
Phosphatidic Acid in Plant Hormonal Signaling: From Target Proteins to Membrane Conformations

 ,
,
Abstract
:1. Introduction
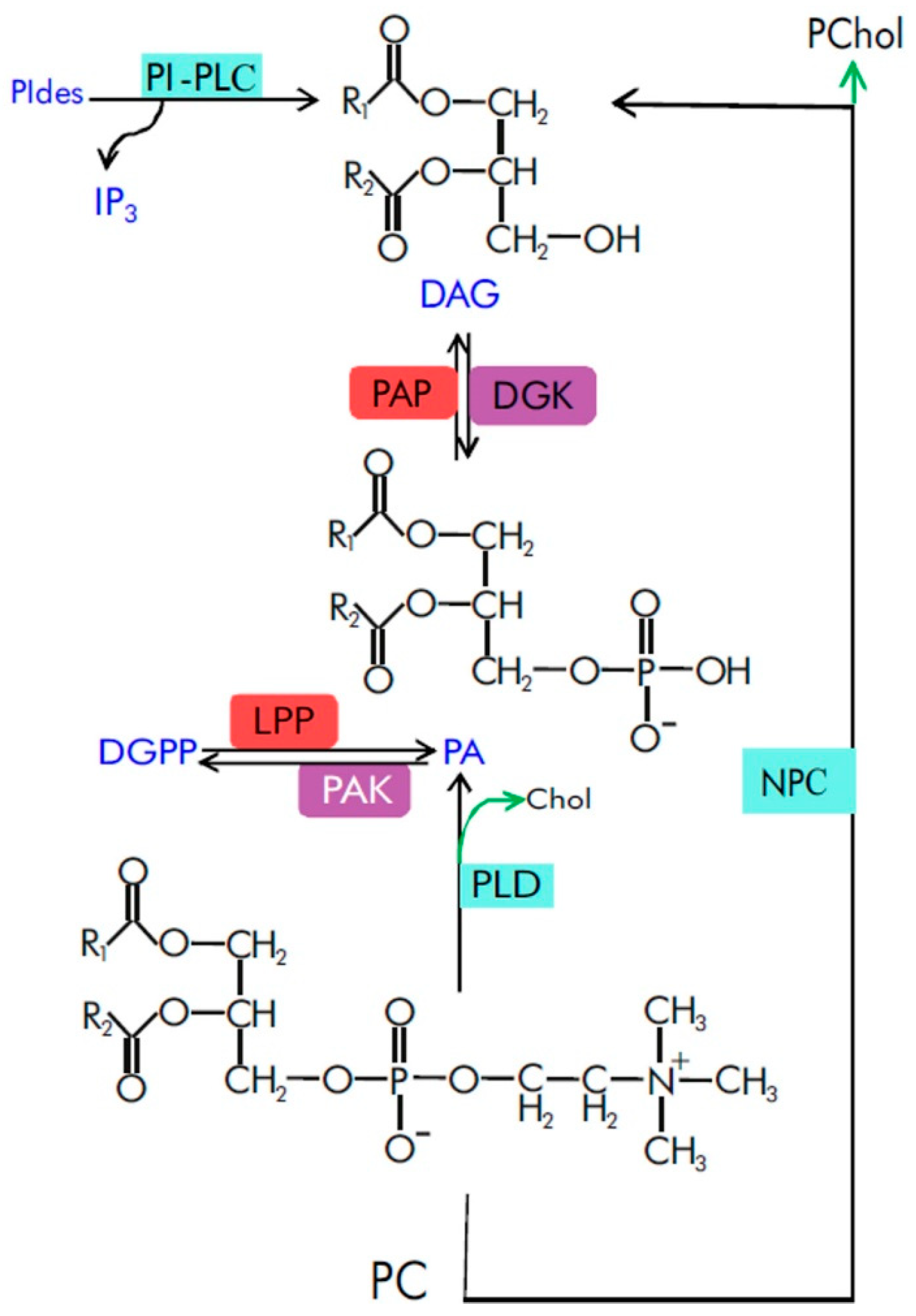
2. Spatiotemporal Characteristics of Phosphatidic Acid as a Signaling Molecule
3. Effect of PA on Membrane Status
4. Binding of Phosphatidic Acid to Target Proteins

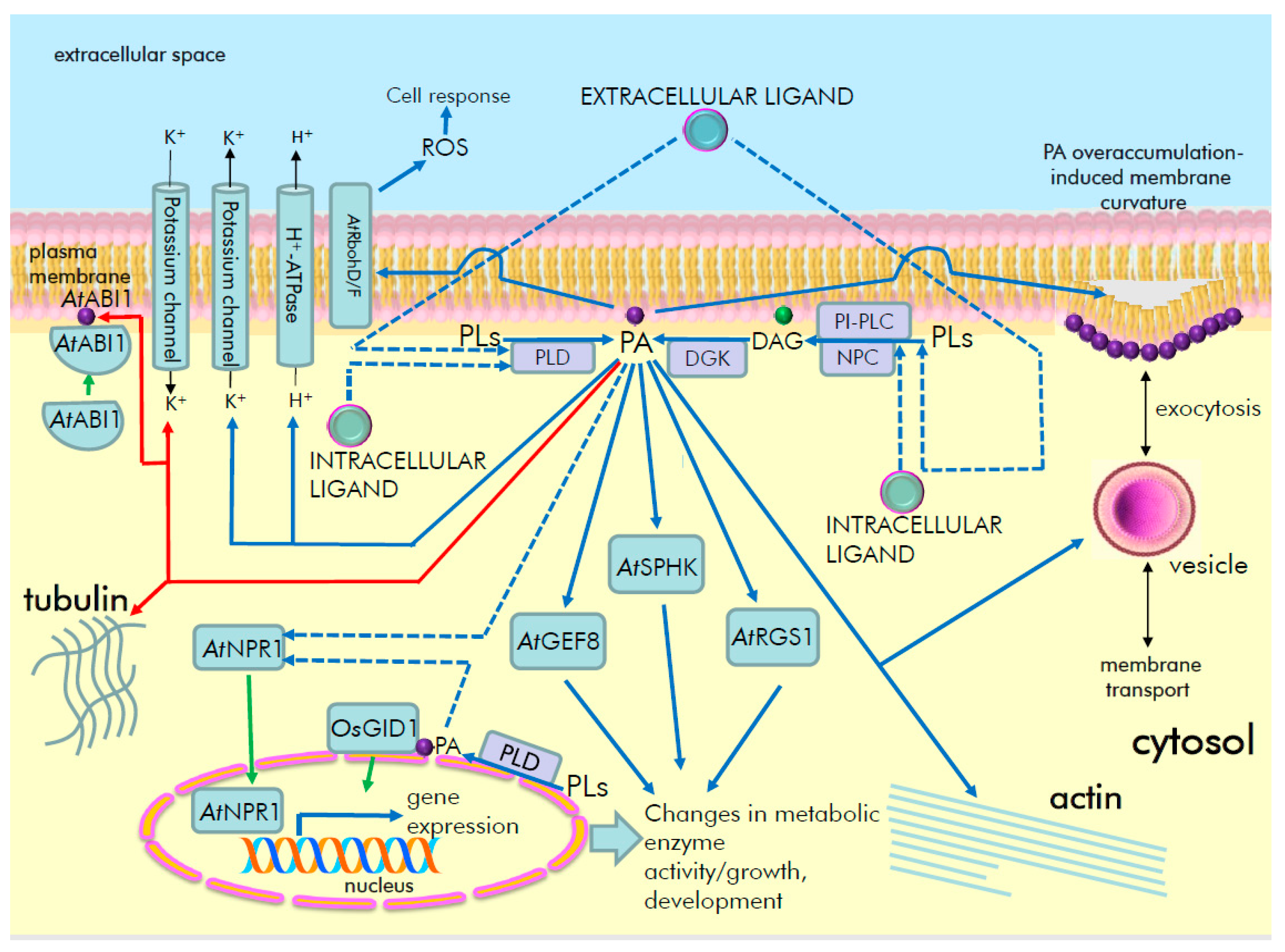
5. Role of PA as a Lipid Second Messenger of Signal Transduction Pathways in Plants
5.1. Phosphatidic Acid Signaling in Hormonal Regulation
5.1.1. Role of Phosphatidic Acid in Auxin Signaling
5.1.2. Role of Phosphatidic Acid in Salicylic Acid Signaling
5.1.3. Role of Phosphatidic Acid in Brassinosteroid Signaling
5.1.4. Role of Phosphatidic Acid in Abscisic Acid Signaling
5.1.5. Role of Phosphatidic Acid in Jasmonic Acid Signaling
5.1.6. Role of Phosphatidic Acid in Gibberellin Signaling
5.2. Phosphatidic Acid as a Signal Transducer for Other Biologically Active Substances

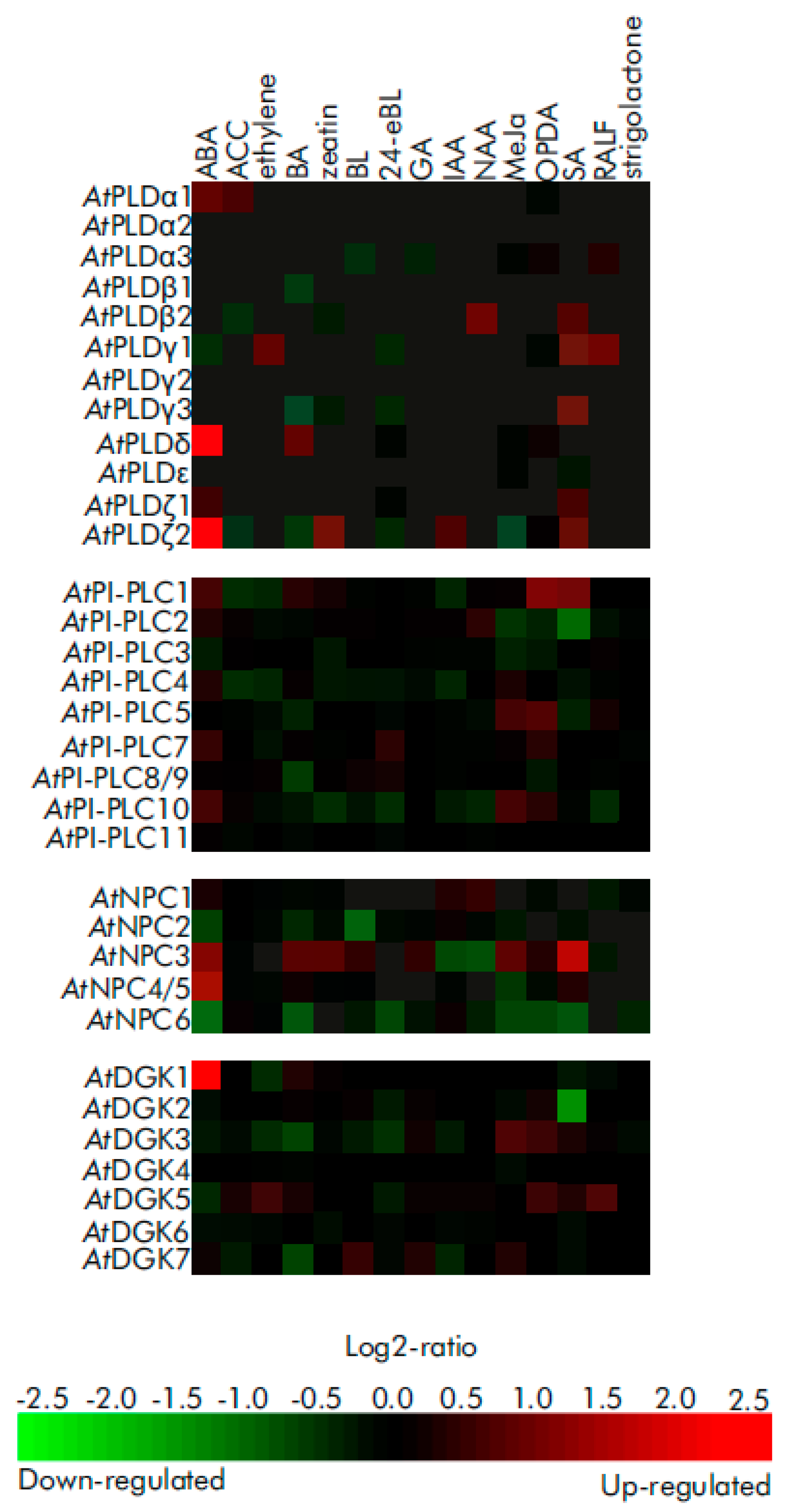
6. The Most Common Methods for the Studying of Phosphatidic Acid Signaling in Plants
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kwiatek, J.M.; Carman, G.M. Yeast phosphatidic acid phosphatase Pah1 hops and scoots along the membrane phospholipid bilayer. J. Lipid Res. 2020, 61, 1232–1243. [Google Scholar] [CrossRef] [PubMed]
- Zhukovsky, M.A.; Filograna, A.; Luini, A.; Corda, D.; Valente, C. Phosphatidic acid in membrane rearrangements. FEBS Lett. 2019, 593, 2428–2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, G.; Kim, S.-C.; Li, J.; Zhou, Y.; Wang, X. Transcriptional regulation of lipid catabolism during seedling establishment. Mol. Plant 2020, 13, 984–1000. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, J.; Chen, X.; Zhao, D.; Zhou, X.; Zhang, Y.; Wang, X.; Zhao, J. Phospholipase D- and phosphatidic acid-mediated phospholipid metabolism and signaling modulate symbiotic interaction and nodulation in soybean (Glycine max). Plant J. 2021, 106, 142–158. [Google Scholar] [CrossRef]
- Lavell, A.A.; Benning, C. Cellular organization and regulation of plant glycerolipid metabolism. Plant Cell Physiol. 2019, 60, 1176–1183. [Google Scholar] [CrossRef]
- Pokotylo, I.; Kravets, V.; Martinec, J.; Ruelland, E. The phosphatidic acid paradox: Too many actions for one molecule class? Lessons from plants. Progress Lipid Res. 2018, 71, 43–53. [Google Scholar] [CrossRef]
- Li, J.; Wang, X. Phospholipase D and phosphatidic acid in plant immunity. Plant Sci. 2019, 279, 45–50. [Google Scholar] [CrossRef]
