
Deciphering the Molecular Mechanisms of Chilling Tolerance in Lsi1-Overexpressing Rice

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Validation of Lsi1-Overexpressing Transgenic Dular
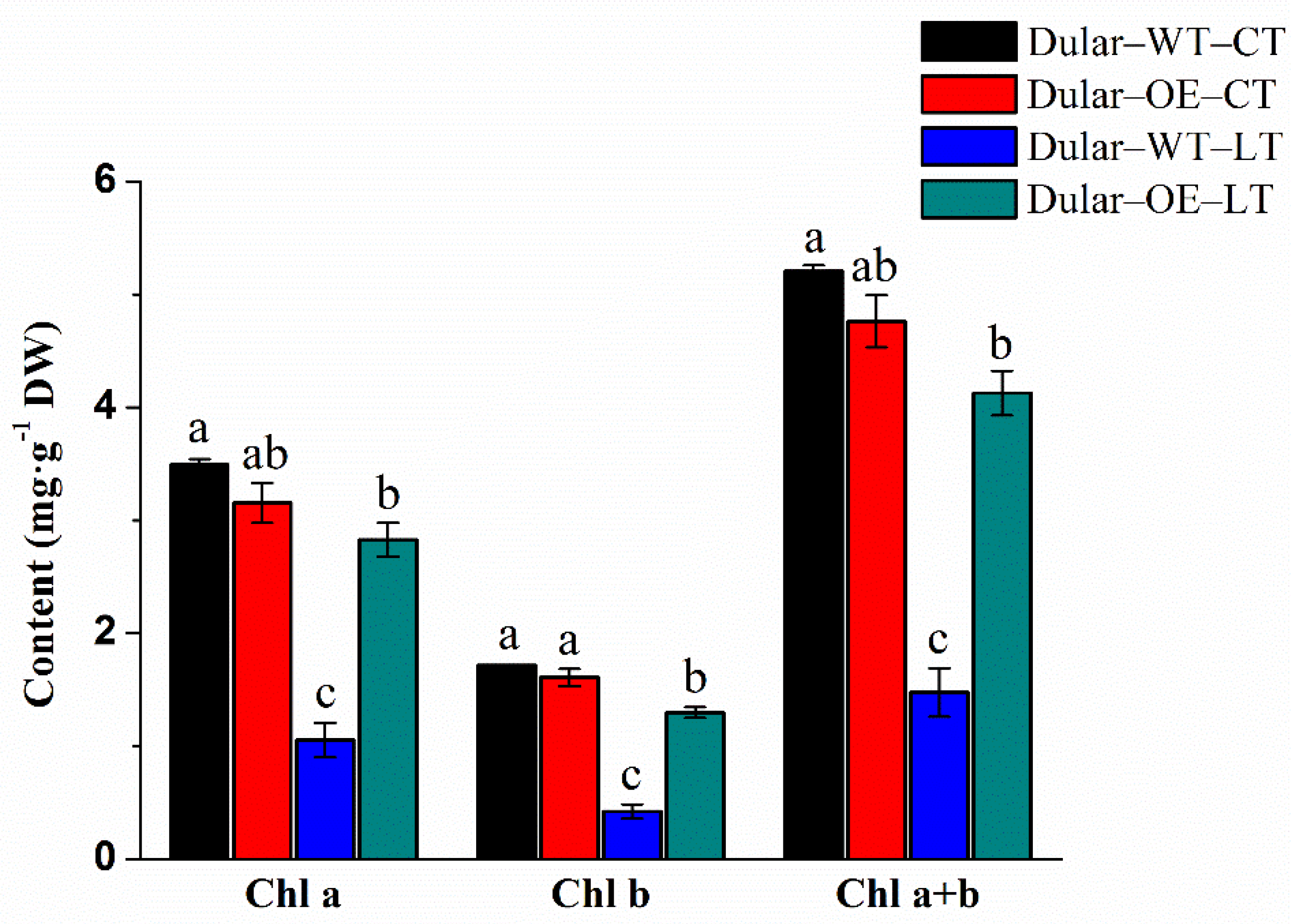
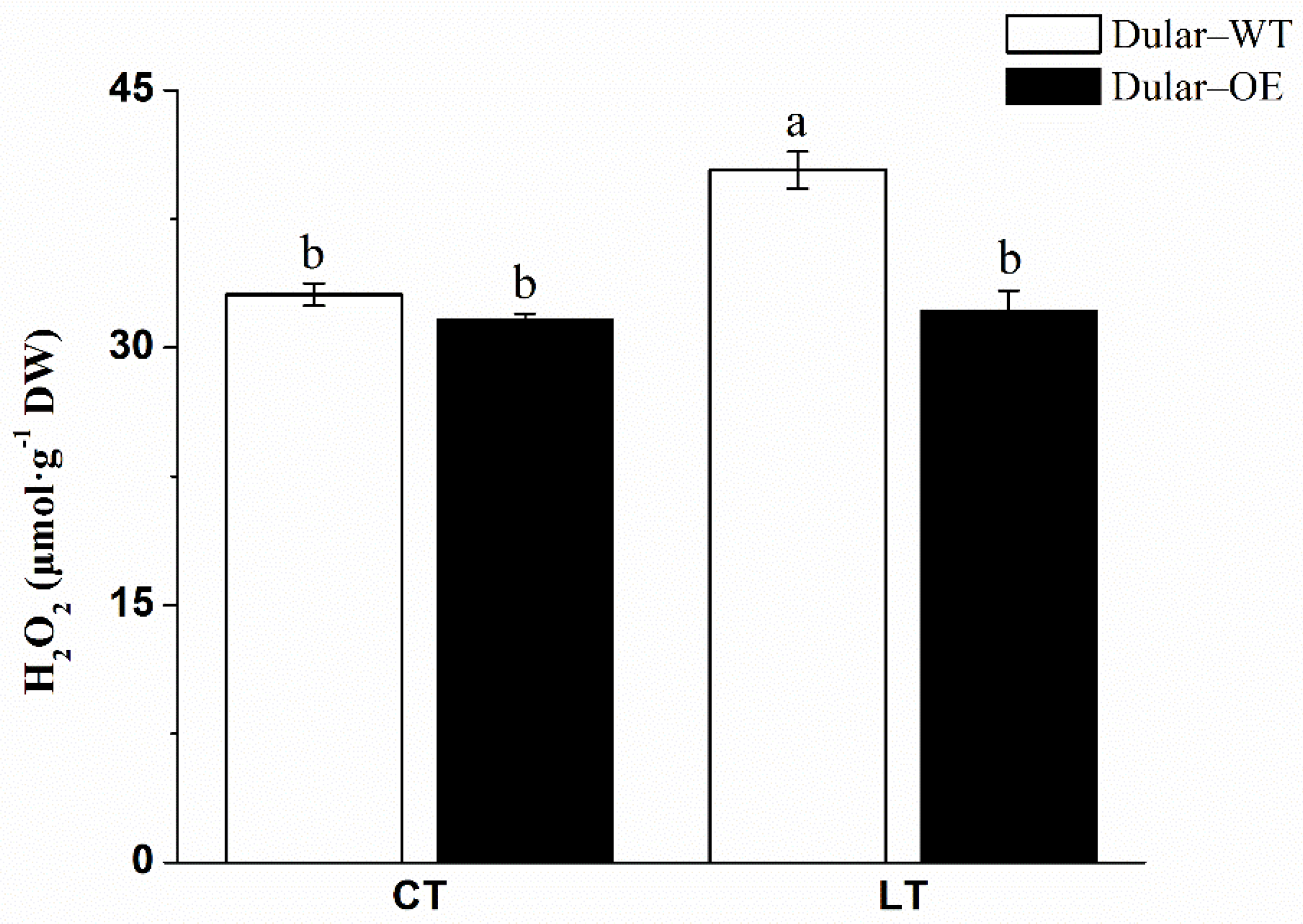
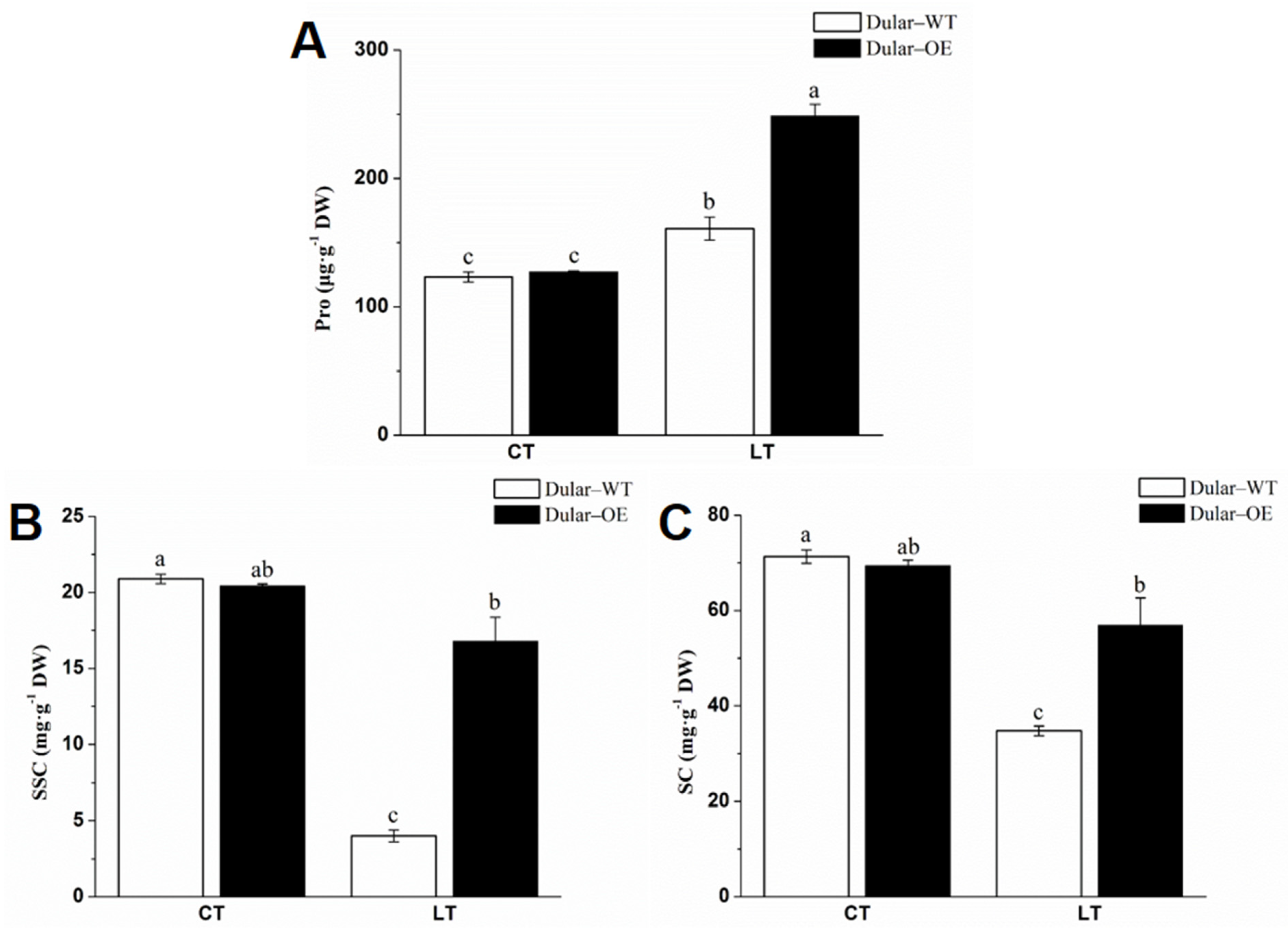
2.2. Cold Response of Dular-WT and Dular-OE
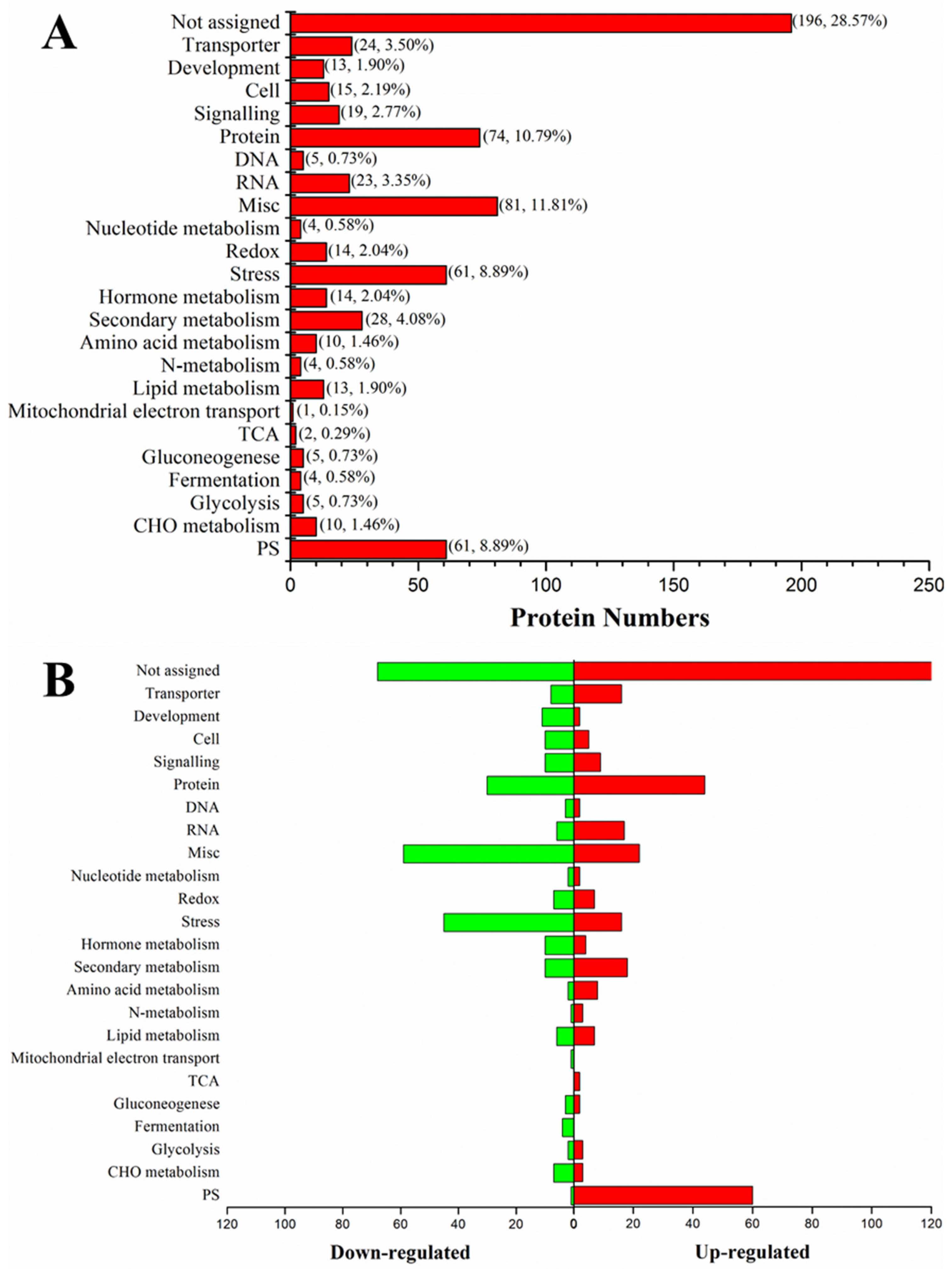
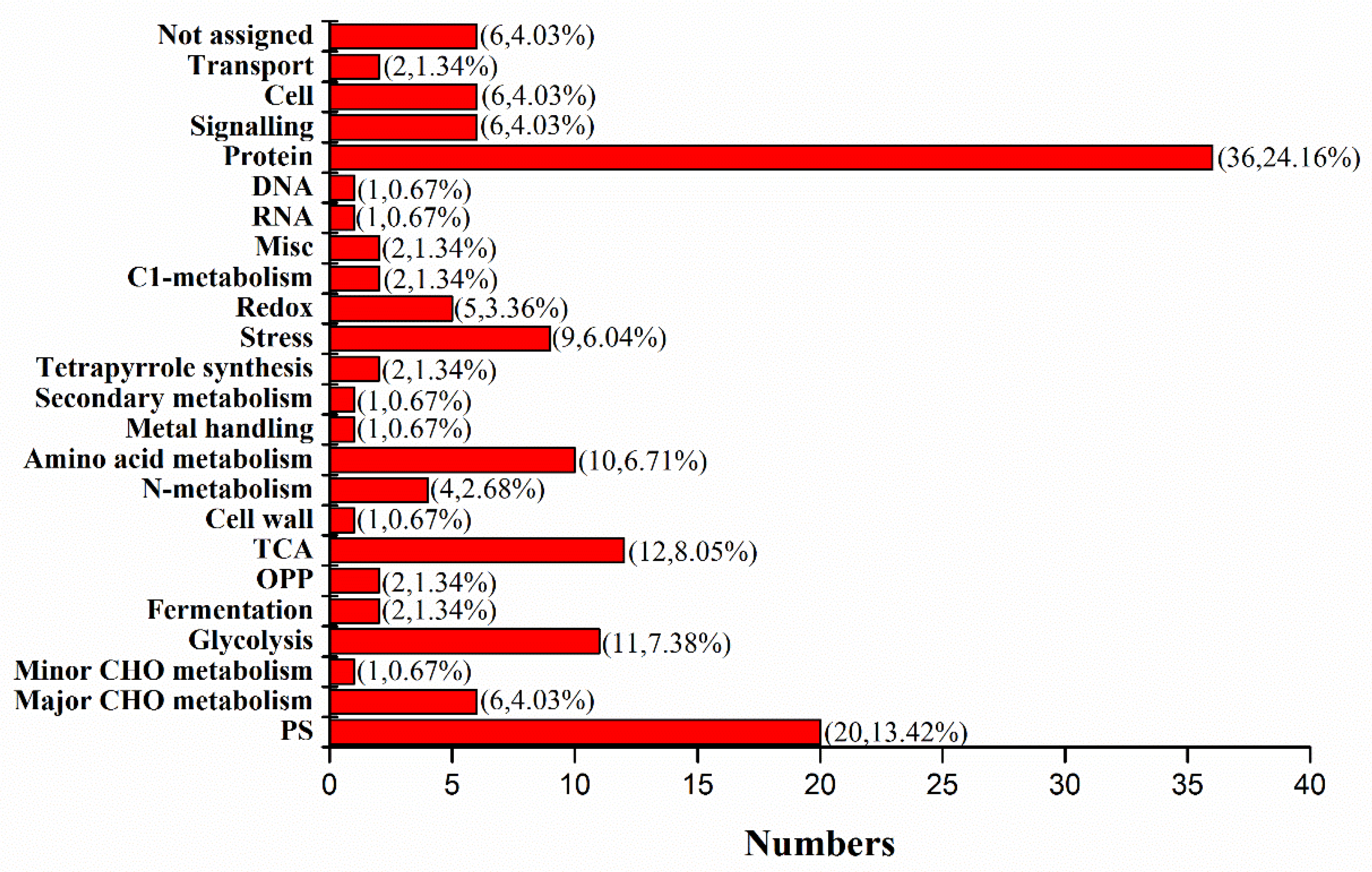
2.3. Differential Protein Expression in Dular-WT and Dular-OE in Response to Low-Temperature Stress
2.4. Effects of Silicon on Rice Growth
2.5. Subcellular Localization of NIP Protein
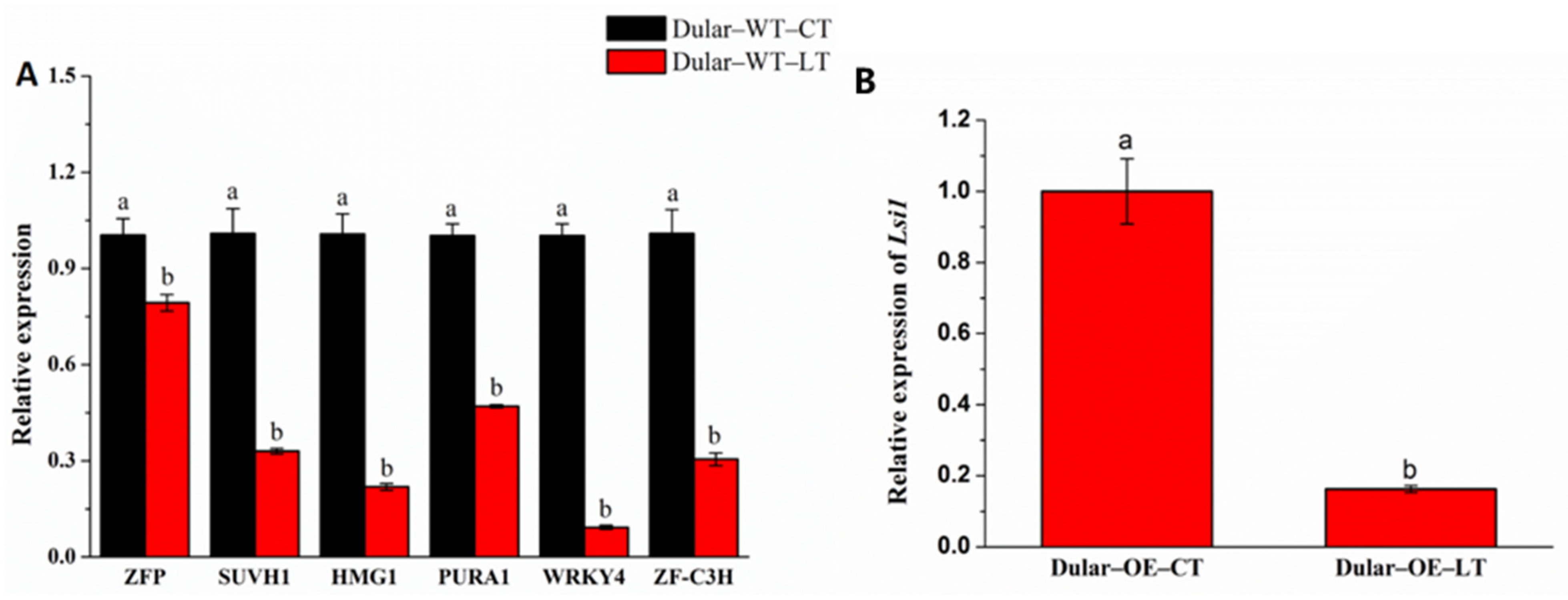
2.6. Identification of Transcription Factors Binding to the Lsi1 Promoter
2.7. Validation of Transcription Factors Binding to the Lsi1 Promoter
2.8. Bioinformatics Screening of Transcription Factors
2.9. Mass-Spectrometry Identification of NIP Interacting Partners
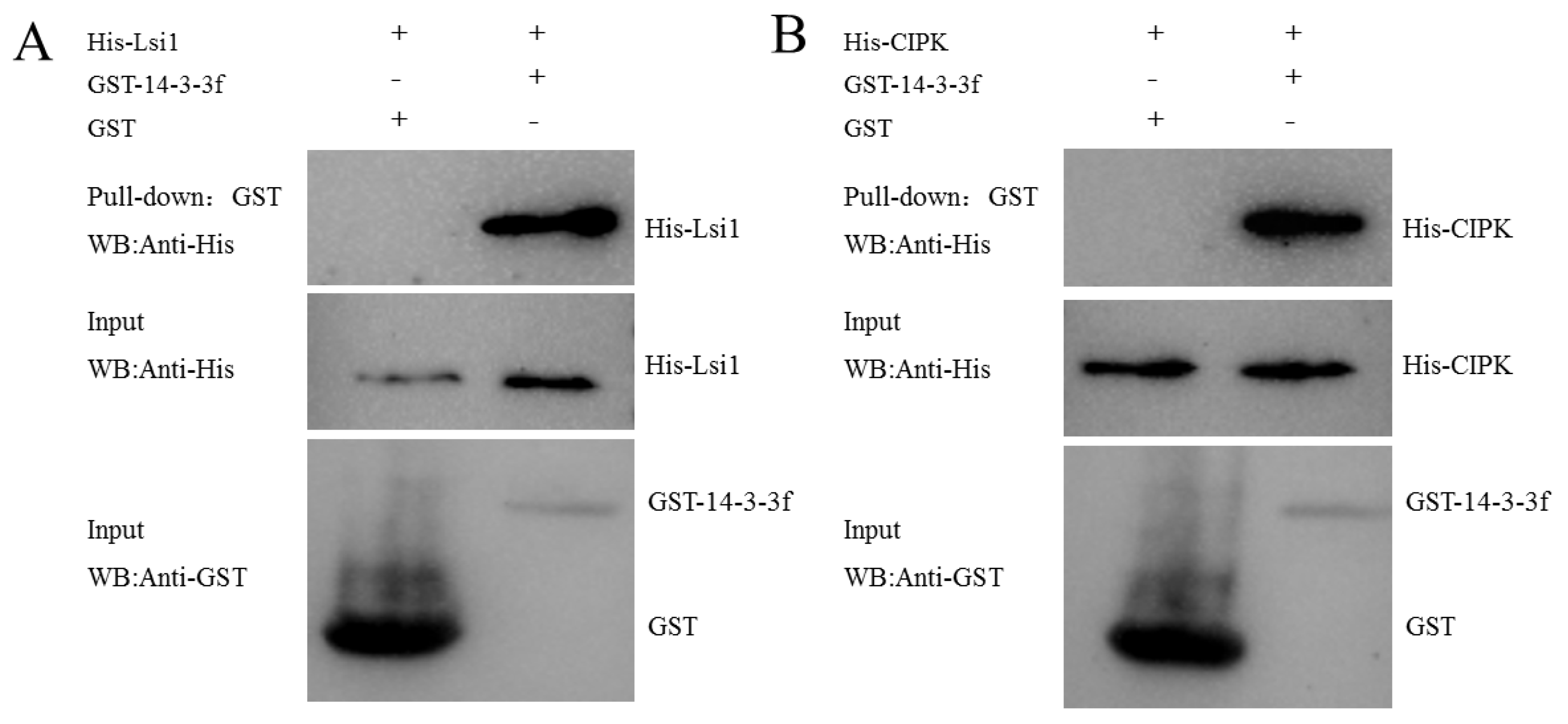
2.10. The Interaction Patterns of NIP Proteins
3. Discussion
3.1. Overexpression of Lsi1 Enhanced Chilling Tolerance in Dular-OE
3.2. Enhanced Silicon Uptake Is Not the Reason for Chilling Tolerance in Dular
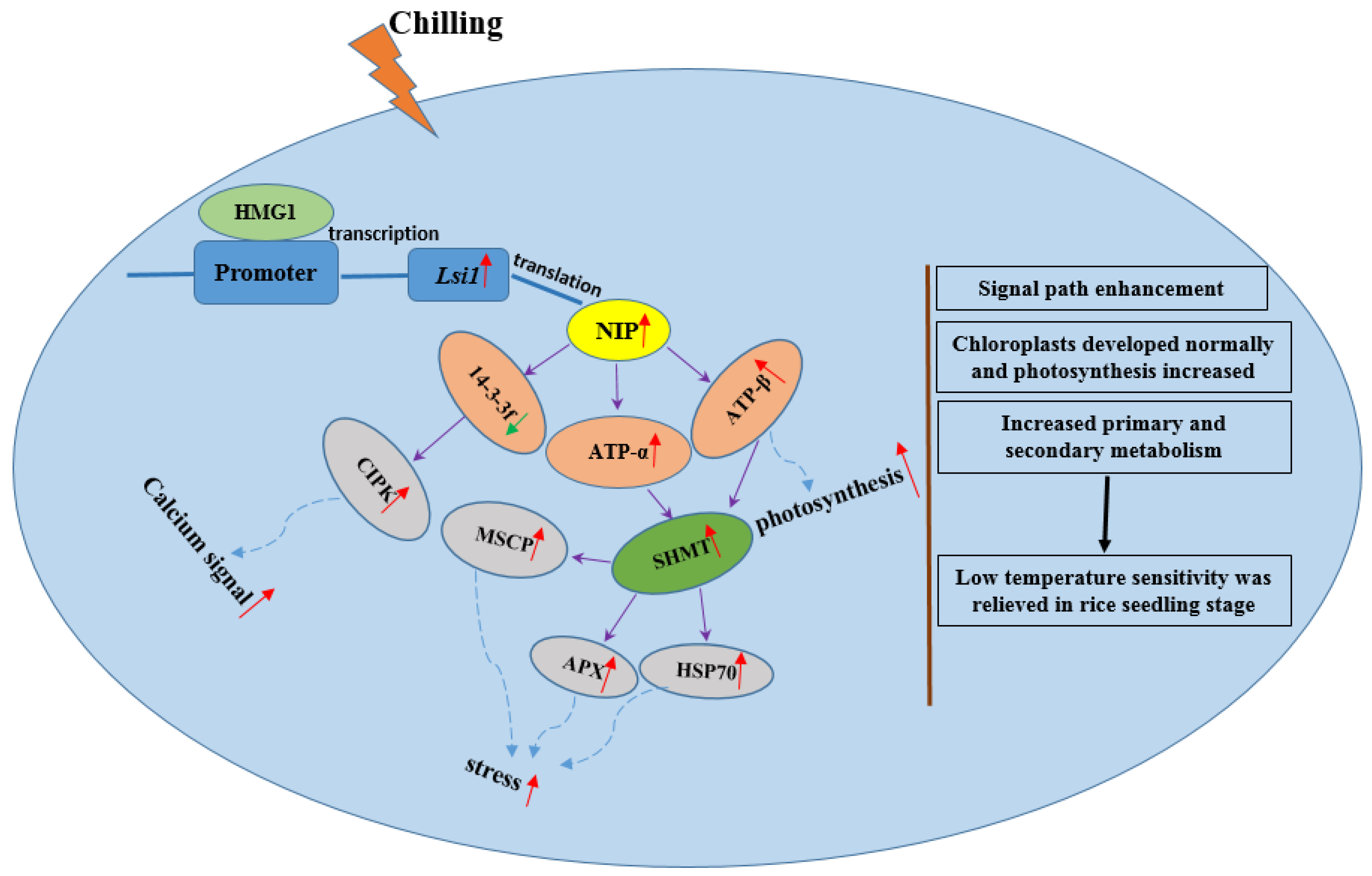
3.3. Molecular Pathways Induced by Lsi1 Overexpression Are the Key to Chilling Tolerance in Transgenic Dular
4. Materials and Methods
4.1. Plant Growth and Treatments
4.2. Protein Extraction and Western Blot Analysis
4.3. Determination of Chlorophyll, H2O2, Enzyme Activity, and Pro and NSC Contents
4.4. Quantitative Proteomics
4.5. Nutrient Solution Experiment
4.6. Subcellular Localization of NIP Protein
4.7. DNA-Pulldown: Transcription Factors Interacting with the Lsi1 Gene Promoter
