back, K. Melatonin-rich transgenic rice plants exhibit resistance to herbicide-induced oxidative stress. J. Pineal Res. 2013, 54, 258–263. [Google Scholar] [CrossRef]
- Gao, W.Y.; Zhang, Y.J.; Feng, Z.; Bai, Q.Q.; He, J.J.; Wang, Y.J. Effects of melatonin on antioxidant capacity in naked oat seedlings under drought stress. Molecules 2018, 23, 1580. [Google Scholar] [CrossRef] [Green Version]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef]
- Wang, L.Y.; Liu, J.L.; Wang, W.X.; Sun, Y. Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress. Photosynthetica 2016, 54, 19–27. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Yang, S.J.; Chen, Y.Y. Effects of melatonin on photosynthetic performance and antioxidants in melon during cold and recovery. Biol. Plant. 2017, 61, 571–578. [Google Scholar] [CrossRef]
- Sharma, A.; Wang, J.F.; Xu, D.B.; Tao, S.C.; Chong, S.L.; Yan, D.L.; Li, Z.; Yuan, H.W.; Zheng, B.S. Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Carya cathayensis plants. Sci. Total Environ. 2020, 713, 136675. [Google Scholar] [CrossRef]
- Wang, P.; Yin, L.H.; Liang, D.; Li, C.; Ma, F.W.; Yue, Z.Y. Delayed senescence of apple leaves by exogenous melatonin treatment: Toward regulating the ascorbate-glutathione cycle. J. Pineal Res. 2012, 53, 11–20. [Google Scholar] [CrossRef]
- Lazar, D.; Murch, S.J.; Beilby, M.J.; Khazaaly, S.A.l. Exogenous melatonin affects photosynthesis in characeae Chara australis. Plant Signal. Behav. 2013, 8, 23279. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yue, C.; Cao, H.L.; Zhou, Y.H.; Zeng, J.M.; Yang, Y.J.; Wang, X.C. Biochemical and transcriptome analyses of a novel chlorophyll-deficient chlorina tea plant cultivar. BMC Plant Biol. 2014, 14, 352. [Google Scholar] [CrossRef] [Green Version]
- Mir, A.R.; Faizan, M.; Bajguz, A.; Sami, F.; Siddiqui, H.; Hayat, S. Occurrence and biosynthesis of melatonin and its exogenous effect on plants. Acta Soc. Bot. Pol. 2020, 89, 1–23. [Google Scholar] [CrossRef]
- Wang, Y.C.; Gao, C.Q.; Liang, Y.N.; Wang, C.; Yang, C.P.; Liu, G.F. A novel bZIP gene from Tamarix hispida mediates physiological responses to salt stress in tobacco plants. J. Plant Physiol. 2010, 167, 222–230. [Google Scholar] [CrossRef]
- Liang, C.Z.; Zheng, G.Y.; Li, W.Z.; Wang, Y.Q.; Hu, B.; Wang, H.R.; Wu, H.K.; Qian, Y.W.; Zhu, X.G.; Tan, D.X.; et al. Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef]
- Wang, P.; Sun, X.; Li, C.; Wei, Z.W.; Liang, D.; Ma, F.W. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. Pineal Res. 2013, 54, 292–302. [Google Scholar] [CrossRef]
- Kuai, B.K.; Chen, J.Y.; Hrtensteiner, S. The biochemistry and molecular biology of chlorophyll breakdown. Environ. Exp. Bot. 2018, 69, 751–767. [Google Scholar] [CrossRef]
- Roca, M.; Pérez-Gálvez, A. Profile of chlorophyll catabolites in senescent leaves of epipremnun aureum includes a catabolite esterified with hydroxytyrosol 1- O -Glucoside. J. Nat. Prod. 2020, 83, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Nilson, S.E.; Assmann, S.M. The control of transpiration. insights from Arabidopsis. Plant Physiol. 2007, 143, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.C.; Laurence, J.A. The response of clones of populus tremuloides differing in sensitivity to ozone in the field. New Phytol. 1999, 141, 411–421. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Mohotti, A.J.; Lawlor, D.W. Diurnal variation of photosynthesis and photoinhibition in tea: Effects of irradiance and nitrogen supply during growth in the field. J. Exp. Bot. 2002, 53, 313–322. [Google Scholar] [CrossRef]