- Meijer, H.J.G.; van Himbergen, J.A.J.; Musgrave, A.; Munnik, T. Acclimation to salt modifies the activation of several osmotic stress-activated lipid signalling pathways in Chlamydomonas. Phytochemistry 2017, 135, 64–72. [Google Scholar] [CrossRef]
- Yao, Y.; Li, J.; Lin, Y.; Zhou, J.; Zhang, P.; Xu, Y. Structural insights into phospholipase D function. Prog. Lipid Res. 2021, 81, 101070. [Google Scholar] [CrossRef]
- Kocourková, D.; Krčková, Z.; Pejchar, P.; Kroumanová, K.; Podmanická, T.; Daněk, M.; Martinec, J. Phospholipase Dα1 mediates the high-Mg2+ stress response partially through regulation of K+ homeostasis. Plant Cell Environ. 2020, 43, 2460–2475. [Google Scholar] [CrossRef]
- Kolesnikov, Y.S.; Nokhrina, K.P.; Kretynin, S.V.; Volotovski, I.D.; Martinec, J.; Romanov, G.A.; Kravets, V.S. Molecular structure of phospholipase D and regulatory mechanisms of its activity in plant and animal cells. Biochemistry 2012, 77, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocourková, D.; Kroumanová, K.; Podmanická, T.; Daněk, M.; Martinec, J. Phospholipase Dα1 acts as a negative regulator of high Mg2+-induced leaf senescence in Arabidopsis. Front. Plant Sci. 2021, 12, 770794. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, Z.; Hu, M.; Pan, Y.; Xu, X.; Zhang, Z. Delay of ripening and senescence in mango fruit by 6-benzylaminopurine is associated with inhibition of ethylene biosynthesis and membrane lipid catabolism. Postharvest Biol. Technol. 2022, 185, 111797. [Google Scholar] [CrossRef]
- Arisz, S.A.; Testerink, C.; Munnik, T. Plant PA signaling via diacylglycerol kinase. Biochim. Biophys. Acta 2009, 1791, 869–875. [Google Scholar] [CrossRef]
- Yuan, S.; Kim, S.-C.; Deng, X.; Hong, Y.; Wang, X. Diacylglycerol kinase and associated lipid mediators modulate rice root architecture. New Phytol. 2019, 223, 261–276. [Google Scholar] [CrossRef]
- Kue Foka, I.C.; Ketehouli, T.; Zhou, Y.; Li, X.-W.; Wang, F.-W.; Li, H. The emerging roles of diacylglycerol kinase (DGK) in plant stress tolerance, growth, and development. Agronomy 2020, 10, 1375. [Google Scholar] [CrossRef]
- Nakamura, Y.; Ngo, A.H. Non-specific phospholipase C (NPC): An emerging class of phospholipase C in plant growth and development. J. Plant Res. 2020, 133, 489–497. [Google Scholar] [CrossRef]
- Sagar, S.; Singh, A. Emerging role of phospholipase C mediated lipid signaling in abiotic stress tolerance and development in plants. Plant Cell Rep. 2021, 40, 2123–2133. [Google Scholar] [CrossRef]
