
Ectopic Expression of Executor Gene Xa23 Enhances Resistance to Both Bacterial and Fungal Diseases in Rice

 , ,
, ,
Abstract
:1. Introduction
2. Results
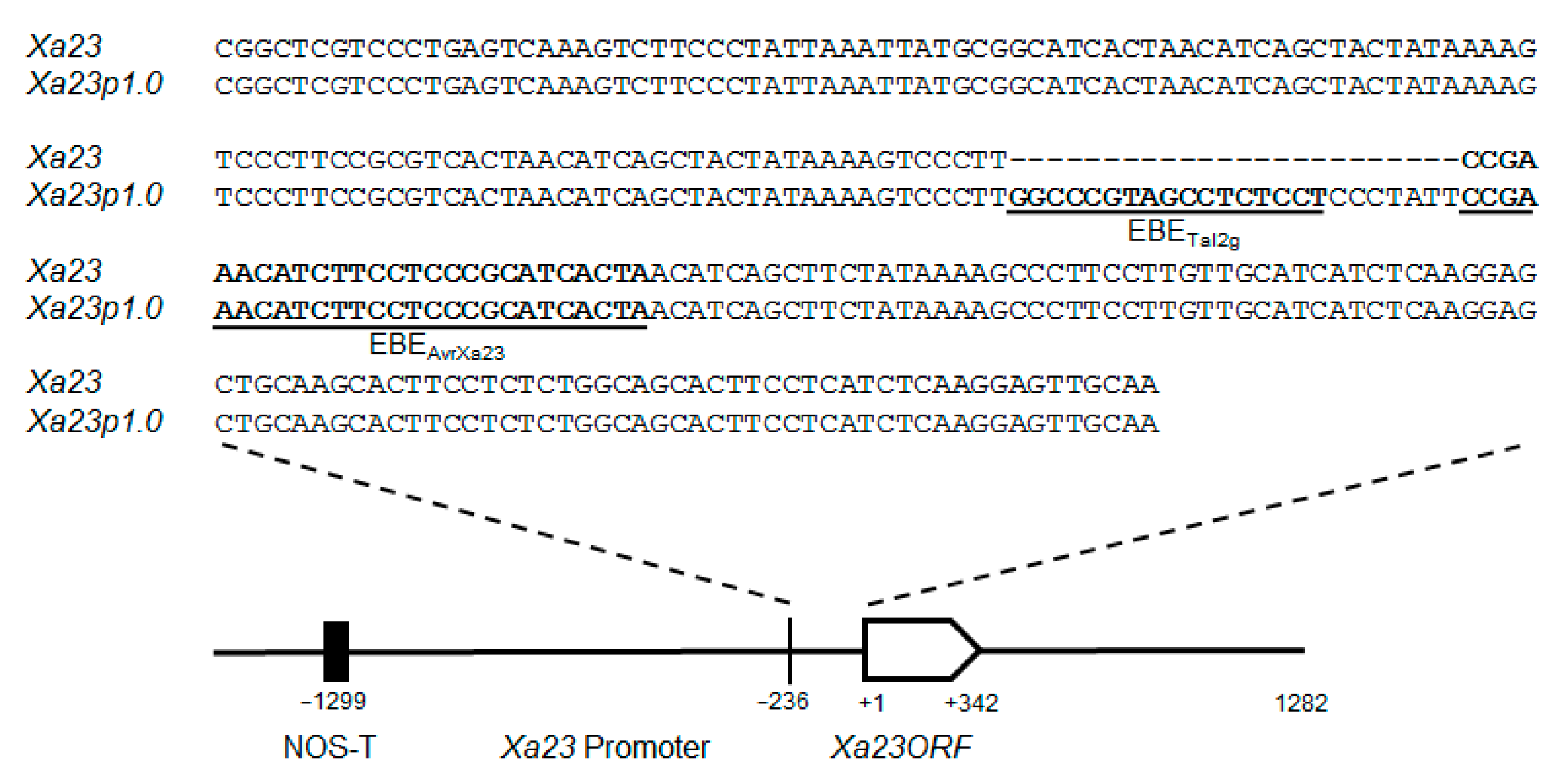
2.1. Generation of EBE-Modified Xa23 Transgenic Rice Plants
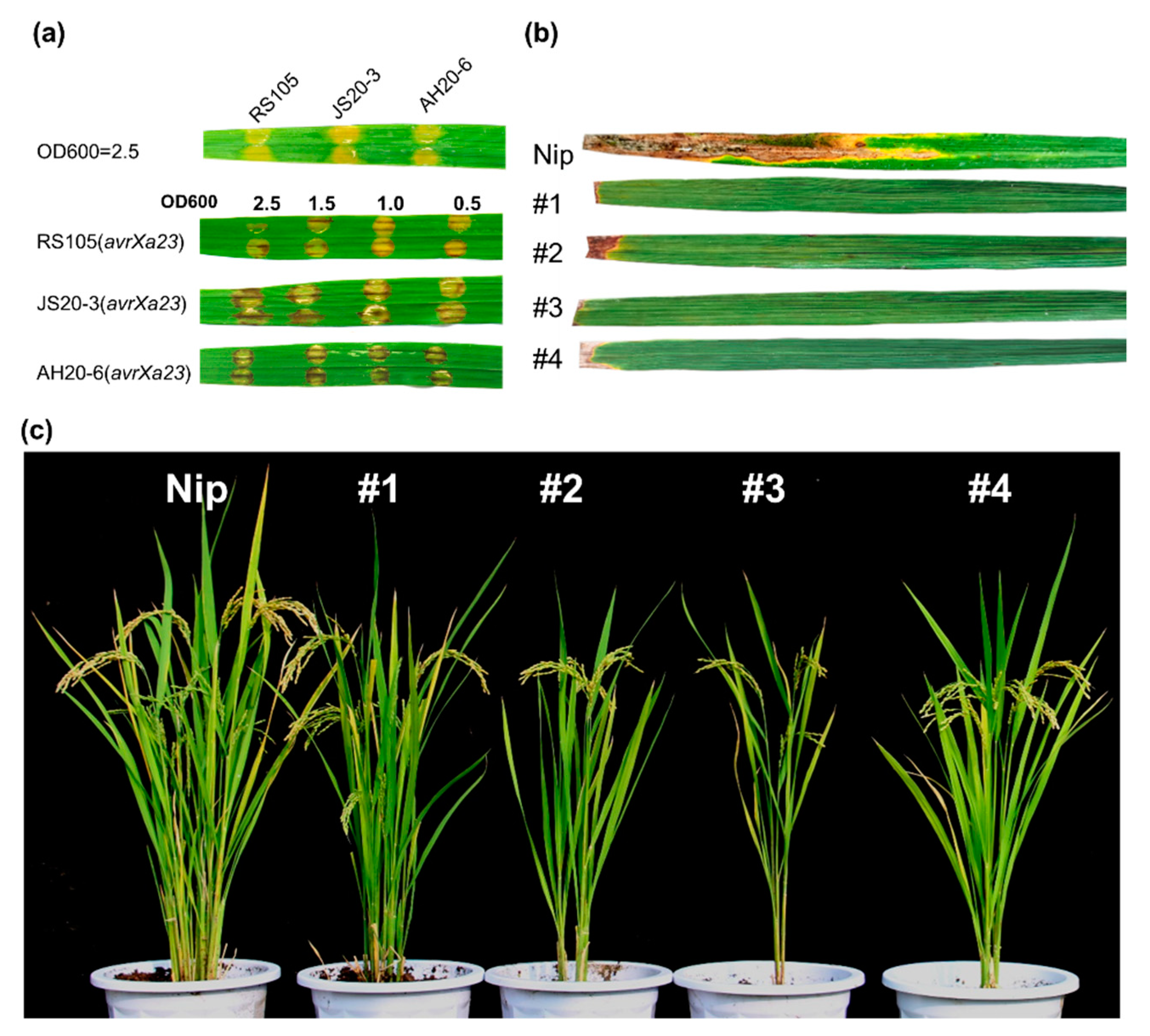
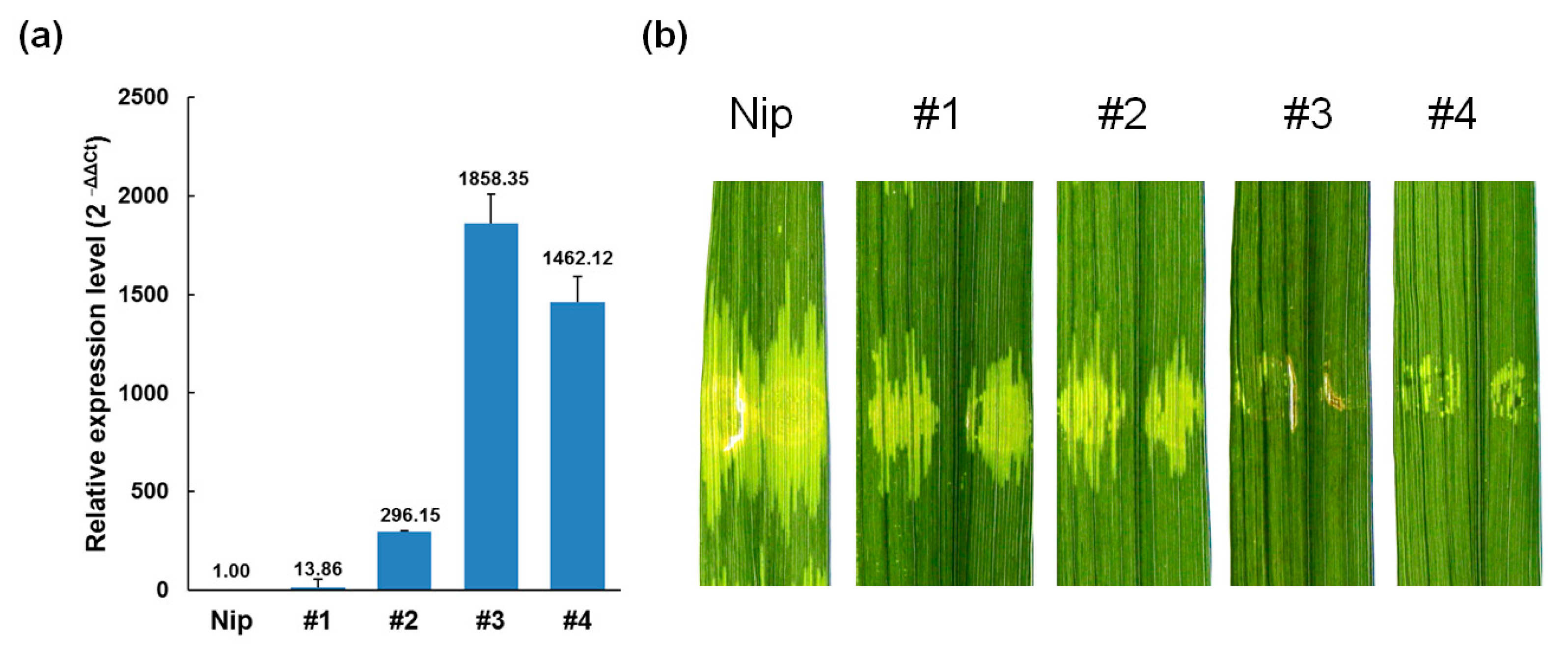
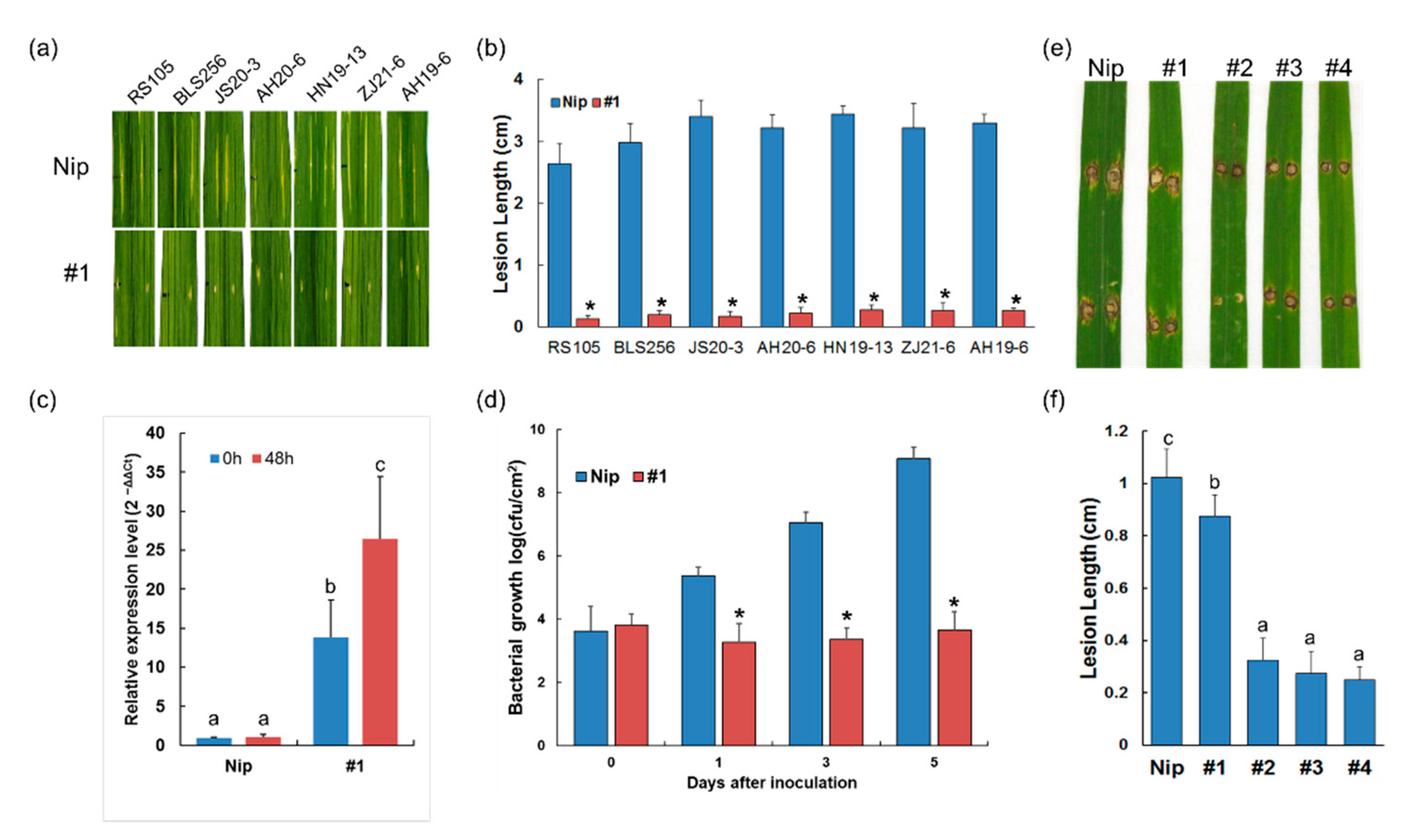
2.2. Xa23p1.0 Transgenic Rice Plants Exhibit Resistance to Xoc and Magnaporthe oryzae
2.3. The Stressed Phenotypes of Xa23p1.0 Transgenic Plants Were Relieved in the Sanya Field Trial
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Bacterial Strains
4.2. Field Experiment
4.3. Plant Inoculations and Disease Assays
4.4. Genes and Constructs
4.5. qRT-PCR Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, Z.; Wang, C.; Zhao, K. Rice Routes of Countering Xanthomonas oryzae. Int. J. Mol. Sci. 2018, 19, 3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nino-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef] [PubMed]
- Nowack, M.K.; Holmes, D.R.; Lahaye, T. TALE-induced cell death executors: An origin outside immunity? Trends Plant Sci. 2021, 27, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant-pathogen interactions. Nat. Rev. Microbiol. 2020, 18, 415–427. [Google Scholar] [CrossRef]
- Boch, J.; Bonas, U.; Lahaye, T. TAL effectors—Pathogen strategies and plant resistance engineering. New Phytol. 2014, 204, 823–832. [Google Scholar] [CrossRef]
- White, F.F.; Yang, B. Host and pathogen factors controlling the rice-Xanthomonas oryzae interaction. Plant Physiol. 2009, 150, 1677–1686. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Guo, W.; Chen, X.; Wang, C.; Zhao, K. Plant Executor Genes. Int. J. Mol. Sci. 2022, 23, 1524. [Google Scholar] [CrossRef]