4.8. Bioinformatics Prediction of Lsi1 Gene-Promoter Transcription-Factor-Binding Sites
4.9. qPCR Validation of Transcription Factors
4.10. NIP-Interacting Proteins
4.11. Bimolecular Fluorescence Complementation to Validate Protein Interactions
4.12. GST Pull-Down Assay
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.; Dong, C.; Zhu, J. Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Curr. Opin. Plant Biol. 2007, 10, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zhang, P.; Jian, X.; Chen, W.; Lin, H.; Li, Y.; Lin, W. Overexpression of Lsi1 in cold-sensitive rice mediates transcriptional regulatory networks and enhances resistance to chilling stress. Plant Sci. 2017, 262, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.; Feng, S.; Amjad, H.; Letuma, P.; Zhan, W.; Li, Z.; Fang, C.; Arafat, Y.; Khan, M.U.; Tayyab, M.; et al. Protein Phosphatase (PP2C9) Induces Protein Expression Differentially to Mediate Nitrogen Utilization Efficiency in Rice under Nitrogen-Deficient Condition. Int. J. Mol. Sci. 2018, 19, 2827. [Google Scholar] [CrossRef] [Green Version]
- Shelton, A.M.; Zhao, J.; Roush, R. Economic, ecological, food safety, and social consequences of the deployment of Bt transgenic plants. Annu. Rev. Entomol. 2002, 47, 845–881. [Google Scholar] [CrossRef]
- Mao, D.; Xin, Y.; Tan, Y.; Hu, X.; Bai, J.; Liu, Z.; Yu, Y.; Li, L.; Peng, C.; Fan, T.; et al. Natural variation in the HAN1 gene confers chilling tolerance in rice and allowed adaptation to a temperate climate. Proc. Natl. Acad. Sci. USA 2019, 116, 3494–3501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, V.; Gupta, A. Overexpression of OsiSAP8, a member of stress associated protein (SAP) gene family of rice confers tolerance to salt, drought and cold stress in