- Craddock, C.P.; Adams, N.; Kroon, J.T.M.; Bryant, F.M.; Hussey, P.J.; Kurup, S.; Eastmond, P.J. Cyclin-dependent kinase activity enhances phosphatidylcholine biosynthesis in Arabidopsis by repressing phosphatidic acid phosphohydrolase activity. Plant J. 2017, 89, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Raza, A.; Zeng, L.; Gao, A.; Lv, Y.; Ding, X.; Cheng, Y.; Zou, X. Genome-wide analysis and expression patterns of lipid phospholipid phospholipase gene family in Brassica napus L. BMC Genom. 2021, 22, 548. [Google Scholar] [CrossRef]
- Zhou, Z.; Yang, Y.; Shan, W.; Zhang, H.; Wei, W.; Kuang, J.; Chen, J.; Lu, W. Ethylene attenuates chilling injury of banana fruit via the MabHLH060/183 module in controlling phosphatidic acid formation genes. Postharvest Biol. Technol. 2022, 183, 111724. [Google Scholar] [CrossRef]
- Takáč, T.; Novák, D.; Šamaj, J. Recent advances in the cellular and developmental biology of phospholipases in plants. Front. Plant Sci. 2019, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Wang, L. MpDGK2, a novel diacylglycerol kinase from Malus prunifolia, confers drought stress tolerance in transgenic Arabidopsis. Plant Mol. Biol. Report. 2020, 38, 452–460. [Google Scholar] [CrossRef]
- Rodas-Junco, B.A.; Racagni-Di-Palma, G.E.; Canul-Chan, M.; Usorach, J.; Hernández-Sotomayor, S.M.T. Link between lipid second messengers and osmotic stress in plants. Int. J. Mol. Sci. 2021, 22, 2658. [Google Scholar] [CrossRef]
- Zhang, Q.; van Wijk, R.; Shahbaz, M.; Roels, W.; Schooten, B.V.; Vermeer, J.E.M.; Zarza, X.; Guardia, A.; Scuffi, D.; García-Mata, C.; et al. Arabidopsis phospholipase C3 is involved in lateral root initiation and ABA responses in seed germination and stomatal closure. Plant Cell Physiol. 2018, 59, 469–486. [Google Scholar] [CrossRef]
- Zhu, L.; Dou, L.; Shang, H.; Li, H.; Yu, J.; Xiao, G. GhPIPLC2D promotes cotton fiber elongation by enhancing ethylene biosynthesis. iScience 2021, 24, 102199. [Google Scholar] [CrossRef]
- Kong, X.-X.; Mei, J.-W.; Zhang, J.; Liu, X.; Wu, J.-Y.; Wang, C.-L. Turnover of diacylglycerol kinase 4 by cytoplasmic acidification induces vacuole morphological change and nuclear DNA degradation in the early stage of pear self-incompatibility response. J. Integr. Plant Biol. 2021, 63, 2123–2135. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, R.; Zhou, Y.; Wang, S.; Zhang, B.; Kong, J.; Zheng, S.; Yang, N. PLDα1 and GPA1 are involved in the stomatal closure induced by oridonin in Arabidopsis thaliana. Funct. Plant Biol. 2021, 48, 1005–1016. [Google Scholar] [CrossRef]
