
Cardiac Progenitor Cells and Adipocyte Stem Cells from Same Patients Exhibit In Vitro Functional Differences

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Characterization of Cells Derived from RAA and EAT
2.2. CPCs Migrate Faster than ASCs
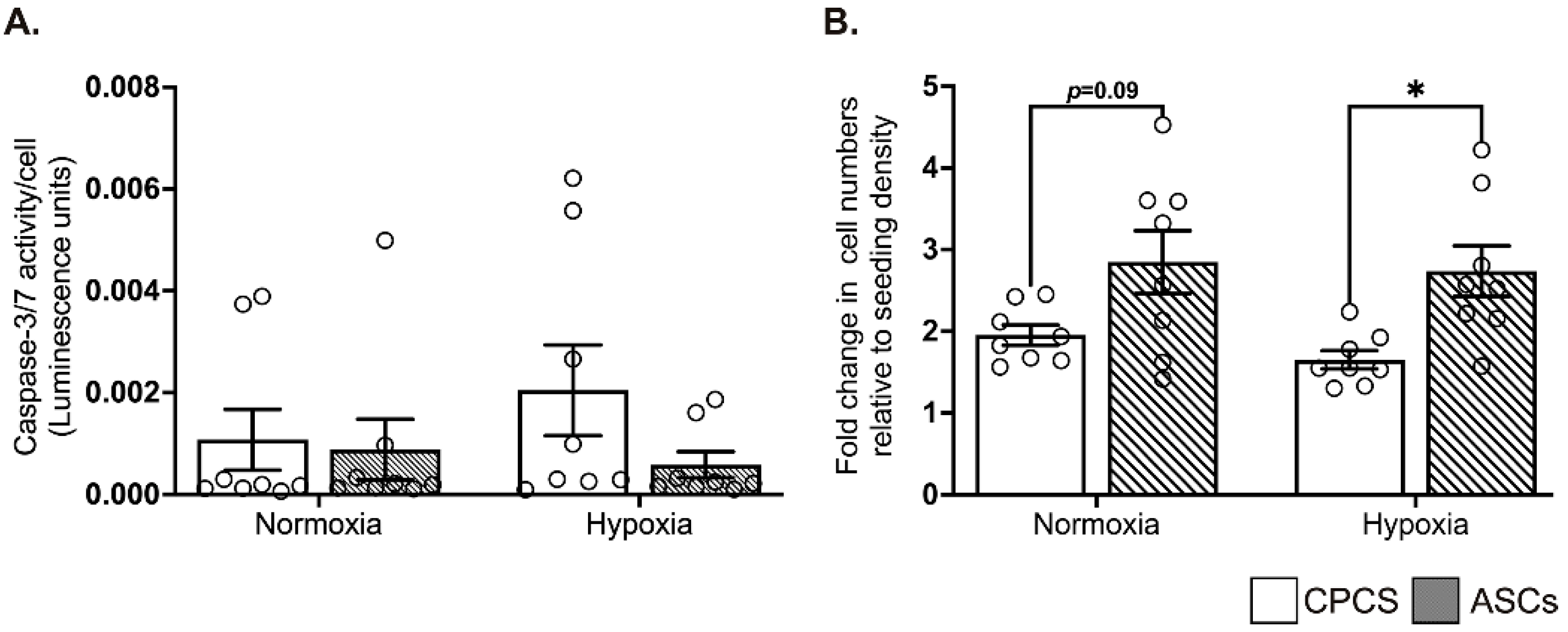
2.3. CPCs and ASCs Were Resistant to Hypoxia-Induced Apoptotic Cell Death
2.4. ASCs Showed Better Proliferation in Serum-Deprived Normoxic and Hypoxic Conditions
2.5. Differential Secretion of Paracrine Cytokines by CPCs and ASCs
2.6. CPC CM Significantly Reduced Hypoxia-Induced Apoptotic Cell Death in AC-16 Cardiomyocytes
2.7. Both CPC and ASC CM Enhanced Tube Formation of HUVECs
3. Discussion
4. Methods
4.1. Ethical Approval
4.2. Tissue Collection
4.3. Cell Culture
4.3.1. Explant Culture of CPCs and ASCs
4.3.2. AC16 Cardiomyocytes
4.3.3. Human Umbilical Vein Endothelial Cells
4.4. Flow Cytometry Characterization of ASCs and CPCs
4.5. Wound-Healing Assay to Determine the Migration Potential of CPCs and ASCs
4.6. Exposure of CPCs and ASCs to Hypoxic Conditions and Collection of Conditioned Media
4.7. Apoptosis and Cell Proliferation Assay
4.8. Enzyme-Linked Immunosorbent Assay (ELISA) for Secretion of Pro-Survival and Angiogenic Factors in CM
4.9. Pro-Survival Effects of CM on Cardiomyocytes
4.10. Pro-Angiogenic Effects of CM on Endothelial Cells
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marketou, M.E.; Parthenakis, F.; Vardas, P.E. Pathological Left Ventricular Hypertrophy and Stem Cells: Current Evidence and New Perspectives. Stem Cells Int. 2016, 2016, 5720758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amini, H.; Rezaie, J.; Vosoughi, A.; Rahbarghazi, R.; Nouri, M. Cardiac progenitor cells application in cardiovascular disease. J. Cardiovasc. Thorac. Res. 2017, 9, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruun, K.; Schermer, E.; Sivendra, A.; Valaik, E.; Wise, R.B.; Said, R.; Bracht, J.R. Therapeutic applications of adipose-derived stem cells in cardiovascular disease. Am. J. Stem Cells 2018, 7, 94–103. [Google Scholar]
- Dixit, P.; Donnelly, H.; Edamatsu, M.; Galvin, I.; Bunton, R.; Katare, R. Progenitor cells from atria, ventricle and peripheral blood of the same patients exhibit functional differences associated with cardiac repair. Int. J. Cardiol. 2017, 228, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H. Adipose stem cells as a clinically available and effective source of adult stem cell therapy. Int. J. Stem Cells 2008, 1, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burchfield, J.S.; Dimmeler, S. Role of paracrine factors in stem and progenitor cell-mediated cardiac repair and tissue fibrosis. Fibrogenesis Tissue Repair 2008, 1, 4. [Google Scholar] [CrossRef] [Green Version]
- Debnath, T.; Chelluri, L.K. Standardization and quality assessment for clinical-grade mesenchymal stem cells from human adipose tissue. Hematol. Transfus. Cell Ther. 2019, 41, 7–16. [Google Scholar] [CrossRef]
