
Mutations of γCOP Gene Disturb Drosophila melanogaster Innate Immune Response to Pseudomonas aeruginosa

 ,
,
Abstract
:1. Introduction
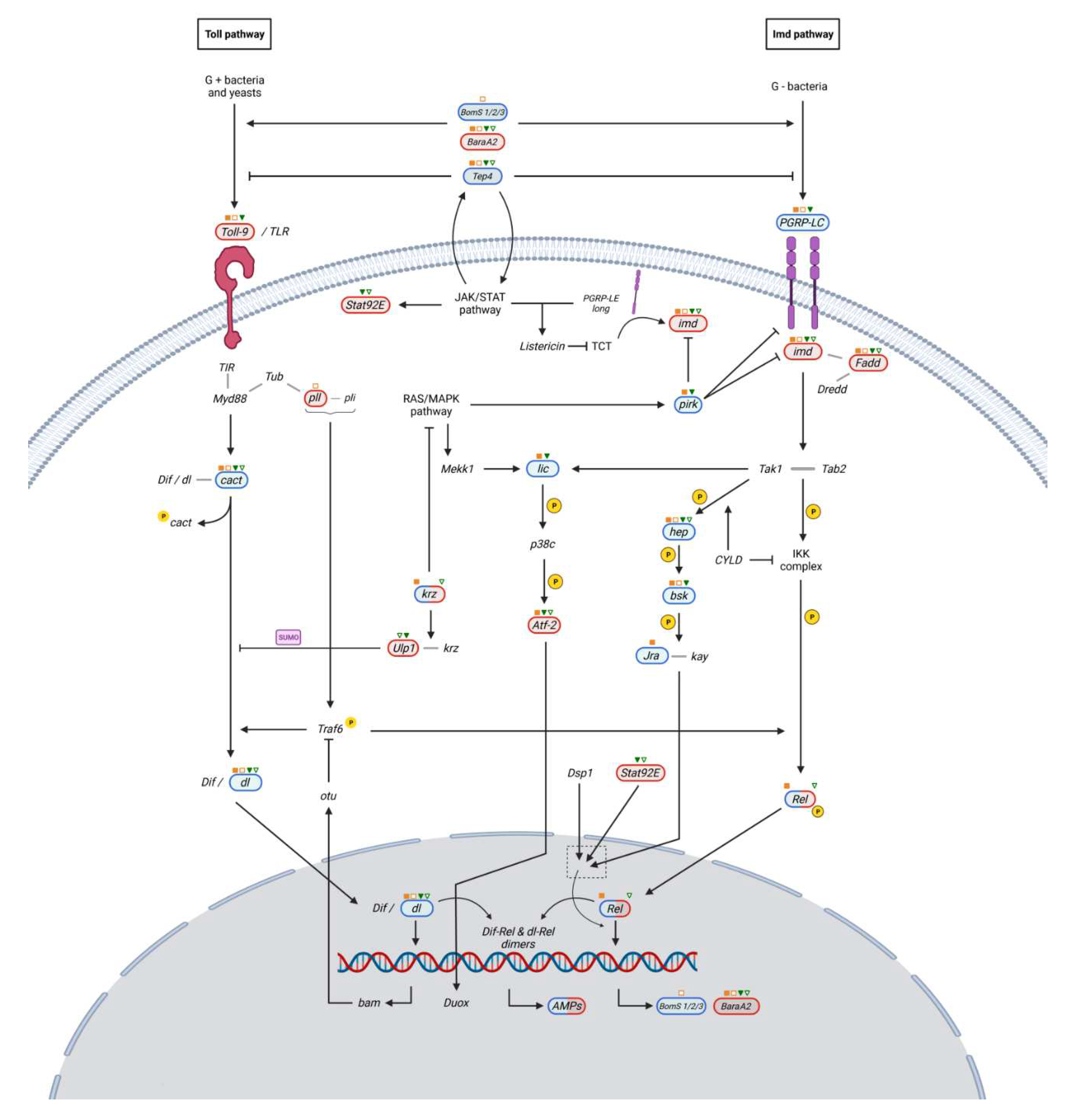
2. Results
3. Discussion
3.1. An Overview of the Mutant γCOP Background in D. melanogaster
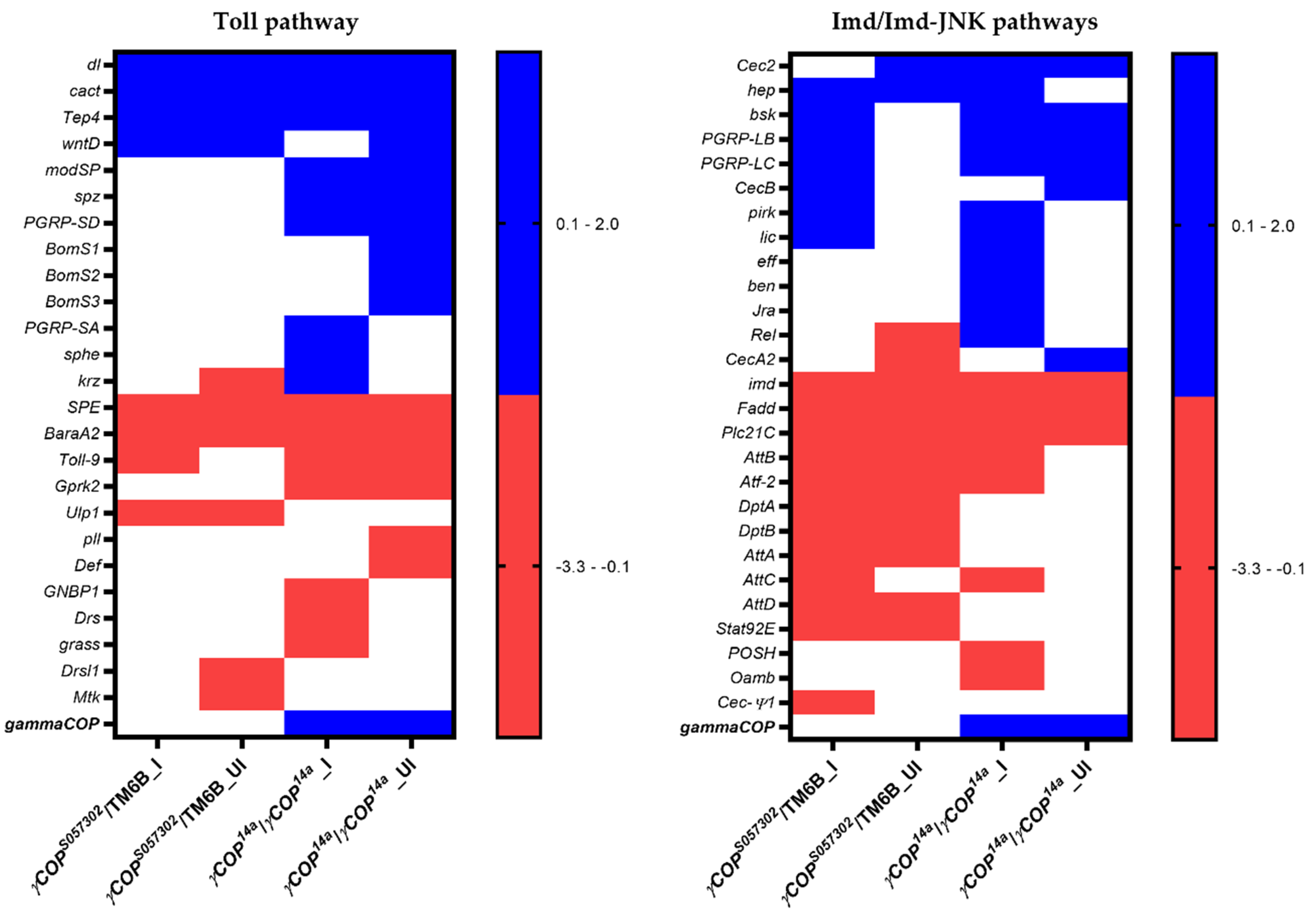
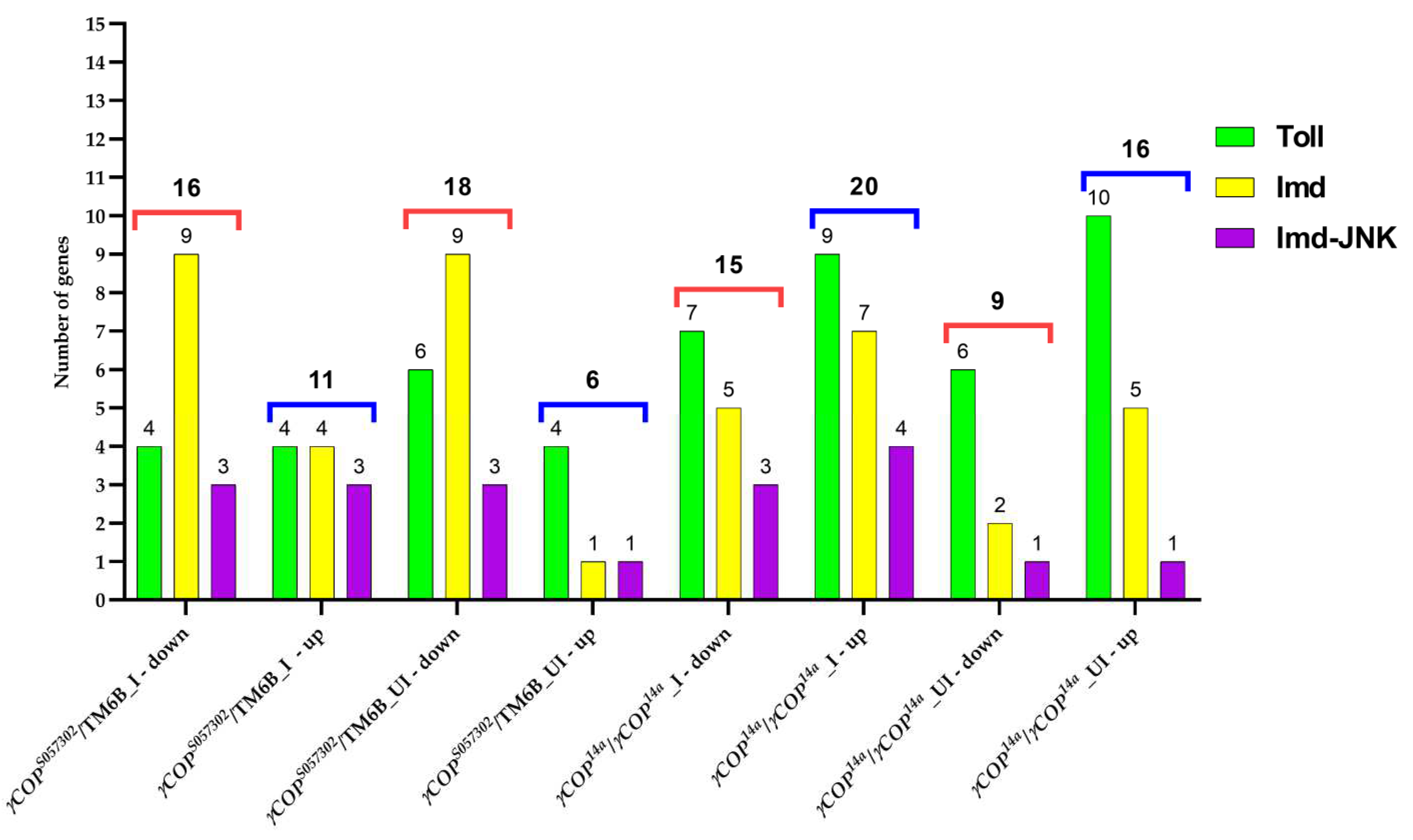
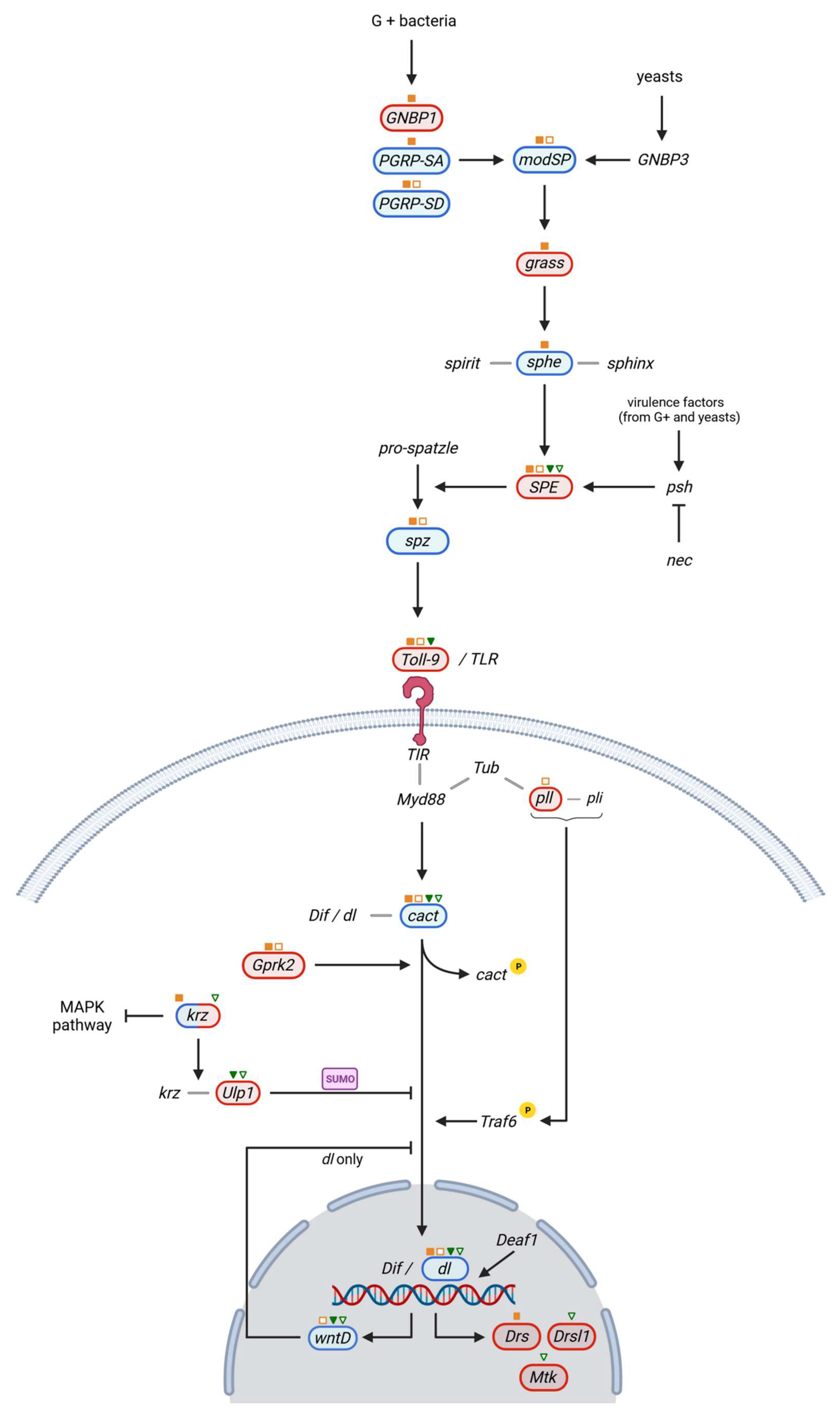
3.2. Toll Pathway
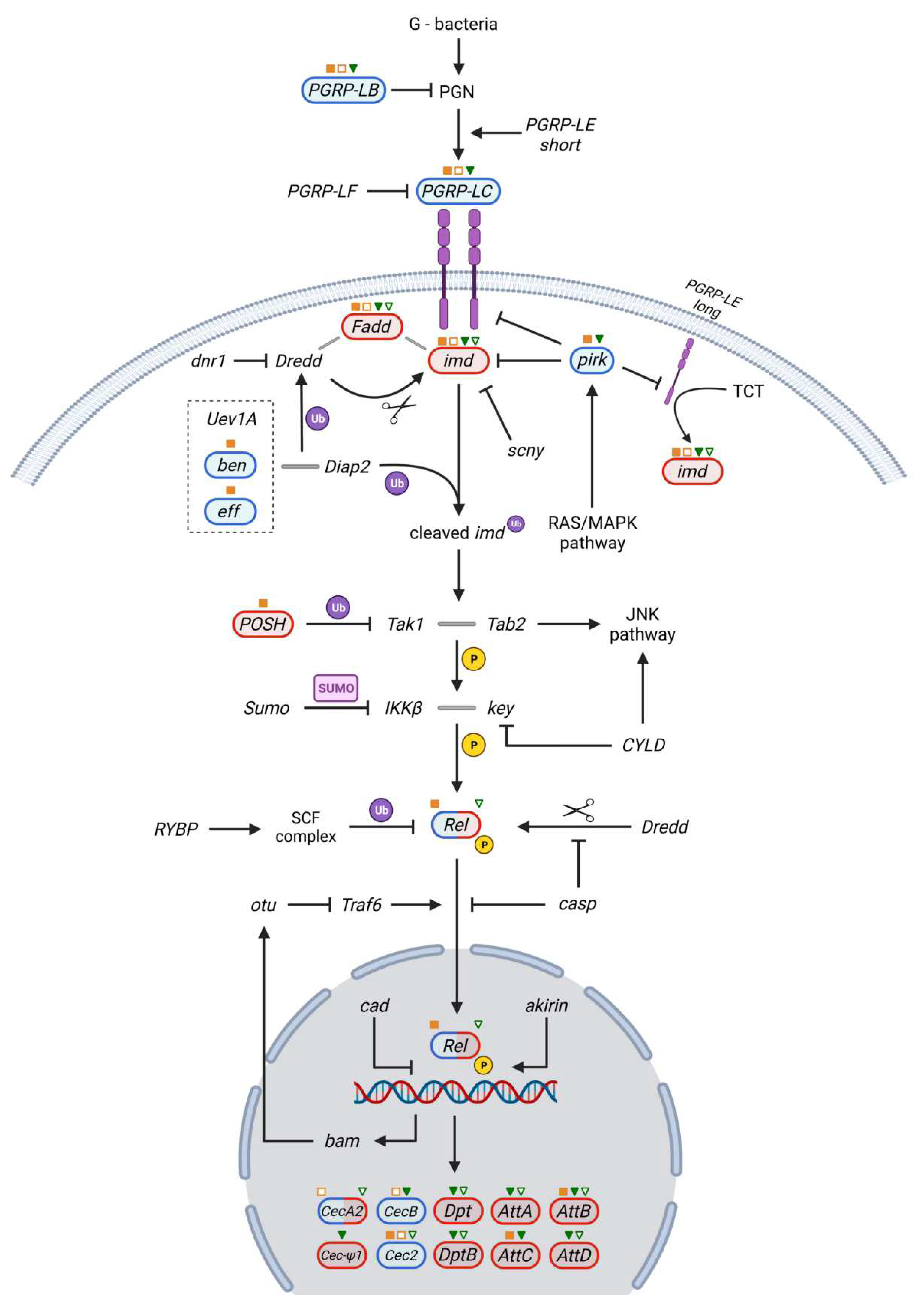
3.3. Imd Pathway
3.4. Toll and Imd Pathways Interplay
4. Materials and Methods
4.1. D. melanogaster Stocks
4.2. Bacterial Strain
4.3. Bacterial Ingestion Assay
4.4. Microarray Project
4.5. Gene Expression Data Set
4.6. Microarray Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavoie, E.G.; Wangdi, T.; Kazmierczak, B.I. Innate immune responses to Pseudomonas aeruginosa infection. Microbes Infect. 2011, 13, 1133–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Vrancianu, C.O.; Popa, L.I.; Bleotu, C.; Chifiriuc, M.C. Targeting Plasmids to Limit Acquisition and Transmission of Antimicrobial Resistance. Front. Microbiol. 2020, 11, 761. [Google Scholar] [CrossRef] [PubMed]
- Pircalabioru, G.G.; Chifiriuc, M.C. Nanoparticulate drug-delivery systems for fighting microbial biofilms: From bench to bedside. Future Microbiol. 2020, 15, 679–698. [Google Scholar] [CrossRef]
- Picard, C.; Casanova, J.L.; Abel, L. Mendelian traits that confer predisposition or resistance to specific infections in humans. Curr. Opin. Immunol. 2006, 18, 383–390. [Google Scholar] [CrossRef]
- Howard, M.W.; Strauss, R.G.; Johnston, R.B., Jr. Infections in patients with neutropenia. Am. J. Dis. Child. 1977, 131, 788–790. [Google Scholar] [CrossRef]
- Bodey, G.P. The changing face of febrile neutropenia-from monotherapy to moulds to mucositis. Fever and neutropenia: The early years. J. Antimicrob. Chemother. 2009, 63 (Suppl. 1), i3–i13. [Google Scholar] [CrossRef] [Green Version]
- Tanji, T.; Hu, X.; Weber, A.N.; Ip, Y.T. Toll and IMD pathways synergistically activate an innate immune response in Drosophila melanogaster. Mol. Cell. Biol. 2007, 27, 4578–4588. [Google Scholar] [CrossRef] [Green Version]
- Ferrandon, D.; Imler, J.L.; Hetru, C.; Hoffmann, J.A. The Drosophila systemic immune response: Sensing and signalling during bacterial and fungal infections. Nat. Rev. Immunol. 2007, 7, 862–874. [Google Scholar] [CrossRef]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas aeruginosa infection: Lessons from a versatile opportunist. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar] [CrossRef]
- Igboin, C.O.; Griffen, A.L.; Leys, E.J. The Drosophila melanogaster host model. J. Oral Microbiol. 2012, 4, 10368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harnish, J.M.; Link, N.; Yamamoto, S. Drosophila as a Model for Infectious Diseases. Int. J. Mol. Sci. 2021, 22, 2724. [Google Scholar] [CrossRef] [PubMed]
- Limmer, S.; Quintin, J.; Hetru, C.; Ferrandon, D. Virulence on the fly: Drosophila melanogaster as a model genetic organism to decipher host-pathogen interactions. Curr. Drug Targets 2011, 12, 978–999. [Google Scholar] [CrossRef] [PubMed]