transgenic tobacco and rice. Plant Mol. Biol. 2008, 66, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Azeem, S.; Li, Z.; Zheng, H.; Lin, W.; Arafat, Y.; Zhang, Z.; Lin, X.; Lin, W. Quantitative proteomics study on Lsi1 in regulation of rice (Oryza sativa L.) cold resistance. Plant Growth Regul. 2016, 78, 307–323. [Google Scholar] [CrossRef]
- Kuk, Y.; Shin, J.; Burgos, N.; Hwang, T.; Han, O.; Cho, B.; Guh, J. Antioxidative Enzymes Offer Protection from Chilling Damage in Rice Plants. Crop Sci. 2003, 43, 2109–2118. [Google Scholar] [CrossRef]
- Mitani, N.; Yamaji, N.; Ma, J. Identification of maize silicon influx transporters. Plant Cell Physiol. 2009, 50, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Mitani, N.; Nagao, S.; Konishi, S.; Tamai, K.; Iwashita, T.; Yano, M. Characterization of the Silicon Uptake System and Molecular Mapping of the Silicon Transporter Gene in Rice. Plant Physiol. 2004, 136, 3284–3289. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. Silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Wang, Q.; Yu, Y.; Li, Q.; Zhang, H.; Wu, X.; Chen, T.; Lin, W. Suppression and overexpression of Lsi1 induce differential gene expression in rice under ultraviolet radiation. Plant Growth Regul. 2011, 65, 1–10. [Google Scholar] [CrossRef]
- Fang, C.; Zhang, P.; Li, L.; Yang, L.; Mu, D.; Yan, X.; Li, Z.; Lin, W. Serine hydroxymethyltransferase localised in the endoplasmic reticulum plays a role in scavenging H2O2 to enhance rice chilling tolerance. BMC Plant Biol. 2020, 20, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, Y.; Wu, J.; Han, X.; Gu, X.; Lu, T.; Zhang, Z. The RNA editing factor DUA1 is crucial to chloroplast development at low temperature in rice. New Phytol. 