
Hydrogen Peroxide Stimulates Dihydrotestosterone Release in C2C12 Myotubes: A New Perspective for Exercise-Related Muscle Steroidogenesis?

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
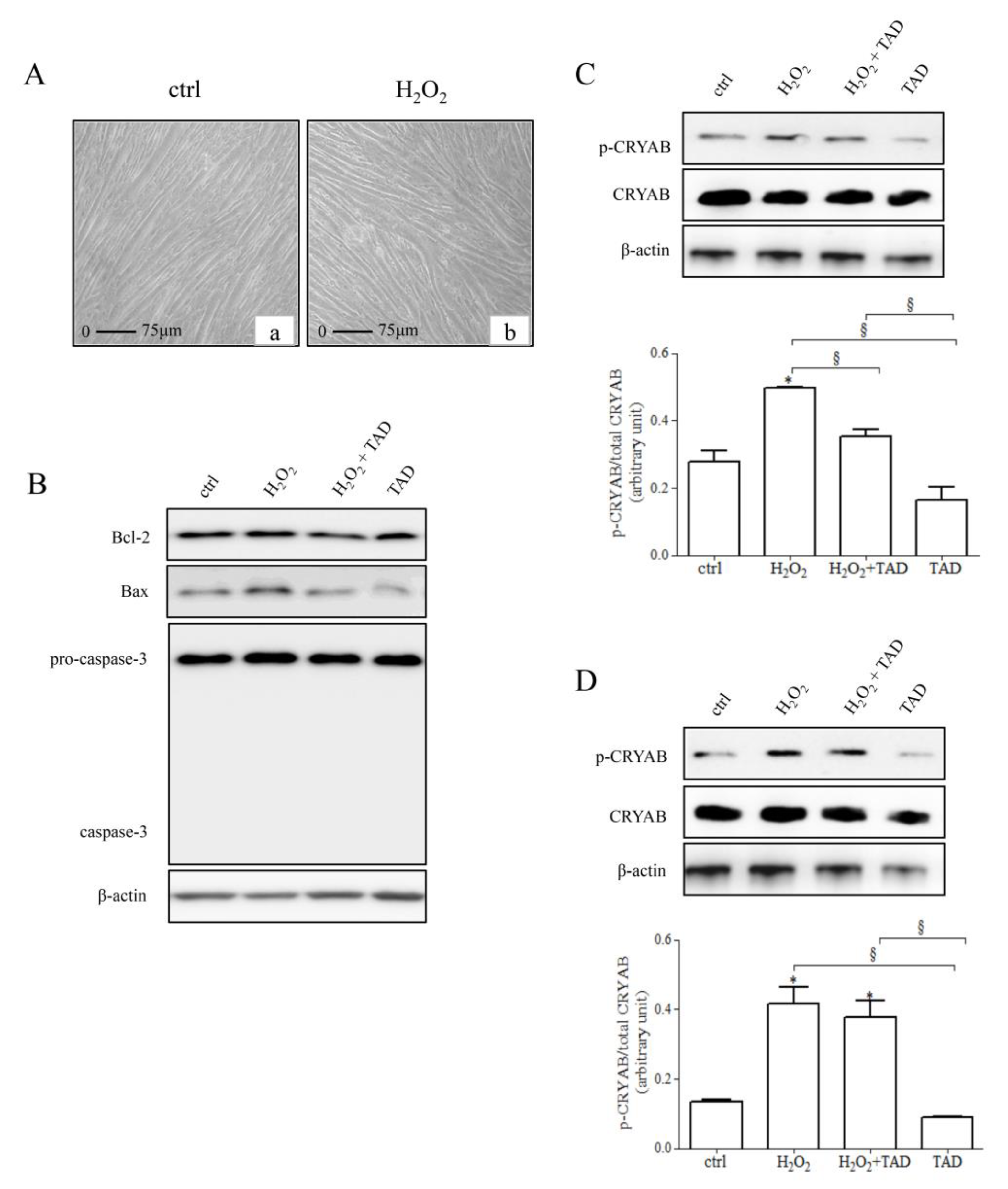
2.1. Analysis of Stress Response Protein and Apoptotic Markers
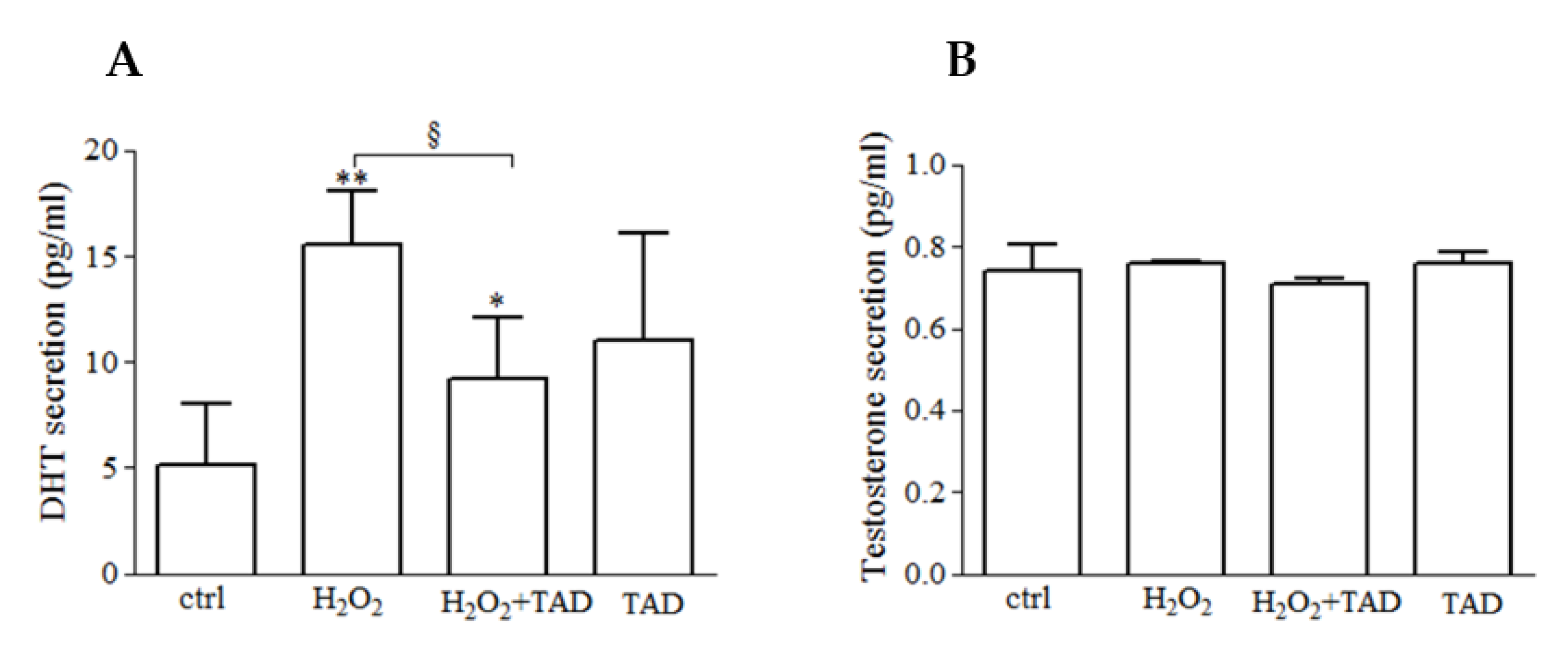
2.2. Dihydrotestosterone (DHT) and Testosterone (T) Release
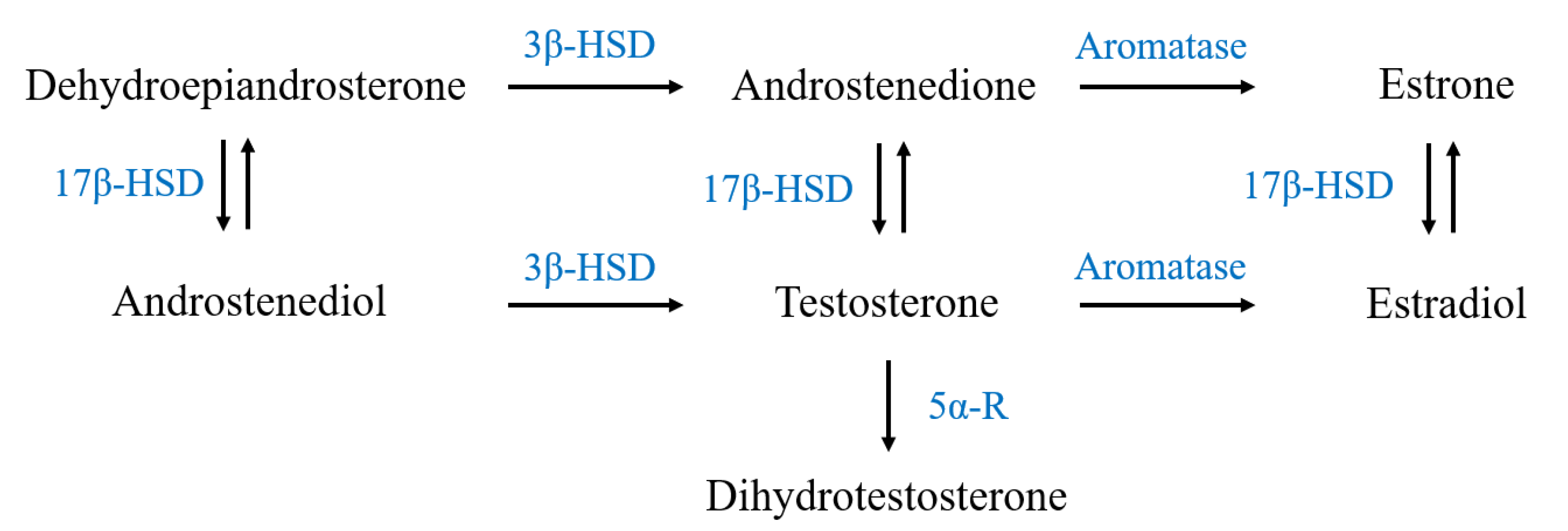
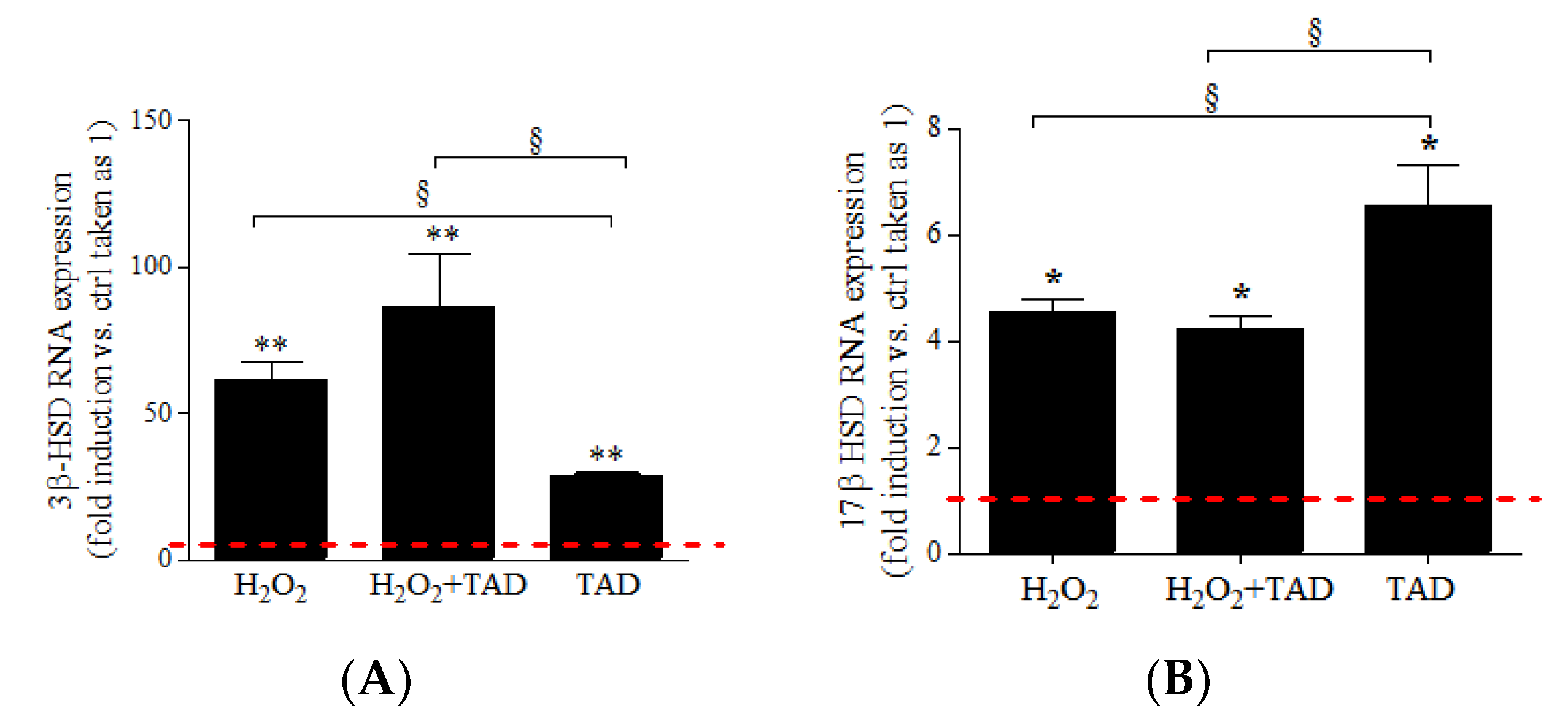
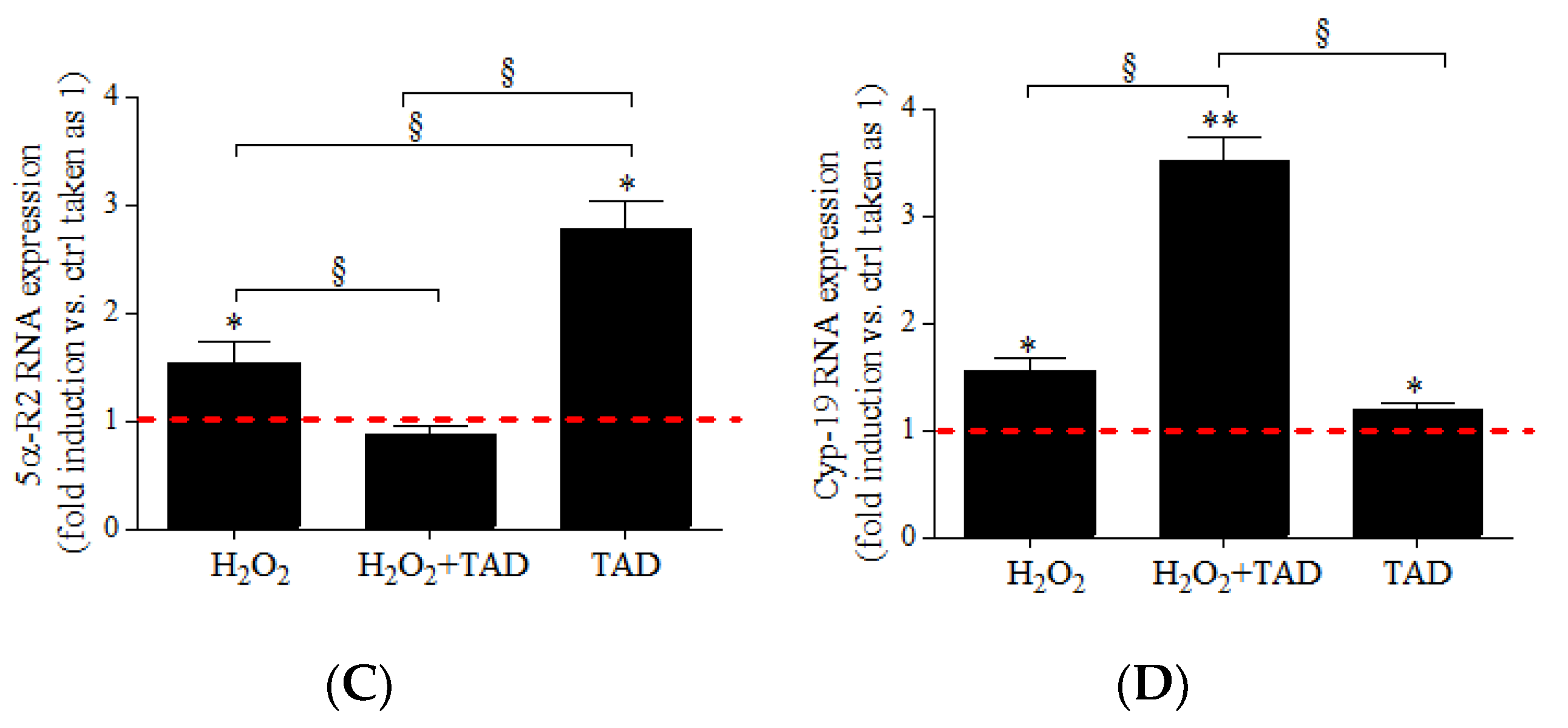
2.3. mRNA Expression of Steroidogenic Enzymes
3. Discussion
3.1. H2O2 Induces the Release of DHT but Not Testosterone
3.2. H2O2 Modulates the Expression of Steroidogenic Enzymes
3.3. TAD Influences Steroidogenesis in C2C12 Myotubes
4. Conclusions
5. Materials and Methods
5.1. Cell Culture
5.2. Cell Treatments
5.3. Morphological Imaging
5.4. Protein Expression Analysis
5.5. Testosterone and Dihydrotestosterone Levels
5.6. RNA Extraction, Reverse Transcription, and Real-Time Quantitative PCR
5.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mendelson, C.; Dufau, M.; Catt, K. Gonadotropin binding and stimulation of cyclic adenosine 3’:5’-monophosphate and testosterone production in isolated Leydig cells. J. Biol. Chem. 1975, 250, 8818–8823. [Google Scholar] [CrossRef]
- Mendelson, C.; Dufau, M.; Catt, K. Dependence of gonadotropin-induced steroidogenesis upon RNA and protein synthesis in the interstitial cells of the rat testis. Biochim. Biophys. Acta 1975, 411, 222–230. [Google Scholar] [CrossRef]
- Sato, K.; Iemitsu, M.; Matsutani, K.; Kurihara, T.; Hamaoka, T.; Fujita, S. Resistance training restores muscle sex steroid hormone steroidogenesis in older men. FASEB J. 2014, 28, 1891–1897. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Iemitsu, M. Exercise and sex steroid hormones in skeletal muscle. J. Steroid Biochem. Mol. Biol. 2015, 145, 200–205. [Google Scholar] [CrossRef]
- Foradori, C.; Weiser, M.; Handa, R. Non-genomic actions of androgens. Front. Neuroendocr. 2008, 29, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felici, F.; Bazzucchi, I.; Sgrò, P.; Quinzi, F.; Conti, A.; Aversa, A.; Gizzi, L.; Mezzullo, M.; Romanelli, F.; Pasquali, R.; et al. Acute severe male hypo-testosteronemia affects central motor command in humans. J. Electromyogr. Kinesiol. 2015, 28, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Crewther, B.T.; Cook, C.; Cardinale, M.; Weatherby, R.P.; Lowe, T. Two emerging concepts for elite athletes: The short-term effects of testosterone and cortisol on the neuromuscular system and the dose-response training role of these endogenous hormones. Sports Med. 2011, 41, 103–123. [Google Scholar] [CrossRef]
- Antinozzi, C.; Marampon, F.; Corinaldesi, C.; Vicini, E.; Sgrò, P.; Vannelli, G.B.; Lenzi, A.; Crescioli, C.; Di Luigi, L. Testosterone insulin-like effects: An in vitro study on the short-term metabolic effects of testosterone in human skeletal muscle cells. J. Endocrinol. Investig. 2017, 40, 1133–1143. [Google Scholar] [CrossRef]
