
Ipsilateral and Contralateral Interactions in Spinal Locomotor Circuits Mediated by V1 Neurons: Insights from Computational Modeling



Abstract
:1. Introduction
2. Results
2.1. Model Description
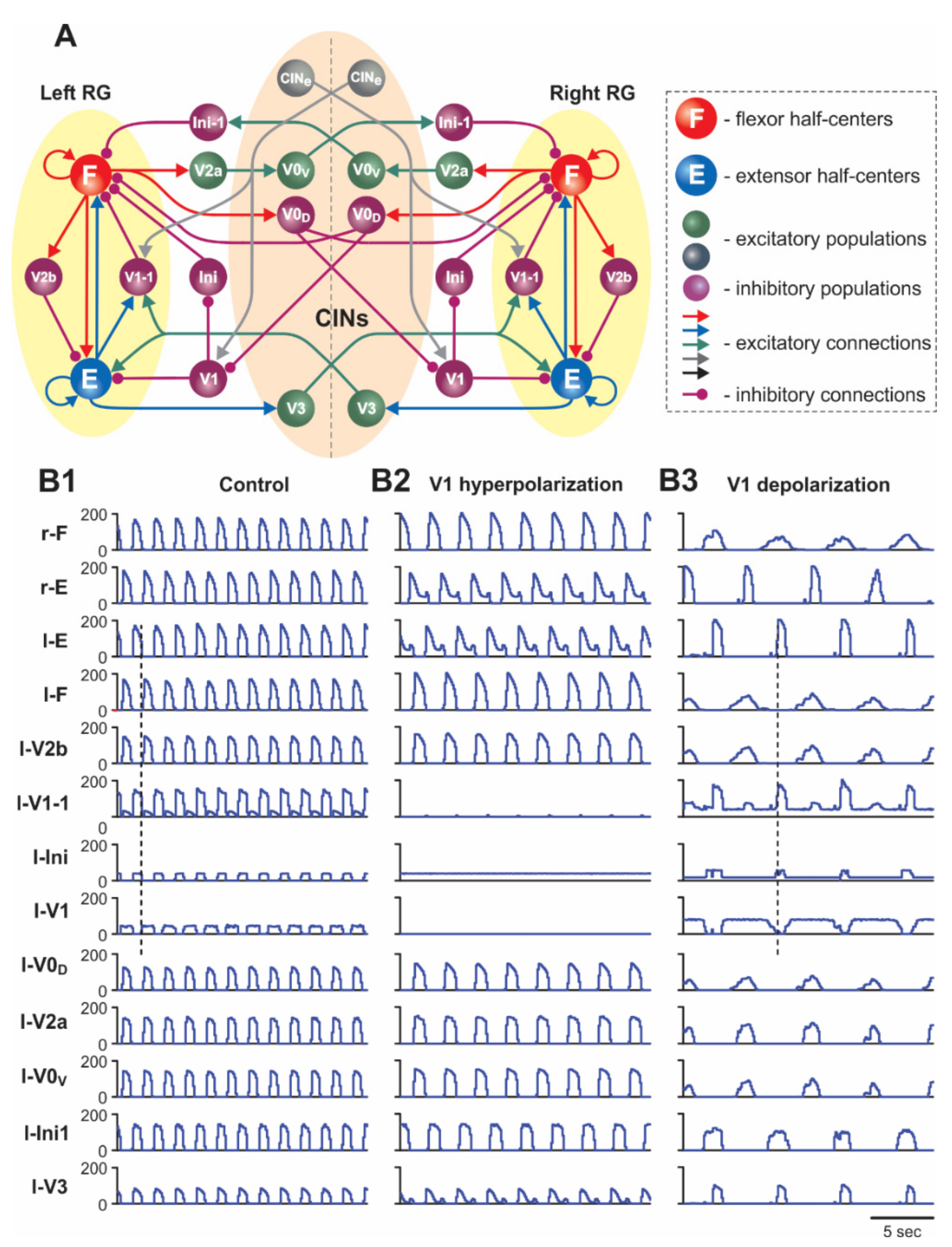
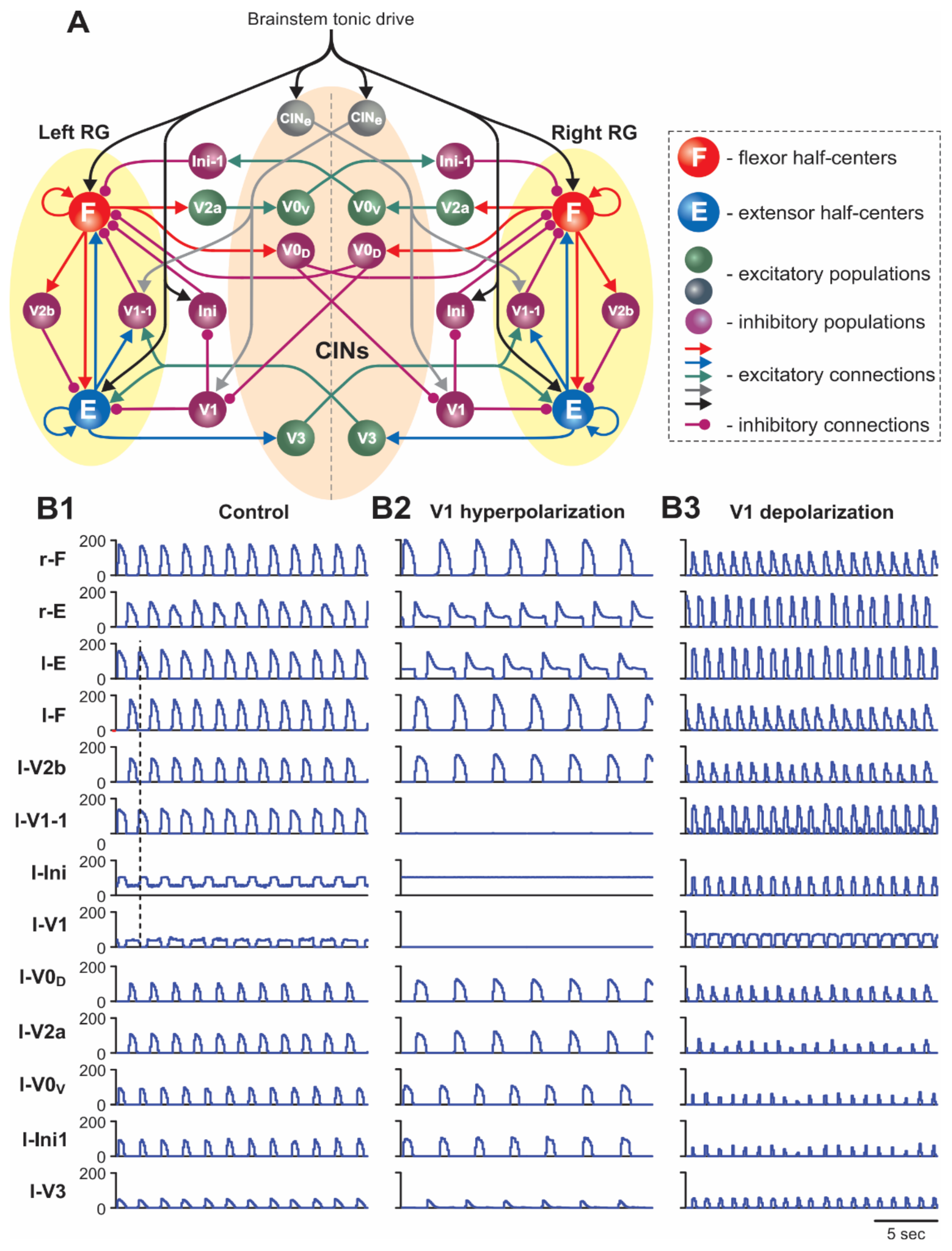
2.1.1. Architecture of Spinal Circuits in the Model
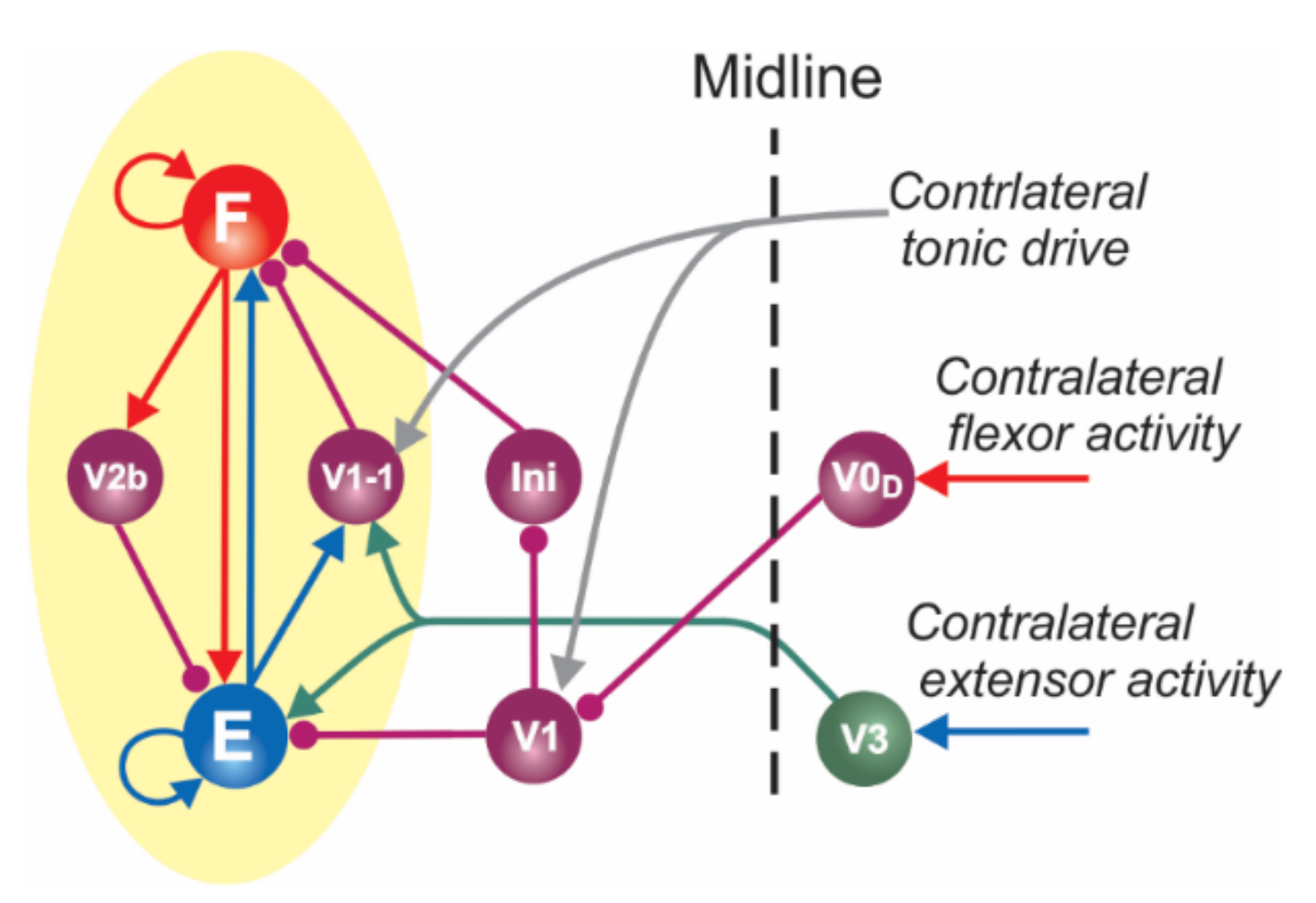
2.1.2. Two Functionally Distinct Types of V1 Populations in the Model and Their Connectivity
2.2. Simulation of Drug-Induced Fictive Locomotion
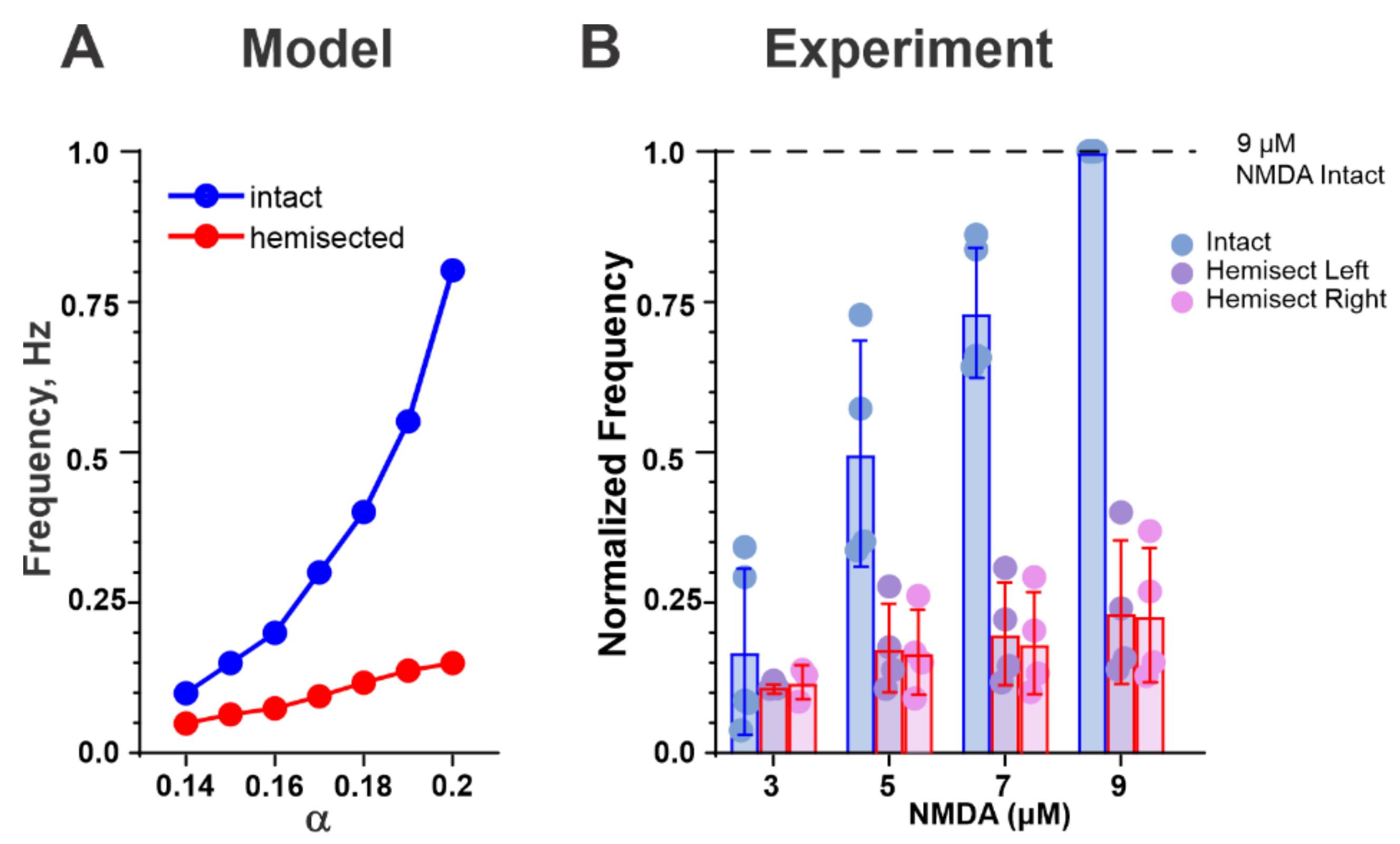
2.3. Changes in the Drug-Induced Locomotor Oscillations after Simulated Hemisection
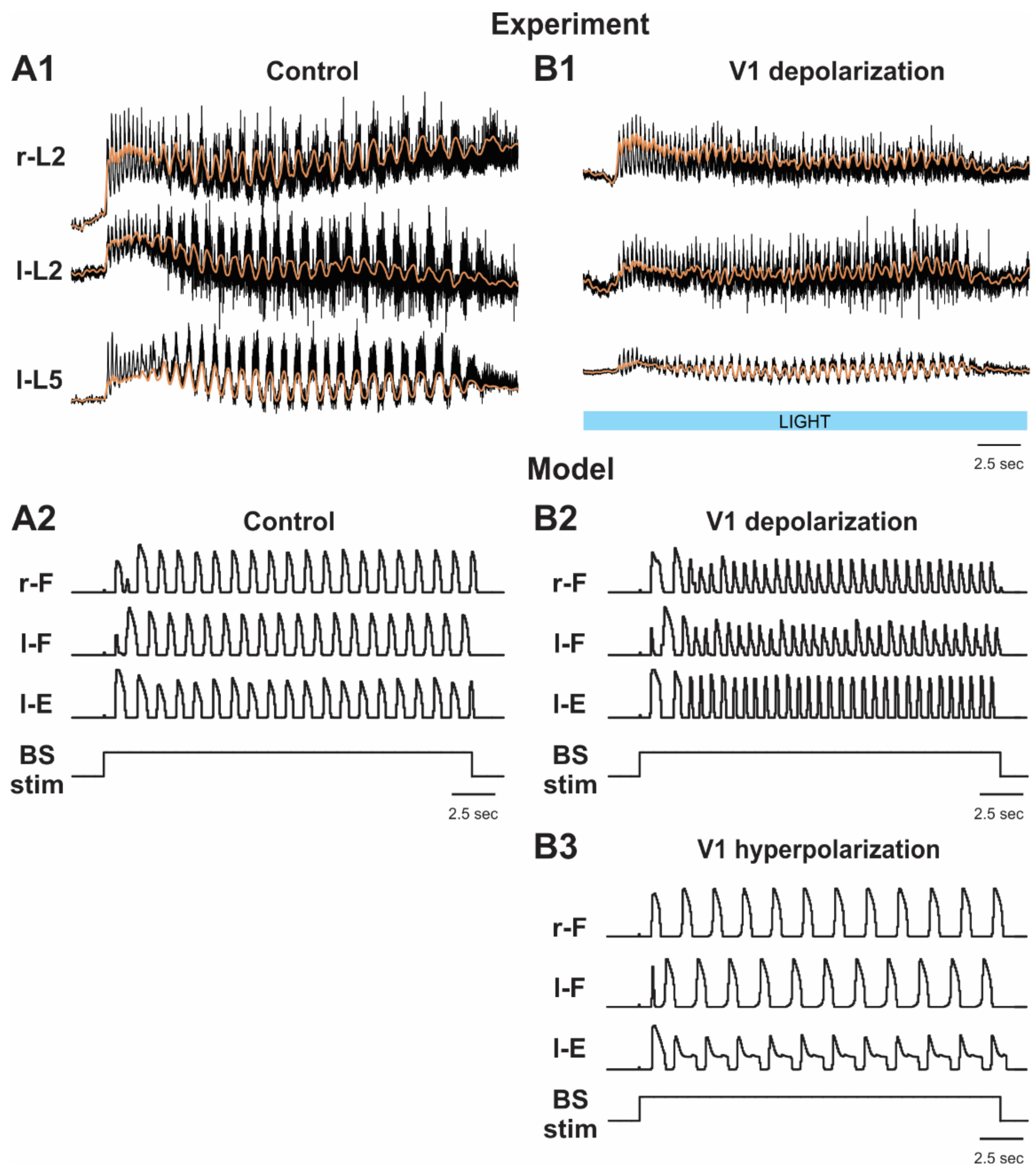
2.4. Simulation of Brainstem Stimulation-Evoked Fictive Locomotion
2.5. Simulating the Effects of Optogenetic Silencing and Activation of V1 Neurons on the Locomotor Rhythm and Pattern
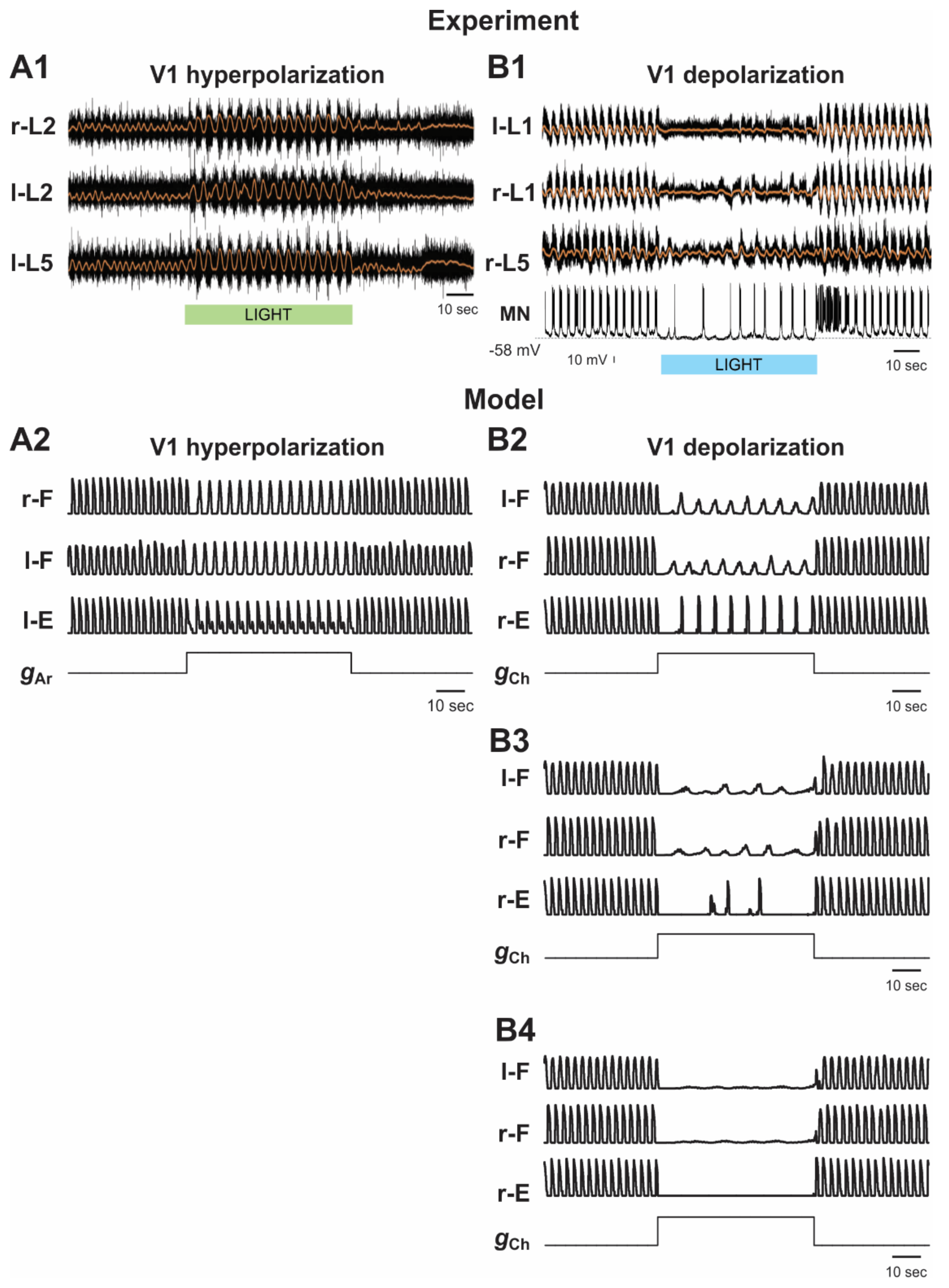
2.5.1. Silencing and Activation of V1 Neurons during Drug-Induced Fictive Locomotion in the Intact Cord
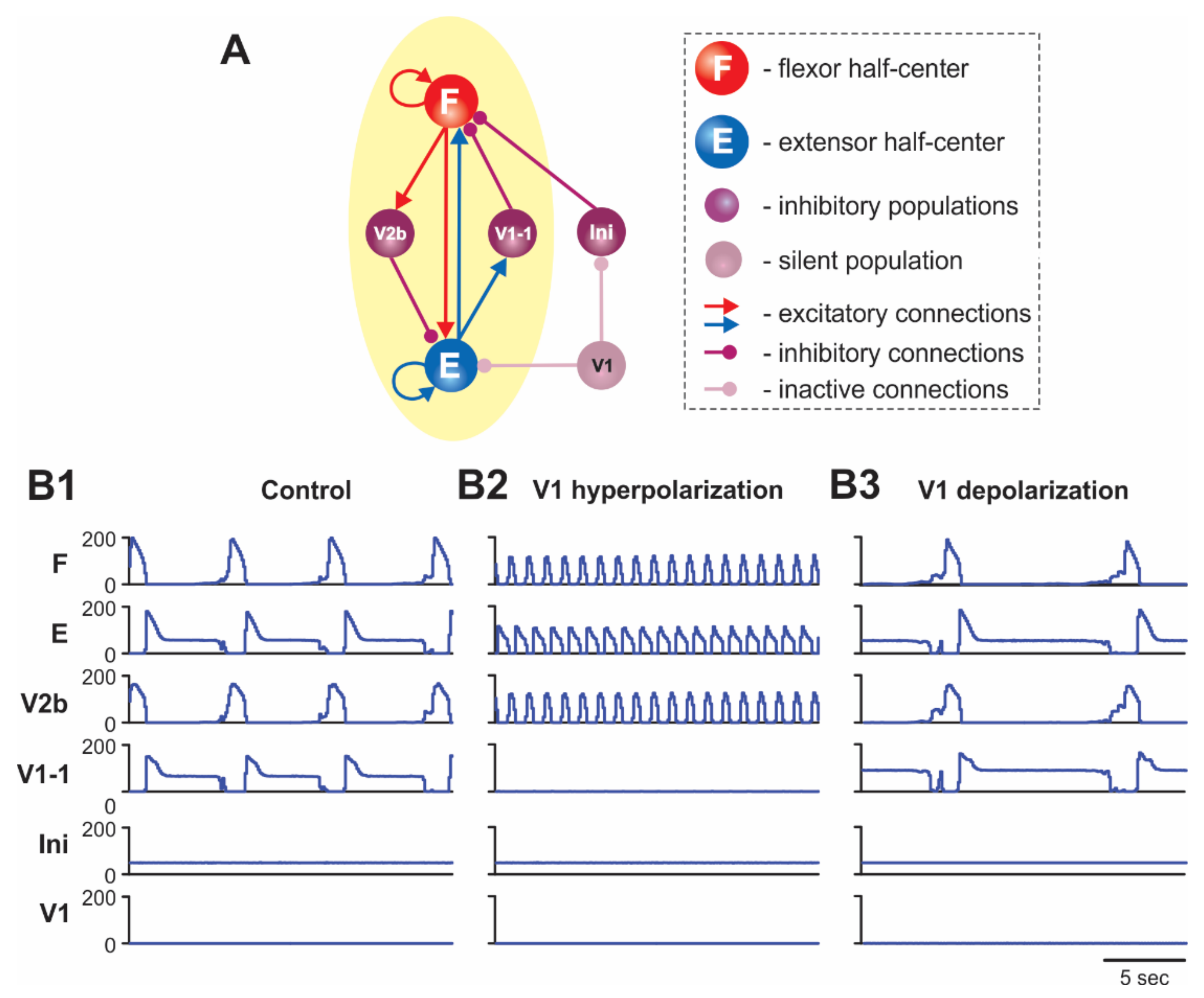
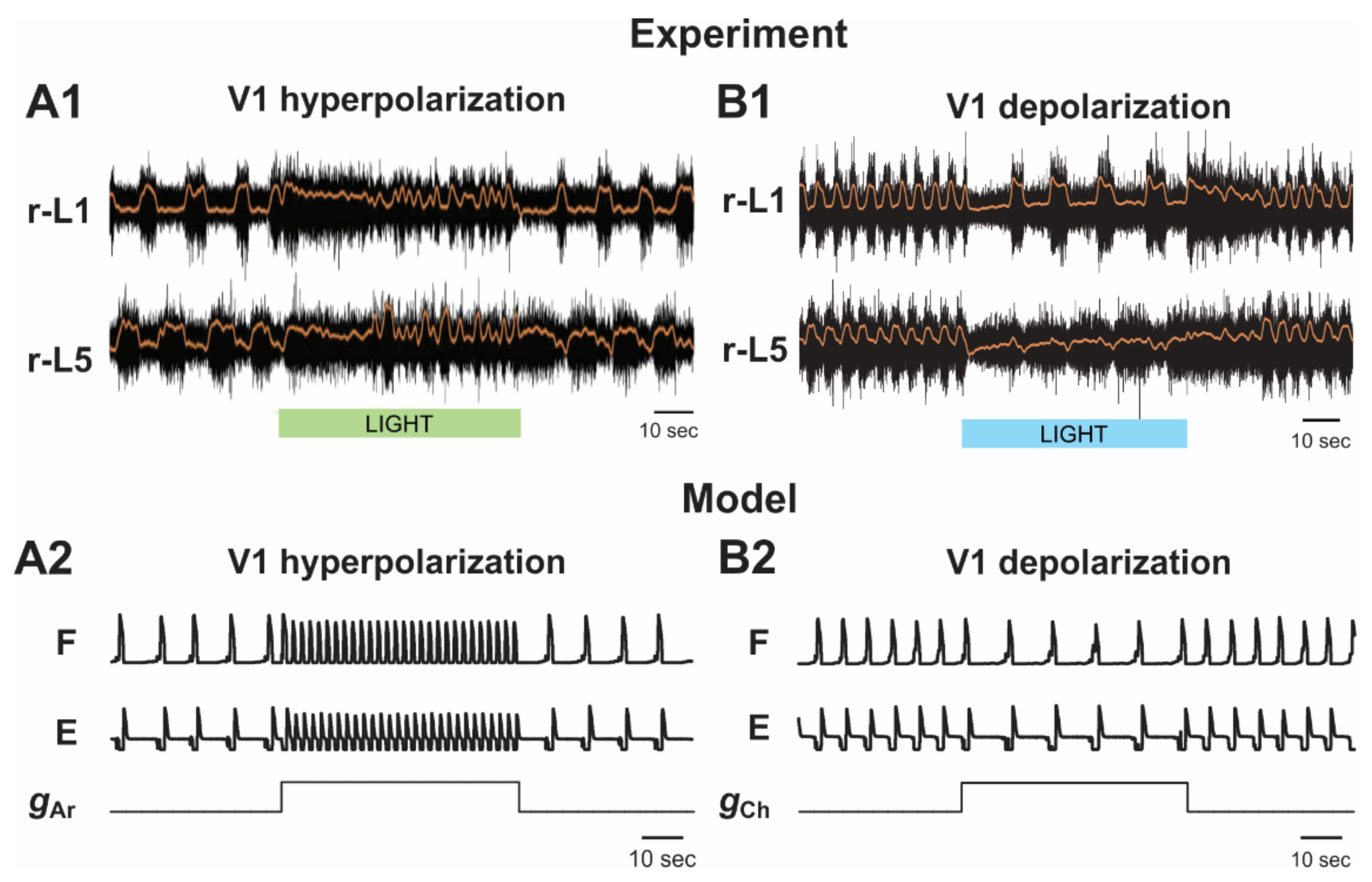
2.5.2. Silencing and Activation of V1 Neurons during Drug-Induced Fictive Locomotion in the Isolated Hemicord
2.5.3. Activation and Silencing of V1 Neurons during Brainstem Stimulation-Evoked Fictive Locomotion
3. Discussion
3.1. Modeling the Locomotor Circuits in the Mammalian Spinal Cord
3.2. Two Distinct Types of Populations of V1 Neurons within the Locomotor Spinal Circuitry and Their Involvement in Flexor–Extensor Alternation and Control of Oscillation Frequency
3.3. Different Effects of V1 Activations during Drug-Induced and Brainstem Stimulation-Evoked Fictive Locomotion
3.4. Reduction of Frequency of Drug-Induced Fictive Locomotion following Midsagittal Hemisection
3.5. Model Limitations
4. Materials and Methods
4.1. Modeling Methods
4.1.1. Modeling Single Neurons and Neuronal Populations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Neuron Type | N, Number of Neurons | , mS/cm2 | , mS/cm2 | , mS/cm2 | , mS/cm2 | , mV |
| F | 200 | 25 | 0.75 (±0.00375) | 2 | 0.07 | −76.8 (±0.77) |
| E | 100 | 25 | 0.75 (±0.00375) | 2 | 0.07 | −72 (±0.72) |
| V2b | 100 | 10 | 5 | 0.1 | −80.4 (±1.68) | |
| V1-1 | 100 | 10 | 5 | 0.1 | −90 (±1.8) | |
| V1 | 100 | 10 | 5 | 0.1 | −90 (±1.8) | |
