
Linear and Circular Long Non-Coding RNAs in Acute Lymphoblastic Leukemia: From Pathogenesis to Classification and Treatment

 ,
, 
Abstract
:1. Introduction
2. Current Challenges in ALL Treatment
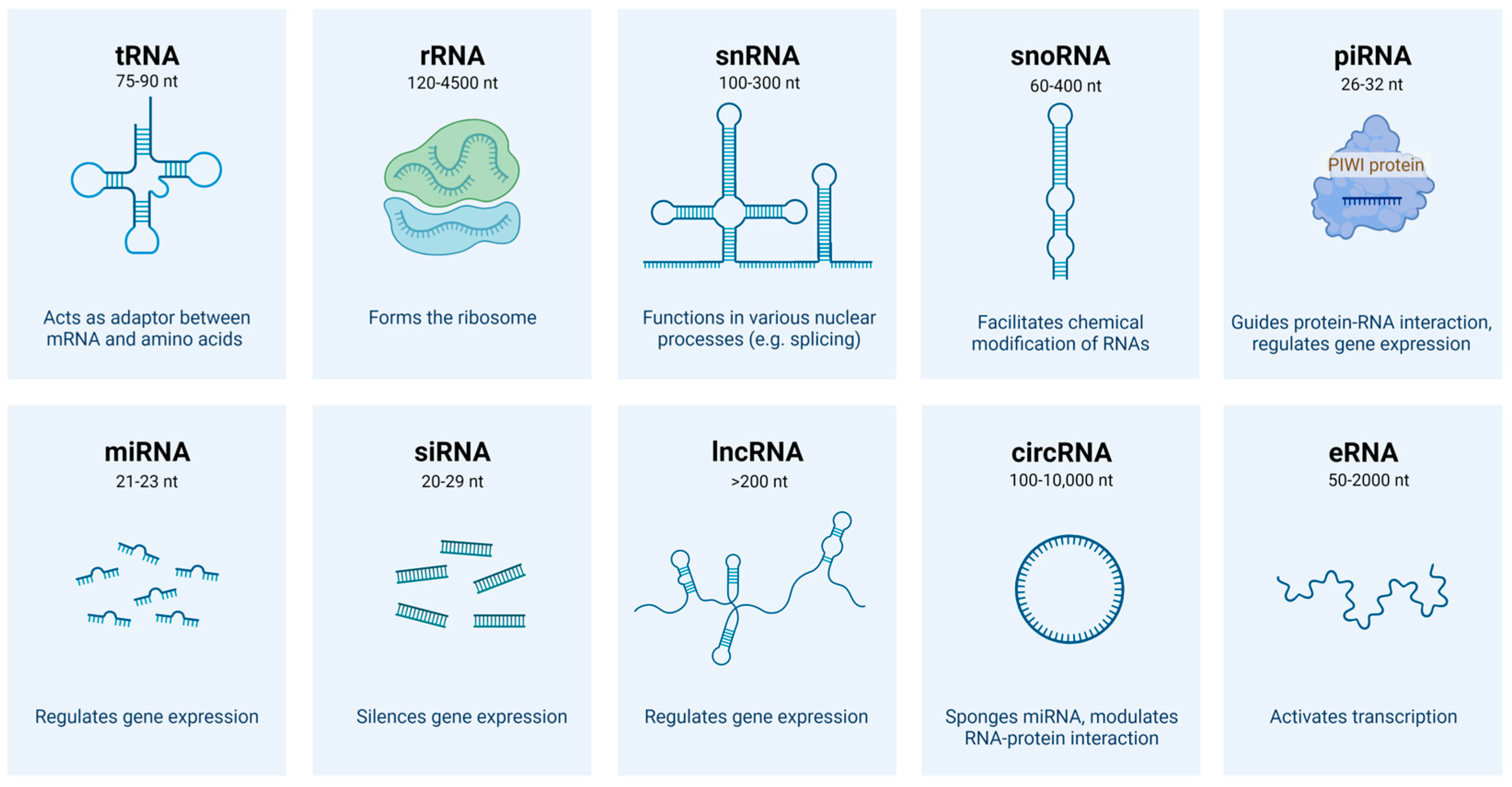
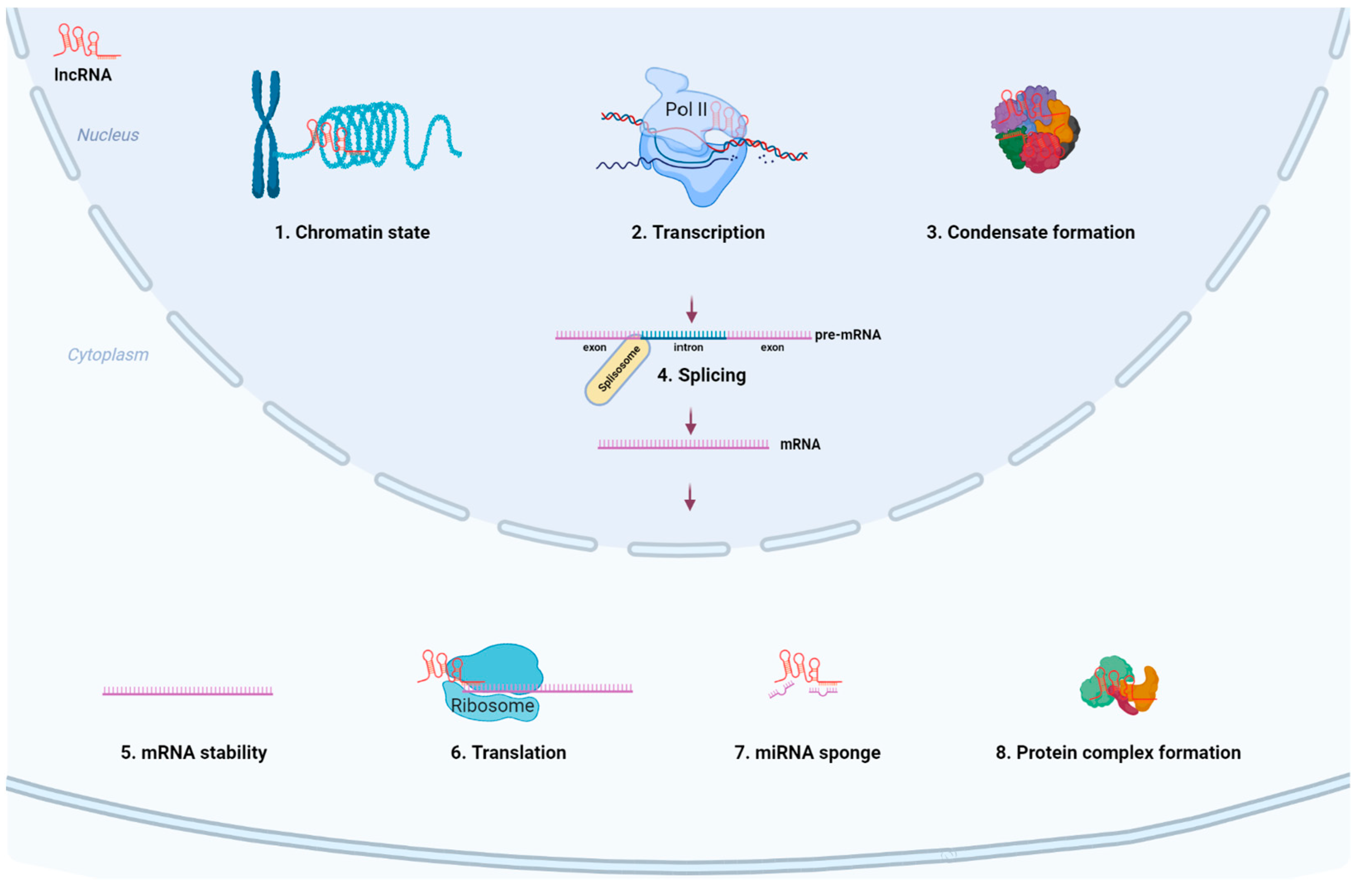
3. Biogenesis and Biological Functions of lncRNAs
4. LncRNAs in ALL Classification, Diagnosis, Pathogenesis, and Resistance to Treatment
4.1. LncRNAs in ALL Classification and Diagnosis
4.2. LncRNAs as Cooperating Events of Genetic Abnormalities in T-ALL Pathogenesis
4.3. LncRNAs as Cooperating Events of Genetic Abnormalities in B-ALL Pathogenesis
4.4. LncRNAs and Immune Response Modulation in ALL Pathogenesis and Treatment
4.5. LncRNAs in Susceptibility, Treatment Response and Prognosis of ALL
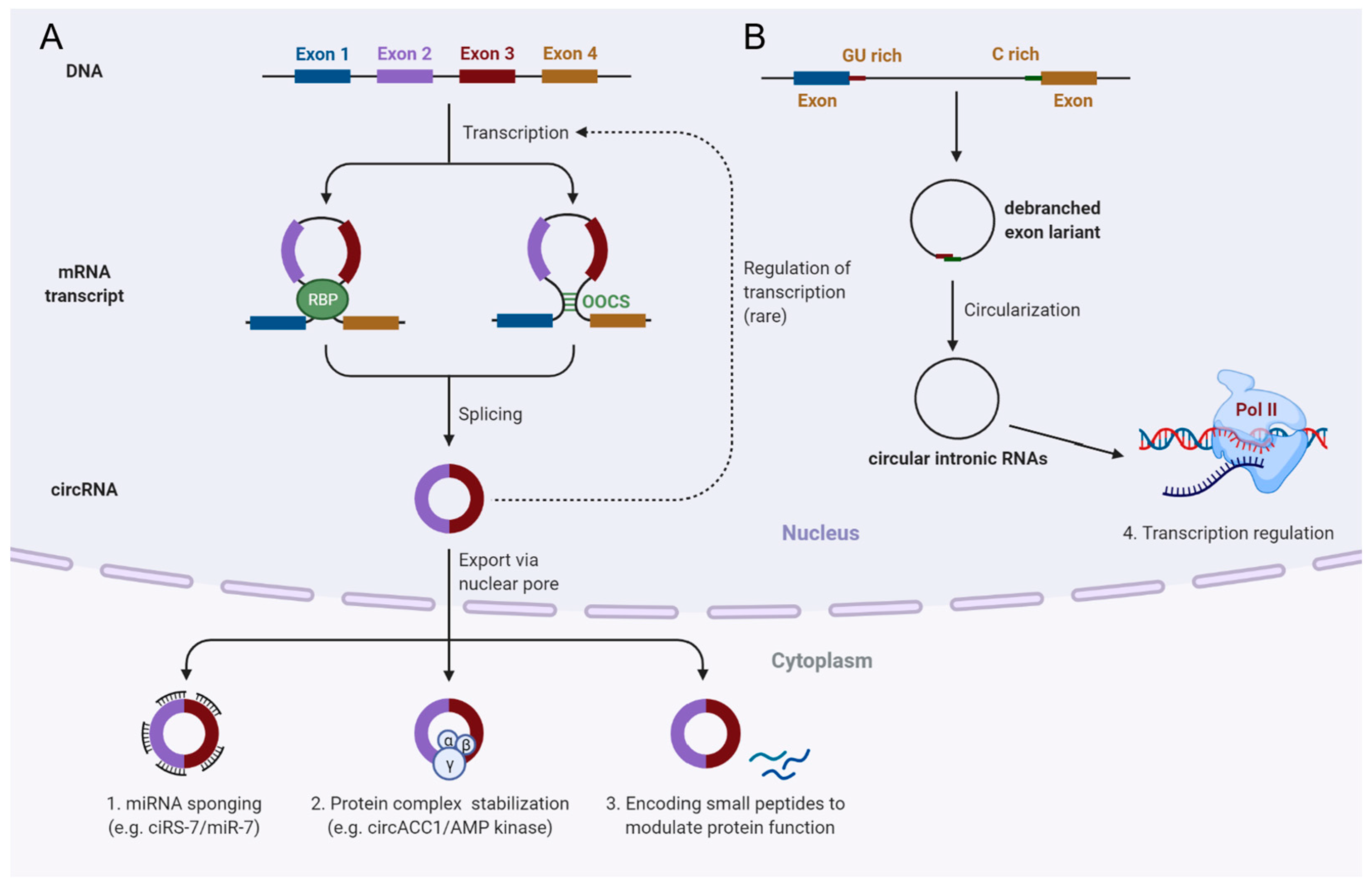
5. Biogenesis and Biological Functions of Circular RNAs
6. CircRNAs in ALL Classification, Pathogenesis and Treatment
6.1. CircRNAs in ALL Classification and Diagnosis
6.2. CircRNAs Acting as Oncogenes in ALL Pathogenesis
6.3. CircRNAs Acting as Tumor Suppressors in ALL Pathogenesis
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ALL | acute lymphoblastic leukemia |
| Allo-SCT | allogenic stem cell transplantation |
| ASOs | antisense oligonucleotides |
| BM | bone marrow |
| CART | chimeric antigen T-cell therapy |
| CNS | central nervous system |
| CR | complete remission |
| circRNA | circular RNA |
| DFS | disease-free survival |
| ENCODE | encyclopedia of DNA elements |
| ETP-ALL | early T-cell precursor ALL |
| GC | glucocorticoid |
| lncRNA | long non-coding RNA |
| MRD | minimal residual disease |
| MLL | mixed lineage leukemia gene |
| ncRNA | non-coding RNA |
| nt | nucleotide |
| OOCS | orientation-opposite complementary sequences |
| ORFs | open reading frames |
| Pol II | RNA polymerase II |
| PRC | polycome repressive complex |
| RBPs | RNA binding proteins |
| RNA-seq | RNA-sequencing |
| SNP | Single nucleotide polymorphism |
| TKIs | tyrosine kinase inhibitors |
| TAL1 | T-cell acute lymphocytic leukemia protein 1 |
| TLX1 | T-Cell Leukemia Homeobox 1 |
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Saw, P.E.; Xu, X.; Chen, J.; Song, E.W. Non-coding RNAs: The new central dogma of cancer biology. Sci. China Life Sci. 2021, 64, 22–50. [Google Scholar] [CrossRef] [PubMed]
- Bindereif, A. Pre-mRNA splicing: From protein-coding to noncoding RNAs. RNA 2015, 21, 571–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [PubMed]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef]
- Malard, F.; Mohty, M. Acute lymphoblastic leukaemia. Lancet 2020, 395, 1146–1162. [Google Scholar] [CrossRef]
- Terwilliger, T.; Abdul-Hay, M. Acute lymphoblastic leukemia: A comprehensive review and 2017 update. Blood Cancer J. 2017, 7, e577. [Google Scholar] [CrossRef] [Green Version]
- van der Valentina Cordo, J.C.; Zwet, K.C.; Rob Pieters, J.P. T-cell Acute Lymphoblastic Leukemia: A Roadmap to Targeted Therapies. Blood Cancer Discov. 2020, 2, 19–31. [Google Scholar] [CrossRef]
- Ratti, S.; Lonetti, A.; Follo, M.Y.; Paganelli, F.; Martelli, A.M.; Chiarini, F.; Evangelisti, C. B-ALL Complexity: Is Targeted Therapy Still A Valuable Approach for Pediatric Patients? Cancers 2020, 12, 3498. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.; Yang, J.J.; Ikawa, T.; Hein, D.; Vicente-Dueñas, C.; Borkhardt, A.; Sánchez-García, I. Cell Fate Decisions: The Role of Transcription Factors in Early B-cell Development and Leukemia. Blood Cancer Discov. 2020, 1, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Van der Zwet, J.C.G.; Cordo, V.; Canté-Barrett, K.; Meijerink, J.P.P. Multi-omic approaches to improve outcome for T-cell acute lymphoblastic leukemia patients. Adv. Biol. Regul. 2019, 74, 100647. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S. Deciphering the Clonal Origin of Relapsed Acute Lymphoblastic Leukemia in Children. Blood Cancer Discov. 2020, 1, 21–22. [Google Scholar] [CrossRef]
- Lee, S.H.R.; Li, Z.; Tai, S.T.; Oh, B.L.Z.; Yeoh, A.E.J. Genetic Alterations in Childhood Acute Lymphoblastic Leukemia: Interactions with Clinical Features and Treatment Response. Cancers 2021, 13, 4068. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Kantarjian, H.; Jabbour, E.J. Adult Acute Lymphoblastic Leukemia. Mayo Clin. Proc. 2016, 91, 1645–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pui, C.H. Precision medicine in acute lymphoblastic leukemia. Front. Med. 2020, 14, 689–700. [Google Scholar] [CrossRef]
- Waanders, E.; Gu, Z.; Dobson, S.M.; Antić, Ž.; Crawford, J.C.; Ma, X.; Edmonson, M.N.; Payne-Turner, D.; van de Vorst, M.; Jongmans, M.; et al. Mutational landscape and patterns of clonal evolution in relapsed pediatric acute lymphoblastic leukemia. Blood Cancer Discov. 2020, 1, 96–111. [Google Scholar] [CrossRef]
- Wiemels, J.L.; Cazzaniga, G.; Daniotti, M.; Eden, O.B.; Addison, G.M.; Masera, G.; Saha, V.; Biondi, A.; Greaves, M.F. Prenatal origin of acute lymphoblastic leukaemia in children. Lancet 1999, 354, 1499–1503. [Google Scholar] [CrossRef]
- Andreasson, P.; Schwaller, J.; Anastasiadou, E.; Aster, J.; Gilliland, D.G. The expression of ETV6/CBFA2 (TEL/AML1) is not sufficient for the transformation of hematopoietic cell lines in vitro or the induction of hematologic disease in vivo. Cancer Genet. Cytogenet. 2001, 130, 93–104. [Google Scholar] [CrossRef]
- Rodriguez, P.D.; Paculova, H.; Kogut, S.; Heath, J.; Schjerven, H.; Frietze, S. Non-Coding RNA Signatures of B-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2021, 22, 2683. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Pan, M.; Zhu, S.; Chao, R.; Wang, L. Emerging roles of microRNAs in acute lymphoblastic leukemia and their clinical prospects. Expert Rev. Hematol. 2021, 14, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Grobbelaar, C.; Ford, A.M. The Role of MicroRNA in Paediatric Acute Lymphoblastic Leukaemia: Challenges for Diagnosis and Therapy. J. Oncol. 2019, 2019, 8941471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malagola, M.; Papayannidis, C.; Baccarani, M. Tyrosine kinase inhibitors in Ph+ acute lymphoblastic leukaemia: Facts and perspectives. Ann. Hematol. 2016, 95, 681–693. [Google Scholar] [CrossRef]
- Hangai, M.; Urayama, K.Y.; Tanaka, J.; Kato, K.; Nishiwaki, S.; Koh, K.; Noguchi, M.; Kato, K.; Yoshida, N.; Sato, M.; et al. Allogeneic Stem Cell Transplantation for Acute Lymphoblastic Leukemia in Adolescents and Young Adults. Biol. Blood Marrow Transplant. 2019, 25, 1597–1602. [Google Scholar] [CrossRef]
- Lato, M.W.; Przysucha, A.; Grosman, S.; Zawitkowska, J.; Lejman, M. The New Therapeutic Strategies in Pediatric T-Cell Acute Lymphoblastic Leukemia. Int. J. Mol. Sci. 2021, 22, 94502. [Google Scholar] [CrossRef]
- Zheng, S.; Gillespie, E.; Naqvi, A.S.; Hayer, K.E.; Ang, Z.; Torres-Diz, M.; Quesnel-Vallières, M.; Hottman, D.A.; Bagashev, A.; Chukinas, J.; et al. Modulation of CD22 Protein Expression in Childhood Leukemia by Pervasive Splicing Aberrations: Implications for CD22-Directed Immunotherapies. Blood Cancer Discov. 2022, 3, 103–115. [Google Scholar] [CrossRef]