- Antinozzi, C.; Marampon, F.; Sgrò, P.; Tombolini, V.; Lenzi, A.; Crescioli, C.; Di Luigi, L. Comparative study of testosterone and vitamin D analogue, elocalcitol, on insulin-controlled signal transduction pathway regulation in human skeletal muscle cells. J. Endocrinol. Investig. 2019, 42, 897–907. [Google Scholar] [CrossRef]
- Spasojevic, I.; Jones, D.R.; Andrades, M.E. Hydrogen Peroxide in Adaptation. Oxidative Med. Cell. Longev. 2012, 2012, 596019. [Google Scholar] [CrossRef]
- Ceci, R.; Valls, M.R.B.; Duranti, G.; Dimauro, I.; Quaranta, F.; Pittaluga, M.; Sabatini, S.; Caserotti, P.; Parisi, P.; Parisi, A.; et al. Oxidative stress responses to a graded maximal exercise test in older adults following explosive-type resistance training. Redox Biol. 2013, 2, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiMauro, I.; Sgura, A.; Pittaluga, M.; Magi, F.; Fantini, C.; Mancinelli, R.; Sgadari, A.; Fulle, S.; Caporossi, D. Regular exercise participation improves genomic stability in diabetic patients: An exploratory study to analyse telomere length and DNA damage. Sci. Rep. 2017, 7, 4137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Dimauro, I.; Paronetto, M.P.; Caporossi, D. Exercise, redox homeostasis and the epigenetic landscape. Redox Biol. 2020, 35, 101477. [Google Scholar] [CrossRef]
- Jackson, M.J.; Stretton, C.; McArdle, A. Hydrogen peroxide as a signal for skeletal muscle adaptations to exercise: What do concentrations tell us about potential mechanisms? Redox Biol. 2020, 35, 101484. [Google Scholar] [CrossRef]
- Dimauro, I.; Antonioni, A.; Mercatelli, N.; Caporossi, D. The Role of αB-Crystallin in Skeletal and Cardiac Muscle Tissues. Cell Stress Chaperones 2018, 23, 491–505. [Google Scholar] [CrossRef]
- Azad, N.; Iyer, A.K.V. Reactive Oxygen Species and Apoptosis. In Systems Biology of Free Radicals and Antioxidants; Laher, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Drobnis, E.Z.; Nangia, A.K. Phosphodiesterase Inhibitors (PDE Inhibitors) and Male Reproduction. Adv. Exp. Med. Biol. 2017, 1034, 29–38. [Google Scholar] [CrossRef]
- Antinozzi, C.; Sgrò, P.; Di Luigi, L. Advantages of Phosphodiesterase Type 5 Inhibitors in the Management of Glucose Metabolism Disorders: A Clinical and Translational Issue. Int. J. Endocrinol. 2020, 2020, 7078108. [Google Scholar] [CrossRef]
- Di Luigi, L.; Baldari, C.; Sgrò, P.G.M.; Emerenziani, G.P.; Gallotta, M.C.; Bianchini, S.; Romanelli, F.; Pigozzi, F.; Lenzi, A.; Guidetti, L. The Type 5 Phosphodiesterase Inhibitor Tadalafil Influences Salivary Cortisol, Testosterone, and Dehydroepiandrosterone Sulphate Responses to Maximal Exercise in Healthy Men. J. Clin. Endocrinol. Metab. 2008, 93, 3510–3514. [Google Scholar] [CrossRef]
