
Protein Profiling of WERI-RB1 and Etoposide-Resistant WERI-ETOR Reveals New Insights into Topoisomerase Inhibitor Resistance in Retinoblastoma

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
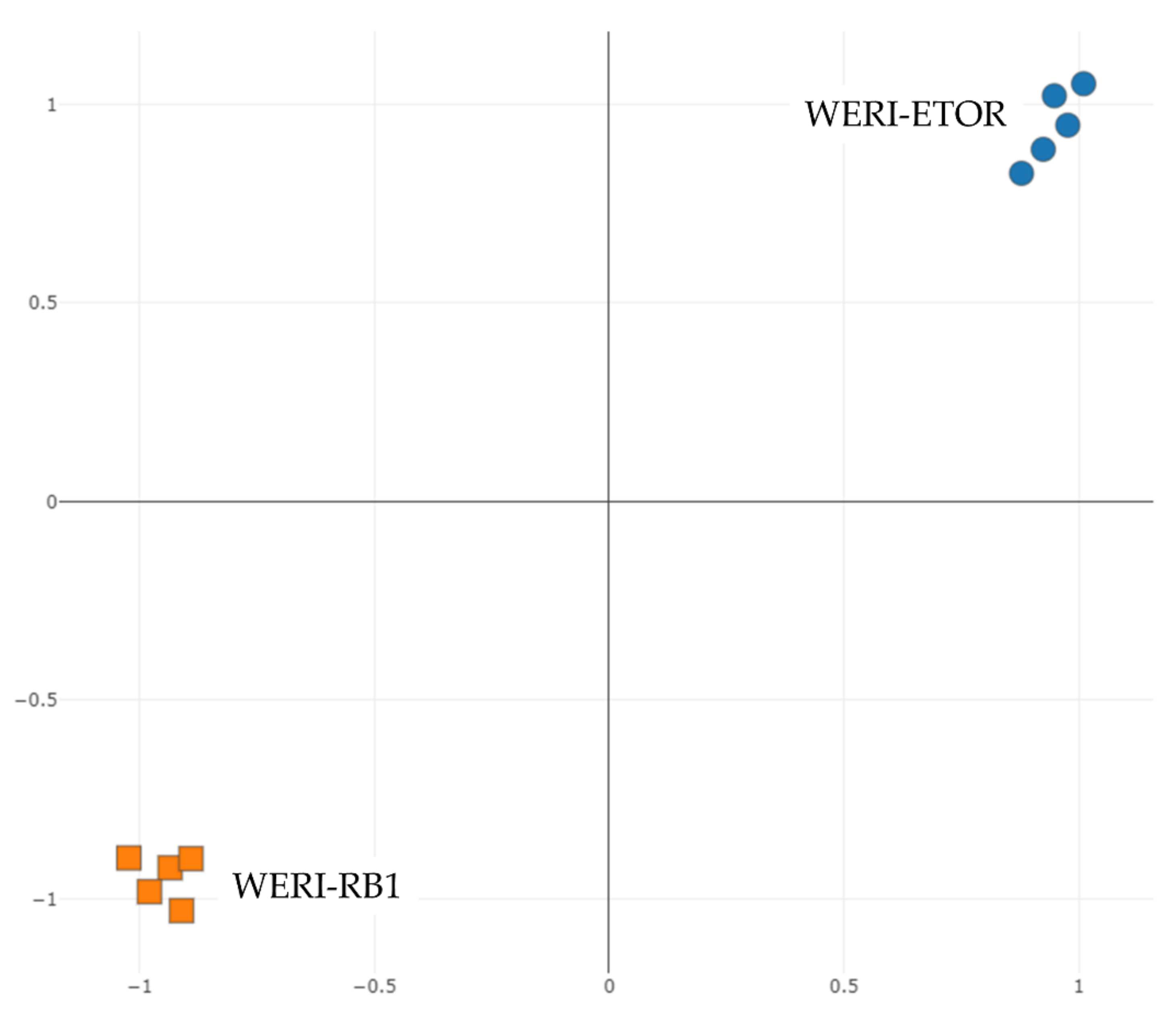
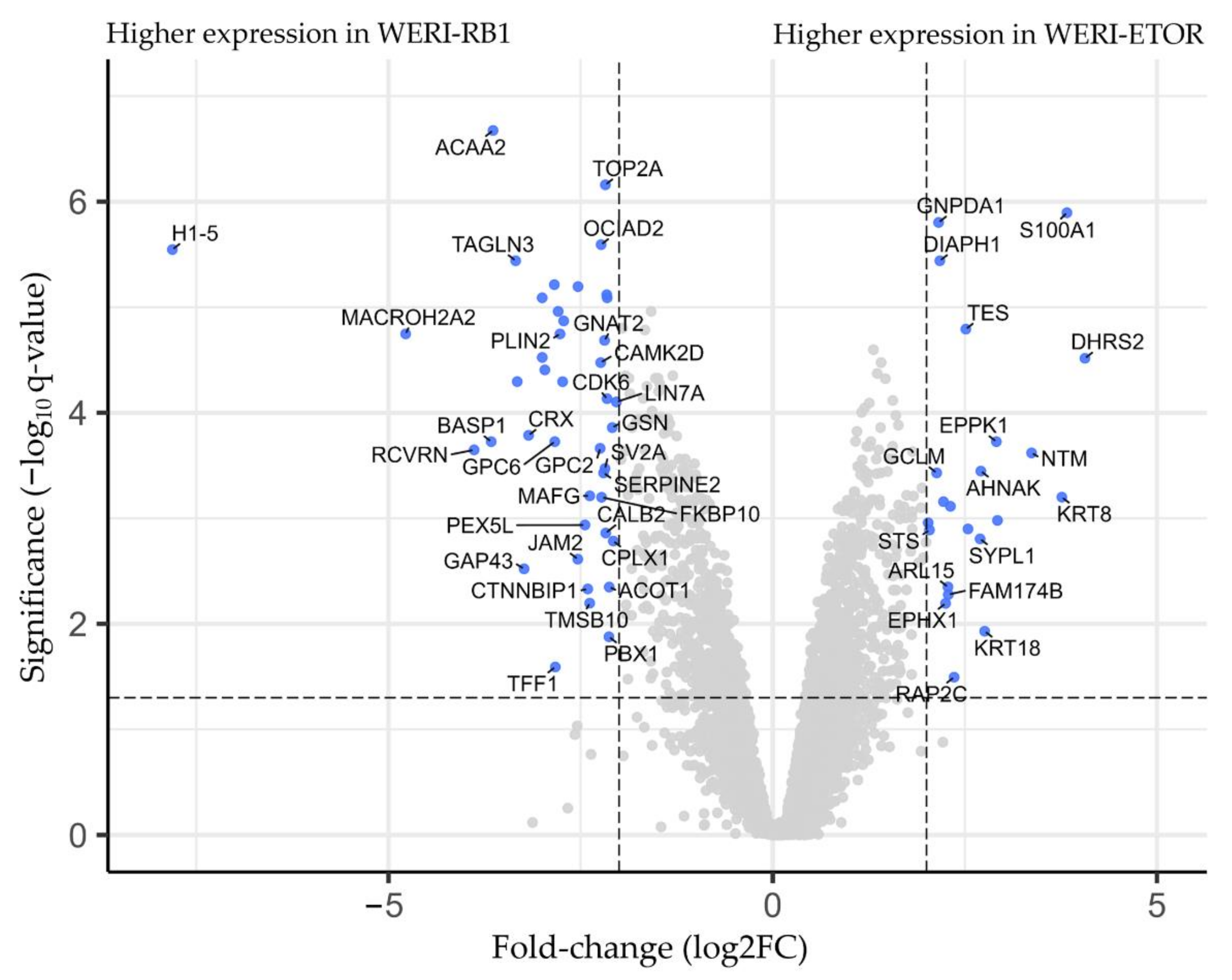
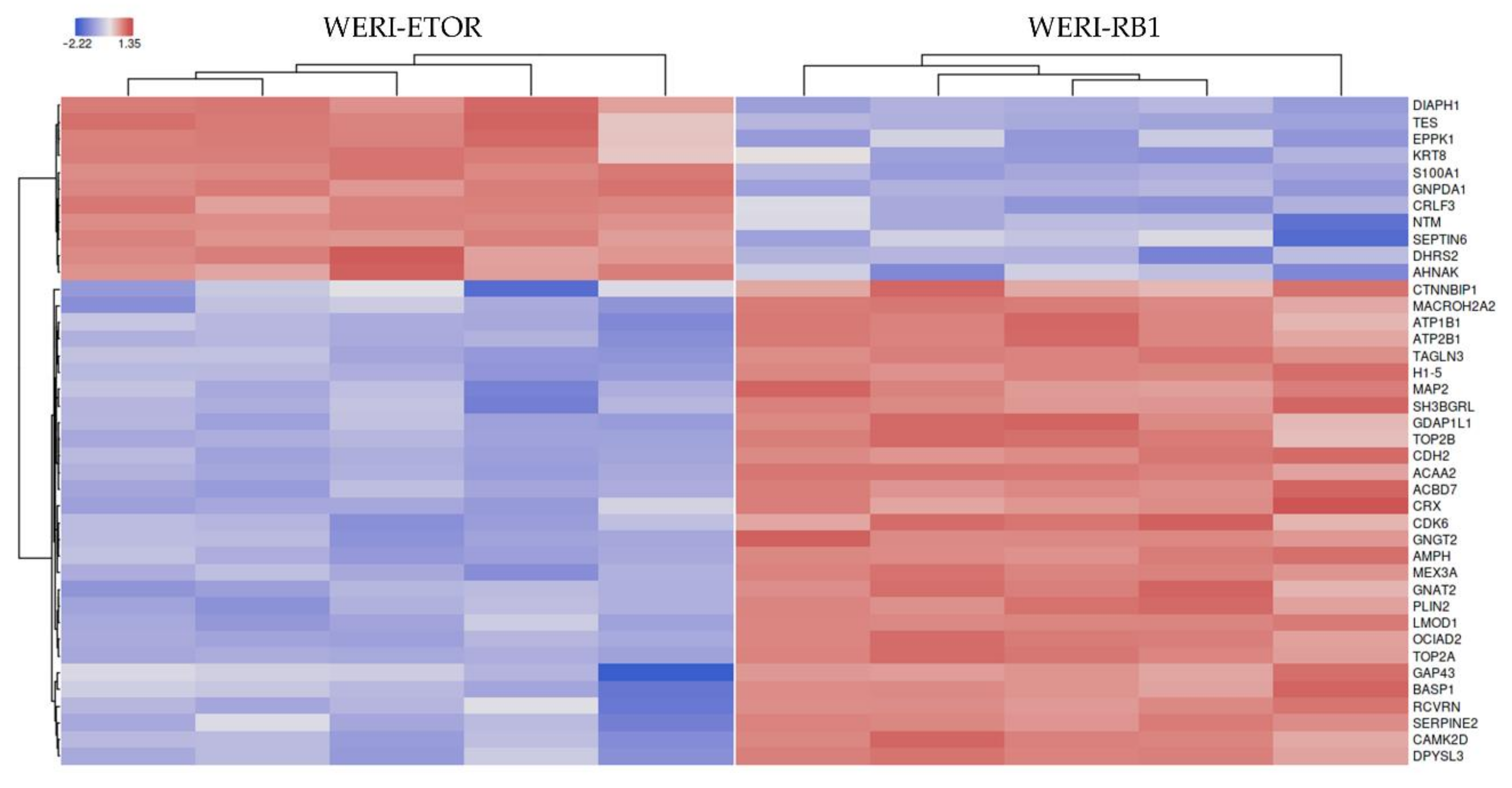
2.1. Protein-Expression Profiles of WERI-RB1 and WERI-ETOR Cell Lines
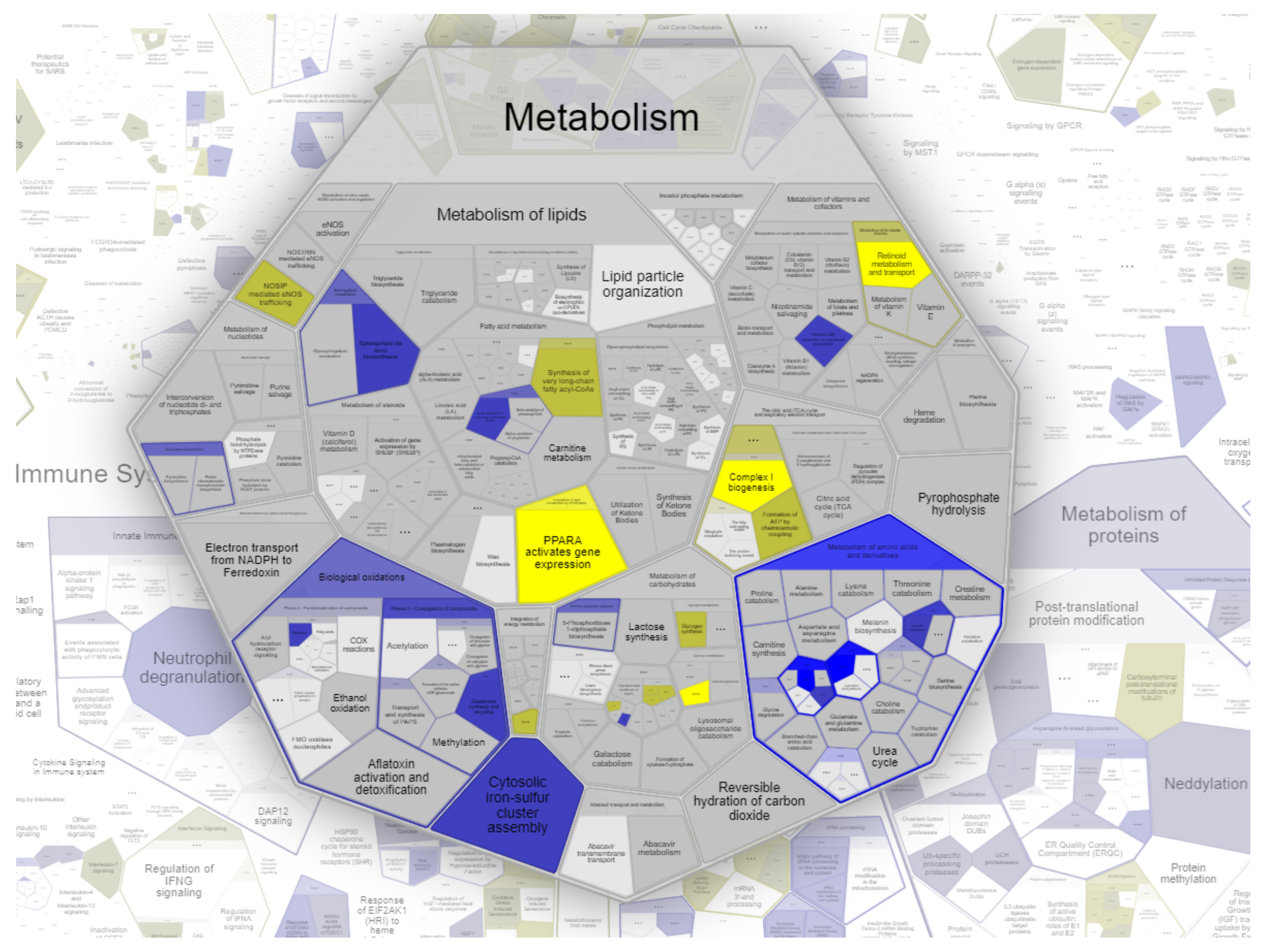
2.2. Enrichment-Pathway Analysis of WERI-RB1 and WERI-ETOR Protein Signatures
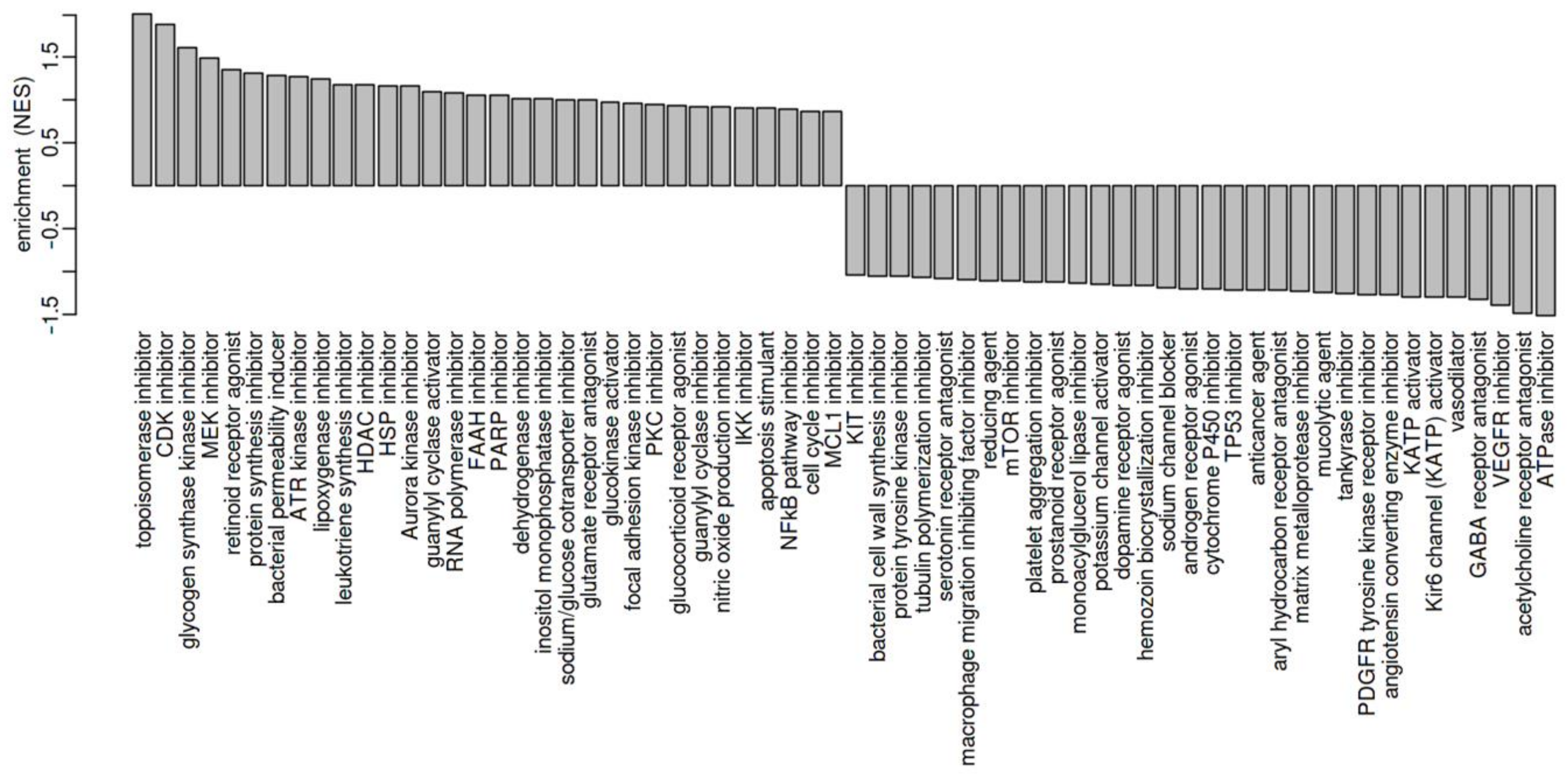
2.3. Drug-Enrichment Analysis of WERI-RB1 and WERI-ETOR Protein Signatures
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Preparation and Mass Spectrometry
4.3. SWATH Data and Quantitative Data Processing
4.4. Data and Statistical Analysis
4.5. Pathway Enrichment and Drug-Connectivity Correlation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimaras, H.; Corson, T.W.; Cobrinik, D.; White, A.; Zhao, J.; Munier, F.L.; Abramson, D.H.; Shields, C.L.; Chantada, G.L.; Njuguna, F.; et al. Retinoblastoma. Nat. Rev. Dis. Primers 2015, 1, 15021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabian, I.D.; Onadim, Z.; Karaa, E.; Duncan, C.; Chowdhury, T.; Scheimberg, I.; Ohnuma, S.I.; Reddy, M.A.; Sagoo, M.S. The management of retinoblastoma. Oncogene 2018, 37, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Kivela, T. The epidemiological challenge of the most frequent eye cancer: Retinoblastoma, an issue of birth and death. Br. J. Ophthalmol. 2009, 93, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Dimaras, H.; Kimani, K.; Dimba, E.A.; Gronsdahl, P.; White, A.; Chan, H.S.; Gallie, B.L. Retinoblastoma. Lancet 2012, 379, 1436–1446. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.; Rojanaporn, D.; Chawla, B.; Sundar, G.; Gopal, L.; Khetan, V. Retinoblastoma in Asia. Eye 2019, 33, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Houston, S.K.; Murray, T.G.; Wolfe, S.Q.; Fernandes, C.E. Current update on retinoblastoma. Int. Ophthalmol. Clin. 2011, 51, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Ward, E.; DeSantis, C.; Robbins, A.; Kohler, B.; Jemal, A. Childhood and adolescent cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 83–103. [Google Scholar] [CrossRef]