- Levitt, J.M.; Howell, D.D.; Rodgers, J.R.; Rich, R.R. Exogenous peptides enter the endoplasmic reticulum of TAP-deficient cells and induce the maturation of nascent MHC class I molecules. Eur. J. Immunol. 2001, 31, 1181–1190. [Google Scholar] [CrossRef]
- Roy, C.R.; Salcedo, S.P.; Gorvel, J.P. Pathogen-endoplasmic-reticulum interactions: In through the out door. Nat. Rev. Immunol. 2006, 6, 136–147. [Google Scholar] [CrossRef]
- Unanue, E.R.; Urano, F. Endoplasmic reticulum: An interface between the immune system and metabolism. Diabetes 2014, 63, 48–49. [Google Scholar] [CrossRef] [Green Version]
- Compagnone, M.; Fruci, D. Peptide Trimming for MHC Class I Presentation by Endoplasmic Reticulum Aminopeptidases. Methods Mol. Biol. 2019, 1988, 45–57. [Google Scholar] [CrossRef]
- Li, A.; Song, N.J.; Riesenberg, B.P.; Li, Z. The Emerging Roles of Endoplasmic Reticulum Stress in Balancing Immunity and Tolerance in Health and Diseases: Mechanisms and Opportunities. Front. Immunol. 2019, 10, 3154. [Google Scholar] [CrossRef] [Green Version]
- Bainter, W.; Platt, C.D.; Park, S.Y.; Stafstrom, K.; Wallace, J.G.; Peters, Z.T.; Massaad, M.J.; Becuwe, M.; Salinas, S.A.; Jones, J.; et al. Combined immunodeficiency due to a mutation in the gamma1 subunit of the coat protein I complex. J. Clin. Investig. 2021, 131, 3. [Google Scholar] [CrossRef]
- Xu, F.; Fu, Y.; Sun, T.Y.; Jiang, Z.; Miao, Z.; Shuai, M.; Gou, W.; Ling, C.W.; Yang, J.; Wang, J.; et al. The interplay between host genetics and the gut microbiome reveals common and distinct microbiome features for complex human diseases. Microbiome 2020, 8, 145. [Google Scholar] [CrossRef]
- De Simone, M.; Spagnuolo, L.; Lore, N.I.; Rossi, G.; Cigana, C.; De Fino, I.; Iraqi, F.A.; Bragonzi, A. Host genetic background influences the response to the opportunistic Pseudomonas aeruginosa infection altering cell-mediated immunity and bacterial replication. PLoS ONE 2014, 9, e106873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lore, N.I.; Iraqi, F.A.; Bragonzi, A. Host genetic diversity influences the severity of Pseudomonas aeruginosa pneumonia in the Collaborative Cross mice. BMC Genet. 2015, 16, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, A.Y.; Priebe, G.P.; Ray, C.; Van Rooijen, N.; Pier, G.B. Inescapable need for neutrophils as mediators of cellular innate immunity to acute Pseudomonas aeruginosa pneumonia. Infect. Immun. 2009, 77, 5300–5310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czobor-Barbu, I.; Ratiu, A.C.; Florescu, A.M.; Chifiriuc, M.C.; Ecovoiu, A.A. Comparative analysis of the native microbiota in gammacop mutant versus wild-type genetic background of Drosophila melanogaster. Rom. Biotechnol. Lett. 2018, 23, 14235–14243. [Google Scholar]
- Liu, H.; Bebu, I.; Li, X. Microarray probes and probe sets. Front. Biosci. 2010, 2, 325–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, A.; Marygold, S.J.; Antonazzo, G.; Attrill, H.; Dos Santos, G.; Garapati, P.V.; Goodman, J.L.; Gramates, L.S.; Millburn, G.; Strelets, V.B.; et al. FlyBase: Updates to the Drosophila melanogaster knowledge base. Nucleic Acids Res. 2021, 49, D899–D907. [Google Scholar] [CrossRef] [PubMed]
