
Nanodosimetric Calculations of Radiation-Induced DNA Damage in a New Nucleus Geometrical Model Based on the Isochore Theory

 , , and
, , and
Abstract
:1. Introduction
2. Results
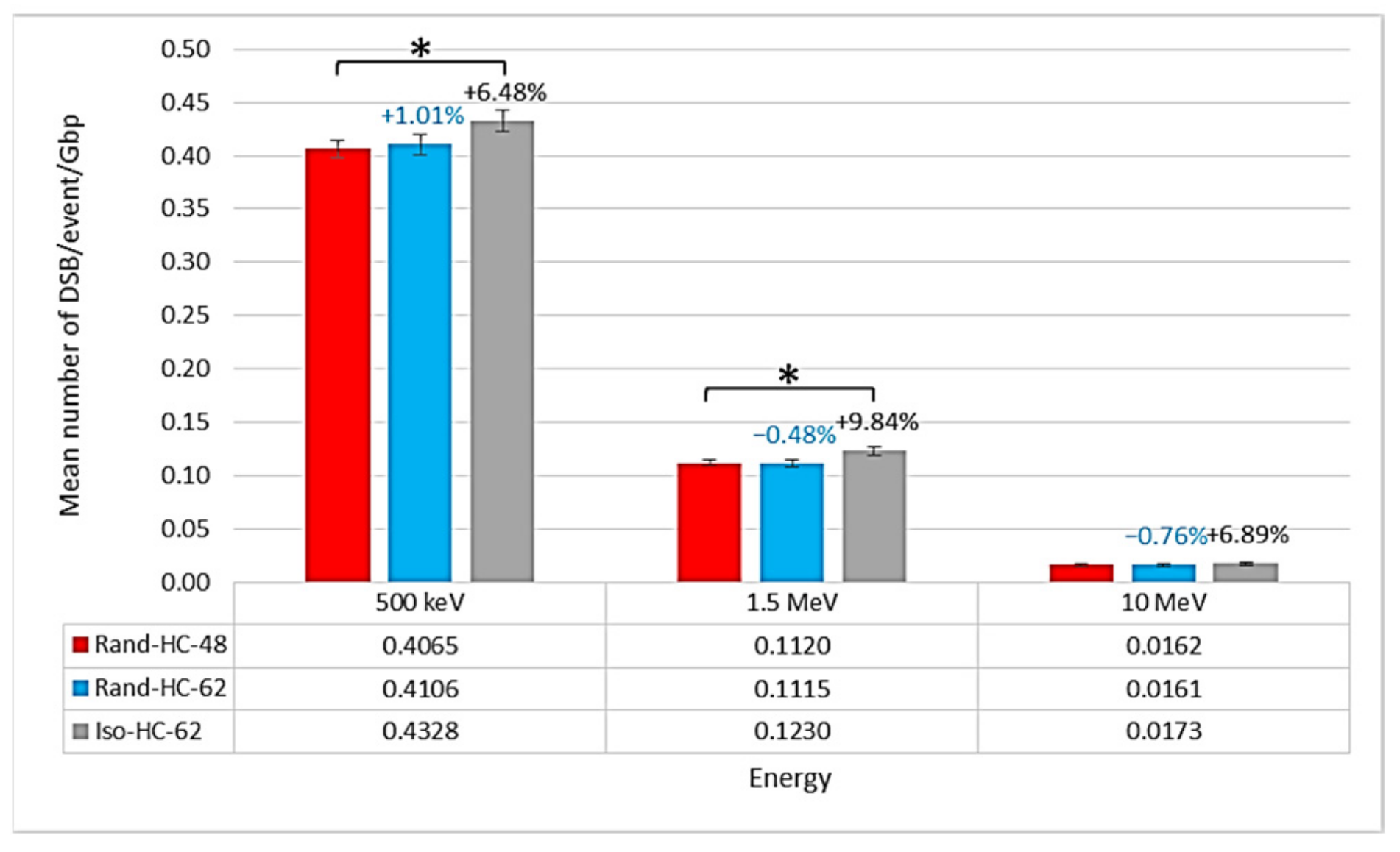
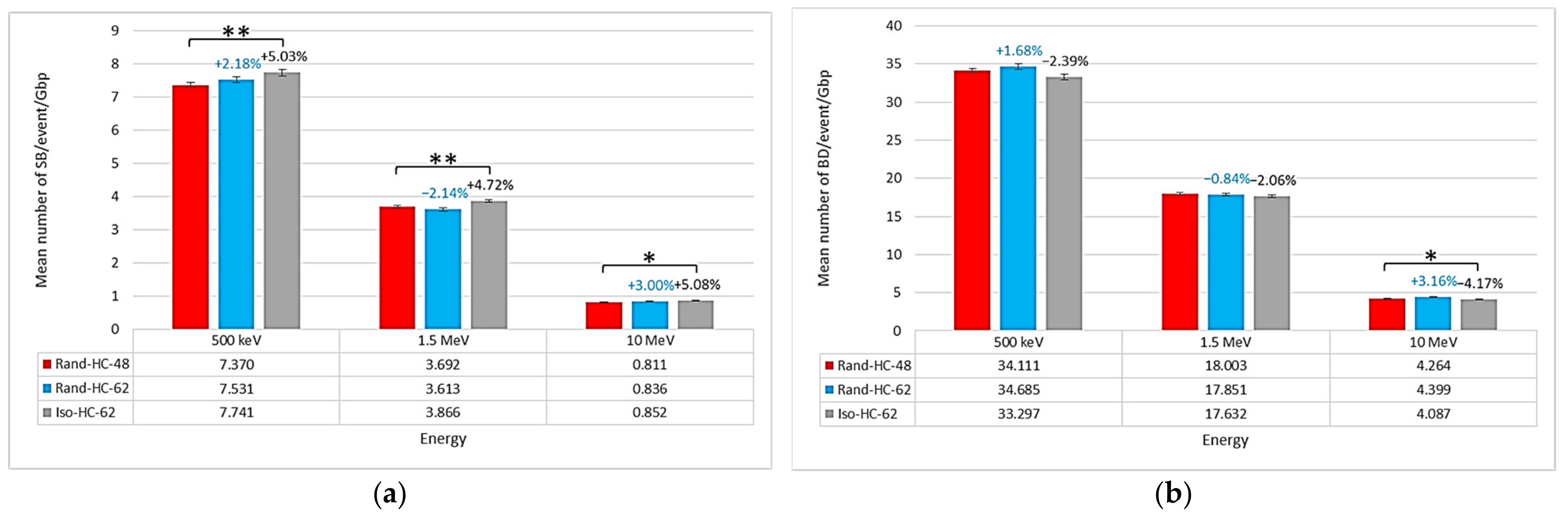