- Jia, Y.; Li, W. Characterisation of lipid changes in ethylene-promoted senescence and its retardation by suppression of phospholipase Dδ in Arabidopsis leaves. Front. Plant Sci. 2015, 6, 1045. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhu, H.; Zhang, Q.; Li, M.; Yan, M.; Wang, R.; Wang, L.; Welti, R.; Zhang, W.; Wang, X. Phospholipase Dα1 and phosphatidic acid regulate NADPH oxidase activity and production of reactive oxygen species in ABA-mediated stomatal closure in Arabidopsis. Plant Cell 2009, 21, 2357–2377. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Devaiah, S.P.; Narasimhan, R.; Pan, X.; Zhang, Y.; Zhang, W.; Wang, X. Cytosolic glyceraldehyde-3-phosphate dehydrogenases interact with phospholipase Dδ to transduce hydrogen peroxide signals in the Arabidopsis response to stress. Plant Cell 2012, 24, 2200–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Qin, C.; Zhao, J.; Wang, X. Phospholipase Dα1-derived phosphatidic acid interacts with ABI1 phosphatase 2C and regulates abscisic acid signaling. Proc. Natl. Acad. Sci. USA 2004, 101, 9508–9513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Mishra, G.; Taylor, K.; Wang, X. Phosphatidic acid binds and stimulates Arabidopsis sphingosine kinases. J. Biol. Chem. 2011, 286, 13336–13345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, F.; Fanella, B.; Guo, L.; Wang, X. Membrane glycerolipidome of soybean root hairs and its response to nitrogen and phosphate availability. Sci. Rep. 2016, 6, 36172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vismans, G.; van der Meer, T.; Langevoort, O.; Schreuder, M.; Bouwmeester, H.; Peisker, H.; Dörman, P.; Ketelaar, T.; van der Krol, A. Low-phosphate induction of plastidal stromules is dependent on strigolactones but not on the canonical strigolactone signaling component MAX2. Plant Physiol. 2016, 172, 2235–2244. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Gong, R.; Yuan, S.; Su, Y.; Lv, W.; Zhou, Y.; Zhang, Q.; Deng, X.; Tong, P.; Liang, S.; et al. Phospholipase Dα6 and phosphatidic acid regulate gibberellin signaling in rice. EMBO Rep. 2021, 22, e51871. [Google Scholar] [CrossRef] [PubMed]
- Sakane, F.; Hoshino, F.; Murakami, C. New era of diacylglycerol kinase, phosphatidic acid and phosphatidic acid-binding protein. Int. J. Mol. Sci. 2020, 21, 6794. [Google Scholar] [CrossRef]