- Deák, P.; Omar, M.M.; Saunders, R.D.; Pál, M.; Komonyi, O.; Szidonya, J.; Maróy, P.; Zhang, Y.; Ashburner, M.; Benos, P.; et al. P-element insertion alleles of essential genes on the third chromosome of Drosophila melanogaster: Correlation of physical and cytogenetic maps in chromosomal region 86E-87F. Genetics 1997, 147, 1697–1722. [Google Scholar] [CrossRef]
- Ecovoiu, A.A.; Mink, M.; Georgescu, L.M.; Graur, M.; Gavrila, L. Genetic data confirm association of P{lacW}gammaCopS057302 insertion in Drosophila melanogaster gammaCop gene with the mutant phenotype of l/3/S057302 transgenic line. An. Univ. Bucur. 2002, 51, 41–47. [Google Scholar]
- Jayaram, S.A.; Senti, K.A.; Tiklová, K.; Tsarouhas, V.; Hemphälä, J.; Samakovlis, C. COPI vesicle transport is a common requirement for tube expansion in Drosophila. PLoS ONE 2008, 3, e1964. [Google Scholar] [CrossRef] [Green Version]
- Ecovoiu, A.A.; Graur, M.; Ratiu, A.C. Hypomorphic alleles of gammaCop gene from Drosophila melanogaster display an unexpected expression pattern in mutant eggs, testes and embryos. Rom. Biotechnol. Lett. 2019, 24, 324–331. [Google Scholar] [CrossRef]
- Miller, D.E.; Cook, K.R.; Hawley, R.S. The joy of balancers. PLoS Genet. 2019, 15, e1008421. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.E.; Cook, K.R.; Arvanitakis, A.V.; Hawley, R.S. Third Chromosome Balancer Inversions Disrupt Protein-Coding Genes and Influence Distal Recombination Events in Drosophila melanogaster. G3 2016, 6, 1959–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bing, X.; Rzezniczak, T.Z.; Bateman, J.R.; Merritt, T.J. Transvection-based gene regulation in Drosophila is a complex and plastic trait. G3 2014, 4, 2175–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lum, T.E.; Merritt, T.J. Nonclassical regulation of transcription: Interchromosomal interactions at the malic enzyme locus of Drosophila melanogaster. Genetics 2011, 189, 837–849. [Google Scholar] [CrossRef] [Green Version]
- Duncan, I.W. Transvection effects in Drosophila. Annu. Rev. Genet. 2002, 36, 521–556. [Google Scholar] [CrossRef]
- Ferreiro, M.J.; Perez, C.; Marchesano, M.; Ruiz, S.; Caputi, A.; Aguilera, P.; Barrio, R.; Cantera, R. Drosophila melanogaster White Mutant w(1118) Undergo Retinal Degeneration. Front. Neurosci. 2017, 11, 732. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Nishimura, T.; Takano, T.; Naito, S.; Yoo, S.K. white regulates proliferative homeostasis of intestinal stem cells during ageing in Drosophila. Nat. Metab. 2021, 3, 546–557. [Google Scholar] [CrossRef]
- Khadilkar, R.J.; Ray, A.; Chetan, D.R.; Sinha, A.R.; Magadi, S.S.; Kulkarni, V.; Inamdar, M.S. Differential modulation of the cellular and humoral immune responses in Drosophila is mediated by the endosomal ARF1-Asrij axis. Sci. Rep. 2017, 7, 118. [Google Scholar] [CrossRef] [Green Version]
- Cronin, S.J.; Nehme, N.T.; Limmer, S.; Liegeois, S.; Pospisilik, J.A.; Schramek, D.; Leibbrandt, A.; Simoes Rde, M.; Gruber, S.; Puc, U.; et al. Genome-wide RNAi screen identifies genes involved in intestinal pathogenic bacterial infection. Science 2009, 325, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, N.; Ehret-Sabatier, L.; Brun, R.; Zachary, D.; Bulet, P.; Imler, J.L. Immune response of Drosophila melanogaster to infection with the flagellate parasite Crithidia spp. Insect. Biochem. Mol. Biol. 2001, 31, 129–137. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, R.; Meade, A.; Potter, S.; Cooper, R.L. Rapid and Direct Action of Lipopolysaccharide (LPS) on Skeletal Muscle of Larval Drosophila. Biology 2021, 10, 1235. [Google Scholar] [CrossRef] [PubMed]