- Zhang, J.; Yin, Z.; White, F. TAL effectors and the executor R genes. Front. Plant Sci. 2015, 6, 641. [Google Scholar] [CrossRef]
- Chen, X.; Liu, P.; Mei, L.; He, X.; Chen, L.; Liu, H.; Shen, S.; Ji, Z.; Zheng, X.; Zhang, Y.; et al. Xa7, a new executor R gene that confers durable and broad-spectrum resistance to bacterial blight disease in rice. Plant Commun. 2021, 2, 100143. [Google Scholar] [CrossRef]
- Luo, D.; Huguet-Tapia, J.C.; Raborn, R.T.; White, F.F.; Brendel, V.P.; Yang, B. The Xa7 resistance gene guards the rice susceptibility gene SWEET14 against exploitation by the bacterial blight pathogen. Plant Commun. 2021, 2, 100164. [Google Scholar] [CrossRef]
- Tian, D.; Wang, J.; Zeng, X.; Gu, K.; Qiu, C.; Yang, X.; Zhou, Z.; Goh, M.; Luo, Y.; Murata-Hori, M.; et al. The rice TAL effector-dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum. Plant Cell 2014, 26, 497–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.L.; Zhang, X.P.; Fan, Y.L.; Gao, Y.; Zhu, Q.L.; Zheng, C.K.; Qin, T.F.; Li, Y.Q.; Che, J.Y.; Zhang, M.W.; et al. XA23 Is an Executor R Protein and Confers Broad-Spectrum Disease Resistance in Rice. Mol. Plant 2015, 8, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Zhang, Q. A climate-resilient R gene in rice traps two pathogen effectors for broad and durable resistance to bacterial blight. Mol. Plant 2021, 14, 366–368. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.Y.; Zakria, M.; Zou, L.F.; Xiong, L.; Li, Z.; Ji, G.H.; Chen, G.Y. Genetic diversity of transcriptional activator-like effector genes in Chinese isolates of Xanthomonas oryzae pv. oryzicola. Phytopathology 2014, 104, 672–682. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Cao, Y.; Xu, Z.; Ma, W.; Zakria, M.; Zou, L.; Cheng, Z.; Chen, G. A Transcription Activator-Like Effector Tal7 of Xanthomonas oryzae pv. oryzicola Activates Rice Gene Os09g29100 to Suppress Rice Immunity. Sci. Rep. 2017, 7, 5089. [Google Scholar] [CrossRef] [Green Version]