- Sgrò, P.; Minganti, C.; Lista, M.; Antinozzi, C.; Cappa, M.; Pitsiladis, Y.; Pigozzi, F.; Di Luigi, L. Dihydrotestosterone (DHT) rapidly increase after maximal aerobic exercise in healthy males: The lowering effect of phosphodiesterase’s type 5 inhibitors on DHT response to exercise-related stress. J. Endocrinol. Investig. 2020, 44, 1219–1228. [Google Scholar] [CrossRef]
- Feng, Y.; Wu, J.; Cheng, Z.; Zhang, J.; Lu, J.; Shi, R. Mechanical stretch enhances sex steroidogenesis in C2C12 skeletal muscle cells. Steroids 2019, 150, 108434. [Google Scholar] [CrossRef] [PubMed]
- Loo, D.T.; Rillema, J.R. Measurement of Cell Death. Methods Cell Biol. 1998, 57, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Fittipaldi, S.; Mercatelli, N.; Dimauro, I.; Jackson, M.J.; Paronetto, M.P.; Caporossi, D. Alpha B-crystallin induction in skeletal muscle cells under redox imbalance is mediated by a JNK-dependent regulatory mechanism. Free Radic. Biol. Med. 2015, 86, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Dimauro, I.; Antonioni, A.; Mercatelli, N.; Grazioli, E.; Fantini, C.; Barone, R.; Macaluso, F.; Di Felice, V.; Caporossi, D. The early response of αB-crystallin to a single bout of aerobic exercise in mouse skeletal muscles depends upon fiber oxidative features. Redox Biol. 2019, 24, 101183. [Google Scholar] [CrossRef]
- Antonioni, A.; Dimauro, I.; Fantini, C.; Barone, R.; Macaluso, F.; Di Felice, V.; Caporossi, D. αB-crystallin response to a pro-oxidant non-cytotoxic environment in murine cardiac cells: An “in vitro” and “in vivo” study. Free Radic. Biol. Med. 2020, 152, 301–312. [Google Scholar] [CrossRef]
- Antinozzi, C.; Sgrò, P.; Marampon, F.; Caporossi, D.; Del Galdo, F.; Dimauro, I.; Di Luigi, L. Sildenafil Counteracts the In Vitro Activation of CXCL-9, CXCL-10 and CXCL-11/CXCR3 Axis Induced by Reactive Oxygen Species in Scleroderma Fibroblasts. Biology 2021, 10, 491. [Google Scholar] [CrossRef]
- Arrigo, A.-P.; Gibert, B. Protein interactomes of three stress inducible small heat shock proteins: HspB1, HspB5 and HspB8. Int. J. Hyperth. 2013, 29, 409–422. [Google Scholar] [CrossRef]
- Hamdi, M.M.; Mutungi, G. Dihydrotestosterone activates the MAPK pathway and modulates maximum isometric force through the EGF receptor in isolated intact mouse skeletal muscle fibres. J. Physiol. 2010, 588, 511–525. [Google Scholar] [CrossRef]
- Hamdi, M.M.; Mutungi, G. Dihydrotestosterone stimulates amino acid uptake and the expression of LAT2 in mouse skeletal muscle fibres through an ERK1/2-dependent mechanism. J. Physiol. 2011, 589, 3623–3640. [Google Scholar] [CrossRef] [Green Version]