- Bartram, J.; Veys, P.; Vora, A. Improvements in outcome of childhood acute lymphoblastic leukaemia (ALL) in the UK—A success story of modern medicine through successive UKALL trials and international collaboration. Br. J. Haematol. 2020, 191, 562–567. [Google Scholar]
- Pulsipher, M.A.; Han, X.; Maude, S.L.; Laetsch, T.W.; Qayed, M.; Rives, S.; Boyer, M.W.; Hiramatsu, H.; Yanik, G.A.; Driscoll, T.; et al. Next-Generation Sequencing of Minimal Residual Disease for Predicting Relapse after Tisagenlecleucel in Children and Young Adults with Acute Lymphoblastic Leukemia. Blood Cancer Discov. 2022, 3, 66–81. [Google Scholar] [CrossRef]
- Ronson, A.; Tvito, A.; Rowe, J.M. Treatment of Relapsed/Refractory Acute Lymphoblastic Leukemia in Adults. Curr. Oncol. Rep. 2016, 18, 39. [Google Scholar] [CrossRef]
- Tottone, L.; Lancho, O.; Loh, J.W.; Singh, A.; Kimura, S.; Roels, J.; Kuchmiy, A.; Strubbe, S.; Lawlor, M.A.; da Silva-Diz, V.; et al. A Tumor Suppressor Enhancer of PTEN in T-cell development and leukemia. Blood Cancer Discov. 2021, 2, 92–109. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.K.; Rowe, J.M.; Buck, G.; Foroni, L.; Gerrard, G.; Litzow, M.R.; Lazarus, H.; Luger, S.M.; Marks, D.I.; McMillan, A.K.; et al. UKALLXII/ECOG2993: Addition of imatinib to a standard treatment regimen enhances long-term outcomes in Philadelphia positive acute lymphoblastic leukemia. Blood 2014, 123, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Foà, R.; Bassan, R.; Vitale, A.; Elia, L.; Piciocchi, A.; Puzzolo, M.C.; Canichella, M.; Viero, P.; Ferrara, F.; Lunghi, M.; et al. Dasatinib-Blinatumomab for Ph-Positive Acute Lymphoblastic Leukemia in Adults. N. Engl. J. Med. 2020, 383, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Maimaitiyiming, Y.; Wang, Q.Q.; Yang, C.; Ogra, Y.; Lou, Y.; Smith, C.A.; Hussain, L.; Shao, Y.M.; Lin, J.; Liu, J.; et al. Hyperthermia Selectively Destabilizes Oncogenic Fusion Proteins. Blood Cancer Discov. 2021, 2, 388–401. [Google Scholar] [CrossRef]
- Jin, M.; An, Q.; Xu, S. Central nervous system disease in childhood acute lymphoblastic leukemia. Minerva Pediatr. 2019, 71, 470–474. [Google Scholar] [CrossRef]
- Simone, J.V. Preventive central-nervous-system therapy in acute leukemia. N. Engl. J. Med. 1973, 289, 1248–1249. [Google Scholar] [CrossRef]
- Pui, C.H.; Howard, S.C. Current management and challenges of malignant disease in the CNS in paediatric leukaemia. Lancet Oncol. 2008, 9, 257–268. [Google Scholar] [CrossRef]
- Frishman-Levy, L.; Izraeli, S. Advances in understanding the pathogenesis of CNS acute lymphoblastic leukaemia and potential for therapy. Br. J. Haematol. 2017, 176, 157–167. [Google Scholar] [CrossRef]
- Bhojwani, D.; Pui, C.H. Relapsed childhood acute lymphoblastic leukaemia. Lancet Oncol. 2013, 14, e205–e217. [Google Scholar] [CrossRef]
- Barredo, J.C.; Devidas, M.; Lauer, S.J.; Billett, A.; Marymont, M.; Pullen, J.; Camitta, B.; Winick, N.; Carroll, W.; Ritchey, A.K. Isolated CNS relapse of acute lymphoblastic leukemia treated with intensive systemic chemotherapy and delayed CNS radiation: A pediatric oncology group study. J. Clin. Oncol. 2006, 24, 3142–3149. [Google Scholar] [CrossRef]
- Lenk, L.; Alsadeq, A.; Schewe, D.M. Involvement of the central nervous system in acute lymphoblastic leukemia: Opinions on molecular mechanisms and clinical implications based on recent data. Cancer Metastasis Rev. 2020, 39, 173–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segers, H.; Cools, J. Genomics, Transcriptomics, and Minimal Residual Disease Detection: The Winning Team to Guide Treatment of Acute Lymphoblastic Leukemia. Blood Cancer Discov. 2021, 2, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Vanner, R.J.; Dobson, S.M.; Gan, O.I.; McLeod, J.; Schoof, E.M.; Grandal, I.; Wintersinger, J.A.; Garcia-Prat, L.; Hosseini, M.; Xie, S.Z.; et al. Multiomic Profiling of Central Nervous System Leukemia Identifies mRNA Translation as a Therapeutic Target. Blood Cancer Discov. 2022, 3, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Yang, L.; Chen, L.L. The Diversity of Long Noncoding RNAs and Their Generation. Trends Genet. 2017, 33, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Lu, T.; Tan, J.; Li, H.; Wang, Q.; Wei, L. Long Non-coding RNAs as Communicators and Mediators Between the Tumor Microenvironment and Cancer Cells. Front. Oncol. 2019, 9, 739. [Google Scholar] [CrossRef] [PubMed]
- Marahrens, Y.; Panning, B.; Dausman, J.; Strauss, W.; Jaenisch, R. Xist-deficient mice are defective in dosage compensation but not spermatogenesis. Genes Dev. 1997, 11, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.H.; Yao, R.W.; Zhang, Y.; Guo, C.J.; Jiang, S.; Xu, G.; Dong, R.; Yang, L.; Chen, L.L. SLERT Regulates DDX21 Rings Associated with Pol I Transcription. Cell 2017, 169, 664–678.e16. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Freier, S.M.; Spector, D.L. 3′ end processing of a long nuclear-retained noncoding RNA yields a tRNA-like cytoplasmic RNA. Cell 2008, 135, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.F.; Yang, L.; Zhang, Y.; Xiang, J.F.; Wu, Y.W.; Carmichael, G.G.; Chen, L.L. Long noncoding RNAs with snoRNA ends. Mol. Cell 2012, 48, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Yin, Q.F.; Luo, Z.; Yao, R.W.; Zheng, C.C.; Zhang, J.; Xiang, J.F.; Yang, L.; Chen, L.L. Unusual Processing Generates SPA LncRNAs that Sequester Multiple RNA Binding Proteins. Mol. Cell 2016, 64, 534–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holdt, L.M.; Hoffmann, S.; Sass, K.; Langenberger, D.; Scholz, M.; Krohn, K.; Finstermeier, K.; Stahringer, A.; Wilfert, W.; Beutner, F.; et al. Alu elements in ANRIL non-coding RNA at chromosome 9p21 modulate atherogenic cell functions through trans-regulation of gene networks. PLoS Genet. 2013, 9, e1003588. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, Y.; Lee, J.T. YY1 tethers Xist RNA to the inactive X nucleation center. Cell 2011, 146, 119–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHugh, C.A.; Chen, C.K.; Chow, A.; Surka, C.F.; Tran, C.; McDonel, P.; Pandya-Jones, A.; Blanco, M.; Burghard, C.; Moradian, A.; et al. The Xist lncRNA interacts directly with SHARP to silence transcription through HDAC3. Nature 2015, 521, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Colognori, D.; Sunwoo, H.; Kriz, A.J.; Wang, C.Y.; Lee, J.T. Xist Deletional Analysis Reveals an Interdependency between Xist RNA and Polycomb Complexes for Spreading along the Inactive, X. Mol. Cell 2019, 74, 101–117.e10. [Google Scholar] [CrossRef] [Green Version]
- Schlackow, M.; Nojima, T.; Gomes, T.; Dhir, A.; Carmo-Fonseca, M.; Proudfoot, N.J. Distinctive Patterns of Transcription and RNA Processing for Human lincRNAs. Mol. Cell 2017, 65, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Zuckerman, B.; Ulitsky, I. Predictive models of subcellular localization of long RNAs. RNA 2019, 25, 557–572. [Google Scholar] [CrossRef]