- van Vliet, P.; Roccio, M.; Smits, A.M.; van Oorschot, A.A.; Metz, C.H.; van Veen, T.A.; Sluijter, J.P.; Doevendans, P.A.; Goumans, M.J. Progenitor cells isolated from the human heart: A potential cell source for regenerative therapy. Neth. Heart J. 2008, 16, 163–169. [Google Scholar] [CrossRef]
- Malliaras, K.; Makkar, R.R.; Smith, R.R.; Cheng, K.; Wu, E.; Bonow, R.O.; Marban, L.; Mendizabal, A.; Cingolani, E.; Johnston, P.V.; et al. Intracoronary cardiosphere-derived cells after myocardial infarction: Evidence of therapeutic regeneration in the final 1-year results of the CADUCEUS trial (CArdiosphere-Derived aUtologous stem CElls to reverse ventricUlar dySfunction). J. Am. Coll. Cardiol. 2014, 63, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Paitazoglou, C.; Bergmann, M.W.; Vrtovec, B.; Chamuleau, S.A.J.; van Klarenbosch, B.; Wojakowski, W.; Michalewska-Wludarczyk, A.; Gyongyosi, M.; Ekblond, A.; Haack-Sorensen, M.; et al. Rationale and design of the European multicentre study on Stem Cell therapy in IschEmic Non-treatable Cardiac diseasE (SCIENCE). Eur. J. Heart Fail. 2019, 21, 1032–1041. [Google Scholar] [CrossRef] [Green Version]
- Perin, E.C.; Sanz-Ruiz, R.; Sanchez, P.L.; Lasso, J.; Perez-Cano, R.; Alonso-Farto, J.C.; Perez-David, E.; Fernandez-Santos, M.E.; Serruys, P.W.; Duckers, H.J.; et al. Adipose-derived regenerative cells in patients with ischemic cardiomyopathy: The Precise Trial. Am. Heart J. 2014, 168, 88–95.e2. [Google Scholar] [CrossRef] [PubMed]
- Dixit, P.; Katare, R.G. Isolation and Characterization of Cardiac Progenitor Cells. Methods Mol. Biol. 2019, 2029, 161–173. [Google Scholar] [PubMed]
- Wen, Z.; Zheng, S.; Zhou, C.; Yuan, W.; Wang, J.; Wang, T. Bone marrow mesenchymal stem cells for post-myocardial infarction cardiac repair: MicroRNAs as novel regulators. J. Cell. Mol. Med. 2012, 16, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise review: Evidence for CD34 as a Common Marker for Diverse Progenitors. Stem Cells. 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Yang, M.; Wang, C.; Zhao, Q.; Liu, J.; Zhan, C.; Liu, Z.; Li, X.; Wang, W.; Yang, X. Effects of cytokines and chemokines on migration of mesenchymal stem cells following spinal cord injury. Neural Regen. Res. 2012, 7, 1106–1112. [Google Scholar]
- Kisselbach, L.; Merges, M.; Bossie, A.; Boyd, A. CD90 Expression on human primary cells and elimination of contaminating fibroblasts from cell cultures. Cytotechnology 2009, 59, 31–44. [Google Scholar] [CrossRef] [Green Version]
- McQuaig, R.; Dixit, P.; Yamauchi, A.; Van Hout, I.; Papannarao, J.B.; Bunton, R.; Parry, D.; Davis, P.; Katare, R. Combination of Cardiac Progenitor Cells From the Right Atrium and Left Ventricle Exhibits Synergistic Paracrine Effects In Vitro. Cell Transplant. 2020, 29, 963689720972328. [Google Scholar] [CrossRef]