| Ini | 50 | 10 | 5 | 0.1 | −60 (±1.2) | |
| Ini1 | 50 | 10 | 5 | 0.1 | −76.8 (±1.57) | |
| V2a | 50 | 40 | 5 | 0.8 | −72.6 (±1.45) | |
| V0V | 50 | 10 | 5 | 0.1 | −74.4 (±1.5) | |
| V0D | 50 | 10 | 5 | 0.1 | −81.6 (±2.45) | |
| V3 | 100 | 10 | 5 | 0.1 | −81.6 (±2.45) | |
| CINe | 100 | 10 | 5 | 0.1 | −60 (±2.04) |
4.1.2. Generation of Rhythmic Activity in the Model and Application of Photostimulation
4.1.3. Computer Simulations
4.2. Experimental Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, T.G. The intrinsic factors in the act of progression in the mammal. Proc. R. Soc. Lond. Ser. B 1911, 84, 308–319. [Google Scholar]
- Brown, T.G. On the nature of the fundamental activity of the nervous centres; together with an analysis of the conditioning of rhythmic activity in progression and a theory of the evolution of function in the nervous system. J. Physiol. 1914, 48, 18–46. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S. Control of locomotion in bipeds, tetrapods, and fish. In Handbook of Physiology, the Nervous System, Motor Control; Waverly Press: Baltimore, MD, USA, 1981. [Google Scholar]
- Rossignol, S. Neural control of stereotypic limb movements. In Handbook of Physiology. Sect 12. Exercise: Regulation and Integration of Multiple Systems; Rowell, L.B., Sheperd, J.T., Eds.; American Physiological Society: Rockville, MD, USA, 1996; pp. 173–216. [Google Scholar]
- Orlovsky, G.N.; Deliagina, T.G.; Grillner, S. Neuronal Control of Locomotion: From Mollusc to Man; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Grillner, S. Biological pattern generation: The cellular and computational logic of networks in motion. Neuron 2006, 52, 751–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiehn, O. Locomotor circuits in the mammalian spinal cord. Annu. Rev. Neurosci. 2006, 29, 279–306. [Google Scholar] [CrossRef] [Green Version]
- Talpalar, A.E.; Kiehn, O. Glutamatergic mechanisms for speed control and network operation in the rodent locomotor CpG. Front. Neural Circuits 2010, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Kiehn, O. Development and functional organization of spinal locomotor circuits. Curr. Opin. Neurobiol. 2011, 21, 100–109. [Google Scholar] [CrossRef]
- Kiehn, O. Decoding the organization of spinal circuits that control locomotion. Nat. Rev. Neurosci. 2016, 17, 224–238. [Google Scholar] [CrossRef]