- Sanders, B.M.; Draper, G.J.; Kingston, J.E. Retinoblastoma in Great Britain 1969-80: Incidence, treatment, and survival. Br. J. Ophthalmol. 1988, 72, 576–583. [Google Scholar] [CrossRef] [Green Version]
- McFall, R.C.; Sery, T.W.; Makadon, M. Characterization of a new continuous cell line derived from a human retinoblastoma. Cancer Res. 1977, 37, 1003–1010. [Google Scholar]
- McFall, R.C.; Nagy, R.M.; Nagle, B.T.; McGreevy, L.M. Scanning electron microscopic observation of two retinoblastoma cell lines. Cancer Res. 1978, 38, 2827–2835. [Google Scholar]
- Busch, M.; Philippeit, C.; Weise, A.; Dunker, N. Re-characterization of established human retinoblastoma cell lines. Histochem. Cell Biol. 2015, 143, 325–338. [Google Scholar] [CrossRef]
- Stephan, H.; Boeloeni, R.; Eggert, A.; Bornfeld, N.; Schueler, A. Photodynamic therapy in retinoblastoma: Effects of verteporfin on retinoblastoma cell lines. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3158–3163. [Google Scholar] [CrossRef] [Green Version]
- Mergler, S.; Cheng, Y.; Skosyrski, S.; Garreis, F.; Pietrzak, P.; Kociok, N.; Dwarakanath, A.; Reinach, P.S.; Kakkassery, V. Altered calcium regulation by thermosensitive transient receptor potential channels in etoposide-resistant WERI-Rb1 retinoblastoma cells. Exp. Eye Res. 2012, 94, 157–173. [Google Scholar] [CrossRef]
- Kakkassery, V.; Skosyrski, S.; Luth, A.; Kleuser, B.; van der Giet, M.; Tate, R.; Reinhard, J.; Faissner, A.; Joachim, S.C.; Kociok, N. Etoposide Upregulates Survival Favoring Sphingosine-1-Phosphate in Etoposide-Resistant Retinoblastoma Cells. Pathol. Oncol. Res. 2019, 25, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Reinhard, J.; Wagner, N.; Kramer, M.M.; Jarocki, M.; Joachim, S.C.; Dick, H.B.; Faissner, A.; Kakkassery, V. Expression Changes and Impact of the Extracellular Matrix on Etoposide Resistant Human Retinoblastoma Cell Lines. Int. J. Mol. Sci. 2020, 21, 4322. [Google Scholar] [CrossRef]