- Yamazaki, T.; Souquere, S.; Chujo, T.; Kobelke, S.; Chong, Y.S.; Fox, A.H.; Bond, C.S.; Nakagawa, S.; Pierron, G.; Hirose, T. Functional Domains of NEAT1 Architectural lncRNA Induce Paraspeckle Assembly through Phase Separation. Mol. Cell 2018, 70, 1038–1053.e7. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, J.N.; Ensminger, A.W.; Clemson, C.M.; Lynch, C.R.; Lawrence, J.B.; Chess, A. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with SC35 splicing domains. BMC Genom. 2007, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Engreitz, J.M.; Sirokman, K.; McDonel, P.; Shishkin, A.A.; Surka, C.; Russell, P.; Grossman, S.R.; Chow, A.Y.; Guttman, M.; Lander, E.S. RNA-RNA interactions enable specific targeting of noncoding RNAs to nascent Pre-mRNAs and chromatin sites. Cell 2014, 159, 188–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maharana, S.; Wang, J.; Papadopoulos, D.K.; Richter, D.; Pozniakovsky, A.; Poser, I.; Bickle, M.; Rizk, S.; Guillén-Boixet, J.; Franzmann, T.M.; et al. RNA buffers the phase separation behavior of prion-like RNA binding proteins. Science 2018, 360, 918–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Maimaitiyiming, Y.; Wang, L.; Cheng, X.; Hsu, C.H. Modulation of Phase Separation by RNA: A Glimpse on N6-Methyladenosine Modification. Front. Cell Dev. Biol. 2021, 9, 786454. [Google Scholar] [CrossRef] [PubMed]
- Lagier-Tourenne, C.; Polymenidou, M.; Cleveland, D.W. TDP-43 and FUS/TLS: Emerging roles in RNA processing and neurodegeneration. Hum. Mol. Genet. 2010, 19, R46–R64. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, B.; Ron, M.; Mikl, M.; Segal, E.; Ulitsky, I. Gene Architecture and Sequence Composition Underpin Selective Dependency of Nuclear Export of Long RNAs on NXF1 and the TREX Complex. Mol. Cell 2020, 79, 251–267.e6. [Google Scholar] [CrossRef]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Nandwani, A.; Rathore, S.; Datta, M. LncRNAs in cancer: Regulatory and therapeutic implications. Cancer Lett. 2021, 501, 162–171. [Google Scholar] [CrossRef]
- Melixetian, M.; Pelicci, P.G.; Lanfrancone, L. Regulation of LncRNAs in Melanoma and Their Functional Roles in the Metastatic Process. Cells 2022, 11, 577. [Google Scholar] [CrossRef]
- Nasrollahzadeh-Khakiani, M.; Emadi-Baygi, M.; Schulz, W.A.; Nikpour, P. Long noncoding RNAs in gastric cancer carcinogenesis and metastasis. Brief. Funct. Genom. 2017, 16, 129–145. [Google Scholar] [CrossRef]
- Liu, H.; Wan, J.; Chu, J. Long non-coding RNAs and endometrial cancer. Biomed. Pharmacother. 2019, 119, 109396. [Google Scholar] [CrossRef]
- Lee, J.S.; Mendell, J.T. Antisense-Mediated Transcript Knockdown Triggers Premature Transcription Termination. Mol. Cell 2020, 77, 1044–1054.e3. [Google Scholar] [CrossRef] [PubMed]
- Stojic, L.; Niemczyk, M.; Orjalo, A.; Ito, Y.; Ruijter, A.E.; Uribe-Lewis, S.; Joseph, N.; Weston, S.; Menon, S.; Odom, D.T.; et al. Transcriptional silencing of long noncoding RNA GNG12-AS1 uncouples its transcriptional and product-related functions. Nat. Commun. 2016, 7, 10406. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lian, Z.; Padden, C.; Gerstein, M.B.; Rozowsky, J.; Snyder, M.; Gingeras, T.R.; Kapranov, P.; Weissman, S.M.; Newburger, P.E. A myelopoiesis-associated regulatory intergenic noncoding RNA transcript within the human HOXA cluster. Blood 2009, 113, 2526–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ørom, U.A.; Derrien, T.; Beringer, M.; Gumireddy, K.; Gardini, A.; Bussotti, G.; Lai, F.; Zytnicki, M.; Notredame, C.; Huang, Q.; et al. Long noncoding RNAs with enhancer-like function in human cells. Cell 2010, 143, 46–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, F.; Dong, F.; Yang, Y.; Huang, A.; Chen, S.; Sun, D.; Xiong, S.; Zhang, J. Dynamic transcription of long non-coding RNA genes during CD4+ T cell development and activation. PLoS ONE 2014, 9, e101588. [Google Scholar] [CrossRef] [Green Version]
- Brazão, T.F.; Johnson, J.S.; Müller, J.; Heger, A.; Ponting, C.P.; Tybulewicz, V.L. Long noncoding RNAs in B-cell development and activation. Blood 2016, 128, e10–e19. [Google Scholar] [CrossRef] [Green Version]
- Trimarchi, T.; Bilal, E.; Ntziachristos, P.; Fabbri, G.; Dalla-Favera, R.; Tsirigos, A.; Aifantis, I. Genome-wide mapping and characterization of Notch-regulated long noncoding RNAs in acute leukemia. Cell 2014, 158, 593–606. [Google Scholar] [CrossRef] [Green Version]
- El-Khazragy, N.; Abdel Aziz, M.A.; Hesham, M.; Matbouly, S.; Mostafa, S.A.; Bakkar, A.; Abouelnile, M.; Noufal, Y.; Mahran, N.A.; Elkhalek, M.A.; et al. Upregulation of leukemia-induced non-coding activator RNA (LUNAR1) predicts poor outcome in pediatric T-acute lymphoblastic leukemia. Immunobiology 2021, 226, 152149. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, H.G.; Lu, C. A novel long non-coding RNA T-ALL-R-LncR1 knockdown and Par-4 cooperate to induce cellular apoptosis in T-cell acute lymphoblastic leukemia cells. Leuk. Lymphoma 2014, 55, 1373–1382. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, P.; Lin, R.; Rong, L.; Xue, Y.; Fang, Y. LncRNA NALT interaction with NOTCH1 promoted cell proliferation in pediatric T cell acute lymphoblastic leukemia. Sci. Rep. 2015, 5, 13749. [Google Scholar] [CrossRef] [Green Version]
- Renou, L.; Boelle, P.Y.; Deswarte, C.; Spicuglia, S.; Benyoucef, A.; Calvo, J.; Uzan, B.; Belhocine, M.; Cieslak, A.; Landman-Parker, J.; et al. Homeobox protein TLX3 activates miR-125b expression to promote T-cell acute lymphoblastic leukemia. Blood Adv. 2017, 1, 733–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.H.; Leong, W.Z.; Ngoc, P.C.T.; Tan, T.K.; Bertulfo, F.C.; Lim, M.C.; An, O.; Li, Z.; Yeoh, A.; Fullwood, M.J.; et al. The enhancer RNA ARIEL activates the oncogenic transcriptional program in T-cell acute lymphoblastic leukemia. Blood 2019, 134, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shi, Y.; Li, J.; Yang, X.; Li, R.; Zhou, X.; Zhu, L. LncRNA CDKN2B-AS1 contributes to tumorigenesis and chemoresistance in pediatric T-cell acute lymphoblastic leukemia through miR-335-3p/TRAF5 axis. Anticancer Drugs 2020. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Guo, W.; Geng, H.; Wang, C.; Yang, S.; Xu, X. LINC00511 exacerbated T-cell acute lymphoblastic leukemia via miR-195-5p/LRRK1 axis. Biosci. Rep. 2020, 40, BSR20193631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Gao, L.; Zhao, J.; Liu, D.; Li, H.; Hu, M. LncRNA ANRIL/miR-7-5p/TCF4 axis contributes to the progression of T cell acute lymphoblastic leukemia. Cancer Cell Int. 2020, 20, 335. [Google Scholar] [CrossRef]
- Li, X.; Song, F.; Sun, H. Long non-coding RNA AWPPH interacts with ROCK2 and regulates the proliferation and apoptosis of cancer cells in pediatric T-cell acute lymphoblastic leukemia. Oncol. Lett. 2020, 20, 239. [Google Scholar] [CrossRef]
- Luo, Y.Y.; Wang, Z.H.; Yu, Q.; Yuan, L.L.; Peng, H.L.; Xu, Y.X. LncRNA-NEAT1 promotes proliferation of T-ALL cells via miR-146b-5p/NOTCH1 signaling pathway. Pathol. Res. Pract. 2020, 216, 153212. [Google Scholar] [CrossRef]