- Kudo, N.; Yamada, T. N-Methyl-d,l-aspartate-induced locomotor activity in a spinal cord-indlimb muscles preparation of the newborn rat studied in vitro. Neurosci. Lett. 1987, 75, 43–48. [Google Scholar] [CrossRef]
- Smith, J.C.; Feldman, J.L. In vitro brainstem-spinal cord preparations for study of motor systems for mammalian respiration and locomotion. J. Neurosci. Methods 1987, 21, 321–333. [Google Scholar] [CrossRef]
- Smith, J.C.; Feldman, J.L.; Schmidt, B.J. Neural mechanisms generating locomotion studied in mammalian brain stem-spinal cord in vitro. FASEB J. 1988, 2, 2283–2288. [Google Scholar] [CrossRef]
- Cazalets, J.R.; Grillner, S.; Menard, I.; Cremieux, J.; Clarac, F. Two types of motor rhythm induced by NMDA and amines in an in vitro spinal cord preparation of neonatal rat. Neurosci. Lett. 1990, 111, 116–121. [Google Scholar] [CrossRef]
- Cazalets, J.R.; Sqalli-Houssaini, Y.; Clarac, F. Activation of the central pattern generators for locomotion by serotonin and excitatory amino acids in neonatal rat. J. Physiol. 1992, 455, 187–204. [Google Scholar] [CrossRef]
- Cazalets, J.R.; Borde, M.; Clarac, F. Localization and organization of the central pattern generator for hindlimb locomotion in newborn rat. J. Neurosci. 1995, 15, 4943–4951. [Google Scholar] [CrossRef] [Green Version]
- Lev-Tov, A.; Delvolve, I.; Kremer, E. Sacrocaudal afferents induce rhythmic efferent bursting in isolated spinal cords of neonatal rats. J. Neurophysiol. 2000, 83, 888–894. [Google Scholar] [CrossRef] [Green Version]
- Whelan, P.; Bonnot, A.; O’Donovan, M.J. Properties of rhythmic activity generated by the isolated spinal cord of the neonatal mouse. J. Neurophysiol. 2000, 84, 2821–2833. [Google Scholar] [CrossRef]
- Zaporozhets, E.; Cowley, K.C.; Schmidt, B.J. A reliable technique for the induction of locomotor-like activity in the in vitro neonatal rat spinal cord using brainstem electrical stimulation. J. Neurosci. Methods 2004, 139, 33–41. [Google Scholar] [CrossRef]
- Zaporozhets, E.; Cowley, K.C.; Schmidt, B.J. Propriospinal neurons contribute to bulbospinal transmission of the locomotor command signal in the neonatal rat spinal cord. J. Physiol. 2006, 572 Pt 2, 443–458. [Google Scholar] [CrossRef]