- Na Lee, M.; Lee, S.H.; Lee, M.Y.; Kim, Y.H.; Park, J.H.; Ryu, J.M.; Yun, S.P.; Lee, Y.J.; Kim, M.O.; Park, K.; et al. Effect of dihydrotestosterone on mouse embryonic stem cells exposed to H2O2-induced oxidative stress. J. Veter. Sci. 2008, 9, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.R.; Marcelino, H.; Gonçalves, I.M.T.M.V.; Quintela, T.; Tomás, J.F.M.; Duarte, A.C.; Fonseca, A.M.; Santos, C.R.A. Sex Hormones Protect Against Amyloid-β Induced Oxidative Stress in the Choroid Plexus Cell Line Z310. J. Neuroendocr. 2016, 28. [Google Scholar] [CrossRef] [PubMed]
- Kajihara, T.; Tochigi, H.; Prechapanich, J.; Uchino, S.; Itakura, A.; Brosens, J.; Ishihara, O. Androgen signaling in decidualizing human endometrial stromal cells enhances resistance to oxidative stress. Fertil. Steril. 2012, 97, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.M.; Jung, H.S.; Kwon, M.J.; Lee, S.H.; Park, J.H. Testosterone Protects Pancreatic β-cells from Apoptosis and Stress-Induced Accelerated Senescence. World J. Men’s Heal. 2021, 39, 724. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, A.M.; Alabiad, M.A.; El Shaer, D.F. Resveratrol Ameliorates the Seminiferous Tubules Damages Induced by Finasteride in Adult Male Rats. Microsc. Microanal. 2020, 26, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Peltola, V.; Huhtaniemi, I.; Metsa-Ketela, T.; Ahotupa, M. Induction of lipid peroxidation during steroidogenesis in the rat testis. Endocrinology 1996, 137, 105–112. [Google Scholar] [CrossRef]
- Chainy, G.B.N.; Samantaray, S.; Samanta, L. Testosterone-induced changes in testicular antioxidant system. Andrologia 2009, 29, 343–349. [Google Scholar] [CrossRef]
- Zhu, X.-D.; Bonet, B.; Knopp, R.H. 17β-Estradiol, progesterone, and testosterone inversely modulate low-density lipoprotein oxidation and cytotoxicity in cultured placental trophoblast and macrophages. Am. J. Obstet. Gynecol. 1997, 177, 196–209. [Google Scholar] [CrossRef]
- Pansarasa, O.; D’Antona, G.; Gualea, M.; Marzani, B.; Pellegrino, M.; Marzatico, F. “Oxidative stress”: Effects of mild endurance training and testosterone treatment on rat gastrocnemius muscle. Eur. J. Appl. Physiol. 2002, 87, 550–555. [Google Scholar] [CrossRef]
- Aydilek, N.; Aksakal, M.; Karakilcik, A.Z.; Karakılçık, A.Z. Effects of testosterone and vitamin E on the antioxidant system in rabbit testis. Andrologia 2004, 36, 277–281. [Google Scholar] [CrossRef]
- Kraemer, W.J.; Ratamess, N.A.; Hymer, W.C.; Nindl, B.C.; Fragala, M.S. Growth Hormone(s), Testosterone, Insulin-Like Growth Factors, and Cortisol: Roles and Integration for Cellular Development and Growth with Exercise. Front. Endocrinol. 2020, 11, 33. [Google Scholar] [CrossRef]
- Aizawa, K.; Iemitsu, M.; Maeda, S.; Jesmin, S.; Otsuki, T.; Mowa, C.N.; Miyauchi, T.; Mesaki, N. Expression of steroidogenic enzymes and synthesis of sex steroid hormones from DHEA in skeletal muscle of rats. Am. J. Physiol. Metab. 2007, 292, E577–E584. [Google Scholar] [CrossRef]