- Oronowicz, J.; Reinhard, J.; Reinach, P.S.; Ludwiczak, S.; Luo, H.; Omar Ba Salem, M.H.; Kraemer, M.M.; Biebermann, H.; Kakkassery, V.; Mergler, S. Ascorbate-induced oxidative stress mediates TRP channel activation and cytotoxicity in human etoposide-sensitive and -resistant retinoblastoma cells. Lab. Investig. 2021, 101, 70–88. [Google Scholar] [CrossRef]
- Busch, M.; Papior, D.; Stephan, H.; Dunker, N. Characterization of etoposide- and cisplatin-chemoresistant retinoblastoma cell lines. Oncol. Rep. 2018, 39, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Zschoche, M.; Skosyrski, S.; Babst, N.; Ranjbar, M.; Rommel, F.; Kurz, M.; Tura, A.; Joachim, S.C.; Kociok, N.; Kakkassey, V. Islet Co-Expression of CD133 and ABCB5 in Human Retinoblastoma Specimens. Klin. Monbl. Augenheilkd. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Pane, K.; Affinito, O.; Zanfardino, M.; Castaldo, R.; Incoronato, M.; Salvatore, M.; Franzese, M. An Integrative Computational Approach Based on Expression Similarity Signatures to Identify Protein-Protein Interaction Networks in Female-Specific Cancers. Front. Genet. 2020, 11, 612521. [Google Scholar] [CrossRef]
- Infante, T.; Franzese, M.; Ruocco, A.; Schiano, C.; Affinito, O.; Pane, K.; Memoli, D.; Rizzo, F.; Weisz, A.; Bontempo, P.; et al. ABCA1, TCF7, NFATC1, PRKCZ, and PDGFA DNA methylation as potential epigenetic-sensitive targets in acute coronary syndrome via network analysis. Epigenetics 2021, 1–17. [Google Scholar] [CrossRef]
- Chater-Diehl, E.; Goodman, S.J.; Cytrynbaum, C.; Turinsky, A.L.; Choufani, S.; Weksberg, R. Anatomy of DNA methylation signatures: Emerging insights and applications. Am. J. Hum. Genet. 2021, 108, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Van Eyk, J.E.; Snyder, M.P. Precision Medicine: Role of Proteomics in Changing Clinical Management and Care. J. Proteome Res. 2019, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jylha, A.; Nattinen, J.; Aapola, U.; Mikhailova, A.; Nykter, M.; Zhou, L.; Beuerman, R.; Uusitalo, H. Comparison of iTRAQ and SWATH in a clinical study with multiple time points. Clin. Proteom. 