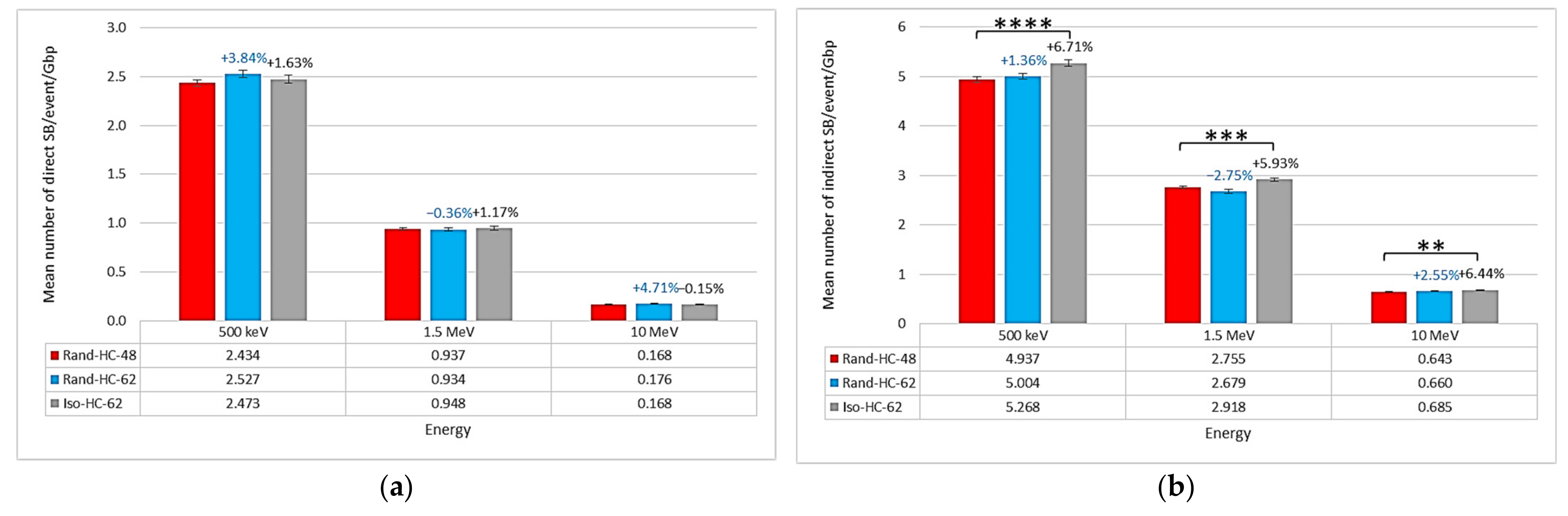
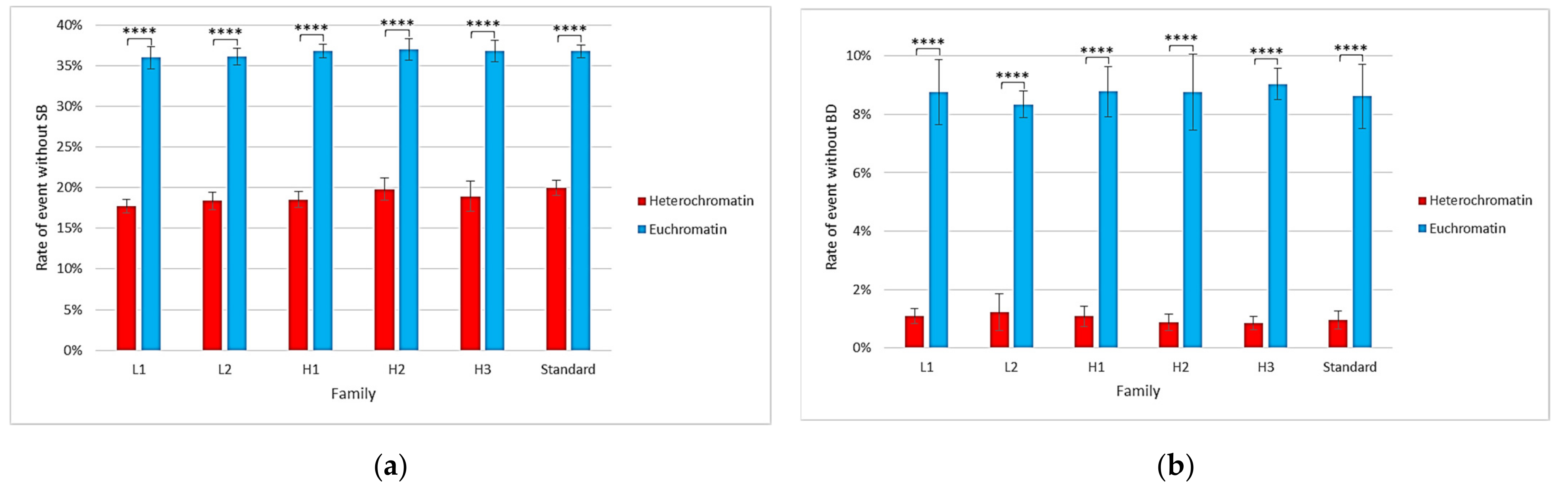
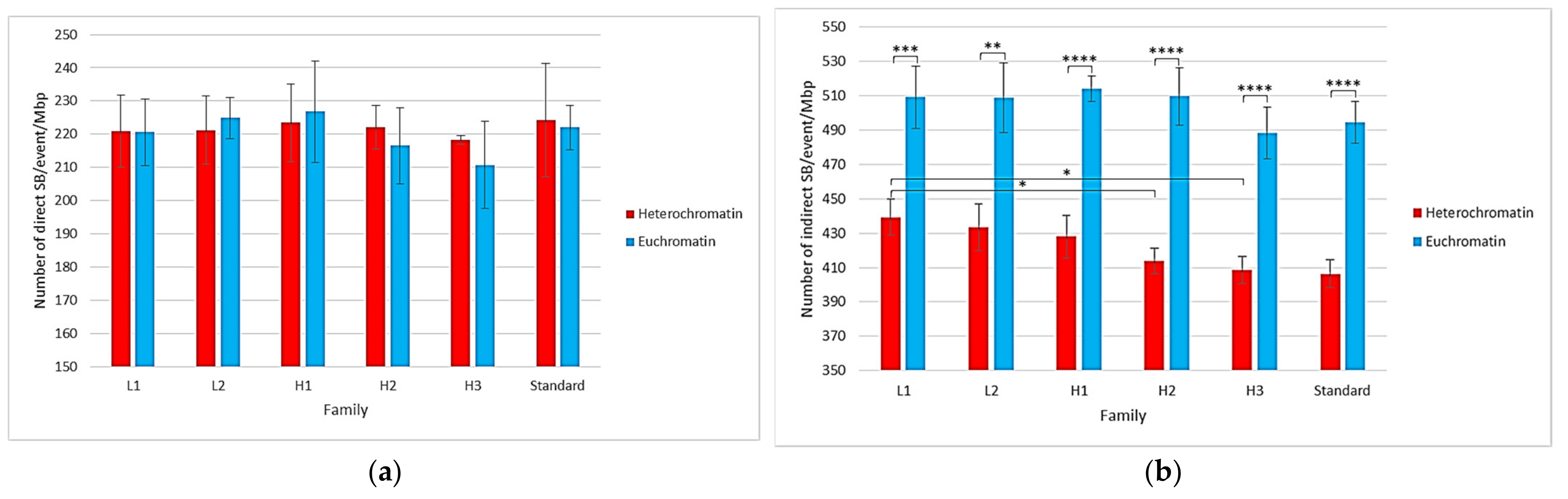
2.1. Impact of the New Geometries on Clustered and Non-Clustered Damages
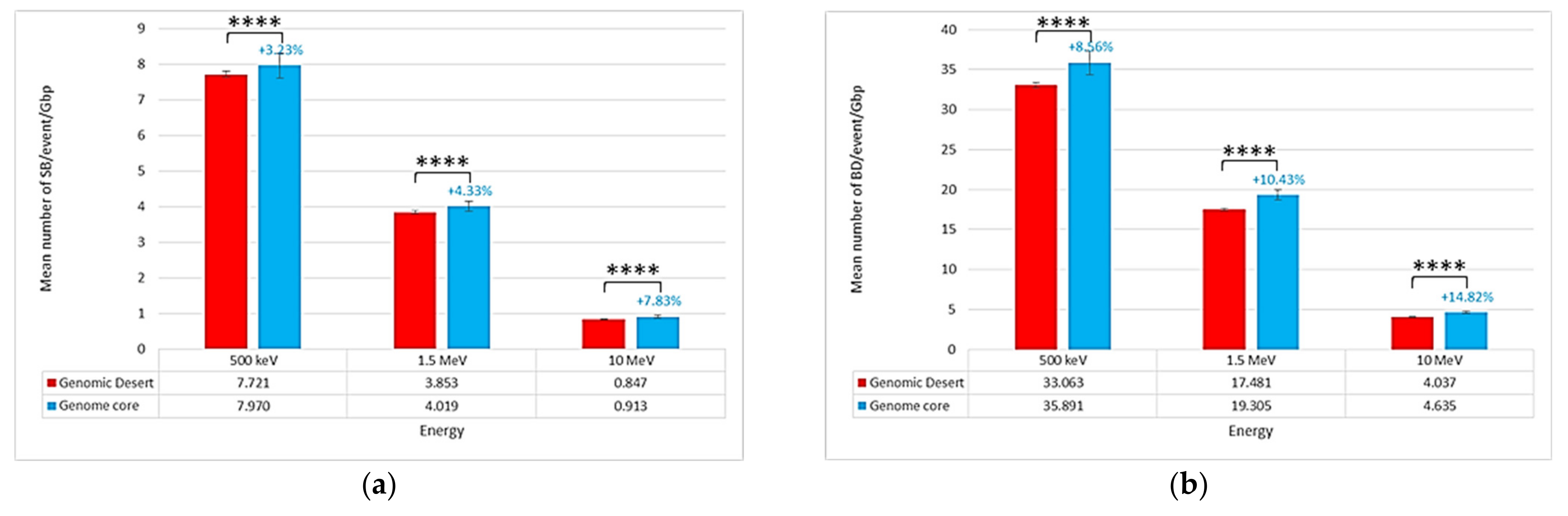