- Dixit, P. Identifying the Best Autologous Progenitors for Cardiac Regeneration. Ph.D. Thesis, University of Otago, Dunedin, New Zealand, 2016. [Google Scholar]
- Hodgkinson, C.P.; Bareja, A.; Gomez, J.A.; Dzau, V.J. Emerging Concepts in Paracrine Mechanisms in Regenerative Cardiovascular Medicine and Biology. Circ. Res. 2016, 118, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Gnecchi, M.; Zhang, Z.; Ni, A.; Dzau, V.J. Paracrine mechanisms in adult stem cell signaling and therapy. Circ. Res. 2008, 103, 1204–1219. [Google Scholar] [CrossRef]
- Borowiec, A.S.; Hague, F.; Harir, N.; Guenin, S.; Guerineau, F.; Gouilleux, F.; Roudbaraki, M.; Lassoued, K.; Ouadid-Ahidouch, H. IGF-1 activates hEAG K(+) channels through an Akt-dependent signaling pathway in breast cancer cells: Role in Cell Proliferation. J. Cell. Physiol. 2007, 212, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Mairet-Coello, G.; Tury, A.; DiCicco-Bloom, E. Insulin-like growth factor-1 promotes G(1)/S cell cycle progression through bidirectional regulation of cyclins and cyclin-dependent kinase inhibitors via the phosphatidylinositol 3-kinase/Akt pathway in developing rat cerebral cortex. J. Neurosci. 2009, 29, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Kakudo, N.; Morimoto, N.; Ogawa, T.; Taketani, S.; Kusumoto, K. Hypoxia Enhances Proliferation of Human Adipose-Derived Stem Cells via HIF-1a Activation. PLoS ONE 2015, 10, e0139890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauretti, A.; Spaans, S.; Bax, N.A.M.; Sahlgren, C.; Bouten, C.V.C. Cardiac Progenitor Cells and the Interplay with Their Microenvironment. Stem Cells Int. 2017, 2017, 7471582. [Google Scholar] [CrossRef] [Green Version]
- Rehman, J.; Traktuev, D.; Li, J.; Merfeld-Clauss, S.; Temm-Grove, C.J.; Bovenkerk, J.E.; Pell, C.L.; Johnstone, B.H.; Considine, R.V.; March, K.L. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation 2004, 109, 1292–1298. [Google Scholar] [CrossRef]
- Grayson, W.L.; Zhao, F.; Izadpanah, R.; Bunnell, B.; Ma, T. Effects of hypoxia on human mesenchymal stem cell expansion and plasticity in 3D constructs. J. Cell. Physiol. 2006, 207, 331–339. [Google Scholar] [CrossRef]
- Yan, X.; Ehnert, S.; Culmes, M.; Bachmann, A.; Seeliger, C.; Schyschka, L.; Wang, Z.; Rahmanian-Schwarz, A.; Stockle, U.; De Sousa, P.A.; et al. 5-azacytidine improves the osteogenic differentiation potential of aged human adipose-derived mesenchymal stem cells by DNA demethylation. PLoS ONE 2014, 9, e90846. [Google Scholar] [CrossRef] [Green Version]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Culmes, M.; Eckstein, H.H.; Burgkart, R.; Nussler, A.K.; Guenther, M.; Wagner, E.; Pelisek, J. Endothelial differentiation of adipose-derived mesenchymal stem cells is improved by epigenetic modifying drug BIX-01294. Eur. J. Cell Biol. 2013, 92, 70–79. [Google Scholar] [CrossRef]