2018, 15, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demichev, V.; Messner, C.B.; Vernardis, S.I.; Lilley, K.S.; Ralser, M. DIA-NN: Neural networks and interference correction enable deep proteome coverage in high throughput. Nat. Methods 2020, 17, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Danda, R.; Ganapathy, K.; Sathe, G.; Madugundu, A.K.; Ramachandran, S.; Krishnan, U.M.; Khetan, V.; Rishi, P.; Keshava Prasad, T.S.; Pandey, A.; et al. Proteomic profiling of retinoblastoma by high resolution mass spectrometry. Clin. Proteom. 2016, 13, 29. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Meng, Q.; Huang, L.; Shi, X.; Hou, J.; Li, X.; Liang, J. iTRAQ-based quantitative proteomic analysis and bioinformatics study of proteins in retinoblastoma. Oncol. Lett. 2017, 14, 8084–8091. [Google Scholar] [CrossRef] [Green Version]
- Danda, R.; Ganapathy, K.; Sathe, G.; Madugundu, A.K.; Krishnan, U.M.; Khetan, V.; Rishi, P.; Gowda, H.; Pandey, A.; Subramanian, K.; et al. Membrane Proteome of Invasive Retinoblastoma: Differential Proteins and Biomarkers. Proteom. Clin. Appl. 2018, 12, e1700101. [Google Scholar] [CrossRef]
- Naru, J.; Aggarwal, R.; Mohanty, A.K.; Singh, U.; Bansal, D.; Kakkar, N.; Agnihotri, N. Identification of differentially expressed proteins in retinoblastoma tumors using mass spectrometry-based comparative proteomic approach. J. Proteom. 2017, 159, 77–91. [Google Scholar] [CrossRef]
- Galardi, A.; Colletti, M.; Lavarello, C.; Di Paolo, V.; Mascio, P.; Russo, I.; Cozza, R.; Romanzo, A.; Valente, P.; De Vito, R.; et al. Proteomic Profiling of Retinoblastoma-Derived Exosomes Reveals Potential Biomarkers of Vitreous Seeding. Cancers 2020, 12, 1555. [Google Scholar] [CrossRef]
- Orlandella, F.M.; Mariniello, R.M.; Iervolino, P.L.C.; Auletta, L.; De Stefano, A.E.; Ugolini, C.; Greco, A.; Mirabelli, P.; Pane, K.; Franzese, M.; et al. Junctional adhesion molecule-A is down-regulated in anaplastic thyroid carcinomas and reduces cancer cell aggressiveness by modulating p53 and GSK3 alpha/beta pathways. Mol. Carcinog. 2019, 58, 1181–1193. [Google Scholar] [CrossRef]