- Cowley, K.C.; Zaporozhets, E.; Schmidt, B.J. Propriospinal neurons are sufficient for bulbospinal transmission of the locomotor command signal in the neonatal rat spinal cord. J. Physiol. 2008, 586, 1623–1635. [Google Scholar] [CrossRef]
- Pujala, A.; Blivis, D.; O’Donovan, M.J. Interactions between Dorsal and Ventral Root Stimulation on the Generation of Locomotor-Like Activity in the Neonatal Mouse Spinal Cord. eNeuro 2016, 3, e0101-16.2016. [Google Scholar] [CrossRef] [Green Version]
- Caldeira, V.; Dougherty, K.J.; Borgius, L.; Kiehn, O. Spinal Hb9: Cre-derived excitatory interneurons contribute to rhythm generation in the mouse. Sci. Rep. 2017, 7, 41369. [Google Scholar] [CrossRef]
- Jessell, T.M. Neuronal specification in the spinal cord: Inductive signals and transcriptional codes. Nat. Rev. Genet. 2000, 1, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Goulding, M. Circuits controlling vertebrate locomotion: Moving in a new direction. Nat. Rev. Neurosci. 2009, 10, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Kiehn, O.; Dougherty, K. Locomotion: Circuits and Physiology. In Neuroscience in the 21st Century; Springer: New York, NY, USA, 2013; pp. 1209–1236. [Google Scholar]
- Talpalar, A.E.; Bouvier, J.; Borgius, L.; Fortin, G.; Pierani, A.; Kiehn, O. Dual-mode operation of neuronal networks involved in left-right alternation. Nature 2013, 500, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Bellardita, C.; Kiehn, O. Phenotypic characterization of speed-associated gait changes in mice reveals modular organization of locomotor networks. Curr. Biol. 2015, 25, 1426–1436. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Narayan, S.; Geiman, E.; Lanuza, G.M.; Velasquez, T.; Shanks, B.; Akay, T.; Dyck, J.; Pearson, K.; Gosgnach, S.; et al. V3 spinal neurons establish a robust and balanced locomotor rhythm during walking. Neuron 2008, 60, 84–96. [Google Scholar] [CrossRef] [Green Version]
- Borowska, J.; Jones, C.T.; Deska-Gauthier, D.; Zhang, Y. V3 interneuron subpopulations in the mouse spinal cord undergo distinctive postnatal maturation processes. Neuroscience 2015, 295, 221–228. [Google Scholar] [CrossRef]