2019, 221, 834–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.; Jiang, J.; Miao, L.; He, C. Over-expression of a C3H-type zinc finger gene contributes to salt stress tolerance in transgenic broccoli plants. Plant Cell Tissue Organ Cult. 2017, 130, 239–354. [Google Scholar] [CrossRef]
- Sato, Y.; Masuta, Y.; Saito, K.; Murayama, S.; Ozawa, K. Enhanced chilling tolerance at the booting stage in rice by transgenic overexpression of the ascorbate peroxidase gene, OsAPXa. Plant Cell Rep. 2011, 30, 399–406. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Niogret, M.; Deleu, C.; Bouchereau, A.; Larher, F. The control of proline consumption by abscisic acid during osmotic stress recovery of canola leaf discs. Physiol. Plant. 2003, 117, 213–221. [Google Scholar] [CrossRef]
- Takemiya, A.; Inoue, S.; Doi, M.; Kinoshita, T.; Shimazaki, K. Phototropins promote plant growth in response to blue light in low light environments. Plant Cell 2005, 17, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- De Carbonnel, M.; Davis, P.; Roelfsema, M.; Inoue, S.; Schepens, I.; Lariguet, P.; Geisler, M.; Shimazaki, K.; Hangarter, R.; Fankhauser, C. The Arabidopsis PHYTOCHROME KINASE SUBSTRATE2 Protein Is a Phototropin Signaling Element That Regulates Leaf Flattening and Leaf Positioning. Plant Physiol. 2010, 152, 1391–1405. [Google Scholar] [CrossRef] [Green Version]
- Lariguet, P.; Schepens, I.; Hodgson, D.; Pedmalt, U.; Fankhauser, C. PHYTOCHROME KINASE SUBSTRATE 1 is a phototropin 1 binding protein required for phototropism. Proc. Natl. Acad. Sci. USA 2006, 103, 10134–10139. [Google Scholar] [CrossRef] [Green Version]