- Nicoud, M.; Kong, J.; Iqball, S.; Kan, O.; Naylor, S.; Gouras, P.; Allikmets, R.; Binley, K. Development of photoreceptor-specific promoters and their utility to investigate EIAV lentiviral vector mediated gene transfer to photoreceptors. J. Gene Med. 2007, 9, 1015–1023. [Google Scholar] [CrossRef]
- Vene, R.; Arena, G.; Poggi, A.; D’Arrigo, C.; Mormino, M.; Noonan, D.M.; Albini, A.; Tosetti, F. Novel cell death pathways induced by N-(4-hydroxyphenyl)retinamide: Therapeutic implications. Mol. Cancer Ther. 2007, 6, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Khanna, H.; Akimoto, M.; Siffroi-Fernandez, S.; Friedman, J.S.; Hicks, D.; Swaroop, A. Retinoic acid regulates the expression of photoreceptor transcription factor NRL. J. Biol. Chem. 2006, 281, 27327–27334. [Google Scholar] [CrossRef] [Green Version]
- Kyritsis, A.; Joseph, G.; Chader, G.J. Effects of butyrate, retinol, and retinoic acid on human Y-79 retinoblastoma cells growing in monolayer cultures. J. Natl. Cancer Inst. 1984, 73, 649–654. [Google Scholar]
- Theocharis, A.D.; Skandalis, S.S.; Tzanakakis, G.N.; Karamanos, N.K. Proteoglycans in health and disease: Novel roles for proteoglycans in malignancy and their pharmacological targeting. FEBS J. 2010, 277, 3904–3923. [Google Scholar] [CrossRef]
- Faissner, A.; Reinhard, J. The extracellular matrix compartment of neural stem and glial progenitor cells. Glia 2015, 63, 1330–1349. [Google Scholar] [CrossRef]
- Reinhard, J.; Brosicke, N.; Theocharidis, U.; Faissner, A. The extracellular matrix niche microenvironment of neural and cancer stem cells in the brain. Int. J. Biochem. Cell Biol. 2016, 81, 174–183. [Google Scholar] [CrossRef]
- Corti, F.; Wang, Y.; Rhodes, J.M.; Atri, D.; Archer-Hartmann, S.; Zhang, J.; Zhuang, Z.W.; Chen, D.; Wang, T.; Wang, Z.; et al. N-terminal syndecan-2 domain selectively enhances 6-O heparan sulfate chains sulfation and promotes VEGFA165-dependent neovascularization. Nat. Commun. 2019, 10, 1562. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, N.; Cheng, H. Fisetin inhibits vascular endothelial growth factor-induced angiogenesis in retinoblastoma cells. Oncol. Lett. 2020, 20, 1239–1244. [Google Scholar] [CrossRef]