- McDermott, M.I.; Wang, Y.; Wakelam, M.J.O.; Bankaitis, V.A. Mammalian phospholipase D: Function, and therapeutics. Prog. Lipid Res. 2020, 78, 101018. [Google Scholar] [CrossRef]
- Lutkewitte, A.J.; Finck, B.N. Regulation of signaling and metabolism by lipin-mediated phosphatidic acid phosphohydrolase activity. Biomolecules 2020, 10, 1386. [Google Scholar] [CrossRef]
- Graber, Z.; Owusu Kwarteng, D.; Lange, S.M.; Koukanas, Y.; Khalifa, H.; Mutambuze, J.W.; Kooijman, E.E. The electrostatic basis of diacylglycerol pyrophosphate-protein interaction. Cells 2022, 11, 290. [Google Scholar] [CrossRef]
- Potocky, M.; Pleskot, R.; Pejchar, P.; Vitale, N.; Kost, B.; Zarsky, V. Live-cell imaging of phosphatidic acid dynamics in pollen tubes visualized by Spo20p-derived biosensor. New Phytol. 2014, 203, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Pejchar, P.; Sekereš, J.; Novotný, O.; Žárský, V.; Potocký, M. Functional analysis of phospholipase Dδ family in tobacco pollen tubes. Plant J. 2020, 103, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Kubátová, Z.; Pejchar, P.; Potocký, M.; Sekereš, J.; Žárský, V.; Kulich, I. Arabidopsis trichome contains two plasma membrane domains with different lipid compositions which attract distinct EXO70 subunits. Int. J. Mol. Sci. 2019, 20, 3803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platre, M.P.; Noack, L.C.; Doumane, M.; Bayle, V.; Simon, M.L.A.; Maneta-Peyret, L.; Fouillen, L.; Stanislas, T.; Armengot, L.; Pejchar, P.; et al. A combinatorial lipid code shapes the electrostatic landscape of plant endomembranes. Dev. Cell 2018, 45, 465–480.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertle, A.P.; García-Cerdán, J.G.; Armbruster, U.; Shih, R.; Lee, J.J.; Wong, W.; Niyogi, K.K. A Sec14 domain protein is required for photoautotrophic growth and chloroplast vesicle formation in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2020, 117, 9101–9111. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Song, T.; Wallrad, L.; Kudla, J.; Wang, X.; Zhang, W. Tissue-specific accumulation of pH-sensing phosphatidic acid determines plant stress tolerance. Nat. Plants 2019, 5, 1012–1021. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Yao, H.-Y.; Xue, H.-W. Lipidomic profiling analysis reveals the dynamics of phospholipid molecules in Arabidopsis thaliana seedling growth. J. Integr. Plant Biol. 2016, 58, 890–902. [Google Scholar] [CrossRef]