- Lintala, M.; Schuck, N.; Thormahlen, I.; Jungfer, A.; Weber, K.; Weber, A.; Geigenberger, P.; Soll, J.; Bölter, B.; Mulo, P. Arabidopsis tic62 trol Mutant Lacking Thylakoid-Bound FerredoxinNADP Oxidoreductase Shows Distinct Metabolic Phenotype. Mol. Plant 2014, 7, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benz, J.; Stengel, A.; Lintala, M.; Lee, Y.; Weber, A.; Philippar, K.; Guegel, I.; Kaieda, S.; Ikegami, T.; Mulo, P.; et al. Arabidopsis Tic62 and Ferredoxin-NADP(H) Oxidoreductase Form Light-Regulated Complexes That Are Integrated into the Chloroplast Redox Poise. Plant Cell 2009, 21, 3965–3983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stengel, A.; Benz, P.; Balsera, M.; Soll, J.; Bolter, B. TIC62 redox-regulated translocon composition and dynamics. J. Biol. Chem. 2008, 283, 6656–6667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, R.; Wang, Z.; Deng, X.; Liu, C.; Yan, B.; Yang, C.; Song, X.; Mo, B.; Cao, X. Ribosomal RNA Biogenesis and Its Response to Chilling Stress in Oryza sativa. Plant Physiol. 2018, 177, 381–397. [Google Scholar] [CrossRef] [Green Version]
- Del Toro, N.; Fernandez-Ruiz, A.; Mignacca, L.; Kalegari, P.; Rowell, M.; Igelmann, S.; Saint-Germain, E.; Benfdil, M.; Lopes-Paciencia, S.; Brakier-Gingras, L.; et al. Ribosomal protein RPL22/eL22 regulates the cell cycle by acting as an inhibitor of the CDK4-cyclin D complex. Cell Cycle 2019, 18, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Hewitson, J.; Shah, K.; Brown, N.; Grevitt, P.; Hain, S.; Newling, K.; Sharp, T.; Kaye, P.; Lagos, D. miR-132 suppresses transcription of ribosomal proteins to promote protective Th1 immunity. EMBO Rep. 2019, 20, 46620–46634. [Google Scholar] [CrossRef]
- Lu, T.; Zhu, Z.; Wu, J.; She, H.; Han, R.; Xu, H.; Qin, Z. DRAM1 regulates autophagy and cell proliferation via inhibition of the phosphoinositide 3-kinase-Akt-mTOR-ribosomal protein S6 pathway. Cell Commun. Signal. 2019, 17, 28–43. [Google Scholar] [CrossRef] [Green Version]