- Fujiki, J.; Maeda, N.; Sato, M.; Ieko, T.; Inoue, H.; Iwasaki, T.; Iwano, H.; Yokota, H. Corticosterone biosynthesis in mouse clonal myoblastic C2C12 cells. Steroids 2018, 138, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, K.; Iemitsu, M.; Maeda, S.; Mesaki, N.; Ushida, T.; Akimoto, T. Endurance Exercise Training Enhances Local Sex Steroidogenesis in Skeletal Muscle. Med. Sci. Sports Exerc. 2011, 43, 2072–2080. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, K.; Iemitsu, M.; Maeda, S.; Otsuki, T.; Sato, K.; Ushida, T.; Mesaki, N.; Akimoto, T. Acute exercise activates local bioactive androgen metabolism in skeletal muscle. Steroids 2010, 75, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.A.; Toone, R.; Peacock, O.; Drawer, S.; Stokes, K.; Cook, C. Dihydrotestosterone is elevated following sprint exercise in healthy young men. J. Appl. Physiol. 2013, 114, 1435–1440. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Iemitsu, M. The Role of Dehydroepiandrosterone (DHEA) in Skeletal Muscle. Vitam. Horm. 2018, 108, 205–221. [Google Scholar] [CrossRef]
- Guidetti, L.; Emerenziani, G.P.; Gallotta, M.C.; Pigozzi, F.; Di Luigi, L.; Baldari, C. Effect of tadalafil on anaerobic performance indices in healthy athletes. Br. J. Sports Med. 2007, 42, 130–133. [Google Scholar] [CrossRef]
- Di Luigi, L.; Baldari, C.; Pigozzi, F.; Emerenziani, G.P.; Gallotta, M.C.; Iellamo, F.; Ciminelli, E.; Sgrò, P.; Romanelli, F.; Lenzi, A.; et al. The Long-Acting Phosphodiesterase Inhibitor Tadalafil does not Influence Athletes’ VO2max, Aerobic, and Anaerobic Thresholds in Normoxia. Int. J. Sports Med. 2008, 29, 110–115. [Google Scholar] [CrossRef]
- Di Luigi, L.; Sgrò, P.G.M.; Baldari, C.; Gallotta, M.C.; Emerenziani, G.P.; Crescioli, C.; Bianchini, S.; Romanelli, F.; Lenzi, A.; Guidetti, L. The phosphodiesterases type 5 inhibitor tadalafil reduces the activation of the hypothalamus-pituitary-adrenal axis in men during cycle ergometric exercise. Am. J. Physiol. Metab. 2012, 302, E972–E978. [Google Scholar] [CrossRef] [Green Version]
- Di Luigi, L.; Botrè, F.; Sabatini, S.; Sansone, M.; Mazzarino, M.; Guidetti, L.; Baldari, C.; Lenzi, A.; Caporossi, D.; Romanelli, F.; et al. Acute effects of physical exercise and phosphodiesterase’s type 5 inhibition on serum 11β-hydroxysteroid dehydrogenases related glucocorticoids metabolites: A pilot study. Endocrine 2014, 47, 952–958. [Google Scholar] [CrossRef]
- Ceci, R.; Duranti, G.; Rossi, A.; Savini, I.; Sabatini, S. Skeletal Muscle Differentiation: Role of Dehydroepiandrosterone Sulfate. Horm. Metab. Res. 2011, 43, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, S.; Sgrò, P.G.M.; Duranti, G.; Ceci, R.; Di Luigi, L. Tadalafil alters energy metabolism in C2C12 skeletal muscle cells. Acta Biochim. Pol. 2011, 58, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isenmann, E.; Ambrosio, G.; Joseph, J.F.; Mazzarino, M.; de la Torre, X.; Zimmer, P.; Kazlauskas, R.; Goebel, C.; Botrè, F.; Diel, P.; et al. Ecdysteroids as non-conventional anabolic agent: Performance enhancement by ecdysterone supplementation in humans. Arch. Toxicol. 2019, 93, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Riedel, W. Role of nitric oxide in the control of the hypothalamic-pituitary-adrenocortical axis. Z. Für Rheumatol. 2000, 59 (Suppl. S2), II36–II42. [Google Scholar] [CrossRef]