- Abu El-Asrar, A.M.; Nawaz, M.I.; De Hertogh, G.; Alam, K.; Siddiquei, M.M.; Van den Eynde, K.; Mousa, A.; Mohammad, G.; Geboes, K.; Opdenakker, G. S100A4 is upregulated in proliferative diabetic retinopathy and correlates with markers of angiogenesis and fibrogenesis. Mol. Vis. 2014, 20, 1209–1224. [Google Scholar]
- Lau, C.S.; Yu, C.B.; Wong, H.K.; Fan, D.S.; Mak, H.T.; Wong, K.W.; Lam, D.S.; Pang, C.P.; Choy, K.W. Allelic imbalance at 13q31 is associated with reduced GPC6 in Chinese with sporadic retinoblastoma. Br. J. Ophthalmol. 2010, 94, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Kim, M.J.; Park, J.; Kim, J.; Kim, J.Y.; An, M.J.; Shin, G.S.; Lee, H.M.; Kim, J.W. Bisphenol A Exposure Changes the Transcriptomic and Proteomic Dynamics of Human Retinoblastoma Y79 Cells. Genes 2021, 12, 264. [Google Scholar] [CrossRef] [PubMed]
- Sauer, T.; Facchinetti, G.; Kohl, M.; Kowal, J.M.; Rozanova, S.; Horn, J.; Schmal, H.; Kwee, I.; Schulz, A.P.; Hartwig, S.; et al. Protein Expression of AEBP1, MCM4, and FABP4 Differentiate Osteogenic, Adipogenic, and Mesenchymal Stromal Stem Cells. Int. J. Mol. Sci. 2022, 23, 2568. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Akhmedov, M.; Martinelli, A.; Geiger, R.; Kwee, I. Omics Playground: A comprehensive self-service platform for visualization, analytics and exploration of Big Omics Data. NAR Genom. Bioinform. 2020, 2, lqz019. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization Paths for Generalized Linear Models via Coordinate Descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.; Hastie, T. Regularization and variable selection via the elastic net. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2005, 67, 301–320. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; pp. 785–794. [Google Scholar]
- Griss, J.; Viteri, G.; Sidiropoulos, K.; Nguyen, V.; Fabregat, A.; Hermjakob, H. ReactomeGSA—Efficient Multi-Omics Comparative Pathway Analysis. Mol. Cell Proteom. 2020, 19, 2115–2125. [Google Scholar] [CrossRef]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e17. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Kakkassery, V.; Gemoll, T.; Krämer, M.M.; Sauer, T.; Tura, A.; Ranjbar, M.; Grisanti, S.; Joachim, S.C.; Mergler, S.; Reinhard, J. Protein profiling of WERI RB1 and etoposide resistant WERI ETOR reveals new insights into topoisomerase inhibitor resistance in retinoblastoma. BioRxiv 2022. preprint. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Full Protein Name | Chromosome | Meta.q-Value | Log Fold-Change [WERI-ETOR/WERI-RB1] |