- Wool, I. Extraribosomal functions of ribosomal proteins. Trends Biochem. Sci. 1996, 21, 164–165. [Google Scholar] [CrossRef]
- Cen, W.; Liu, J.; Lu, S.; Jia, P.; Yu, K.; Han, Y.; Li, R.; Luo, J. Comparative proteomic analysis of QTL CTS-12 derived from wild rice (Oryza rufipogon Griff.), in the regulation of cold acclimation and de-acclimation of rice (Oryza sativa L.) in response to severe chilling stress. BMC Plant Biol. 2018, 18, 163–176. [Google Scholar] [CrossRef]
- Kessler, F.; Blobel, G. Interaction of the protein import and folding machineries of the chloroplast. Proc. Natl. Acad. Sci. USA 1996, 93, 7684–7689. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Chen, L.; Yeh, Y.; Hsiao, C.; Li, H. Chloroplast Preproteins Bind to the Dimer Interface of the Toc159 Receptor during Import. Plant Physiol. 2017, 173, 2148–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban, R.; Moran, J.; Becerril, J.; García-Plazaola, J. Versatility of carotenoids: An integrated view on diversity, evolution, functional roles and environmental interactions. Environ. Exp. Bot. 2015, 119, 63–75. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, W.; Pwee, K.; Kumar, P. Cloning and characterization of rice HMGB1 gene. Gene 2003, 312, 103–109. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, W.; Pwee, K.; Kumar, P. Rice HMGB1 protein recognizes DNA structures and bends DNA efficiently. Arch. Biochem. Biophys. 2003, 411, 105–111. [Google Scholar] [CrossRef]