- Bugajski, J.; Gądek-Michalska, A.; Bugajski, A.J. Nitric oxide and prostaglandin systems in the stimulation of hypothalamic-pituitary-adrenal axis by neurotransmitters and neurohormones. J. Physiol. Pharmacol. 2004, 55, 679–703. [Google Scholar]
- Jobgen, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.; Wu, G. Regulatory role for the arginine–nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem. 2006, 17, 571–588. [Google Scholar] [CrossRef]
- Gądek-Michalska, A.; Bugajski, J. Nitric oxide in the adrenergic-and CRH-induced activation of hypothalamic-pituitary-adrenal axis. J. Physiol. Pharmacol. 2008, 59, 365–378. [Google Scholar]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Vinciguerra, I.; De Lutiis, M.; Grilli, A.; Felaco, M.; Patruno, A. Phosphodiesterase Type-5 Inhibitor and Oxidative Stress. Int. J. Immunopathol. Pharmacol. 2008, 21, 879–889. [Google Scholar] [CrossRef]
- Ceci, R.; Duranti, G.; Sgrò, P.; Sansone, M.; Guidetti, L.; Baldari, C.; Sabatini, S.; Di Luigi, L. Effects of tadalafil administration on plasma markers of exercise-induced muscle damage, IL6 and antioxidant status capacity. Eur. J. Appl. Physiol. 2015, 115, 531–539. [Google Scholar] [CrossRef]
- Ceci, R.; Duranti, G.; Sgrò, P.; Sabatini, S.; Di Luigi, L. Acute tadalafil administration increases plasma fatty acids without changes in the inflammatory response in healthy men. Acta Biochim. Pol. 2017, 64, 687–691. [Google Scholar] [CrossRef]
- Duranti, G.; Ceci, R.; Sgrò, P.; Sabatini, S.; Di Luigi, L. Influence of the PDE5 inhibitor tadalafil on redox status and antioxidant defense system in C2C12 skeletal muscle cells. Cell Stress Chaperones 2017, 22, 389–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, P.; Singh, N. Tadalafil ameliorates memory deficits, oxidative stress, endothelial dysfunction and neuropathological changes in rat model of hyperhomocysteinemia induced vascular dementia. Int. J. Neurosci. 2020, 132, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Di Luigi, L.; Duranti, G.; Antonioni, A.; Sgrò, P.; Ceci, R.; Crescioli, C.; Sabatini, S.; Lenzi, A.; Caporossi, D.; Del Galdo, F.; et al. The Phosphodiesterase Type 5 Inhibitor Sildenafil Improves DNA Stability and Redox Homeostasis in Systemic Sclerosis Fibroblasts Exposed to Reactive Oxygen Species. Antioxidants 2020, 9, 786. [Google Scholar] [CrossRef] [PubMed]