- Lawlor, D.W. Photosynthesis, productivity and environment. J. Exp. Bot. 1995, 46, 1449–1461. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Yu, Q.; Xu, C.Y.; Li, J.H.; Qin, G.M. Rapid estimation of stomatal density and stomatal area of plant leaves based on object-oriented classification and its ecological trade-off strategy analysis. Forests 2018, 9, 616. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.Z.; Zhang, D.Y.; Zhou, L.; Zhang, X.Z.; Liao, J.R.; Duan, Y.; Wen, B.; Ma, Y.C.; Wang, Y.H.; Fang, W.P.; et al. Transcriptomic and metabolomic profiling of Camellia sinensis L. cv. ‘Suchazao’ exposed to temperature stresses reveals modification in protein synthesis and photosynthetic and anthocyanin biosynthetic pathways. Tree Physiol. 2019, 39, 1583–1599. [Google Scholar] [CrossRef]
- Roach, T.; Krieger-Liszkay, A. The role of the PsbS protein in the protection of photosystems I and II against high light in Arabidopsis thaliana. BBA Bioenerg. 2012, 1817, 2158–2165. [Google Scholar] [CrossRef] [Green Version]
- Palm, D.M.; Agostini, A.; Averesch, V.; Girr, P.; Werwie, M.; Takahashi, S.; Satoh, H.; Jaenicke, E.; Paulsen, H. Chlorophyll a/b binding-specificity in water-soluble chlorophyll protein. Nat. Plants 2018, 4, 920–929. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Ni, Z.Y.; Xia, H.; Xie, Y.; Lv, X.L.; Wang, J.; Lin, L.J.; Deng, Q.X.; Luo, X. Exogenous melatonin promotes biomass accumulation and photosynthesis of kiwifruit seedlings under drought stress. Sci. Hortic. 2019, 246, 34–43. [Google Scholar] [CrossRef]
- Wu, S.Q.; Wang, Y.; Zhang, J.K.; Gong, X.J.; Zhang, Z.; Sun, J.J.; Chen, X.S.; Wang, Y.L. Exogenous melatonin improves physiological characteristics and promotes growth of strawberry seedlings under cadmium stress. Hortic. Plant J. 2020, 7, 13–22. [Google Scholar] [CrossRef]
- Zhuang, K.Y.; Kong, F.Y.; Zhang, S.; Meng, C.; Yang, M.M.; Liu, Z.B.; Wang, Y.; Ma, N.; Meng, Q.W. Whirly1 enhances tolerance to chilling stress in tomato via protection of photosystem II and regulation of starch degradation. New Phytol. 2019, 221, 1998–2012. [Google Scholar] [CrossRef]
- Fan, J.B.; Hu, Z.R.; Xie, Y.; Chan, Z.L.; Chen, K.; Amombo, E.; Chen, L.; Fu, J.M. Alleviation of cold damage to photosystem II and metabolisms by melatonin in Bermudagrass. Front. Plant Sci. 2014, 6, 925. [Google Scholar] [CrossRef] [Green Version]
- Aghdam, M.S.; Moradi, M.; Razavi, F.; Rabiei, V. Exogenous phenylalanine application promotes chilling tolerance in tomato fruits during cold storage by ensuring supply of NADPH for activation of ROS scavenging systems. Sci. Hortic. 2019, 246, 818–825. [Google Scholar] [CrossRef]
- Haskirli, H.; Ylmaz, O.; Ozgur, R.; Uzilday, B.; Turkan, I. Melatonin mitigates UV-B stress via regulating oxidative stress response, cellular redox and alternative electron sinks in Arabidopsis thaliana. Phytochemistry 2020, 182, 112592. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Jannatizadeh, A.; Nojadeh, M.S.; Ebrahimzadeh, A. Exogenous melatonin ameliorates chilling injury in cut anthurium flowers during low temperature storage. Postharvest Biol. Technol. 2019, 148, 184–191. [Google Scholar] [CrossRef]