|---|---|---|---|---|
| DHRS2 | Dehydrogenase/reductase SDR family member 2 | 14 | 3.051 × 10−5 | 4.062 |
| S100A1 | Protein S100-A1 | 1 | 1.275 × 10−6 | 3.828 |
| KRT8 | Keratin 8 | 12 | 6.307 × 10−4 | 3.762 |
| NTM | NAC domain-containing protein 69 | 11 | 2.402 × 10−4 | 3.368 |
| EPPK1 | Epiplakin | 8 | 1.879 × 10−4 | 2.910 |
| AHNAK | AHNAK nucleoprotein | 11 | 3.570 × 10−4 | 2.708 |
| SEPTIN6 | Septin 6 | X | 1.262 × 10−4 | 2.540 |
| TES | Testin | 7 | 1.615 × 10−5 | 2.513 |
| DIAPH1 | Protein diaphanous homolog 1 | 5 | 3.634 × 10−6 | 2.174 |
| GNPDA1 | Glucosamine-6-phosphate isomerase 1 | 5 | 1.576 × 10−6 | 2.158 |
| CRLF3 | Cytokine receptor like factor 3 | 17 | 5.155 × 10−4 | 1.942 |
| MEX3A | RNA-binding protein MEX3A | 1 | 4.712 × 10−5 | −1.871 |
| CDH2 | Cadherin-2 | 18 | 1.581 × 10−5 | −1.955 |
| TOP2B | DNA topoisomerase 2-beta | 3 | 8.151 × 10−6 | −2.155 |
| CDK6 | Cyclin-dependent kinase 6 | 7 | 7.362 × 10−5 | −2.156 |
| ATP2B1 | Plasma membrane calcium-transporting ATPase 1 | 12 | 7.630 × 10−6 | −2.160 |
| TOP2A | DNA topoisomerase 2-alpha | 17 | 6.932 × 10−7 | −2.178 |
| GNAT2 | Guanine nucleotide-binding protein G(t) subunit alpha-2 | 1 | 2.064 × 10−5 | −2.189 |
| SERPINE2 | Serpin family E member 2 | 2 | 3.716 × 10−4 | −2.201 |
| OCIAD2 | OCIA domain-containing protein 2 | 4 | 2.556 × 10−6 | −2.234 |
| CAMK2D | Calcium-dependent protein kinase II | 4 | 3.350 × 10−5 | −2.238 |
| CTNNBIP1 | Catenin-beta-interacting protein 1 | 1 | 4.668 × 10−3 | −2.408 |
| LMOD1 | Leiomodin-1 | 1 | 6.385 × 10−6 | −2.534 |
| ACBD7 | Acyl-CoA-binding domain-containing protein 7 | 10 | 1.349 × 10−5 | −2.720 |
| GDAP1L1 | Ganglioside-induced differentiation-associated protein 1-like 1 | 20 | 5.081 × 10−5 | −2.734 |
| PLIN2 | Perilipin-2 | 9 | 1.793 × 10−5 | −2.769 |
| DPYSL3 | Dihydropyrimidinase like 3 | 5 | 1.096 × 10−5 | −2.792 |