- Hummel, A.W.; Wilkins, K.E.; Wang, L.; Cernadas, R.A.; Bogdanove, A.J. A transcription activator-like effector from Xanthomonas oryzae pv. oryzicola elicits dose-dependent resistance in rice. Mol. Plant Pathol. 2017, 18, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Zhang, H.; Yuan, B.; Liu, H.; Kong, L.; Chu, Z.; Ding, X. Tal2b targets and activates the expression of OsF3H03g to hijack OsUGT74H4 and synergistically interfere with rice immunity. New Phytol. 2021, 223, 1864–1880. [Google Scholar] [CrossRef]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 Years of Resistance Gene Cloning Identifies Nine Mechanisms for R Protein Function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.M.; Zhang, Y. Plant Immunity: Danger Perception and Signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef]
- Ji, C.; Ji, Z.; Liu, B.; Cheng, H.; Liu, H.; Liu, S.; Yang, B.; Chen, G. Xa1 Allelic R Genes Activate Rice Blight Resistance Suppressed by Interfering TAL Effectors. Plant Commun. 2020, 1, 100087. [Google Scholar] [CrossRef]
- Ji, Z.; Ji, C.; Liu, B.; Zou, L.; Chen, G.; Yang, B. Interfering TAL effectors of Xanthomonas oryzae neutralize R-gene-mediated plant disease resistance. Nat. Commun. 2016, 7, 13435. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pruitt, R.N.; Nurnberger, T.; Wang, Y. Evasion of plant immunity by microbial pathogens. Nat. Rev. Microbiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Antony, G.; Li, T.; Liu, B.; Obasa, K.; Yang, B.; White, F.F. The broadly effective recessive resistance gene xa5 of rice is a virulence effector-dependent quantitative trait for bacterial blight. Plant J. 2016, 86, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Ke, Y.G.; Huang, R.Y.; Ma, L.; Yang, Z.Y.; Chu, Z.H.; Xiao, J.H.; Li, X.H.; Wang, S.P. A host basal transcription factor is a key component for infection of rice by TALE-carrying bacteria. eLife 2016, 5, e19605. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.; Shi, Y.; Tian, J.; Wang, L.; Li, Y.; Wang, S.; Yuan, M. TALE-carrying bacterial pathogens trap host nuclear import receptors for facilitation of infection of rice. Mol. Plant Pathol. 2019, 20, 519–532. [Google Scholar] [CrossRef]
- Ma, L.; Wang, Q.; Yuan, M.; Zou, T.; Yin, P.; Wang, S. Xanthomonas TAL effectors hijack host basal transcription factor IIA alpha and gamma subunits for invasion. Biochem. Biophys. Res. Commun. 2018, 496, 608–613. [Google Scholar] [CrossRef]
- Ma, W.; Zou, L.; Ji, Z.; Xu, X.; Xu, Z.; Yang, Y.; Alfano, J.R.; Chen, G. Xanthomonas oryzae pv. oryzae TALE proteins recruit OsTFIIAgamma1 to compensate for the absence of OsTFIIAgamma5 in bacterial blight in rice. Mol. Plant Pathol. 2018, 19, 2248–2262. [Google Scholar] [CrossRef] [Green Version]
- Boch, J.; Scholze, H.; Schornack, S.; Landgraf, A.; Hahn, S.; Kay, S.; Lahaye, T.; Nickstadt, A.; Bonas, U. Breaking the code of DNA binding specificity of TAL-type III effectors. Science 2009, 326, 1509–1512. [Google Scholar] [CrossRef]
- Moscou, M.J.; Bogdanove, A.J. A simple cipher governs DNA recognition by TAL effectors. Science 2009, 326, 1501. [Google Scholar] [CrossRef]
- Hummel, A.W.; Doyle, E.L.; Bogdanove, A.J. Addition of transcription activator-like effector binding sites to a pathogen strain-specific rice bacterial blight resistance gene makes it effective against additional strains and against bacterial leaf streak. New Phytol. 2012, 195, 883–893. [Google Scholar] [CrossRef]