- Danner, S.M.; Zhang, H.; Shevtsova, N.A.; Borowska-Fielding, J.; Deska-Gauthier, D.; Rybak, I.A.; Zhang, Y. Spinal V3 Interneurons and Left–Right Coordination in Mammalian Locomotion. Front. Cell. Neurosci. 2019, 13, 516. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lanuza, G.M.; Britz, O.; Wang, Z.; Siembab, V.C.; Zhang, Y.; Velasquez, T.; Alvarez, F.J.; Frank, E.; Goulding, M. V1 and V2b interneurons secure the alternating flexor-extensor motor activity mice require for limbed locomotion. Neuron 2014, 82, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Britz, O.; Zhang, J.; Grossmann, K.S.; Dyck, J.; Kim, J.C.; Dymecki, S.; Gosgnach, S.; Goulding, M. A genetically defined asymmetry underlies the inhibitory control of flexor-extensor locomotor movements. Elife 2015, 4, e04718. [Google Scholar] [CrossRef]
- Gosgnach, S.; Lanuza, G.M.; Butt, S.J.; Saueressig, H.; Zhang, Y.; Velasquez, T.; Riethmacher, D.; Callaway, E.M.; Kiehn, O.; Goulding, M. V1 spinal neurons regulate the speed of vertebrate locomotor outputs. Nature 2006, 440, 215–219. [Google Scholar] [CrossRef]
- Falgairolle, M.; O’Donovan, M.J. V1 interneurons regulate the pattern and frequency of locomotor-like activity in the neonatal mouse spinal cord. PLoS Biol. 2019, 17, e3000447. [Google Scholar] [CrossRef]
- Falgairolle, M.; O’Donovan, M.J. Optogenetic Activation of V1 Interneurons Reveals the Multimodality of Spinal Locomotor Networks in the Neonatal Mouse. J. Neurosci. 2021, 41, 8545–8561. [Google Scholar] [CrossRef]
- Crone, S.A.; Quinlan, K.A.; Zagoraiou, L.; Droho, S.; Restrepo, C.E.; Lundfald, L.; Endo, T.; Setlak, J.; Jessell, T.M.; Kiehn, O.; et al. Genetic ablation of V2a ipsilateral interneurons disrupts left-right locomotor coordination in mammalian spinal cord. Neuron 2008, 60, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Crone, S.A.; Zhong, G.; Harris-Warrick, R.; Sharma, K. In mice lacking V2a interneurons, gait depends on speed of locomotion. J. Neurosci. 2009, 29, 7098–7109. [Google Scholar] [CrossRef]
- Kimura, Y.; Higashijima, S.I. Regulation of locomotor speed and selection of active sets of neurons by V1 neurons. Nat. Commun. 2019, 10, 2268. [Google Scholar] [CrossRef] [Green Version]
- Rybak, I.A.; Dougherty, K.J.; Shevtsova, N.A. Organization of the Mammalian Locomotor CPG: Review of Computational Model and Circuit Architectures Based on Genetically Identified Spinal Interneurons(1,2,3). eNeuro 2015, 2, e0069-15.2015. [Google Scholar] [CrossRef]