- Singh, N.; Padi, S.K.R.; Bearss, J.J.; Pandey, R.; Okumura, K.; Beltran, H.; Song, J.H.; Kraft, A.S.; Olive, V. PIM protein kinases regulate the level of the long noncoding RNA H19 to control stem cell gene transcription and modulate tumor growth. Mol. Oncol. 2020, 14, 974–990. [Google Scholar] [CrossRef] [Green Version]
- Mofidi, M.; Rahgozar, S.; Pouyanrad, S. Increased level of long non coding RNA H19 is correlated with the downregulation of miR-326 and BCL-2 genes in pediatric acute lymphoblastic leukemia, a possible hallmark for leukemogenesis. Mol. Biol. Rep. 2021, 48, 1531–1538. [Google Scholar] [CrossRef]
- Norouzi, A.; Motaghi, M.; Hassanshahi, G.; Nazari-Robati, M. Exploring the expression profile of vitamin D receptor and its related long non-coding RNAs in patients with acute lymphoblastic leukemia. Rev. Assoc. Med. Bras. 2021, 67, 1113–1117. [Google Scholar] [CrossRef]
- Li, J.; Muhammad, J.; Xie, T.; Sun, J.; Lei, Y.; Wei, Z.; Pan, S.; Qin, H.; Shao, L.; Jiang, D.; et al. LINC00853 restrains T cell acute lymphoblastic leukemia invasion and infiltration by regulating CCR9/CCL25. Mol. Immunol. 2021, 140, 267–275. [Google Scholar] [CrossRef]
- Xiao, S.; Xu, N.; Ding, Q.; Huang, S.; Zha, Y.; Zhu, H. LncRNA VPS9D1-AS1 promotes cell proliferation in acute lymphoblastic leukemia through modulating GPX1 expression by miR-491-5p and miR-214-3p evasion. Biosci. Rep. 2020, 40, BSR20193461. [Google Scholar] [CrossRef]
- Huang, M.; Zheng, J.; Ren, Y.; Zhu, J.; Kou, L.; Nie, J. LINC00221 suppresses the malignancy of children acute lymphoblastic leukemia. Biosci. Rep. 2020, 40, BSR20194070. [Google Scholar] [CrossRef] [Green Version]
- Ouimet, M.; Drouin, S.; Lajoie, M.; Caron, M.; St-Onge, P.; Gioia, R.; Richer, C.; Sinnett, D. A childhood acute lymphoblastic leukemia-specific lncRNA implicated in prednisolone resistance, cell proliferation, and migration. Oncotarget 2017, 8, 7477–7488. [Google Scholar] [CrossRef] [Green Version]
- Fernando, T.R.; Contreras, J.R.; Zampini, M.; Rodriguez-Malave, N.I.; Alberti, M.O.; Anguiano, J.; Tran, T.M.; Palanichamy, J.K.; Gajeton, J.; Ung, N.M.; et al. The lncRNA CASC15 regulates SOX4 expression in RUNX1-rearranged acute leukemia. Mol. Cancer. 2017, 16, 126. [Google Scholar] [CrossRef]
- Wang, Q.; Du, X.; Yang, M.; Xiao, S.; Cao, J.; Song, J.; Wang, L. LncRNA ZEB1-AS1 contributes to STAT3 activation by associating with IL-11 in B-lymphoblastic leukemia. Biotechnol. Lett. 2017, 39, 1801–1810. [Google Scholar] [CrossRef]
- Rodríguez-Malavé, N.I.; Fernando, T.R.; Patel, P.C.; Contreras, J.R.; Palanichamy, J.K.; Tran, T.M.; Anguiano, J.; Davoren, M.J.; Alberti, M.O.; Pioli, K.T.; et al. BALR-6 regulates cell growth and cell survival in B-lymphoblastic leukemia. Mol. Cancer 2015, 14, 214. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, J.; Guo, G.; Feng, R.; Chen, K.; Liao, Y.; Zhang, L.; Sun, L.; Huang, S.; Chen, J.L. Novel lncRNA-IUR suppresses Bcr-Abl-induced tumorigenesis through regulation of STAT5-CD71 pathway. Mol. Cancer 2019, 18, 84. [Google Scholar] [CrossRef]
- Wang, W.; Wu, F.; Ma, P.; Gan, S.; Li, X.; Chen, L.; Sun, L.; Sun, H.; Jiang, Z.; Guo, F. LncRNA CRNDE Promotes the Progression of B-cell Precursor Acute Lymphoblastic Leukemia by Targeting the miR-345-5p/CREB Axis. Mol. Cells 2020, 43, 718–727. [Google Scholar]
- Wang, W.T.; Chen, T.Q.; Zeng, Z.C.; Pan, Q.; Huang, W.; Han, C.; Fang, K.; Sun, L.Y.; Yang, Q.Q.; Wang, D.; et al. The lncRNA LAMP5-AS1 drives leukemia cell stemness by directly modulating DOT1L methyltransferase activity in MLL leukemia. J. Hematol. Oncol. 2020, 13, 78. [Google Scholar] [CrossRef]
- Orlandella, F.M.; Smaldone, G.; Salvatore, G.; Vitagliano, L.; Cianflone, A.; Parasole, R.; Beneduce, G.; Menna, G.; Salvatore, M.; Mirabelli, P. The lncRNA TEX41 is upregulated in pediatric B-Cells Acute Lymphoblastic Leukemia and it is necessary for leukemic cell growth. Biomark. Res. 2021, 9, 54. [Google Scholar] [CrossRef]
- Fernando, T.R.; Rodriguez-Malave, N.I.; Waters, E.V.; Yan, W.; Casero, D.; Basso, G.; Pigazzi, M.; Rao, D.S. LncRNA Expression Discriminates Karyotype and Predicts Survival in B-Lymphoblastic Leukemia. Mol. Cancer Res. 2015, 13, 839–851. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhou, S.; Zhou, T.; Li, X.; Tang, J. Targeting the lncRNA DUXAP8/miR-29a/PIK3CA Network Restores Doxorubicin Chemosensitivity via PI3K-AKT-mTOR Signaling and Synergizes With Inotuzumab Ozogamicin in Chemotherapy-Resistant B-Cell Acute Lymphoblastic Leukemia. Front. Oncol. 2022, 12, 773601. [Google Scholar] [CrossRef]
- Zhao, D.; Xing, Q.; Song, H.; Zhao, Y.; Guo, G. LINC00265/miR-4500 Axis Accelerates Acute Lymphoblastic Leukemia Progression by Enhancing STAT3 Signals. Cancer Manag. Res. 2021, 13, 8147–8156. [Google Scholar] [CrossRef]
- Li, G.; Lei, X.; Zhang, Y.; Liu, Z.; Zhu, K. LncRNA PPM1A-AS Regulate Tumor Development Through Multiple Signal Pathways in T-Cell Acute Lymphoblastic Leukemia. Front. Oncol. 2021, 11, 761205. [Google Scholar] [CrossRef]
- Zeng, P.; Chai, Y.; You, C.; Yue, L.; Wu, C.; Chen, H.; Li, L.; Li, J.; Liu, H.; Zhang, Y.; et al. Correlation analysis of long non-coding RNA TUG1 with disease risk, clinical characteristics, treatment response, and survival profiles of adult Ph− Acute lymphoblastic leukemia. J. Clin. Lab. Anal. 2021, 35, e23583. [Google Scholar] [CrossRef]
- Abuduer, M.; A, E.Z.G. LINC00665 promotes the viability, migration and invasion of T cell acute lymphoblastic leukemia cells by targeting miR-101 via modulating PI3K/Akt pathway. Tissue Cell 2021, 71, 101579. [Google Scholar] [CrossRef]
- Mao, J.; Gao, W.; Xue, L.; Wang, J.; Zhao, L. The lncRNA SLCO4A1-AS1/miR-876-3p/RBBP6 axis regulates cell proliferation and apoptosis in acute lymphocytic leukemia via the JNK signaling pathway. Int. J. Lab. Hematol. 2021, 43, 1050–1061. [Google Scholar] [CrossRef]
- Ju, J.K.; Han, W.N.; Shi, C.L. Long non-coding RNA (lncRNA) plasmacytoma variant translocation 1 gene (PVT1) modulates the proliferation and apoptosis of acute lymphoblastic leukemia cells by sponging miR-486-5p. Bioengineered 2022, 13, 4587–4597. [Google Scholar] [CrossRef]
- Yazdi, N.; Houshmand, M.; Atashi, A.; Kazemi, A.; Najmedini, A.A.; Zarif, M.N. Long noncoding RNA PVT1: Potential oncogene in the development of acute lymphoblastic leukemia. Turk. J. Biol. 2018, 42, 405–413. [Google Scholar] [CrossRef]
- Yang, J.; Yang, Y. Long noncoding RNA endogenous bornavirus-like nucleoprotein acts as an oncogene by regulating microRNA-655-3p expression in T-cell acute lymphoblastic leukemia. Bioengineered 2022, 13, 6409–6419. [Google Scholar] [CrossRef]
- Garitano-Trojaola, A.; José-Enériz, E.S.; Ezponda, T.; Unfried, J.P.; Carrasco-León, A.; Razquin, N.; Barriocanal, M.; Vilas-Zornoza, A.; Sangro, B.; Segura, V.; et al. Deregulation of linc-PINT in acute lymphoblastic leukemia is implicated in abnormal proliferation of leukemic cells. Oncotarget 2018, 9, 12842–12852. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Huang, S.H.; Zhou, H.R.; Chen, C.J.; Tian, L.H.; Shen, J.Z. Role of HOTAIR in the diagnosis and prognosis of acute leukemia. Oncol. Rep. 2016, 36, 3113–3122. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Zhao, S.; Li, J.; Zhang, H.; Qian, C.; Wang, H.; Liu, J.; Zhao, Y. TCF7L2 activated HOXA-AS2 decreased the glucocorticoid sensitivity in acute lymphoblastic leukemia through regulating HOXA3/EGFR/Ras/Raf/ MEK/ERK pathway. Biomed. Pharmacother. 2019, 109, 1640–1649. [Google Scholar] [CrossRef]