| AMPH | D-alanyl-D-alanine-carboxypeptidase/endopeptidase AmpH | 7 | 6.121 × 10−6 | −2.842 |
| MAP2 | Microtubule-associated protein 2 | 2 | 3.928 × 10−5 | −2.966 |
| ATP1B1 | Sodium/potassium-transporting ATPase subunit beta-1 | 1 | 2.994 × 10−5 | −3.000 |
| GNGT2 | Guanine nucleotide-binding protein subunit gamma-T2 | 17 | 8.151 × 10−6 | −3.000 |
| CRX | Cone-rod homeobox protein | 19 | 1.636 × 10−4 | −3.177 |
| GAP43 | Neuromodulin | 3 | 3.011 × 10−3 | −3.236 |
| SH3BGRL | SH3 domain-binding glutamic acid-rich-like protein 3 | X | 5.071 × 10−5 | −3.325 |
| TAGLN3 | Transgelin-3 | 3 | 3.634 × 10−6 | −3.346 |
| ACAA2 | 3-ketoacyl-CoA thiolase | 18 | 2.119 × 10−7 | −3.640 |
| BASP1 | Brain-acid-soluble protein 1 | 5 | 1.879 × 10−4 | −3.664 |
| RCVRN | Recoverin | 17 | 2.248 × 10−4 | −3.886 |
| MACROH2A2 | Core histone macro-H2A.2 | 10 | 1.791 × 10−5 | −4.777 |
| H1-5 | Histone H1.5 | 6 | 2.846 × 10−6 | −7.812 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakkassery, V.; Gemoll, T.; Kraemer, M.M.; Sauer, T.; Tura, A.; Ranjbar, M.; Grisanti, S.; Joachim, S.C.; Mergler, S.; Reinhard, J. Protein Profiling of WERI-RB1 and Etoposide-Resistant WERI-ETOR Reveals New Insights into Topoisomerase Inhibitor Resistance in Retinoblastoma. Int. J. Mol. Sci. 2022, 23, 4058. https://doi.org/10.3390/ijms23074058
Kakkassery V, Gemoll T, Kraemer MM, Sauer T, Tura A, Ranjbar M, Grisanti S, Joachim SC, Mergler S, Reinhard J. Protein Profiling of WERI-RB1 and Etoposide-Resistant WERI-ETOR Reveals New Insights into Topoisomerase Inhibitor Resistance in Retinoblastoma. International Journal of Molecular Sciences. 2022; 23(7):4058. https://doi.org/10.3390/ijms23074058
Chicago/Turabian StyleKakkassery, Vinodh, Timo Gemoll, Miriam M. Kraemer, Thorben Sauer, Aysegül Tura, Mahdy Ranjbar, Salvatore Grisanti, Stephanie C. Joachim, Stefan Mergler, and Jacqueline Reinhard. 2022. "Protein Profiling of WERI-RB1 and Etoposide-Resistant WERI-ETOR Reveals New Insights into Topoisomerase Inhibitor Resistance in Retinoblastoma" International Journal of Molecular Sciences 23, no. 7: 4058. https://doi.org/10.3390/ijms23074058