- Lau, G.W.; Goumnerov, B.C.; Walendziewicz, C.L.; Hewitson, J.; Xiao, W.; Mahajan-Miklos, S.; Tompkins, R.G.; Perkins, L.A.; Rahme, L.G. The Drosophila melanogaster toll pathway participates in resistance to infection by the gram-negative human pathogen Pseudomonas aeruginosa. Infect. Immun. 2003, 71, 4059–4066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linhartova, I.; Bumba, L.; Masin, J.; Basler, M.; Osicka, R.; Kamanova, J.; Prochazkova, K.; Adkins, I.; Hejnova-Holubova, J.; Sadilkova, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef] [Green Version]
- Sebald, J.; Morettini, S.; Podhraski, V.; Lass-Florl, C.; Lusser, A. CHD1 contributes to intestinal resistance against infection by P. aeruginosa in Drosophila melanogaster. PLoS ONE 2012, 7, e43144. [Google Scholar] [CrossRef]
- Imler, J.L.; Hoffmann, J.A. Toll receptors in innate immunity. Trends Cell. Biol. 2001, 11, 304–311. [Google Scholar] [CrossRef]
- Elrod-Erickson, M.; Mishra, S.; Schneider, D. Interactions between the cellular and humoral immune responses in Drosophila. Curr. Biol. 2000, 10, 781–784. [Google Scholar] [CrossRef] [Green Version]
- Valanne, S.; Wang, J.H.; Ramet, M. The Drosophila Toll signaling pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Valanne, S.; Myllymaki, H.; Kallio, J.; Schmid, M.R.; Kleino, A.; Murumagi, A.; Airaksinen, L.; Kotipelto, T.; Kaustio, M.; Ulvila, J.; et al. Genome-wide RNA interference in Drosophila cells identifies G protein-coupled receptor kinase 2 as a conserved regulator of NF-kappaB signaling. J. Immunol. 2010, 184, 6188–6198. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Liu, H.; Skolnik, E.Y.; Manley, J.L. Physical and functional interactions between Drosophila TRAF2 and Pelle kinase contribute to Dorsal activation. Proc. Natl. Acad. Sci. USA 2001, 98, 8596–8601. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, K.; Silverman, N. Positive and negative regulation of the Drosophila immune response. BMB Rep. 2008, 41, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjum, S.G.; Xu, W.; Nikkholgh, N.; Basu, S.; Nie, Y.; Thomas, M.; Satyamurti, M.; Budnik, B.A.; Ip, Y.T.; Veraksa, A. Regulation of Toll signaling and inflammation by beta-arrestin and the SUMO protease Ulp1. Genetics 2013, 195, 1307–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchon, N.; Poidevin, M.; Kwon, H.M.; Guillou, A.; Sottas, V.; Lee, B.L.; Lemaitre, B. A single modular serine protease integrates signals from pattern-recognition receptors upstream of the Drosophila Toll pathway. Proc. Natl. Acad. Sci. USA 2009, 106, 12442–12447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooi, J.Y.; Yagi, Y.; Hu, X.; Ip, Y.T. The Drosophila Toll-9 activates a constitutive antimicrobial defense. EMBO Rep. 2002, 3, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Parthier, C.; Stelter, M.; Ursel, C.; Fandrich, U.; Lilie, H.; Breithaupt, C.; Stubbs, M.T. Structure of the Toll-Spatzle complex, a molecular hub in Drosophila development and innate immunity. Proc. Natl. Acad. Sci. USA 2014, 111, 6281–6286. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.; Li, C.F.; He, Z.; Lu, Y.; Liu, X.S.; Wang, Y.F.; Ip, Y.T.; Strand, M.R.; Yu, X.Q. Toll family members bind multiple Spatzle proteins and activate antimicrobial peptide gene expression in Drosophila. J. Biol. Chem. 2019, 294, 10172–10181. [Google Scholar] [CrossRef]