- Yang, T.; Jin, X.; Lan, J.; Wang, W. Long non-coding RNA SNHG16 has Tumor suppressing effect in acute lymphoblastic leukemia by inverse interaction on hsa-miR-124-3p. IUBMB Life 2019, 71, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Guo, N.H.; Zheng, J.F. LncRNA-MALAT1 regulates proliferation and apoptosis of acute lymphoblastic leukemia cells via miR-205-PTK7 pathway. Pathol. Int. 2020, 70, 724–732. [Google Scholar] [CrossRef]
- Melo, C.P.; Campos, C.B.; Rodrigues, J.; Aguirre-Neto, J.C.; Atalla, Â.; Pianovski, M.A.; Carbone, E.K.; Lares, L.B.; Moraes-Souza, H.; Octacílio-Silva, S.; et al. Long non-coding RNAs: Bioarkers for acute leukaemia subtypes. Br. J. Haematol. 2016, 173, 318–320. [Google Scholar] [CrossRef]
- Wallaert, A.; Durinck, K.; Van Loocke, W.; Van de Walle, I.; Matthijssens, F.; Volders, P.J.; Avila Cobos, F.; Rombaut, D.; Rondou, P.; Mestdagh, P.; et al. Long noncoding RNA signatures define oncogenic subtypes in T-cell acute lymphoblastic leukemia. Leukemia 2016, 30, 1927–1930. [Google Scholar] [CrossRef]
- Arthur, G.; Almamun, M.; Taylor, K. Hypermethylation of antisense long noncoding RNAs in acute lymphoblastic leukemia. Epigenomics 2017, 9, 635–645. [Google Scholar] [CrossRef]
- Cheng, H.; Huang, C.M.; Wang, Y.; Hu, X.X.; Xu, X.Q.; Song, X.M.; Tang, G.S.; Chen, L.; Yang, J.M. Microarray profiling and co-expression network analysis of the lncRNAs and mRNAs associated with acute leukemia in adults. Mol. Biosyst. 2017, 13, 1102–1108. [Google Scholar] [CrossRef]
- Lajoie, M.; Drouin, S.; Caron, M.; St-Onge, P.; Ouimet, M.; Gioia, R.; Lafond, M.H.; Vidal, R.; Richer, C.; Oualkacha, K.; et al. Specific expression of novel long non-coding RNAs in high-hyperdiploid childhood acute lymphoblastic leukemia. PLoS ONE 2017, 12, e0174124. [Google Scholar] [CrossRef] [Green Version]
- Caron, M.; St-Onge, P.; Drouin, S.; Richer, C.; Sontag, T.; Busche, S.; Bourque, G.; Pastinen, T.; Sinnett, D. Very long intergenic non-coding RNA transcripts and expression profiles are associated to specific childhood acute lymphoblastic leukemia subtypes. PLoS ONE 2018, 13, e0207250. [Google Scholar] [CrossRef]
- Cuadros, M.; Andrades, Á.; Coira, I.F.; Baliñas, C.; Rodríguez, M.I.; Álvarez-Pérez, J.C.; Peinado, P.; Arenas, A.M.; García, D.J.; Jiménez, P.; et al. Expression of the long non-coding RNA TCL6 is associated with clinical outcome in pediatric B-cell acute lymphoblastic leukemia. Blood Cancer J. 2019, 9, 93. [Google Scholar] [CrossRef] [Green Version]
- James, A.R.; Schroeder, M.P.; Neumann, M.; Bastian, L.; Eckert, C.; Gökbuget, N.; Tanchez, J.O.; Schlee, C.; Isaakidis, K.; Schwartz, S.; et al. Long non-coding RNAs defining major subtypes of B cell precursor acute lymphoblastic leukemia. J. Hematol. Oncol. 2019, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Bian, H.; Cao, Y.; Juan, C.; Cao, Q.; Zhou, G.; Fang, Y. Identification of novel lncRNAs involved in the pathogenesis of childhood acute lymphoblastic leukemia. Oncol. Lett. 2019, 17, 2081–2090. [Google Scholar] [CrossRef] [Green Version]
- Affinito, O.; Pane, K.; Smaldone, G.; Orlandella, F.M.; Mirabelli, P.; Beneduce, G.; Parasole, R.; Ripaldi, M.; Salvatore, M.; Franzese, M. lncRNAs-mRNAs Co-Expression Network Underlying Childhood B-Cell Acute Lymphoblastic Leukaemia: A Pilot Study. Cancers 2020, 12, 2489. [Google Scholar] [CrossRef]
- Bárcenas-López, D.A.; Núñez-Enríquez, J.C.; Hidalgo-Miranda, A.; Beltrán-Anaya, F.O.; May-Hau, D.I.; Jiménez-Hernández, E.; Bekker-Méndez, V.C.; Flores-Lujano, J.; Medina-Sansón, A.; Tamez-Gómez, E.L.; et al. Transcriptome Analysis Identifies LINC00152 as a Biomarker of Early Relapse and Mortality in Acute Lymphoblastic Leukemia. Genes 2020, 11, 302. [Google Scholar] [CrossRef] [Green Version]
- Cuadros, M.; García, D.J.; Andrades, A.; Arenas, A.M.; Coira, I.F.; Baliñas-Gavira, C.; Peinado, P.; Rodríguez, M.I.; Álvarez-Pérez, J.C.; Ruiz-Cabello, F.; et al. LncRNA-mRNA Co-Expression Analysis Identifies AL133346.1/CCN2 as Biomarkers in Pediatric B-Cell Acute Lymphoblastic Leukemia. Cancers 2020, 12, 3803. [Google Scholar] [CrossRef]
- Yu, W.; Wang, W.; Yu, X. Investigation of lncRNA-mRNA co-expression network in ETV6-RUNX1-positive pediatric B-cell acute lymphoblastic leukemia. PLoS ONE 2021, 16, e0253012. [Google Scholar] [CrossRef]
- Das Chagas, P.F.; de Sousa, G.R.; Kodama, M.H.; de Biagi Junior, C.; Yunes, J.A.; Brandalise, S.R.; Calin, G.A.; Tone, L.G.; Scrideli, C.A.; de Oliveira, J.C. Ultraconserved long non-coding RNA uc.112 is highly expressed in childhood T versus B-cell acute lymphoblastic leukemia. Hematol. Transfus. Cell Ther. 2021, 43, 28–34. [Google Scholar] [CrossRef]
- Wang, W.; Lyu, C.; Wang, F.; Wang, C.; Wu, F.; Li, X.; Gan, S. Identification of Potential Signatures and Their Functions for Acute Lymphoblastic Leukemia: A Study Based on the Cancer Genome Atlas. Front. Genet. 2021, 12, 656042. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wang, M.; Zhu, Y.; Bu, C.; Li, T. Differential mRNA and long noncoding RNA expression profiles in pediatric B-cell acute lymphoblastic leukemia patients. BMC Pediatr. 2022, 22, 10. [Google Scholar] [CrossRef] [PubMed]
- Ngoc, P.C.T.; Tan, S.H.; Tan, T.K.; Chan, M.M.; Li, Z.; Yeoh, A.; Tenen, D.G.; Sanda, T. Identification of novel lncRNAs regulated by the TAL1 complex in T-cell acute lymphoblastic leukemia. Leukemia 2018, 32, 2138–2151. [Google Scholar] [CrossRef] [PubMed]
- Verboom, K.; Van Loocke, W.; Volders, P.J.; Decaesteker, B.; Cobos, F.A.; Bornschein, S.; de Bock, C.E.; Atak, Z.K.; Clappier, E.; Aerts, S.; et al. A comprehensive inventory of TLX1 controlled long non-coding RNAs in T-cell acute lymphoblastic leukemia through polyA+ and total RNA sequencing. Haematologica 2018, 103, e585–e589. [Google Scholar] [CrossRef]
- Sanda, T.; Lawton, L.N.; Barrasa, M.I.; Fan, Z.P.; Kohlhammer, H.; Gutierrez, A.; Ma, W.; Tatarek, J.; Ahn, Y.; Kelliher, M.A.; et al. Core transcriptional regulatory circuit controlled by the TAL1 complex in human T cell acute lymphoblastic leukemia. Cancer Cell 2012, 22, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Grabher, C.; von Boehmer, H.; Look, A.T. Notch 1 activation in the molecular pathogenesis of T-cell acute lymphoblastic leukaemia. Nat. Rev. Cancer 2006, 6, 347–359. [Google Scholar] [CrossRef]
- Durinck, K.; Wallaert, A.; Van de Walle, I.; Van Loocke, W.; Volders, P.J.; Vanhauwaert, S.; Geerdens, E.; Benoit, Y.; Van Roy, N.; Poppe, B.; et al. The Notch driven long non-coding RNA repertoire in T-cell acute lymphoblastic leukemia. Haematologica 2014, 99, 1808–1816. [Google Scholar] [CrossRef] [Green Version]
- Renneville, A.; Gasser, J.A.; Grinshpun, D.E.; Jean Beltran, P.M.; Udeshi, N.D.; Matyskiela, M.E.; Clayton, T.; McConkey, M.; Viswanathan, K.; Tepper, A.; et al. Avadomide induces degradation of ZMYM2 fusion oncoproteins in hematologic malignancies. Blood Cancer Discov. 2021, 2, 250–265. [Google Scholar] [CrossRef]