2.2. Location of Damages in the Isochore Nucleus
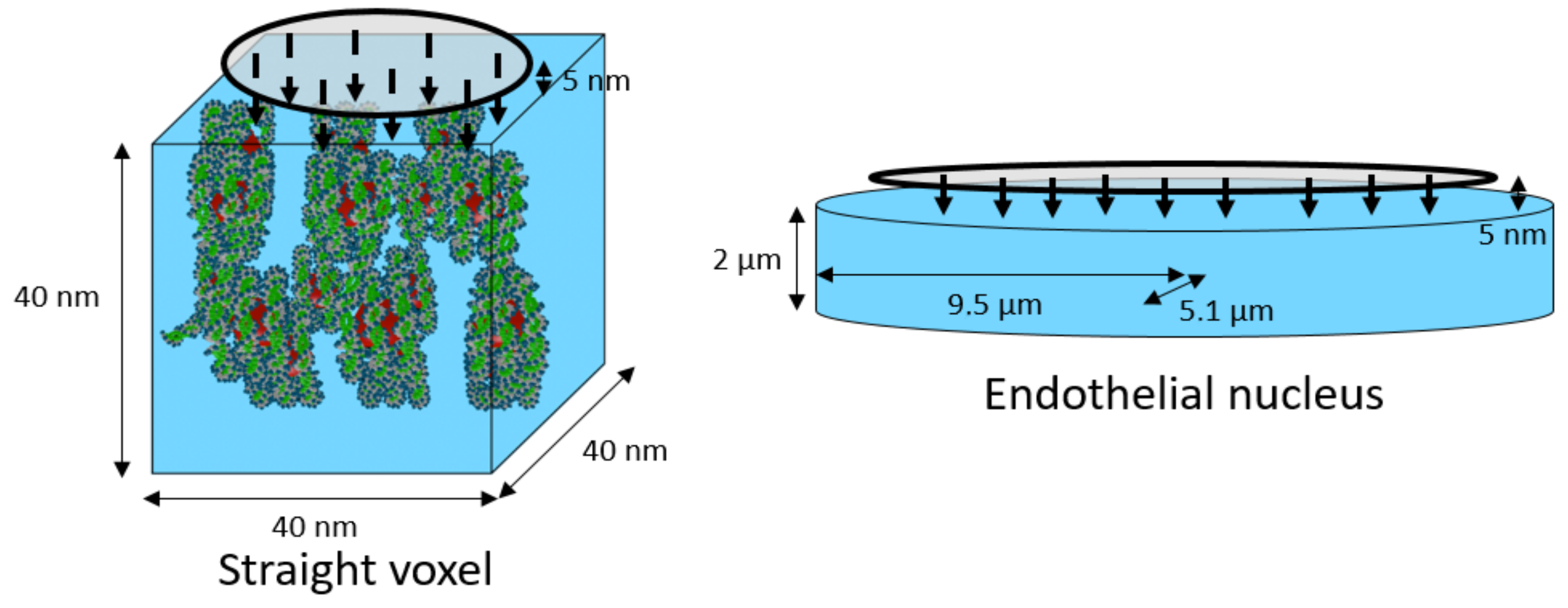
2.3. Single-Voxel Study
3. Discussion
4. Materials and Methods
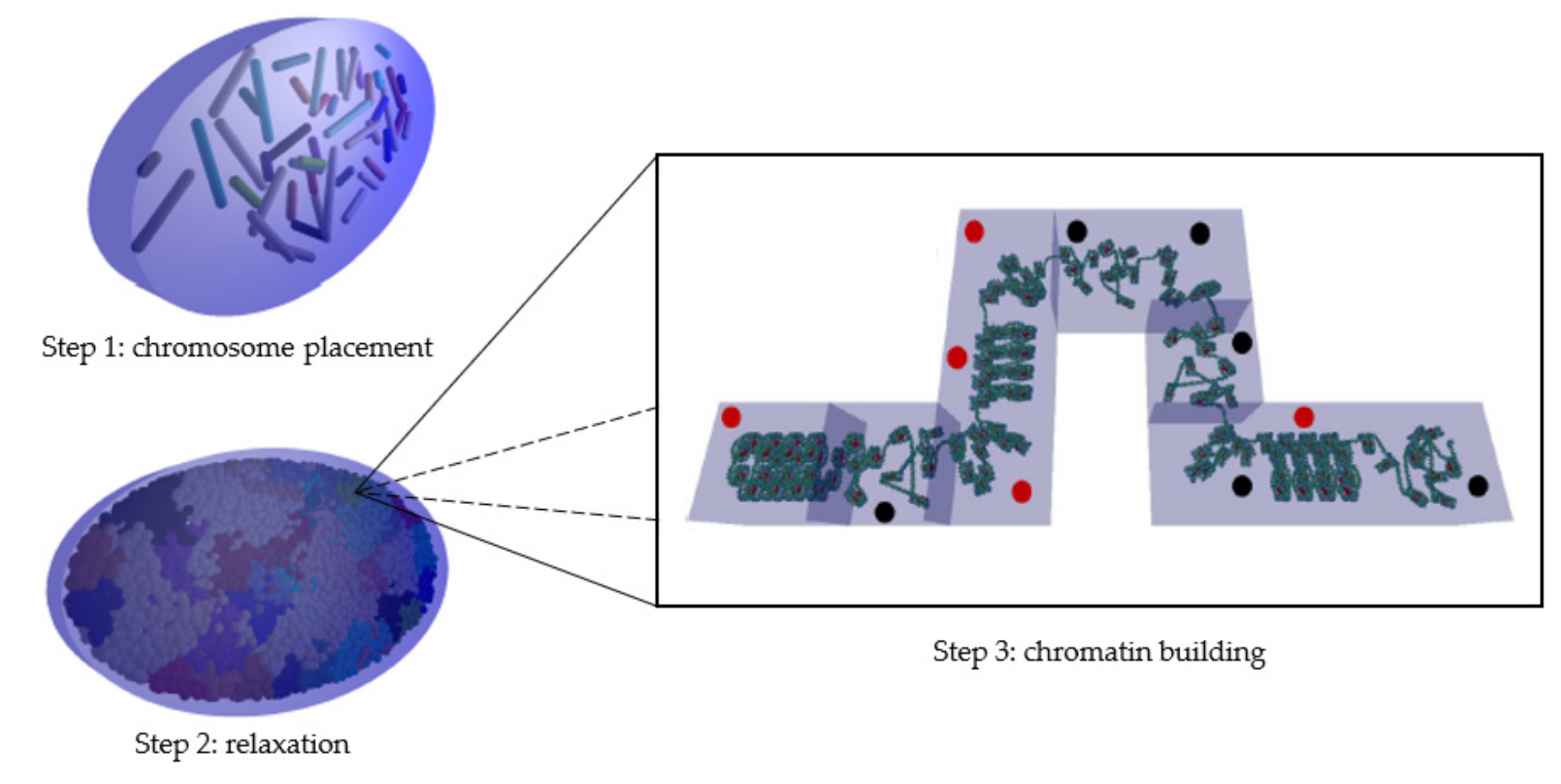
4.1. Geometry Modeling Tool
- The first step consists of placing the 46 chromosomes in a condensed form (not biological; for more information, please see [6]) at their preferred position within the nucleus.
- The second step expands these condensed chromosomes into connected spherical domains of 1 Mbp to occupy maximum space in the nucleus [46].
- The third step consists of building the chromatin fiber placing one by one the cubic voxels (40 nm sides) in the spherical domains and ensuring that they are continuously connected with each other.