- Devaiah, S.P.; Roth, M.R.; Baughman, E.; Li, M.; Tamura, P.; Jeannotte, R.; Welti, R.; Wang, X. Quantitative profiling of polar glycerolipid species from organs of wild-type Arabidopsis and a phospholipase Dalpha1 knockout mutant. Phytochemistry 2006, 67, 1907–1924. [Google Scholar] [CrossRef]
- Yunus, I.S.; Cazenave-Gassiot, A.; Liu, Y.-C.; Lin, Y.-C.; Wenk, M.R.; Nakamura, Y. Phosphatidic acid is a major phospholipid class in reproductive organs of Arabidopsis thaliana. Plant Signal. Behav. 2015, 10, e1049790. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Teo, N.Z.; Shui, G.; Chua, C.H.; Cheong, W.F.; Parameswaran, S.; Koizumi, R.; Ohta, H.; Wenk, M.R.; Ito, T. Transcriptomic and lipidomic profiles of glycerolipids during Arabidopsis flower development. New Phytol. 2014, 203, 310–322. [Google Scholar] [CrossRef]
- Arisz, S.A.; van Wijk, R.; Roels, W.; Zhu, J.-K.; Haring, M.A.; Munnik, T. Rapid phosphatidic acid accumulation in response to low temperature stress in Arabidopsis is generated through diacylglycerol kinase. Front. Plant Sci. 2013, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarza, X.; Shabala, L.; Fujita, M.; Shabala, S.; Haring, M.A.; Tiburcio, A.F.; Munnik, T. Extracellular spermine triggers a rapid intracellular phosphatidic acid response in Arabidopsis, involving PLDδ activation and stimulating ion flux. Front. Plant Sci. 2019, 10, 601. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, H.; Li, K.; Wu, Y.; Liu, Z.; Huang, C. Genome-wide proteomic profiling reveals the role of dominance protein expression in heterosis in immature maize ears. Sci. Rep. 2017, 7, 16130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.-J.; Xiao, G.-H.; Liu, N.-J.; Liu, D.; Chen, P.-S.; Qin, Y.-M.; Zhu, Y.-X. Targeted lipidomics studies reveal that linolenic acid promotes cotton fiber elongation by activating phosphatidylinositol and phosphatidylinositol monophosphate biosynthesis. Mol. Plant 2015, 8, 911–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hruz, T.; Laule, O.; Szabo, G.; Wessendorp, F.; Bleuler, S.; Oertle, L.; Widmayer, P.; Gruissem, W.; Zimmermann, P. Genevestigator V3: A reference expression database for the meta-analysis of transcriptomes. Adv. Bioinform. 2008, 2008, 420747. [Google Scholar] [CrossRef]
- Stenzel, I.; Ischebeck, T.; Vu-Becker, L.H.; Riechmann, M.; Krishnamoorthy, P.; Fratini, M.; Heilmann, I. Coordinated localization and antagonistic function of NtPLC3 and PI4P 5-Kinases in the subapical plasma membrane of tobacco pollen tubes. Plants 2020, 9, 452. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Shi, Y.; Liu, G.; Guo, Y.; Yang, Y. Activation of ROP6 GTPase by phosphatidylglycerol in Arabidopsis. Front. Plant Sci. 2018, 9, 347. [Google Scholar] [CrossRef]