- Shevtsova, N.A.; Talpalar, A.E.; Markin, S.N.; Harris-Warrick, R.M.; Kiehn, O.; Rybak, I.A. Organization of left-right coordination of neuronal activity in the mammalian spinal cord: Insights from computational modelling. J. Physiol. 2015, 593, 2403–2426. [Google Scholar] [CrossRef]
- Shevtsova, N.A.; Rybak, I.A. Organization of flexor-extensor interactions in the mammalian spinal cord: Insights from computational modelling. J. Physiol. 2016, 594, 6117–6131. [Google Scholar] [CrossRef]
- Danner, S.M.; Wilshin, S.D.; Shevtsova, N.A.; Rybak, I.A. Central control of interlimb coordination and speed-dependent gait expression in quadrupeds. J. Physiol. 2016, 594, 6947–6967. [Google Scholar] [CrossRef] [Green Version]
- Danner, S.M.; Shevtsova, N.A.; Frigon, A.; Rybak, I.A. Computational modeling of spinal circuits controlling limb coordination and gaits in quadrupeds. Elife 2017, 6, e31050. [Google Scholar] [CrossRef]
- Ausborn, J.; Shevtsova, N.A.; Caggiano, V.; Danner, S.M.; Rybak, I.A. Computational modeling of brainstem circuits controlling locomotor frequency and gait. Elife 2019, 8, e43587. [Google Scholar] [CrossRef]
- Hinckley, C.A.; Hartley, R.; Wu, L.; Todd, A.; Ziskind-Conhaim, L. Locomotor-like rhythms in a genetically distinct cluster of interneurons in the mammalian spinal cord. J. Neurophysiol. 2005, 93, 1439–1449. [Google Scholar] [CrossRef] [Green Version]
- Zhong, G.; Shevtsova, N.A.; Rybak, I.A.; Harris-Warrick, R.M. Neuronal activity in the isolated mouse spinal cord during spontaneous deletions in fictive locomotion: Insights into locomotor central pattern generator organization. J. Physiol. 2012, 590, 4735–4759. [Google Scholar] [CrossRef]
- Bikoff, J.B.; Gabitto, M.I.; Rivard, A.F.; Drobac, E.; Machado, T.A.; Miri, A.; Brenner-Morton, S.; Famojure, E.; Diaz, C.; Alvarez, F.J.; et al. Spinal Inhibitory Interneuron Diversity Delineates Variant Motor Microcircuits. Cell 2016, 165, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Bikoff, J.B. Interneuron diversity and function in the spinal motor system. Curr. Opin. Physiol. 2019, 8, 36–43. [Google Scholar] [CrossRef]
- Cowley, K.C.; Schmidt, B.J. Regional distribution of the locomotor pattern-generating network in the neonatal rat spinal cord. J. Neurophysiol. 1997, 77, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Kjaerulff, O.; Kiehn, O. Distribution of networks generating and coordinating locomotor activity in the neonatal rat spinal cord in vitro: A lesion study. J. Neurosci. 1996, 16, 5777–5794. [Google Scholar] [CrossRef] [Green Version]