- Gordon, M.D.; Dionne, M.S.; Schneider, D.S.; Nusse, R. WntD is a feedback inhibitor of Dorsal/NF-kappaB in Drosophila development and immunity. Nature 2005, 437, 746–749. [Google Scholar] [CrossRef]
- Haghayeghi, A.; Sarac, A.; Czerniecki, S.; Grosshans, J.; Schock, F. Pellino enhances innate immunity in Drosophila. Mech. Dev. 2010, 127, 301–307. [Google Scholar] [CrossRef]
- Hoffmann, J.A.; Reichhart, J.M. Drosophila innate immunity: An evolutionary perspective. Nat. Immunol. 2002, 3, 121–126. [Google Scholar] [CrossRef]
- Kleino, A.; Silverman, N. The Drosophila IMD pathway in the activation of the humoral immune response. Dev. Comp. Immunol. 2014, 42, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Choe, K.M.; Werner, T.; Stoven, S.; Hultmark, D.; Anderson, K.V. Requirement for a peptidoglycan recognition protein (PGRP) in Relish activation and antibacterial immune responses in Drosophila. Science 2002, 296, 359–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballinger-Boone, C.; Anyagaligbo, O.; Bernard, J.; Bierbower, S.M.; Dupont-Versteegden, E.E.; Ghoweri, A.; Greenhalgh, A.; Harrison, D.; Istas, O.; McNabb, M.; et al. The effects of bacterial endotoxin (LPS) on cardiac and synaptic function in various animal models: Larval Drosophila, crayfish, crab, and rodent. Int. J. Zool. Res. 2020, 16, 33–62. [Google Scholar] [CrossRef]
- Myllymaki, H.; Valanne, S.; Ramet, M. The Drosophila imd signaling pathway. J. Immunol. 2014, 192, 3455–3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, A.; Yano, T.; Terashima, J.; Iwashita, S.; Oshima, Y.; Kurata, S. Cooperative regulation of the induction of the novel antibacterial Listericin by peptidoglycan recognition protein LE and the JAK-STAT pathway. J. Biol. Chem. 2010, 285, 15731–15738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinander, A.; Runchel, C.; Tenev, T.; Chen, L.; Kim, C.H.; Ribeiro, P.S.; Broemer, M.; Leulier, F.; Zvelebil, M.; Silverman, N.; et al. Ubiquitylation of the initiator caspase DREDD is required for innate immune signalling. EMBO J. 2012, 31, 2770–2783. [Google Scholar] [CrossRef] [Green Version]
- Kleino, A.; Myllymaki, H.; Kallio, J.; Vanha-aho, L.M.; Oksanen, K.; Ulvila, J.; Hultmark, D.; Valanne, S.; Ramet, M. Pirk is a negative regulator of the Drosophila Imd pathway. J. Immunol. 2008, 180, 5413–5422. [Google Scholar] [CrossRef] [Green Version]
- Ji, S.; Luo, Y.; Cai, Q.; Cao, Z.; Zhao, Y.; Mei, J.; Li, C.; Xia, P.; Xie, Z.; Xia, Z.; et al. LC Domain-Mediated Coalescence Is Essential for Otu Enzymatic Activity to Extend Drosophila Lifespan. Mol. Cell. 2019, 74, 363–377.e5. [Google Scholar] [CrossRef]
- Shibata, T.; Sekihara, S.; Fujikawa, T.; Miyaji, R.; Maki, K.; Ishihara, T.; Koshiba, T.; Kawabata, S. Transglutaminase-catalyzed protein-protein cross-linking suppresses the activity of the NF-kappaB-like transcription factor relish. Sci. Signal. 2013, 6, ra61. [Google Scholar] [CrossRef]