- Di Luigi, L.; Sgrò, P.; Duranti, G.; Sabatini, S.; Caporossi, D.; Del Galdo, F.; DiMauro, I.; Antinozzi, C. Sildenafil Reduces Expression and Release of IL-6 and IL-8 Induced by Reactive Oxygen Species in Systemic Sclerosis Fibroblasts. Int. J. Mol. Sci. 2020, 21, 3161. [Google Scholar] [CrossRef] [PubMed]
- Duranti, G.; Ceci, R.; Di Luigi, L.; Antinozzi, C.; Dimauro, I.; Sabatini, S.; Cervelli, M.; Sgrò, P. Effect of Tadalafil Administration on Redox Homeostasis and Polyamine Levels in Healthy Men with High Level of Physical Activity. Int. J. Environ. Res. Public Health 2021, 18, 9962. [Google Scholar] [CrossRef]
- Khantham, C.; Yooin, W.; Sringarm, K.; Sommano, S.; Jiranusornkul, S.; Carmona, F.; Nimlamool, W.; Jantrawut, P.; Rachtanapun, P.; Ruksiriwanich, W. Effects on Steroid 5-Alpha Reductase Gene Expression of Thai Rice Bran Extracts and Molecular Dynamics Study on SRD5A2. Biology 2021, 10, 319. [Google Scholar] [CrossRef]
- Persky, A.M.; Green, P.S.; Stubley, L.; Howell, C.O.; Zaulyanov, L.; Brazeau, G.A.; Simpkins, J.W. Protective Effect of Estrogens Against Oxidative Damage to Heart and Skeletal Muscle In Vivo and In Vitro. Proc. Soc. Exp. Boil. Med. 2000, 223, 59–66. [Google Scholar] [CrossRef]
- Baltgalvis, K.A.; Greising, S.M.; Warren, G.; Lowe, D.A. Estrogen Regulates Estrogen Receptors and Antioxidant Gene Expression in Mouse Skeletal Muscle. PLoS ONE 2010, 5, e10164. [Google Scholar] [CrossRef] [Green Version]
- MacNeil, L.G.; Baker, S.K.; Stevic, I.; Tarnopolsky, M.A. 17β-estradiol attenuates exercise-induced neutrophil infiltration in men. Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R1443–R1451. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.-L.; Lin, T.-M.; Wang, S.-W.; Wang, P.S. Antisteroidogenic effects of hydrogen peroxide on rat granulosa cells. Free Radic. Res. 2012, 46, 718–725. [Google Scholar] [CrossRef]
- Angeloni, C.; Teti, G.; Barbalace, M.C.; Malaguti, M.; Falconi, M.; Hrelia, S. 17β-Estradiol enhances sulforaphane cardioprotection against oxidative stress. J. Nutr. Biochem. 2017, 42, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Burattini, S.; Ferri, P.; Battistelli, M.; Curci, R.; Luchetti, F.; Falcieri, E. C2C12 murine myoblasts as a model of skeletal muscle development: Morpho-functional characterization. Eur. J. Histochem. 2004, 48, 223–233. [Google Scholar] [PubMed]
- Duranti, G.; LA Rosa, P.; Dimauro, I.; Wannenes, F.; Bonini, S.; Sabatini, S.; Parisi, P.; Caporossi, D. Effects of Salmeterol on Skeletal Muscle Cells. Med. Sci. Sports Exerc. 2011, 43, 2259–2273. [Google Scholar] [CrossRef] [PubMed]
- Duranti, G.; Maldini, M.; Crognale, D.; Horner, K.; Dimauro, I.; Sabatini, S.; Ceci, R. Moringa oleifera Leaf Extract Upregulates Nrf2/HO−1 Expression and Ameliorates Redox Status in C2C12 Skeletal Muscle Cells. Molecules 2021, 26, 5041. [Google Scholar] [CrossRef]
- Dimauro, I.; Mercatelli, N.; Caporossi, D. Exercise-induced ROS in heat shock proteins response. Free Radic. Biol. Med. 2016, 98, 46–55. [Google Scholar] [CrossRef]