- Butt, S.J.; Lebret, J.M.; Kiehn, O. Organization of left-right coordination in the mammalian locomotor network. Brain Res. Rev. 2002, 40, 107–117. [Google Scholar] [CrossRef]
- Bannatyne, B.A.; Edgley, S.A.; Hammar, I.; Jankowska, E.; Maxwell, D.J. Networks of inhibitory and excitatory commissural interneurons mediating crossed reticulospinal actions. Eur. J. Neurosci. 2003, 18, 2273–2284. [Google Scholar] [CrossRef]
- Jankowska, E.; Hammar, I.; Slawinska, U.; Maleszak, K.; Edgley, S.A. Neuronal basis of crossed actions from the reticular formation on feline hindlimb motoneurons. J. Neurosci. 2003, 23, 1867–1878. [Google Scholar] [CrossRef] [Green Version]
- Krutki, P.; Jankowska, E.; Edgley, S.A. Are crossed actions of reticulospinal and vestibulospinal neurons on feline motoneurons mediated by the same or separate commissural neurons? J. Neurosci. 2003, 23, 8041–8050. [Google Scholar] [CrossRef] [PubMed]
- Edgley, S.A.; Jankowska, E.; Hammar, I. Ipsilateral actions of feline corticospinal tract neurons on limb motoneurons. J. Neurosci. 2004, 24, 7804–7813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowska, E.; Edgley, S.A.; Krutki, P.; Hammar, I. Functional differentiation and organization of feline midlumbar commissural interneurones. J. Physiol. 2005, 565 Pt 2, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, K.A.; Kiehn, O. Segmental, synaptic actions of commissural interneurons in the mouse spinal cord. J. Neurosci. 2007, 27, 6521–6530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossignol, S.; Dubuc, R.; Gossard, J.P. Dynamic sensorimotor interactions in locomotion. Physiol. Rev. 2006, 86, 89–154. [Google Scholar] [CrossRef] [PubMed]
- Rybak, I.A.; Shevtsova, N.A.; Lafreniere-Roula, M.; McCrea, D.A. Modelling spinal circuitry involved in locomotor pattern generation: Insights from deletions during fictive locomotion. J. Physiol. 2006, 577 Pt 2, 617–639. [Google Scholar] [CrossRef]
- Rybak, I.A.; Stecina, K.; Shevtsova, N.A.; McCrea, D.A. Modelling spinal circuitry involved in locomotor pattern generation: Insights from the effects of afferent stimulation. J. Physiol. 2006, 577 Pt 2, 641–658. [Google Scholar] [CrossRef]