- Tsuzuki, S.; Yasuda, T.; Kojima, S.; Kawazu, M.; Akahane, K.; Inukai, T.; Imaizumi, M.; Morishita, T.; Miyamura, K.; Ueno, T.; et al. Targeting MEF2D-fusion Oncogenic Transcriptional Circuitries in B-cell Precursor Acute Lymphoblastic Leukemia. Blood Cancer Discov. 2020, 1, 82–95. [Google Scholar] [CrossRef]
- Montaño, A.; Ordoñez, J.L.; Alonso-Pérez, V.; Hernández-Sánchez, J.; Santos, S.; González, T.; Benito, R.; García-Tuñón, I.; Hernández-Rivas, J.M. ETV6/RUNX1 Fusion Gene Abrogation Decreases the Oncogenicity of Tumour Cells in a Preclinical Model of Acute Lymphoblastic Leukaemia. Cells 2020, 9, 215. [Google Scholar] [CrossRef] [Green Version]
- Ghazavi, F.; De Moerloose, B.; Van Loocke, W.; Wallaert, A.; Helsmoortel, H.H.; Ferster, A.; Bakkus, M.; Plat, G.; Delabesse, E.; Uyttebroeck, A.; et al. Unique long non-coding RNA expression signature in ETV6/RUNX1-driven B-cell precursor acute lymphoblastic leukemia. Oncotarget 2016, 7, 73769–73780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Chaer, F.; Keng, M.; Ballen, K.K. MLL-Rearranged Acute Lymphoblastic Leukemia. Curr. Hematol. Malig. Rep. 2020, 15, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Han, B.W.; Chen, Z.H.; Lin, K.Y.; Zeng, C.W.; Li, X.J.; Li, J.H.; Luo, X.Q.; Chen, Y.Q. A distinct set of long non-coding RNAs in childhood MLL-rearranged acute lymphoblastic leukemia: Biology and epigenetic target. Hum. Mol. Genet. 2014, 23, 3278–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, J.C.; Andersen, M.H.; Schrama, D.; Thor Straten, P. Immune-suppressive properties of the tumor microenvironment. Cancer Immunol. Immunother. 2013, 62, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Niesel, K.; Schulz, M.; Anthes, J.; Alekseeva, T.; Macas, J.; Salamero-Boix, A.; Möckl, A.; Oberwahrenbrock, T.; Lolies, M.; Stein, S.; et al. The immune suppressive microenvironment affects efficacy of radio-immunotherapy in brain metastasis. EMBO Mol. Med. 2021, 13, e13412. [Google Scholar] [CrossRef]
- Haloupek, N. The Landscape of Blood Cancer Research Today—And Where the Field Is Headed. Blood Cancer Discov. 2020, 1, 1–4. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Sun, X.; Rong, L.; Kang, M.; Wu, P.; Ji, X.; Lin, R.; Huang, J.; Xue, Y.; et al. Bone marrow infiltrated Lnc-INSR induced suppressive immune microenvironment in pediatric acute lymphoblastic leukemia. Cell Death Dis. 2018, 9, 1043. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, H.; Chen, S.Y.; Liu, C.J.; Hu, F.F.; Yu, J.; Wu, Y.; Guo, A.Y. Transcriptome and Regulatory Network Analyses of CD19-CAR-T Immunotherapy for B-ALL. Genom. Proteom. Bioinform. 2019, 17, 190–200. [Google Scholar] [CrossRef]
- Hashemi, M.; Bahari, G.; Naderi, M.; Sadeghi Bojd, S.; Taheri, M. Association of lnc-LAMC2-1:1 rs2147578 and CASC8 rs10505477 Polymorphisms with Risk of Childhood Acute Lymphoblastic Leukemia. Asian Pac. J. Cancer Prev. 2016, 17, 4985–4989. [Google Scholar]
- Han, J.; Zhou, W.; Jia, M.; Wen, J.; Jiang, J.; Shi, J.; Zhang, K.; Ma, H.; Liu, J.; Ren, J.; et al. Expression quantitative trait loci in long non-coding RNA PAX8-AS1 are associated with decreased risk of cervical cancer. Mol. Genet. Genom. 2016, 291, 1743–1748. [Google Scholar] [CrossRef]
- Bahari, G.; Hashemi, M.; Naderi, M.; Sadeghi-Bojd, S.; Taheri, M. Long non-coding RNA PAX8-AS1 polymorphisms increase the risk of childhood acute lymphoblastic leukemia. Biomed. Rep. 2018, 8, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.S.; Chang, W.S.; Chen, C.C.; Mong, M.C.; Hsu, S.W.; Hsu, P.C.; Hsu, Y.N.; Wang, Y.C.; Tsai, C.W.; Bau, D.T. Novel Contribution of Long Non-coding RNA MEG3 Genotype to Prediction of Childhood Leukemia Risk. Cancer Genom. Proteom. 2022, 19, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Gasic, V.; Stankovic, B.; Zukic, B.; Janic, D.; Dokmanovic, L.; Krstovski, N.; Lazic, J.; Milosevic, G.; Lucafò, M.; Stocco, G.; et al. Expression Pattern of Long Non-coding RNA Growth Arrest-specific 5 in the Remission Induction Therapy in Childhood Acute Lymphoblastic Leukemia. J. Med. Biochem. 2019, 38, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Gioia, R.; Drouin, S.; Ouimet, M.; Caron, M.; St-Onge, P.; Richer, C.; Sinnett, D. LncRNAs downregulated in childhood acute lymphoblastic leukemia modulate apoptosis, cell migration, and DNA damage response. Oncotarget 2017, 8, 80645–80650. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Chi, L.; Wang, X.; Jin, X.; Wang, W.; Lan, J. Identification of a Seven-lncRNA-mRNA Signature for Recurrence and Prognostic Prediction in Relapsed Acute Lymphoblastic Leukemia Based on WGCNA and LASSO Analyses. Anal. Cell. Pathol. 2021, 2021, 6692022. [Google Scholar] [CrossRef]
- Pouyanrad, S.; Rahgozar, S.; Ghodousi, E.S. Dysregulation of miR-335-3p, targeted by NEAT1 and MALAT1 long non-coding RNAs, is associated with poor prognosis in childhood acute lymphoblastic leukemia. Gene 2019, 692, 35–43. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, F.; Liu, H.; Fan, F.; Lin, Y.; Zhou, J.; Cai, Y.; Zhang, X.; Wu, Y.; Mao, R.; et al. Identification of a nomogram based on an 8-lncRNA signature as a novel diagnostic biomarker for childhood acute lymphoblastic leukemia. Aging 2021, 13, 15548–15568. [Google Scholar] [CrossRef]
- Gao, W. Long non-coding RNA MEG3 as a candidate prognostic factor for induction therapy response and survival profile in childhood acute lymphoblastic leukemia patients. Scand. J. Clin. Lab. Investig. 2021, 81, 194–200. [Google Scholar] [CrossRef]
- Xagorari, M.; Marmarinos, A.; Kossiva, L.; Baka, M.; Doganis, D.; Servitzoglou, M.; Tsolia, M.; Scorilas, A.; Avgeris, M.; Gourgiotis, D. Overexpression of the GR Riborepressor LncRNA GAS5 Results in Poor Treatment Response and Early Relapse in Childhood B-ALL. Cancers 2021, 13, 6064. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Verduci, L.; Tarcitano, E.; Strano, S.; Yarden, Y.; Blandino, G. CircRNAs: Role in human diseases and potential use as biomarkers. Cell Death Dis. 2021, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starke, S.; Jost, I.; Rossbach, O.; Schneider, T.; Schreiner, S.; Hung, L.H.; Bindereif, A. Exon circularization requires canonical splice signals. Cell Rep. 2015, 10, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xue, W.; Li, X.; Zhang, J.; Chen, S.; Zhang, J.L.; Yang, L.; Chen, L.L. The Biogenesis of Nascent Circular RNAs. Cell Rep. 2016, 15, 611–624. [Google Scholar] [CrossRef] [Green Version]
- Van Zonneveld, A.J.; Kölling, M.; Bijkerk, R.; Lorenzen, J.M. Circular RNAs in kidney disease and cancer. Nat. Rev. Nephrol. 2021, 17, 814–826. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N6-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [Green Version]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Rong, Y.; Tang, X.; Yi, K.; Wu, J.; Wang, F. Circular RNAs Are Promising Biomarkers in Liquid Biopsy for the Diagnosis of Non-small Cell Lung Cancer. Front. Mol. Biosci. 2021, 8, 625722. [Google Scholar] [CrossRef]