- Magi, F.; DiMauro, I.; Margheritini, F.; Duranti, G.; Mercatelli, N.; Fantini, C.; Ripani, F.R.; Sabatini, S.; Caporossi, D. Telomere length is independently associated with age, oxidative biomarkers, and sport training in skeletal muscle of healthy adult males. Free Radic. Res. 2018, 52, 639–647. [Google Scholar] [CrossRef]
- Duranti, G.; Maldini, M.; Crognale, D.; Sabatini, S.; Corana, F.; Horner, K.; Ceci, R. Moringa oleifera leaf extract influences oxidative metabolism in C2C12 myotubes through SIRT1-PPARα pathway. Phyto. Plus 2020, 1, 100014. [Google Scholar] [CrossRef]
- Marampon, F.; Antinozzi, C.; Corinaldesi, C.; Vannelli, G.B.; Sarchielli, E.; Migliaccio, S.; Di Luigi, L.; Lenzi, A.; Crescioli, C. The phosphodiesterase 5 inhibitor tadalafil regulates lipidic homeostasis in human skeletal muscle cell metabolism. Endocrine 2017, 59, 602–613. [Google Scholar] [CrossRef]
- Testa, E.; Nardozi, D.; Antinozzi, C.; Faieta, M.; di Cecca, S.; Caggiano, C.; Fukuda, T.; Bonanno, E.; Zhenkun, L.; Maldonado, A.; et al. H2AFX and MDC1 promote maintenance of genomic integrity in male germ cells. J. Cell Sci. 2018, 131, jcs214411. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DHT (pg/mL) | |
|---|---|
| ctrl | n.d. |
| H2O2 | n.d. |
| H2O2 + TAD | n.d. |
| TAD | n.d. |
| DHEA | 1547.5 ± 395.7 * |
| Gene Name | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| 5α-R2 | TGGAGGGCATGGTGCTAAAG | TCTCTCACTTAGCACGGGGA |
| 17β-HSD | TTTGCGCTCGAAGGTTTGTG | GCAGTCAAGAAGAGCTCCGT |
| 3β-HSD | ACCTTGTGGCTGACCATCTC | TGCTCTTCCTCGTTGCCATT |
| CYP-19 | AACCCCATGCAGTATAATGTCAC | AGGACCTGGTATTGAAGACGAG |
| β-actin | CTGAACCCCAAGGCCAAC | AGCCTGGATAGCAACGTACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antinozzi, C.; Duranti, G.; Ceci, R.; Lista, M.; Sabatini, S.; Caporossi, D.; Di Luigi, L.; Sgrò, P.; Dimauro, I. Hydrogen Peroxide Stimulates Dihydrotestosterone Release in C2C12 Myotubes: A New Perspective for Exercise-Related Muscle Steroidogenesis? Int. J. Mol. Sci. 2022, 23, 6566. https://doi.org/10.3390/ijms23126566
Antinozzi C, Duranti G, Ceci R, Lista M, Sabatini S, Caporossi D, Di Luigi L, Sgrò P, Dimauro I. Hydrogen Peroxide Stimulates Dihydrotestosterone Release in C2C12 Myotubes: A New Perspective for Exercise-Related Muscle Steroidogenesis? International Journal of Molecular Sciences. 2022; 23(12):6566. https://doi.org/10.3390/ijms23126566
Chicago/Turabian StyleAntinozzi, Cristina, Guglielmo Duranti, Roberta Ceci, Marco Lista, Stefania Sabatini, Daniela Caporossi, Luigi Di Luigi, Paolo Sgrò, and Ivan Dimauro. 2022. "Hydrogen Peroxide Stimulates Dihydrotestosterone Release in C2C12 Myotubes: A New Perspective for Exercise-Related Muscle Steroidogenesis?" International Journal of Molecular Sciences 23, no. 12: 6566. https://doi.org/10.3390/ijms23126566