- Li, X.; Wei, J.P.; Scott, E.R.; Liu, J.W.; Guo, S.; Li, Y.; Zhang, L.; Han, W.Y. Exogenous melatonin alleviates cold stress by promoting antioxidant defense and redox homeostasis in Camellia sinensis L. Molecules 2018, 23, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.C.; Feng, H.; Chang, Y.X.; Ma, C.L.; Wang, L.Y.; Hao, X.Y.; Li, A.L.; Cheng, H.; Wang, L.; Cui, P.; et al. Population sequencing enhances understanding of tea plant evolution. Nat. Commun. 2020, 11, 4447. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Feng, K.; Hou, X.L.; Jiang, Q.; Xu, Z.S.; Wang, G.L.; Liu, J.X.; Wang, F.; Xiong, A.S. The genome sequence of celery (Apium graveolens L.), an important leaf vegetable crop rich in apigenin in the Apiaceae family. Hortic. Res. 2020, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, E.H.; Li, F.D.; Tong, W.; Li, P.H.; Wu, Q.; Zhao, H.J.; Ge, R.H.; Li, R.P.; Li, Y.Y.; Zhang, Z.Z.; et al. Tea Plant Information Archive: A comprehensive genomics and bioinformatics platform for tea plant. Plant Biotechnol. J. 2019, 17, 1938–1953. [Google Scholar] [CrossRef]
- Liu, J.X.; Jiang, Q.; Tao, J.P.; Feng, K.; Li, T.; Duan, A.Q.; Wang, H.; Xu, Z.S.; Liu, H.; Xiong, A.S. Integrative genome, transcriptome, microRNA, and degradome analysis of water dropwort (Oenanthe javanica) in response to water stress. Hortic. Res. 2021, 8, 262. [Google Scholar] [CrossRef]
- Yang, N.; Wan, Q.W.; Li, Y.M.; Han, M.H.; Teng, R.M.; Liu, J.X.; Zhuang, J. Effects of exogenous spermidine on photosynthetic characteristics and gene expression of key enzymes under salt stress in tea plant. Acta Hortic. Sinica 2022, 49, 378–394. [Google Scholar]
- Li, T.; Huang, Y.; Khadr, A.; Wang, Y.H.; Xu, Z.S.; Xiong, A.S. DcDREB1A, a DREB-binding transcription factor from Daucus carota, enhances drought tolerance in transgenic Arabidopsis thaliana and modulates lignin levels by regulating lignin-biosynthesis-related genes. Environ. Exp. Bot. 2020, 169, 103896. [Google Scholar] [CrossRef]
- Li, J.H.; Yang, Y.Q.; Sun, K.; Chen, Y.; Chen, X.; Li, X.H. Exogenous melatonin enhances cold, salt and drought stress tolerance by improving antioxidant defense in tea plant [Camellia sinensis (L.) O. Kuntze]. Molecules 2019, 24, 1826. [Google Scholar] [CrossRef] [Green Version]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Han, M.H.; Yang, N.; Wan, Q.W.; Teng, R.M.; Duan, A.Q.; Wang, Y.H.; Zhuang, J. Exogenous melatonin positively regulates lignin biosynthesis in Camellia sinensis. Int. J. Biol. Macromol. 2021, 179, 485–499. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wu, Z.J.; Tian, C.; Jiang, Q.; Li, X.H.; Zhuang, J. Selection of suitable reference genes for qRT-PCR normalization during leaf development and hormonal stimuli in tea plant (Camellia sinensis). Sci. Rep. 2016, 6, 19748. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Chl a Content (mg/g FW) | Chl b Content (mg/g FW) | Total Chl Content (mg/g FW) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 8 | Day 16 | Day 0 | Day 8 | Day 16 | Day 0 | Day 8 | Day 16 | |
| Control | 1.073 ± 0.008 b | 0.967 ± 0.020 d | 0.975 ± 0.042 d | 0.593 ± 0.004 c | 0.497 ± 0.004 f | 0.427 ± 0.005 g | 1.667 ± 0.001 b | 1.464 ± 0.018 f | 1.403 ± 0.046 g |
| 0.2 MT | 0.980 ± 0.031 d | 1.072 ± 0.001 b | 1.115 ± 0.013 a | 0.388 ± 0.006 g | 0.550 ± 0.036 de | 0.718 ± 0.012 a | 1.368 ± 0.026 gh | 1.621 ± 0.035 c | 1.832 ± 0.025 a |
| 0.3 MT | 1.037 ± 0.002 c | 0.867 ± 0.007 e | 1.089 ± 0.010 ab | 0.477 ± 0.009 f | 0.473 ± 0.013 f | 0.718 ± 0.012 a | 1.515 ± 0.007 e | 1.340 ± 0.006 h | 1.807 ± 0.002 a |