- Shantharaj, D.; Romer, P.; Figueiredo, J.F.L.; Minsavage, G.V.; Kronauer, C.; Stall, R.E.; Moore, G.A.; Fisher, L.C.; Hu, Y.; Horvath, D.M.; et al. An engineered promoter driving expression of a microbial avirulence gene confers recognition of TAL effectors and reduces growth of diverse Xanthomonas strains in citrus. Mol. Plant Pathol. 2017, 18, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Tian, D.; Gu, K.; Zhou, Z.; Yang, X.; Luo, Y.; White, F.F.; Yin, Z. Genetic engineering of the Xa10 promoter for broad-spectrum and durable resistance to Xanthomonas oryzae pv. oryzae. Plant Biotechnol. J. 2015, 13, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Makino, S.; Sugio, A.; White, F.; Bogdanove, A.J. Inhibition of resistance gene-mediated defense in rice by Xanthomonas oryzae pv. oryzicola. Mol. Plant Microbe Interact. 2006, 19, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Mucke, S.; Reschke, M.; Erkes, A.; Schwietzer, C.A.; Becker, S.; Streubel, J.; Morgan, R.D.; Wilson, G.G.; Grau, J.; Boch, J. Transcriptional Reprogramming of Rice Cells by Xanthomonas oryzae TALEs. Front. Plant Sci. 2019, 10, 162. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, K.E.; Booher, N.J.; Wang, L.; Bogdanove, A.J. TAL effectors and activation of predicted host targets distinguish Asian from African strains of the rice pathogen Xanthomonas oryzae pv. oryzicola while strict conservation suggests universal importance of five TAL effectors. Front. Plant Sci. 2015, 6, 536. [Google Scholar] [CrossRef]
- Cernadas, R.A.; Doyle, E.L.; Nino-Liu, D.O.; Wilkins, K.E.; Bancroft, T.; Wang, L.; Schmidt, C.L.; Caldo, R.; Yang, B.; White, F.F.; et al. Code-assisted discovery of TAL effector targets in bacterial leaf streak of rice reveals contrast with bacterial blight and a novel susceptibility gene. PLoS Pathog. 2014, 10, e1003972. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Xu, Z.Y.; Li, Z.Y.; Zakria, M.; Zou, L.F.; Chen, G.Y. Increasing resistance to bacterial leaf streak in rice by editing the promoter of susceptibility gene OsSULRT3; 6. Plant Biotechnol. J. 2021, 19, 1101–1103. [Google Scholar] [CrossRef]