- Goto, A.; Matsushita, K.; Gesellchen, V.; El Chamy, L.; Kuttenkeuler, D.; Takeuchi, O.; Hoffmann, J.A.; Akira, S.; Boutros, M.; Reichhart, J.M. Akirins are highly conserved nuclear proteins required for NF-kappaB-dependent gene expression in Drosophila and mice. Nat. Immunol. 2008, 9, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Zaidman-Remy, A.; Herve, M.; Poidevin, M.; Pili-Floury, S.; Kim, M.S.; Blanot, D.; Oh, B.H.; Ueda, R.; Mengin-Lecreulx, D.; Lemaitre, B. The Drosophila amidase PGRP-LB modulates the immune response to bacterial infection. Immunity 2006, 24, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Naitza, S.; Rosse, C.; Kappler, C.; Georgel, P.; Belvin, M.; Gubb, D.; Camonis, J.; Hoffmann, J.A.; Reichhart, J.M. The Drosophila immune defense against gram-negative infection requires the death protein dFADD. Immunity 2002, 17, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Langmann, C.; Harden, N.; Aigaki, T. The RING-finger scaffold protein Plenty of SH3s targets TAK1 to control immunity signalling in Drosophila. EMBO Rep. 2005, 6, 1082–1087. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.H.; Kim, S.H.; Lee, H.Y.; Bai, J.Y.; Nam, Y.D.; Bae, J.W.; Lee, D.G.; Shin, S.C.; Ha, E.M.; Lee, W.J. Innate immune homeostasis by the homeobox gene caudal and commensal-gut mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, T.; Reichhart, J.M.; Hoffmann, J.A.; Royet, J. Drosophila Toll is activated by Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature 2001, 414, 756–759. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, E.; Spellman, P.T.; Tzou, P.; Rubin, G.M.; Lemaitre, B. The Toll and Imd pathways are the major regulators of the immune response in Drosophila. EMBO J. 2002, 21, 2568–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, U.M.; Bjorklund, G.; Ip, Y.T.; Engstrom, Y. The dorsal-related immunity factor, Dif, is a sequence-specific trans-activator of Drosophila Cecropin gene expression. EMBO J. 1995, 14, 3146–3158. [Google Scholar] [CrossRef]
- Kim, L.K.; Choi, U.Y.; Cho, H.S.; Lee, J.S.; Lee, W.B.; Kim, J.; Jeong, K.; Shim, J.; Kim-Ha, J.; Kim, Y.J. Downregulation of NF-kappaB target genes by the AP-1 and STAT complex during the innate immune response in Drosophila. PLoS Biol. 2007, 5, e238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.S.; Ip, Y.T. Interaction and specificity of Rel-related proteins in regulating Drosophila immunity gene expression. J. Biol. Chem. 1999, 274, 21355–21361. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, S.A.; Lin, S.J.H.; Wasserman, S.A. Short-Form Bomanins Mediate Humoral Immunity in Drosophila. J. Innate Immun. 2018, 10, 306–314. [Google Scholar] [CrossRef]
- Hanson, M.A.; Cohen, L.B.; Marra, A.; Iatsenko, I.; Wasserman, S.A.; Lemaitre, B. The Drosophila Baramicin polypeptide gene protects against fungal infection. PLoS Pathog. 2021, 17, e1009846. [Google Scholar] [CrossRef]