- de Jong, F.; Munnik, T. Attracted to membranes: Lipid-binding domains in plants. Plant Physiol. 2021, 185, 707–723. [Google Scholar] [CrossRef]
- Hempel, F.; Stenzel, I.; Heilmann, M.; Krishnamoorthy, P.; Menzel, W.; Golbik, R.; Helm, S.; Dobritzsch, D.; Baginsky, S.; Lee, J.; et al. MAPKs influence pollen tube growth by controlling the formation of phosphatidylinositol 4,5-bisphosphate in an apical plasma membrane domain. Plant Cell 2017, 29, 3030–3050. [Google Scholar] [CrossRef] [Green Version]
- Janda, M.; Šašek, V.; Chmelařová, H.; Andrejch, J.; Nováková, M.; Hajšlová, J.; Burketová, L.; Valentová, O. Phospholipase D affects translocation of NPR1 to the nucleus in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 59. [Google Scholar] [CrossRef] [Green Version]
- Scandola, S.; Samuel, M.A. A flower-specific phospholipase D is a stigmatic compatibility factor targeted by the self-incompatibility response in Brassica napus. Curr. Biol. 2019, 29, 506–512.e4. [Google Scholar] [CrossRef] [Green Version]
- Pleskot, R.; Pejchar, P.; Bezvoda, R.; Lichtscheidl, I.; Wolters-Arts, M.; Marc, J.; Žárský, V.; Potocký, M. Turnover of phosphatidic acid through distinct signaling pathways affects multiple aspects of pollen tube growth in tobacco. Front. Plant Sci. 2012, 3, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvan-Ampudia, C.S.; Julkowska, M.M.; Darwish, E.; Gandullo, J.; Korver, R.A.; Brunoud, G.; Haring, M.A.; Munnik, T.; Vernoux, T.; Testerink, C. Halotropism Is a Response of Plant Roots to Avoid a Saline Environment. Curr. Biol. 2013, 23, 2044–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaz Dias, F.; Serrazina, S.; Vitorino, M.; Marchese, D.; Heilmann, I.; Godinho, M.; Rodrigues, M.; Malhó, R. A role for diacylglycerol kinase 4 in signalling crosstalk during Arabidopsis pollen tube growth. New Phytol. 2019, 222, 1434–1446. [Google Scholar] [CrossRef] [PubMed]
- Wattelet-Boyer, V.; Le Guédard, M.; Dittrich-Domergue, F.; Maneta-Peyret, L.; Kriechbaumer, V.; Boutté, Y.; Bessoule, J.-J.; Moreau, P. Lysophosphatidic acid acyltransferases: A link with intracellular protein trafficking in Arabidopsis root cells? J. Exp. Bot. 2021, 73, 1327–1343. [Google Scholar] [CrossRef] [PubMed]