4.2. New Model Geometry Based on the Isochore Theory
4.3. Damage Calculation
4.4. Configurations, Cell Nucleus Geometries, and Simulation Parameters
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khanna, K.; Jackson, S. DNA double-strand breaks: Signaling, repair and the cancer connection. Nat. Genet. 2001, 27, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Rothkamm, K.; Lobrich, M. Misrepair of radiation-induced DNA double-strand breaks and its relevance for tumorigenesis and cancer treatment. Int. J. Oncol. 2002, 21, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Friedland, W.; Jacob, P.; Bernhardt, P.; Paretzke, H.; Dingfelder, M. Simulation of DNA Damage after Proton Irradiatin. Rad. Res. 2003, 159, 401–410. [Google Scholar] [CrossRef]
- Uehara, S.; Nikjoo, H.; Goodhead, D.T. Cross-sections for water vapour for the Monte Carlo electron track structure code from 10 eV to the MeV region. Phys. Med. Biol. 1993, 38, 1841–1858. [Google Scholar] [CrossRef]
- Lampe, N.; Karamitros, M.; Breton, V.; Brown, J.M.C.; Sakata, D.; Sarramia, D.; Incerti, S. Mechanistic DNA damage simulations in Geant4-DNA Part 1: A parameter study in a simplified geometry. Phys. Med. 2018, 48, 135–145. [Google Scholar] [CrossRef]
- Meylan, S.; Vimont, U.; Incerti, S.; Clairand, I.; Villagrasa, C. Geant4-DNA simulations using complex DNA geometries generated by the DnaFabric tool. Comput. Phys. Commun. 2016, 204, 159–169. [Google Scholar] [CrossRef]
- Costes, S.; Chiolo, I.; Pluth, J.; Barcellos-Hoff, M.; Jakob, B. Spatiotemporal characterization of ionizing radiation induced DNA damage foci and their relation to chromatin organization. Mutat. Res.-Rev. Mutat. 2010, 704, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Falk, M.; Lukasova, E.; Kozubek, S. Higher-order chromatin structure in DSB induction, repair and misrepair. Mutat. Res.-Rev. Mutat. 2010, 704, 88–100. [Google Scholar] [CrossRef]
- Lafon-Hughes, L.; Di Tomaso, M.; Liddle, P.; Toledo, A.; Reyes-Ábalos, A.; Folle, G. Preferential localization of γH2AX foci in euchromatin of retina rod cells after DNA damage induction. Chromosome Res. 2013, 21, 789–803. [Google Scholar] [CrossRef]
- Magnander, K.; Hultborn, R.; Claesson, K.; Elmroth, K. Clustered DNA damage in irradiated human diploid fibroblasts: Influence of chromatin organization. Rad. Res. 2009, 173, 272–282. [Google Scholar] [CrossRef]
- Murray, J.; Stiff, T.; Jeggo, P. DNA double-strand break repair within heterochromatic regions. Biochem. Soc. Trans. 2012, 10, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Radulescu, I.; Elmroth, K.; Stenerlöw, B. Chromatin organization contributes to non-randomly distributed Double-Strand Breaks after exposure to high-LET radiation. Rad. Res. 2004, 161, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Storch, K.; Eke, I.; Borgmann, K.; Krause, M.; Richter, C.; Becker, K.; Schröck, E.; Cordes, N. Three-dimensional cell growth confers radioresistance by chromatin density modification. Cancer Res. 2010, 70, 3925–3934. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, P.; Panyutin, I.V.; Remeeva, E.; Neumann, R.; Panyutin, I.G. Effect of chromatin structure on the extent and distribution of DNA Double Strand Breaks produced by ionizing radiation; Comparative study of hESC and differentiated cells lines. Int. J. Mol. Sci. 2016, 17, 58–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olins, D.; Olins, A. Chromatin history: Our view from the bridge. Nat. Rev. Mol. Cell Biol. 2003, 4, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Reinberg, D. Facultative heterochromatin: Is there a distinctive molecular signature? Mol. Cell 2007, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gross, D. Heterochromatin: Dark matter or variation on a theme? Curr. Biol. 2015, 25, R462–R465. [Google Scholar] [CrossRef] [Green Version]
- Grewal, S.; Jia, S. Heterochromatin revisited. Nat. Rev. Genet. 2007, 8, 35–46. [Google Scholar] [CrossRef]
- Friedland, W.; Kundrát, P. Track structure based modelling of chromosome aberrations after photon and alpha-particle irradiation. Mutat. Res.-Genet. Toxicol. Environ. Mutagen. 2013, 756, 213–223. [Google Scholar] [CrossRef]
- Tang, N.; Bueno, M.; Meylan, S.; Incerti, S.; Tran, H.N.; Vaurijoux, A.; Gruel, G.; Villagrasa, C. Influence of chromatin compaction on simulated early radiation-induced DNA damage using Geant4-DNA. Med. Phys. 2019, 46, 1501–1511. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, G. The vertebrate genome: Isochores and evolution. Mol. Biol. Evol. 1993, 10, 186–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekrutenko, A.; Li, W. Assessment of compositional heterogeneity within and between eukaryotic genomes. Genome Res. 2000, 10, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Dagan, T.; Stone, L.; Graur, D. GC composition of the human genome: In search of isochores. Mol. Biol. Evol. 2005, 22, 1260–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häring, D.; Kypr, J. No Isochores in the human chromosomes 21 and 22? Biochem. Biophys. Res. Commun. 2001, 280, 567–573. [Google Scholar] [CrossRef]