- Nicolet, B.P.; Engels, S.; Aglialoro, F.; van den Akker, E.; von Lindern, M.; Wolkers, M.C. Circular RNA expression in human hematopoietic cells is widespread and cell-type specific. Nucleic Acids Res. 2018, 46, 8168–8180. [Google Scholar] [CrossRef]
- Yu, C.Y.; Li, T.C.; Wu, Y.Y.; Yeh, C.H.; Chiang, W.; Chuang, C.Y.; Kuo, H.C. The circular RNA circBIRC6 participates in the molecular circuitry controlling human pluripotency. Nat. Commun. 2017, 8, 1149. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, D.; Pu, W.; Wang, J.; Peng, Y. Circular RNAs in Cancer: Biogenesis, Function, and Clinical Significance. Trends Cancer 2020, 6, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Jeha, S.; Choi, J.; Roberts, K.G.; Pei, D.; Coustan-Smith, E.; Inaba, H.; Rubnitz, J.E.; Ribeiro, R.C.; Gruber, T.A.; Raimondi, S.C.; et al. Clinical significance of novel subtypes of acute lymphoblastic leukemia in the context of minimal residual disease-directed therapy. Blood Cancer Discov. 2021, 2, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.; Hilton, L.K.; Tang, J.; Rushton, C.K.; Grande, B.M.; Scott, D.W.; Morin, R.D. PRPS-ST: A protocol-agnostic self-training method for gene expression-based classification of blood cancers. Blood Cancer Discov. 2020, 1, 244–257. [Google Scholar] [CrossRef]
- Gaffo, E.; Boldrin, E.; Dal Molin, A.; Bresolin, S.; Bonizzato, A.; Trentin, L.; Frasson, C.; Debatin, K.M.; Meyer, L.H.; Te Kronnie, G.; et al. Circular RNA differential expression in blood cell populations and exploration of circRNA deregulation in pediatric acute lymphoblastic leukemia. Sci. Rep. 2019, 9, 14670. [Google Scholar] [CrossRef] [Green Version]
- Buratin, A.; Paganin, M.; Gaffo, E.; Dal Molin, A.; Roels, J.; Germano, G.; Siddi, M.T.; Serafin, V.; De Decker, M.; Gachet, S.; et al. Large-scale circular RNA deregulation in T-ALL: Unlocking unique ectopic expression of molecular subtypes. Blood Adv. 2020, 4, 5902–5914. [Google Scholar] [CrossRef]
- Guo, S.; Li, B.; Chen, Y.; Zou, D.; Yang, S.; Zhang, Y.; Wu, N.; Sheng, L.; Huang, H.; Ouyang, G.; et al. Hsa_circ_0012152 and Hsa_circ_0001857 Accurately Discriminate Acute Lymphoblastic Leukemia From Acute Myeloid Leukemia. Front. Oncol. 2020, 10, 1655. [Google Scholar] [CrossRef]
- Panda, A.C.; Grammatikakis, I.; Kim, K.M.; De, S.; Martindale, J.L.; Munk, R.; Yang, X.; Abdelmohsen, K.; Gorospe, M. Identification of senescence-associated circular RNAs (SAC-RNAs) reveals senescence suppressor CircPVT1. Nucleic Acids Res. 2017, 45, 4021–4035. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Zheng, Q.; Bao, C.; He, J.; Chen, B.; Lyu, D.; Zheng, B.; Xu, Y.; Long, Z.; et al. Circular RNA profile identifies circPVT1 as a proliferative factor and prognostic marker in gastric cancer. Cancer Lett. 2017, 388, 208–219. [Google Scholar] [CrossRef]
- Hu, J.; Han, Q.; Gu, Y.; Ma, J.; McGrath, M.; Qiao, F.; Chen, B.; Song, C.; Ge, Z. Circular RNA PVT1 expression and its roles in acute lymphoblastic leukemia. Epigenomics 2018, 10, 723–732. [Google Scholar] [CrossRef]
- Jia, Y.; Hu, H.; Gu, W. Regulation Effects of Circular RNA CircPVT1 and miR-125b on NF-κB Signal Pathway in Childhood ALL. Clin. Lab. 2021, 67. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Gu, W. Up-regulation of circPVT1 in T cell acute lymphoblastic leukemia promoted cell proliferation via miR-30e/DLL4 induced activating NOTCH signaling. Pathol. Res. Pract. 2021, 224, 153536. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, C.; Li, Y.; Deng, Y.; Lu, W.; Li, J. Upregulation of circ-0000745 in acute lymphoblastic leukemia enhanced cell proliferation by activating ERK pathway. Gene 2020, 751, 144726. [Google Scholar] [CrossRef]
- Feng, H.; Li, F.; Tang, P. Circ_0000745 regulates NOTCH1-mediated cell proliferation and apoptosis in pediatric T-cell acute lymphoblastic leukemia through adsorbing miR-193b-3p. Hematology 2021, 26, 885–895. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, Y.; Zhang, Y.; Liu, J. Circ_0000745 promotes acute lymphoblastic leukemia progression through mediating miR-494-3p/NET1 axis. Hematology 2022, 27, 11–22. [Google Scholar] [CrossRef]
- Zheng, Y.; Niu, B.; Zhang, W.; Ru, X.; Gao, Y.; Li, C.; Wu, X. Circular RNA circPRKCI contributes to malignant progression of T-cell acute lymphoblastic leukemia by modulating miR-20a-5p/SOX4 axis. Aging 2021, 13, 23757–23768. [Google Scholar] [CrossRef]
- Ling, Z.; Fang, Z.G.; Wu, J.Y.; Liu, J.J. The depletion of Circ-PRKDC enhances autophagy and apoptosis in T-cell acute lymphoblastic leukemia via microRNA-653-5p/Reelin mediation of the PI3K/AKT/mTOR signaling pathway. Kaohsiung J. Med. Sci. 2021, 37, 392–401. [Google Scholar] [CrossRef]
- Hou, Y.; Sun, J.; Huang, J.; Yao, F.; Chen, X.; Zhu, B.; Zhao, D. Circular RNA circRNA_0000094 sponges microRNA-223-3p and up-regulate F-box and WD repeat domain containing 7 to restrain T cell acute lymphoblastic leukemia progression. Hum Cell. 2021, 34, 977–989. [Google Scholar] [CrossRef]
- Tong, J.; Liu, H.; Yao, W.; Zhang, L.; Song, K.; Sun, G.; Sun, Z. Down-regulation of circRNA_0000143 regulates miR-142-3p/GRα axis to facilitate the progression of T-ALL. Cancer Sci. 2021. Epub ahead of print. [Google Scholar] [CrossRef]
- Zhu, Y.; Ma, X.; Zhang, H.; Wu, Y.; Kang, M.; Fang, Y.; Xue, Y. Mechanism of circADD2 as ceRNA in Childhood Acute Lymphoblastic Leukemia. Front. Cell Dev. Biol. 2021, 9, 639910. [Google Scholar] [CrossRef]
- He, A.T.; Liu, J.; Li, F.; Yang, B.B. Targeting circular RNAs as a therapeutic approach: Current strategies and challenges. Signal Transduct. Target. Ther. 2021, 6, 185. [Google Scholar] [CrossRef] [PubMed]
- Roels, J.; Thénoz, M.; Szarzyńska, B.; Landfors, M.; De Coninck, S.; Demoen, L.; Provez, L.; Kuchmiy, A.; Strubbe, S.; Reunes, L.; et al. Aging of preleukemic thymocytes drives CpG island hypermethylation in T-cell acute lymphoblastic leukemia. Blood Cancer Discov. 2020, 1, 274–289. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| lncRNA | Subject | Function | Alteration | Target Regulation | Effect on All Cell Phenotype | References |
|---|---|---|---|---|---|---|
| LUNAR1 | T-ALL cell lines and patient samples | oncogene | upregulated | enhancing IGF1R mRNA expression, sustaining IGF1 signaling | promoting cell growth via acting as NOTCH effector | [77,78] |
| T-ALL-R-LncR1 | T-ALL cell lines and patient samples | oncogene | upregulated | inhibiting pro-apoptotic factor Par-4/THAP1 protein complex | inhibiting apoptosis | [79] |
| NALT1 | T-ALL cell lines and patient samples | oncogene | upregulated | causing transcriptional activation of NOTCH1 signaling | promoting cell proliferation | [80] |
| LINC00478 | T-ALL cell lines and patient samples | oncogene | upregulated | upregulating miR-125b production | promoting cell growth and invasiveness | [81] |
| ARIEL | T-ALL cell lines and patient samples | oncogene | upregulated | enhancing ARID5B gene expression | promoting cell growth and survival | [82] |