- Kasamo, K.; Yamaguchi, M.; Nakamura, Y. Mechanism of the chilling-induced decrease in proton pumping across the tonoplast of rice cells. Plant Cell Physiol. 2000, 41, 840–849. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhao, H.; Huang, F.; Long, J.; Song, G.; Lin, W. The 14-3-3 protein GF14f negatively affects grain filling of inferior spikelets of rice (Oryza sativa L.). Plant J. 2019, 99, 344–358. [Google Scholar]
- Liu, Z.; Jia, Y.; Ding, Y.; Shi, Y.; Li, Z.; Guo, Y.; Gong, Z.; Yang, S. Plasma Membrane CRPK1-Mediated Phosphorylation of 14-3-3 Proteins Induces Their Nuclear Import to Fine-Tune CBF Signaling during Cold Response. Mol. Cell. 2017, 66, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Xu, S.; Chong, K. Cold Signal Shuttles from Membrane to Nucleus. Mol. Cell. 2017, 66, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Li, P.; Amjad, H.; Khan, A.; Arafat, Y.; Waqas, M.; Li, Z.; Noman, A.; Islam, W.; Wu, L.; et al. Exploring the potential of overexpressed OsCIPK2 rice as a nitrogen utilization efficient crop and analysis of its associated rhizo-compartmental microbial communities. Int. J. Mol. Sci. 2019, 15, 3636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Guo, X.; Xu, Y.; Li, H.; Ma, L. OsCIPK7 point-mutation leads to conformation and kinase-activity change for sensing cold response. J. Integr. Plant Biol. 2019, 61, 1194–1200. [Google Scholar] [CrossRef]
- Ma, Y.; Dai, X.; Xu, Y.; Luo, W.; Zheng, X.; Zeng, D.; Pan, Y.; Lin, X.; Liu, H.; Zhang, D.; et al. COLD1 confers chilling tolerance in rice. Cell 2015, 6, 1209–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Feng, S.; Zhan, W.; Xu, L.; Fang, C.; Zhang, Z.; Lin, W. Lsi1 plays an active role in enhancing the chilling tolerance of rice roots. Plant Growth Regul. 2020, 90, 529–543. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nahakpam, S.; Shah, K. Expression of key antioxidant enzymes under combined effect of heat and cadmium toxicity in growing rice seedlings. Plant Growth Regul. 2011, 63, 23–35. [Google Scholar] [CrossRef]