- Bernardi, G. Misunderstandings about isochores. Part 1. Gene 2001, 276, 3–16. [Google Scholar] [CrossRef]
- Clay, O.; Bernardi, G. How Not to Search for Isochores: A Reply to Cohen et al. Mol. Biol. Evol. 2005, 22, 2315–2317. [Google Scholar] [CrossRef] [Green Version]
- Elhaik, E.; Graur, D.; Josić, K. Comparative testing of DNA segmentation algorithms using benchmark simulations. Mol. Biol. Evol. 2010, 27, 1015–1024. [Google Scholar] [CrossRef]
- Pačes, J.; Zíka, R.; Pačes, V.; Pavlíček, A.; Clay, O.; Bernardi, G. Representing GC variation along eukaryotic chromosomes. Gene 2004, 333, 135–141. [Google Scholar] [CrossRef]
- Costantini, M.; Clay, O.; Auletta, F.; Bernardi, G. An isochore map of human chromosomes. Genome Res. 2006, 16, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Costantini, M.; Musto, H. The isochores as a fundamental level of genome structure and organization: A General Overview. J. Mol. Evol. 2017, 84, 93–103. [Google Scholar] [CrossRef]
- Meylan, S.; Incerti, S.; Karamitros, M.; Tang, N.; Bueno, M.; Clairand, I.; Villagrasa, C. Simulation of early DNA damage after the irradiation of a fibroblast cell nucleus using Geant4-DNA. Sci. Rep. 2017, 7, 11923. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Bueno, M.; Meylan, S.; Perrot, Y.; Tran, H.; Freneau, A.; Dos Santos, M.; Vaurijoux, A.; Gruel, G.; Bernal, M.; et al. Assessment of radio-induced damage in endothelial cells irradiated with 40 kVp, 220 kVp, and 4 MV X-rays by means of micro and nanodosimetric calculations. Int. J. Mol. Sci. 2019, 20, 6204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Incerti, S.; Baldacchino, G.; Bernal, M.; Capra, R.; Champion, C.; Francis, Z.; Guatelli, S.; Gueye, P.; Mantero, A.; Mascialino, B.; et al. The Geant4-DNA project. Int. J. Model. Simul. Sci. Comput. 2010, 1, 157–178. [Google Scholar] [CrossRef]
- Incerti, S.; Ivanchenko, A.; Karamitros, M.; Mantero, A.; Moretto, P.; Tran, H.N.; Mascialino, B.; Champion, C.; Ivanchenko, V.; Bernal, M.; et al. Comparison of Geant4 very low energy cross section models with experimental data in water. Med. Phys. 2010, 37, 4692–4708. [Google Scholar] [CrossRef]
- Bernal, M.; Bordage, M.C.; Brown, J.M.C.; Davídková, M.; Delage, E.; El Bitar, Z.; Enger, S.A.; Francis, Z.; Guatelli, S.; Ivanchenko, V.; et al. Track structure modelling in liquid water: A review of the Geant4-DNAvery low energy extension of the Geant4 Monte Carlo simulation toolkit. Phys. Med. 2015, 31, 861–874. [Google Scholar] [CrossRef]
- Incerti, S.; Kyriakou, I.; Bernal, M.; Bordage, M.C.; Francis, Z.; Guatelli, S.; Ivanchenko, V.; Karamitros, M.; Lampe, N.; Lee, S.B.; et al. Geant4-DNA example applications for track structure simulations in liquid water: A report from the Geant4-DNA Project. Med. Phys. 2018, 45, e722–e739. [Google Scholar] [CrossRef] [Green Version]
- Nikitaki, Z.; Nikolov, V.; Mavragani, I.; Plante, I.; Emfietzoglou, D.; Iliakis, G.; Georgakilas, A. Non-DSB clustered DNA lesions. Does theory colocalize with the experiment? Rad. Phys. Chem. 2016, 128, 26–35. [Google Scholar] [CrossRef]
- Frankenberg, D.; Brede, H.J.; Schrewe, U.J.; Steinmetz, C.; Frankenberg-Schwager, M.; Kasten, G.; Pralle, E. Induction of DNA Double-Strand Breaks by 1 H and 4 He Ions in Primary Human Skin Fibroblasts in the LET Range of 8 to 124 keV/μm. Rad. Res. 1999, 151, 540–549. [Google Scholar] [CrossRef]
- Tang, N. Évaluation à Partir de Modélisations Nanodosimétriques, de L’influence de la Compaction de la Chromatine sur les Effets Radio-Induits Précoces et Extension aux Effets Tardifs (Réparation des Dommages à l’ADN et Mort Cellulaire). Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, 2019. [Google Scholar]
- Steenken, S.; Jovanovic, S.V. How easily oxidizable is DNA? One-electron reduction potentials of adenosine and guanosine radicals in aqueous solution. J. Am. Chem. Soc. 1997, 119, 617–618. [Google Scholar] [CrossRef]
- Steenken, S. Purine Bases, nucleosides and nucleotides: Aqueous solution redox chemistry and transformation reactions of their radical cations and e- and OH adducts. Chem. Rev. 1989, 89, 503–520. [Google Scholar] [CrossRef]
- Pratviel, G.; Meunier, B. Guanine oxidation: One- and two-electron reactions. Chem. Eur. J. 2006, 12, 6018–6030. [Google Scholar] [CrossRef] [PubMed]
- Gimisis, T.; Cismas, C. Isolation, characterization, and independent synthesis of guanine oxidation products. Eur. J. Org. Chem. 2006, 2006, 1351–1378. [Google Scholar] [CrossRef]