| CDKN2B-AS1 | T-ALL cell lines and patient samples | oncogene | upregulated | sponging miR-335-3p to upregulate TRAF5 | promoting cell proliferation and cell cycle progression | [83] |
| LINC00511 | T-ALL cell lines and patient samples | oncogene | upregulated | sponging miR-195-5p to upregulate LRRK1 | boosting cell proliferation and invasion, inhibiting apoptosis | [84] |
| ANRIL | T-ALL cell lines and patient samples | oncogene | upregulated | suppressing miR-7-5p to upregulate TCF4 | promoting cell viability, migration, and invasion | [85] |
| AWPPH | T-ALL cell lines and patient samples | oncogene | upregulated | upregulation of ROCK2 expression | promoting proliferation and inhibiting apoptosis | [86] |
| NEAT1 | T-ALL cell lines and patient samples | oncogene | upregulated | sponging miR-146b-5p to upregulate expression of NOTCH1 | promoting cell proliferation and growth | [87] |
| H19 | T-ALL cell lines and patient samples | oncogene | upregulated | inducing SOX2, OCT-4, and NANOG expression; decreasing miR-326 level to upregulate BCL-2 expression | maintaining stemness and promoting cell proliferation; inhibiting apoptosis | [88,89,90] |
| LINC00853 | T-ALL cell lines and patient samples | tumor suppressor | downregulated | upregulating CCR9 expression | inhibiting cell proliferation, migration, and invasion | [91] |
| LNC00221 | T-ALL cell lines Jurkat, CCRF-CEM, CEM/C1 | tumor suppressor | downregulated | sponging miR-152-3p to upregulate ATPA2A | inhibiting cell proliferation and apoptosis | [92] |
| VPS9D1-AS1 | T-ALL cell lines Molt-3 and Molt-4 | oncogene | upregulated | inhibiting miR-419-5p, miR-214-3p to upregulate GPX1 | inhibiting apoptosis and promoting survival | [93] |
| lncRNA | Subject | Function | Alteration | Target Regulation | Effect on ALL Cell Phenotype | Reference |
|---|---|---|---|---|---|---|
| RP11-137H2.4 | B-ALL cell lines and patient samples | oncogene | upregulated | modulated expression of NRAS/BRAF/NF-κB MAPK cascade and cell cycle pathways | enhancing cell proliferation, migration, and apoptosis, inducing GC resistance | [94] |
| CASC15 | RUNX1/AML+ B-ALL cell lines and patient samples | oncogene | upregulated | enhancing YY1-mediated regulation of the SOX4 promoter | promoting cell proliferation and survival | [95] |
| ZEB1-AS1 | B-ALL cell lines and patient samples | oncogene | upregulated | binding to IL-11 and promoting IL-11 stability, activating STAT3 | increasing cell proliferation | [96] |
| BALR-6 | B-ALL cell lines and patient samples | oncogene | upregulated | enhancing SP1-mediated transcription of CREB1 | increasing proliferation and decreasing apoptosis | [97] |
| IUR | Ph+ B-ALL patient samples and cell lines | tumor suppressor | downregulated | inhibiting STAT5-CD71 pathway | suppression of BCR-ABL1-mediated tumorigenesis | [98] |
| CRNDE | B-ALL cell lines and patient samples | oncogene | upregulated | sequestering miR-345-5p to upregulate CREB | promoting cell proliferation and inhibiting apoptosis | [99] |
| LAMP5-AS1 | MLL rearranged B-ALL patient samples and cell lines | oncogene | upregulated | promoting methyltransferase activity of DOT1L to facilitate H3K9me2/me3 and increase HOXA expression | increasing colony formation and inhibiting differentiation | [100] |
| TEX41 | B-ALL patient samples and cell lines | oncogene | upregulated | decreasing CDK4, CDK6 and p27 level, increasing p21 and p53 level | promoting cell growth and inhibiting cell cycle | [101] |
| BALR-2 | B-ALL patient samples and cell lines | oncogene | upregulated | decreasing JUN and BIM expression | increasing cell growth and conferring resistance to prednisone treatment | [102] |
| DUXAP8 | B-ALL patient samples and cell lines | oncogene | upregulated | sequestering miR-29a to increase PIK3CA expression | boosting proliferation, inhibiting apoptosis, and conferring Dox resistance | [103] |
| lncRNA | Subject | Function | Alteration | Target Regulation | Effect on ALL Cell Phenotype | References |
|---|---|---|---|---|---|---|
| LINC00265 | ALL patient samples and cell lines | oncogene | upregulated | sponging miR-4500 to enhance STAT3 expression | facilitating cell growth, proliferation, and migration | [104] |
| PPM1A-AS | ALL patient samples and cell lines | oncogene | upregulated | increasing phosphorylation of STAT3, Akt, and Notch4 | promoting cell proliferation and inhibiting apoptosis | [105] |
| TUG1 | Ph- ALL patient samples | oncogene | upregulated | unknown | promoting CNS infiltration | [106] |
| LINC00665 | ALL patient samples and cell lines | oncogene | upregulated | suppressing miR-101 to activate PI3K/Akt pathway | promoting cell viability, migration, and invasion | [107] |
| SLCO4A1-AS1 | ALL patient samples and cell lines | oncogene | upregulated | sponging miR-876-3p to upregulate RBBP6 and activate JNK | inhibited cell proliferation and promoted apoptosis | [108] |
| PVT1 | ALL patient samples and cell lines | oncogene | upregulated | sponging miR-486-5p to increase MAML3 expression; increasing NOP2 and c-Myc, decreasing p15, p16 | increasing cell viability restraining apoptosis; deregulating cell cycle, inhibiting apoptosis | [109,110] |
| EBLN3P | ALL patient samples and cell lines | oncogene | upregulated | negatively regulating miR-655-3p | promoting cell proliferation, invasion, and migration | [111] |
| PINT | ALL patient samples and cell lines | tumor suppressor | downregulated | increasing transcription of HMOX1 | inducing apoptosis and causing cell cycle arrest | [112] |
| HOTAIR | ALL patient samples | oncogene | upregulated | increasing EZH2, LSD1, DNMT3A and DNMT3B level | promoting cell proliferation, survival, and migration | [113] |
| HOXA-AS2 | ALL cell lines and patient samples | oncogene | upregulated | enhancing GC resistance via promoting HOXA3 expression | promotion of cell proliferation, inhibition of apoptosis | [114] |
| SNHG16 | ALL cell lines and patient samples | oncogene | upregulated | sponging miR-124-3p | promoting ALL cell proliferation and migration | [115] |
| MALAT1 | ALL samples and cell lines | oncogene | upregulated | sponging miR-205 to increase PTK7 expression | promoting cell proliferation and apoptosis | [116] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maimaitiyiming, Y.; Ye, L.; Yang, T.; Yu, W.; Naranmandura, H. Linear and Circular Long Non-Coding RNAs in Acute Lymphoblastic Leukemia: From Pathogenesis to Classification and Treatment. Int. J. Mol. Sci. 2022, 23, 4442. https://doi.org/10.3390/ijms23084442
Maimaitiyiming Y, Ye L, Yang T, Yu W, Naranmandura H. Linear and Circular Long Non-Coding RNAs in Acute Lymphoblastic Leukemia: From Pathogenesis to Classification and Treatment. International Journal of Molecular Sciences. 2022; 23(8):4442. https://doi.org/10.3390/ijms23084442
Chicago/Turabian StyleMaimaitiyiming, Yasen, Linyan Ye, Tao Yang, Wenjuan Yu, and Hua Naranmandura. 2022. "Linear and Circular Long Non-Coding RNAs in Acute Lymphoblastic Leukemia: From Pathogenesis to Classification and Treatment" International Journal of Molecular Sciences 23, no. 8: 4442. https://doi.org/10.3390/ijms23084442