- Shokal, U.; Eleftherianos, I. The Drosophila Thioester containing Protein-4 participates in the induction of the cellular immune response to the pathogen Photorhabdus. Dev. Comp. Immunol. 2017, 76, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, A.W.; Lindsay, S.A.; Wasserman, S.A. An effector Peptide family required for Drosophila toll-mediated immunity. PLoS Pathog. 2015, 11, e1004876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokal, U.; Kopydlowski, H.; Harsh, S.; Eleftherianos, I. Thioester-Containing Proteins 2 and 4 Affect the Metabolic Activity and Inflammation Response in Drosophila. Infect. Immun. 2018, 86, e00810-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bou Aoun, R.; Hetru, C.; Troxler, L.; Doucet, D.; Ferrandon, D.; Matt, N. Analysis of thioester-containing proteins during the innate immune response of Drosophila melanogaster. J. Innate Immun. 2011, 3, 52–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, C.; Silverman, N. p38b and JAK-STAT signaling protect against Invertebrate iridescent virus 6 infection in Drosophila. PLoS Pathog. 2018, 14, e1007020. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Poidevin, M.; Lemaitre, B. The Drosophila MAPK p38c regulates oxidative stress and lipid homeostasis in the intestine. PLoS Genet. 2014, 10, e1004659. [Google Scholar] [CrossRef]
- Huber, W.; von Heydebreck, A.; Sultmann, H.; Poustka, A.; Vingron, M. Variance stabilization applied to microarray data calibration and to the quantification of differential expression. Bioinformatics 2002, 18 (Suppl. 1), S96–S104. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Soldano, A.; Alpizar, Y.A.; Boonen, B.; Franco, L.; Lopez-Requena, A.; Liu, G.; Mora, N.; Yaksi, E.; Voets, T.; Vennekens, R.; et al. Gustatory-mediated avoidance of bacterial lipopolysaccharides via TRPA1 activation in Drosophila. Elife 2016, 5, e13133. [Google Scholar] [CrossRef]
- Boonen, B.; Alpizar, Y.A.; Meseguer, V.M.; Talavera, K. TRP Channels as Sensors of Bacterial Endotoxins. Toxins 2018, 10, 326. [Google Scholar] [CrossRef] [Green Version]
- Khalil, S.; Jacobson, E.; Chambers, M.C.; Lazzaro, B.P. Systemic bacterial infection and immune defense phenotypes in Drosophila melanogaster. J. Vis. Exp. 2015, 13, e52613. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toll Pathway | ||
|---|---|---|
| Gene Name | Gene Symbol | Biological Process |
| Baramicin A2 | BaraA2 | defense response |
| Bomanin Short 1 | BomS1 | response to bacteria |
| Bomanin Short 2 | BomS2 | defense response |
| Bomanin Short 3 | BomS3 | response to bacteria |
| cactus | cact | negative regulation of Toll signaling pathway |
| Defensin | Def | humoral immune response |
| dorsal | dl | dorsal/ventral axis specification |
| Drosomycin | Drs | defense response to protozoa |
| Drosomycin-like 1 | Drsl1 | defense response to fungi |
| Gram-negative bacteria binding protein 1 | GNBP1 | carbohydrate metabolic process |
| G protein-coupled receptor kinase 2 | Gprk2 | protein phosphorylation |
| Gram-positive Specific Serine protease | grass | proteolysis |
| kurtz | krz | locomotory exploration behavior |
| modular serine protease | modSP | proteolysis |
| Metchnikowin | Mtk | defense response to Gram-positive bacteria |
| Peptidoglycan recognition protein SA | PGRP-SA | innate immune response |
| Peptidoglycan recognition protein SD | PGRP-SD | innate immune response |
| pelle | pll | Toll signaling pathway |