- Shukla, L.; Adhikary, A.; Pazdro, R.; Becker, D.; Sevilla, M. Formation of 8-oxo-7,8-dihydroguanine-radicals in γ-irradiated DNA by multiple one-electron oxidations. Nucleic Acids Res. 2004, 32, 6565–6574. [Google Scholar] [CrossRef]
- Von Sonntag, C. (Ed.) Nucleobases, Nucleosides and Nucleotides. In Free-Radical-Induced DNA Damage and Its Repair; Springer: Berlin/Heidelberg, Germany, 2006; pp. 213–371. [Google Scholar]
- Kreth, G.; Finsterle, J.; Von Hase, J.; Cremer, M.; Cremer, C. Radial arrangement of chromosome territories in human cell nuclei: A computer model approach based on gene density indicates a probabilistic global positioning code. Biophys. J. 2004, 86, 2803–2812. [Google Scholar] [CrossRef] [Green Version]
- Kent, W.; Sugnet, C.; Furey, T.; Roskin, K.; Pringle, T.; Zahler, A.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Connelly, N.; Damhus, T.; Hartshorn, R.; Hutton, T. Nomenclature of Inorganic Chemistry—IUPAC Recommendations 2005. Chem. Int. 2005, 27, 25–26. [Google Scholar] [CrossRef]
- Karamitros, M.; Luan, S.; Bernal, M.; Allison, J.; Baldacchino, G.; Davidkova, M.; Francis, Z.; Friedland, W.; Ivantchenko, V.; Ivantchenko, A.; et al. Diffusion-controlled reactions modelling in Geant4-DNA. J. Comput. Phys. 2014, 274, 841–882. [Google Scholar] [CrossRef]
- Roots, R.; Okada, S. Estimation of life times and diffusion distances of radicals involved in X-Ray-induced DNA strand breaks or killing of mammalian cells. Rad. Res. 1975, 64, 306–320. [Google Scholar] [CrossRef]
- Stewart, R. Induction of DNA damage by light ions relative to 60Co γ-rays. Int. J. Part. Ther. 2018, 5, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Buxton, G.; Greenstock, C.; Hellman, F.; Ross, A. Critical review of rate constants for reactions of hydrated electrons, hydrogen atoms and hydroxyl radicals (•OH/ •O−) in aqueous solution. J. Phys. Chem. Ref. Data 1988, 17, 513–886. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Shinohara, H.; Kondo, M. Reactions of Hydroxyl Radicals with Nucleic Acid Bases and the Related Compounds in Gamma-irradiated Aqueous Solution. J. Radiat. Res. 1975, 16, 153–161. [Google Scholar] [CrossRef]
- Davídková, M.; Spotheim-Maurizot, M. Radiation damage to DNA in DNA–protein complexes. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2011, 711, 41–48. [Google Scholar] [CrossRef]
- Kreipl, M.; Friedland, W.; Paretzke, H. Interaction of ion tracks in spatial and temporal proximity. Radiat. Environ. Biophys. 2009, 48, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Sakata, D.; Lampe, N.; Karamitros, M.; Kyriakou, I.; Belov, O.; Bernal, M.; Bolst, D.; Bordage, M.C.; Breton, V.; Brown, J.; et al. Evaluation of early radiation DNA damage in a fractal cell nucleus model using Geant4-DNA. Phys. Med. 2019, 62, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Adhikary, A.; Dingfelder, M.; Dizdaroglu, M. Hydroxyl radical is a significant player in oxidative DNA damage in vivo. Chem. Soc. Rev. 2021, 50, 8355–8360. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | GC Rate | Heterochromatin Content |
|---|---|---|
| L1 | <37.5% | 100% |
| L2 | 37.5–42.5% | 70% |
| H1 | 42.5–47.5% | 50% |
| H2 | 47.5–52.5% | 20% |
| H3 | >52.5% | 0% |
| Reactions | Reaction Rates (109 M−1·s−1) |
|---|---|
| 2-Deoxyribose + OH• | 1.80 |
| Adenine + OH• | 6.10 |
| Guanine + OH• | 9.20 |
| Thymine + OH• | 6.40 |
| Cytosine + OH• | 6.10 |
| 2-Deoxyribose + e−aq | 0.01 |
| Adenine + e−aq | 9.00 |
| Guanine + e−aq | 14.00 |
| Thymine + e−aq | 18.00 |
| Cytosine + e−aq | 13.00 |
| 2-Deoxyribose + H• | 0.029 |
| Adenine + H• | 0.10 |
| Thymine + H• | 0.57 |
| Cytosine + H• | 0.092 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thibaut, Y.; Tang, N.; Tran, H.N.; Vaurijoux, A.; Villagrasa, C.; Incerti, S.; Perrot, Y. Nanodosimetric Calculations of Radiation-Induced DNA Damage in a New Nucleus Geometrical Model Based on the Isochore Theory. Int. J. Mol. Sci. 2022, 23, 3770. https://doi.org/10.3390/ijms23073770
Thibaut Y, Tang N, Tran HN, Vaurijoux A, Villagrasa C, Incerti S, Perrot Y. Nanodosimetric Calculations of Radiation-Induced DNA Damage in a New Nucleus Geometrical Model Based on the Isochore Theory. International Journal of Molecular Sciences. 2022; 23(7):3770. https://doi.org/10.3390/ijms23073770
Chicago/Turabian StyleThibaut, Yann, Nicolas Tang, Hoang Ngoc Tran, Aurélie Vaurijoux, Carmen Villagrasa, Sébastien Incerti, and Yann Perrot. 2022. "Nanodosimetric Calculations of Radiation-Induced DNA Damage in a New Nucleus Geometrical Model Based on the Isochore Theory" International Journal of Molecular Sciences 23, no. 7: 3770. https://doi.org/10.3390/ijms23073770