| 0.4 MT | 1.099 ± 0.006 ab | 1.017 ± 0.005 c | 0.896 ± 0.009 e | 0.532 ± 0.011 e | 0.562 ± 0.007 d | 0.680 ± 0.016 b | 1.631 ± 0.004 bc | 1.579 ± 0.001 d | 1.576 ± 0.011 d |
| Treatments | Fo | Fm | Fv/Fm | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 8 | Day 16 | Day 0 | Day 8 | Day 16 | Day 0 | Day 8 | Day 16 | |
| Control | 0.109 ± 0.005 e | 0.141 ± 0.011 a | 0.142 ± 0.005 a | 0.516 ± 0.020 e | 0.688 ± 0.043 a | 0.629 ± 0.034 b | 0.789 ± 0.002 ab | 0.795 ± 0.003 a | 0.775 ± 0.005 cde |
| 0.2 MT | 0.133 ± 0.004 bc | 0.129 ± 0.009 bcd | 0.125 ± 0.005 d | 0.580 ± 0.014 cd | 0.595 ± 0.038 cd | 0.602 ± 0.019 c | 0.771 ± 0.002 e | 0.783 ± 0.002 bcd | 0.793 ± 0.001 a |
| 0.3 MT | 0.127 ± 0.007 cd | 0.133 ± 0.013 bc | 0.141 ± 0.007 a | 0.567 ± 0.025 d | 0.588 ± 0.032 cd | 0.591 ± 0.023 cd | 0.776 ± 0.004 de | 0.775 ± 0.012 de | 0.761 ± 0.006 f |
| 0.4 MT | 0.111 ± 0.005 e | 0.135 ± 0.009 ab | 0.135 ± 0.008 ab | 0.517 ± 0.023 e | 0.521 ± 0.052 e | 0.527 ± 0.032 e | 0.786 ± 0.001 abc | 0.738 ± 0.032 g | 0.743 ± 0.008 g |
| Transcriptome ID | Gene Symbol | Molecular Function | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|---|
| CSS0047948 | CsPsaG | Photosystem I subunit G | GTCATACCGTTCTGTTCG | GCACTGGTCTGTCCCT |
| CSS0043679 | CsPsbS | Photosystem II subunit S | TCTCACATCTTACTACCTCCAC | AAACCCAATCCCACCC |
| CSS0018496 | CsATPase-a | F-type H+-transporting ATPase subunit a | ATCAATACTCGCTTCGC | CATCCTGGTCTTCCTCT |
| CSS0040087 | CsPetF | Ferredoxin | TGGGATGATGGTTTCTG | TGAGCACGTACCCTTTC |
| CSS0038112 | CsLHCA | Light-harvesting complex I chlorophyll a/b binding protein | GGACCCTGAGAACCTAAA | GACGATGACGCGAAGTA |
| CSS0010537 | CsLHCB | Light-harvesting complex II chlorophyll a/b binding protein | CGGAAAGGCGGTGAAA | CAGACAATGGACCCAAATAC |
| CSS0028383 | CsALDO | Fructose-bisphosphate aldolase | AAAGCGAGGAAGAGGC | CCGAATGAGAACGAAAGT |
| CSS0042699 | CsFBP | Fructose-bisphosphatase | GTATTTCAATGGCGTGTA | TTATCCCTGTCCTCCC |
| CSS0032234 | CsSBPase | Sedoheptulose-bisphosphatase | TGAGGCTGCTGTTTGA | ATCGGATAATCTCGTTCT |
| CSS0016317 | CsCHLH | Magnesium chelatase H subunit | GCTGCTTACTACTCGTTT | CTCATTCCCACCTGCT |
| CSS0008645 | CsDVR | Divinyl chlorophyllide a 8-vinyl-reductase | TGGGTGGACAGGTTGA | CACAGTCTGCGATAAATG |
| CSS0004684 | CsCLH | Chlorophyllase | ATCATCACCACCTAAACC | CTGAGGAGCAACGACTA |
| CSS0042634 | CsPAO | Pheophorbide a oxygenase | GACTTGACCCTGTTGC | AAGCCAGTACCGATGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, N.; Han, M.-H.; Teng, R.-M.; Yang, Y.-Z.; Wang, Y.-H.; Xiong, A.-S.; Zhuang, J. Exogenous Melatonin Enhances Photosynthetic Capacity and Related Gene Expression in A Dose-Dependent Manner in the Tea Plant (Camellia sinensis (L.) Kuntze). Int. J. Mol. Sci. 2022, 23, 6694. https://doi.org/10.3390/ijms23126694
Yang N, Han M-H, Teng R-M, Yang Y-Z, Wang Y-H, Xiong A-S, Zhuang J. Exogenous Melatonin Enhances Photosynthetic Capacity and Related Gene Expression in A Dose-Dependent Manner in the Tea Plant (Camellia sinensis (L.) Kuntze). International Journal of Molecular Sciences. 2022; 23(12):6694. https://doi.org/10.3390/ijms23126694
Chicago/Turabian StyleYang, Ni, Miao-Hua Han, Rui-Min Teng, Ya-Zhuo Yang, Ya-Hui Wang, Ai-Sheng Xiong, and Jing Zhuang. 2022. "Exogenous Melatonin Enhances Photosynthetic Capacity and Related Gene Expression in A Dose-Dependent Manner in the Tea Plant (Camellia sinensis (L.) Kuntze)" International Journal of Molecular Sciences 23, no. 12: 6694. https://doi.org/10.3390/ijms23126694