- Liu, Y.G.; Shirano, Y.; Fukaki, H.; Yanai, Y.; Tasaka, M.; Tabata, S.; Shibata, D. Complementation of plant mutants with large genomic DNA fragments by a transformation-competent artificial chromosome vector accelerates positional cloning. Proc. Natl. Acad. Sci. USA 1999, 96, 6535–6540. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.Q.; Xue, J.Y.; Wang, Q.; Wang, B.; Chen, J.Q. Revisiting the Origin of Plant NBS-LRR Genes. Trends Plant Sci. 2019, 24, 9–12. [Google Scholar] [CrossRef]
- Wei, Z.; Abdelrahman, M.; Gao, Y.; Ji, Z.; Mishra, R.; Sun, H.; Sui, Y.; Wu, C.; Wang, C.; Zhao, K. Engineering broad-spectrum resistance to bacterial blight by CRISPR-Cas9-mediated precise homology directed repair in rice. Mol. Plant 2021, 14, 1215–1218. [Google Scholar] [CrossRef]
- Li, T.; Huang, S.; Zhou, J.; Yang, B. Designer TAL effectors induce disease susceptibility and resistance to Xanthomonas oryzae pv. oryzae in rice. Mol. Plant 2013, 6, 781–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant. Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitsili, E.; Phukan, U.J.; Coll, N.S. Cell Death in Plant Immunity. Cold Spring Harb. Perspect. Biol. 2020, 12, a036483. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Zhang, A.; Ren, Y.; Wu, F.; Wang, G.; Xu, Y.; Lei, C.; Zhu, S.; Pan, T.; et al. A cyclic nucleotide-gated channel mediates cytoplasmic calcium elevation and disease resistance in rice. Cell Res. 2019, 29, 820–831. [Google Scholar] [CrossRef]