- Yoshida, M.; Kawakami, A.; Sato, Y. Analyses of cold tolerance and sugar content of rice seedlings expressing wheat fructosyltransferase. Plant Cell Physiol. 2007, 45, 237. [Google Scholar]
- Khare, T.; Srivastava, A.; Suprasanna, P.; Kumar, V. Individual and additive stress impacts of Na+ and Cl− on proline metabolism and nitrosative responses in rice. Plant Physiol. Biochem. 2020, 152, 44–52. [Google Scholar] [CrossRef]
- Thimm, O.; Bläsing, O.; Gibon, Y.; Nagel, A.; Stitt, M. Mapman: A User-Driven Tool To Display Genomics Data Sets Onto Diagrams of Metabolic Pathways and Other Biological Processes. Plant J. 2004, 37, 914–939. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Umar Khan, M.; Yan, X.; Mu, D.; Xie, Y.; Waqas, M.; Wu, X.; Letuma, P.; Fang, C.; Lin, W. Deciphering the Molecular Mechanisms of Chilling Tolerance in Lsi1-Overexpressing Rice. Int. J. Mol. Sci. 2022, 23, 4667. https://doi.org/10.3390/ijms23094667
Li Z, Umar Khan M, Yan X, Mu D, Xie Y, Waqas M, Wu X, Letuma P, Fang C, Lin W. Deciphering the Molecular Mechanisms of Chilling Tolerance in Lsi1-Overexpressing Rice. International Journal of Molecular Sciences. 2022; 23(9):4667. https://doi.org/10.3390/ijms23094667
Chicago/Turabian StyleLi, Zhong, Muhammad Umar Khan, Xue Yan, Dan Mu, Yuebin Xie, Muhammad Waqas, Xin Wu, Puleng Letuma, Changxun Fang, and Wenxiong Lin. 2022. "Deciphering the Molecular Mechanisms of Chilling Tolerance in Lsi1-Overexpressing Rice" International Journal of Molecular Sciences 23, no. 9: 4667. https://doi.org/10.3390/ijms23094667