- Campagnolo, P.; Katare, R.; Madeddu, P. Realities and misconceptions on the pericytes role in tissue repair. Regen. Med. 2018, 13, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Brugarolas, J.; Kaelin, W.G. Dysregulation of HIF and VEGF is a unifying feature of the familial hamartoma syndromes. Cancer Cell. 2004, 6, 7–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottet, D.; Dumont, V.; Deccache, Y.; Demazy, C.; Ninane, N.; Raes, M.; Michiels, C. Regulation of hypoxia-inducible factor-1alpha protein level during hypoxic conditions by the phosphatidylinositol 3-kinase/Akt/glycogen synthase kinase 3beta pathway in HepG2 cells. J. Biol. Chem. 2003, 278, 31277–31285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Ko, J.; Ju, C.; Eltzschig, H.K. Hypoxia signaling in human diseases and therapeutic targets. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kocan, B.; Maziarz, A.; Tabarkiewicz, J.; Ochiya, T.; Banas-Zabczyk, A. Trophic Activity and Phenotype of Adipose Tissue-Derived Mesenchymal Stem Cells as a Background of Their Regenerative Potential. Stem Cells Int. 2017, 2017, 1653254. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.M.; Nesti, C.; Palenzuela, L.; Walker, W.F.; Hernandez, E.; Protas, L.; Hirano, M.; Isaac, N.D. Novel cell lines derived from adult human ventricular cardiomyocytes. J. Mol. Cell. Cardiol. 2005, 39, 133–147. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Number | Age | Sex | BMI | Diabetes | Hypertension | Ejection Fraction (%) |
|---|---|---|---|---|---|---|
| 487 | 72 | F | 39.0 | N | N | 55–60 |
| 493 | 64 | F | 26.4 | N | Y | 60 |
| 500 | 69 | M | 35.1 | N | N | 53.1 |
| 765 | 61 | M | 34.4 | N | N | 57.3 |
| 769 | 70 | M | 25.7 | N | Y | 55–60 |
| 772 | 67 | F | 30.5 | N | Y | 71.6 |
| 779 | 67 | M | 27.3 | N | N | 50.6 |
| 782 | 67 | M | 34.5 | N | Y | 45–50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yee-Goh, A.S.; Yamauchi, A.; van Hout, I.; Bellae Papannarao, J.; Sugunesegran, R.; Parry, D.; Davis, P.; Katare, R. Cardiac Progenitor Cells and Adipocyte Stem Cells from Same Patients Exhibit In Vitro Functional Differences. Int. J. Mol. Sci. 2022, 23, 5588. https://doi.org/10.3390/ijms23105588
Yee-Goh AS, Yamauchi A, van Hout I, Bellae Papannarao J, Sugunesegran R, Parry D, Davis P, Katare R. Cardiac Progenitor Cells and Adipocyte Stem Cells from Same Patients Exhibit In Vitro Functional Differences. International Journal of Molecular Sciences. 2022; 23(10):5588. https://doi.org/10.3390/ijms23105588
Chicago/Turabian StyleYee-Goh, Anthony Soonseng, Atsushi Yamauchi, Isabelle van Hout, Jayanthi Bellae Papannarao, Ramanen Sugunesegran, Dominic Parry, Philip Davis, and Rajesh Katare. 2022. "Cardiac Progenitor Cells and Adipocyte Stem Cells from Same Patients Exhibit In Vitro Functional Differences" International Journal of Molecular Sciences 23, no. 10: 5588. https://doi.org/10.3390/ijms23105588