| Spatzle-Processing Enzyme | SPE | defense response to Gram-positive bacteria |
| spheroide | sphe | proteolysis |
| spatzle | spz | defense response to Gram-positive bacteria |
| Thioester-containing protein 4 | Tep4 | innate immune response |
| Toll-9 | Toll-9 | signal transduction |
| Ulp1 | Ulp1 | negative regulation of Toll signaling pathway |
| wnt inhibitor of Dorsal | wntD | defense response to Gram-positive bacteria |
| Imd Pathway | ||
| Gene Name | Gene Symbol | Biological Process |
| Attacin-A | AttA | humoral immune response |
| Attacin-B | AttB | humoral immune response |
| Attacin-C | AttC | antibacterial humoral response |
| Attacin-D | AttD | response to wounding |
| bendless | ben | positive regulation of tumor necrosis factor-mediated signaling pathway |
| Cecropin pseudogene 1 | Cec-Ψ1 | pseudogene |
| Cecropin 2 | Cec2 | pseudogene |
| Cecropin A2 | CecA2 | antibacterial humoral response |
| Cecropin B | CecB | defense response to Gram-positive bacteria |
| Diptericin A | DptA | response to bacteria |
| Diptericin B | DptB | response to wounding |
| effete | eff | germ-line stem cell population maintenance |
| Fas-associated death domain | Fadd | peptidoglycan recognition protein signaling pathway |
| Immune deficiency | imd | response to bacteria |
| Peptidoglycan recognition protein LB | PGRP-LB | innate immune response |
| Peptidoglycan recognition protein LC | PGRP-LC | regulation of synaptic plasticity |
| poor Imd response upon knock-in | pirk | negative regulation of peptidoglycan recognition protein signaling pathway |
| Plenty of SH3s | POSH | response to peptidoglycan |
| Relish | Rel | peripheral nervous system neuron development |
| Imd-JNK Pathway | ||
| Gene Name | Gene Symbol | Biological Process |
| Activating transcription factor-2 | Atf-2 | positive regulation of transcription by RNA polymerase II |
| Octopamine receptor in mushroom bodies | Oamb | cellular calcium ion homeostasis |
| basket | bsk | cellular response to oxidative stress |
| hemipterous | hep | positive regulation of cell death |
| Jun-related antigen | Jra | wound healing |
| licorne | lic | JNK cascade |
| Phospholipase C at 21C | Plc21C | lipid metabolic process |
| Signal-transducer and activator of transcription protein at 92E | Stat92E | receptor signaling pathway via JAK-STAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chifiriuc, M.C.; Bologa, A.M.; Ratiu, A.C.; Ionascu, A.; Ecovoiu, A.A. Mutations of γCOP Gene Disturb Drosophila melanogaster Innate Immune Response to Pseudomonas aeruginosa. Int. J. Mol. Sci. 2022, 23, 6499. https://doi.org/10.3390/ijms23126499
Chifiriuc MC, Bologa AM, Ratiu AC, Ionascu A, Ecovoiu AA. Mutations of γCOP Gene Disturb Drosophila melanogaster Innate Immune Response to Pseudomonas aeruginosa. International Journal of Molecular Sciences. 2022; 23(12):6499. https://doi.org/10.3390/ijms23126499
Chicago/Turabian StyleChifiriuc, Mariana Carmen, Alexandru Marian Bologa, Attila Cristian Ratiu, Adrian Ionascu, and Alexandru Al. Ecovoiu. 2022. "Mutations of γCOP Gene Disturb Drosophila melanogaster Innate Immune Response to Pseudomonas aeruginosa" International Journal of Molecular Sciences 23, no. 12: 6499. https://doi.org/10.3390/ijms23126499