- Salzberg, S.L.; Sommer, D.D.; Schatz, M.C.; Phillippy, A.M.; Rabinowicz, P.D.; Tsuge, S.; Furutani, A.; Ochiai, H.; Delcher, A.L.; Kelley, D.; et al. Genome sequence and rapid evolution of the rice pathogen Xanthomonas oryzae pv. oryzae PXO99A. BMC Genomics. 2008, 9, 204. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rice Lines | Plant Height (cm) | Number of Tillers | Number of Panicles | Panicle Length (cm) | Seed Setting Rate (%) | Thousand Seed Weight (g) | Seed Weight per Plant (g) | |
|---|---|---|---|---|---|---|---|---|
| BJ | Nip | 67.88 ± 4.91 c | 22.50 ± 1.66 c | 21.00 ± 1.80 b | 17.34 ± 0.95 c | 79.45 ± 2.36 c | 25.33 ± 0.87 a | 26.21 ± 0.95 c |
| #3 | 52.75 ± 2.45 a | 7.50 ± 0.87 a | 6.50 ± 1.22 a | 12.93 ± 1.17 a | 58.36 ± 4.27 a | 26.25 ± 0.48 a | 12.28 ± 2.19 a | |
| SY | Nip | 64.80 ± 1.55 c | 26.30 ± 1.73 d | 25.20 ± 2.32 c | 16.40 ± 0.81 bc | 69.69 ± 3.99 b | 27.37 ± 0.92 a | 25.80 ± 3.13 c |
| #3 | 59.00 ± 2.45 b | 19.50 ± 2.06 b | 18.60 ± 2.41 b | 16.05 ± 0.56 b | 68.87 ± 6.93 b | 26.57 ± 1.01 a | 20.05 ± 1.54 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, Z.; Sun, H.; Wei, Y.; Li, M.; Wang, H.; Xu, J.; Lei, C.; Wang, C.; Zhao, K. Ectopic Expression of Executor Gene Xa23 Enhances Resistance to Both Bacterial and Fungal Diseases in Rice. Int. J. Mol. Sci. 2022, 23, 6545. https://doi.org/10.3390/ijms23126545
Ji Z, Sun H, Wei Y, Li M, Wang H, Xu J, Lei C, Wang C, Zhao K. Ectopic Expression of Executor Gene Xa23 Enhances Resistance to Both Bacterial and Fungal Diseases in Rice. International Journal of Molecular Sciences. 2022; 23(12):6545. https://doi.org/10.3390/ijms23126545
Chicago/Turabian StyleJi, Zhiyuan, Hongda Sun, Yena Wei, Man Li, Hongjie Wang, Jiangmin Xu, Cailin Lei, Chunlian Wang, and Kaijun Zhao. 2022. "Ectopic Expression of Executor Gene Xa23 Enhances Resistance to Both Bacterial and Fungal Diseases in Rice" International Journal of Molecular Sciences 23, no. 12: 6545. https://doi.org/10.3390/ijms23126545