- McCrea, D.A.; Rybak, I.A. Modeling the mammalian locomotor CPG: Insights from mistakes and perturbations. Prog. Brain Res. 2007, 165, 235–253. [Google Scholar]
- Alvarez, F.J.; Fyffe, R.E. The continuing case for the Renshaw cell. J. Physiol. 2007, 584 Pt 1, 31–45. [Google Scholar] [CrossRef]
- McCrea, D.A.; Rybak, I.A. Organization of mammalian locomotor rhythm and pattern generation. Brain Res. Rev. 2008, 57, 134–146. [Google Scholar] [CrossRef] [Green Version]








| Ionic Channels | m∞(V) and h∞(V), V in mV | τm(V) and τh(V), ms |
| Fast sodium, Na | ||
| Persistent sodium, NaP | ||
| Potassium delayed rectifier, K | ||
| hK = 1 |
| Source Population | Target Populations |
|---|---|
| F | i-F * (0.0075, p = 0.1) i-V2b (0.5, p = 0.1) i-V2a (0.3, p = 0.1) i-V0D (0.6, p = 0.1) |
| E | i-E (0.018, p = 0.1) i-V1-1 (0.57, p = 0.1) i-V3 (0.5, p = 0.05) |
| V2b | i-E (−0.3, p = 0.1) |
| V1-1 | i-F (−0.0165, p = 0.1) |
| V1 | i-E (−0.05, p = 0.1) i-Ini (−0.06, p = 0.1) |
| Ini | i-F (−0.005, p = 0.1) |
| Ini1 | i-F (−0.0025, p = 0.1) |
| V2a | i-V0V (0.1, p = 0.1) |
| V0V | c-Ini (0.1, p = 0.1) |
| V0D | c-F (−0.02, p = 0.1) c-V1 (−0.07, p = 0.1) |
| V3 | c-E (0.05, p = 0.05) c-V1-1 (0.3, p = 0.05) |
| CINe | i-V1 (0.14, p = 0.1) i-V1-1 (0.03, p = 0.1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shevtsova, N.A.; Li, E.Z.; Singh, S.; Dougherty, K.J.; Rybak, I.A. Ipsilateral and Contralateral Interactions in Spinal Locomotor Circuits Mediated by V1 Neurons: Insights from Computational Modeling. Int. J. Mol. Sci. 2022, 23, 5541. https://doi.org/10.3390/ijms23105541
Shevtsova NA, Li EZ, Singh S, Dougherty KJ, Rybak IA. Ipsilateral and Contralateral Interactions in Spinal Locomotor Circuits Mediated by V1 Neurons: Insights from Computational Modeling. International Journal of Molecular Sciences. 2022; 23(10):5541. https://doi.org/10.3390/ijms23105541
Chicago/Turabian StyleShevtsova, Natalia A., Erik Z. Li, Shayna Singh, Kimberly J. Dougherty, and Ilya A. Rybak. 2022. "Ipsilateral and Contralateral Interactions in Spinal Locomotor Circuits Mediated by V1 Neurons: Insights from Computational Modeling" International Journal of Molecular Sciences 23, no. 10: 5541. https://doi.org/10.3390/ijms23105541