- Novák, D.; Vadovič, P.; Ovečka, M.; Šamajová, O.; Komis, G.; Colcombet, J.; Šamaj, J. Gene expression pattern and protein localization of Arabidopsis phospholipase D alpha 1 revealed by advanced light-sheet and super-resolution microscopy. Front. Plant Sci. 2018, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, M.; van Oostende-Triplet, C.; Chebli, Y.; Testerink, C.; Bednarek, S.Y.; Geitmann, A. Plant AP180 N-Terminal Homolog proteins are involved in clathrin-dependent endocytosis during pollen tube growth in Arabidopsis thaliana. Plant Cell Physiol. 2019, 60, 1316–1330. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, F.; Arisz, S.A.; Dekker, H.L.; Kramer, G.; De Koster, C.G.; Haring, M.A.; Munnik, T.; Testerink, C. Identification of novel candidate phosphatidic acid-binding proteins involved in the salt-stress response of Arabidopsis thaliana roots. Biochem. J. 2013, 450, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekereš, J.; Pejchar, P.; Šantrůček, J.; Vukašinović, N.; Žárský, V.; Potocký, M. Analysis of exocyst subunit EXO70 family reveals distinct membrane polar domains in tobacco pollen tubes. Plant Physiol. 2017, 173, 1659–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putta, P.; Rankenberg, J.; Korver, R.A.; van Wijk, R.; Munnik, T.; Testerink, C.; Kooijman, E.E. Phosphatidic acid binding proteins display differential binding as a function of membrane curvature stress and chemical properties. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 2709–2716. [Google Scholar] [CrossRef]
- Putta, P.; Creque, E.; Piontkivska, H.; Kooijman, E.E. Lipid-protein interactions for ECA1 an N-ANTH domain protein involved in stress signaling in plants. Chem. Phys. Lipids 2020, 231, 104919. [Google Scholar] [CrossRef] [PubMed]
- Kulig, W.; Korolainen, H.; Zatorska, M.; Kwolek, U.; Wydro, P.; Kepczynski, M.; Róg, T. Complex behavior of phosphatidylcholine–phosphatidic acid bilayers and monolayers: Effect of acyl chain unsaturation. Langmuir 2019, 35, 5944–5956. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.A.F.; Vila-Viçosa, D.; Teixeira, V.H.; Baptista, A.M.; Machuqueiro, M. Constant-pH MD simulations of DMPA/DMPC lipid bilayers. J. Chem. Theory Comput. 2015, 11, 5973–5979. [Google Scholar] [CrossRef] [PubMed]
- Tanguy, E.; Kassas, N.; Vitale, N. Protein-phospholipid interaction motifs: A focus on phosphatidic acid. Biomolecules 2018, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Wang, P.; Song, H.; Jing, W.; Shen, L.; Zhang, Q.; Zhang, W. Phosphatidic acid binds to and regulates guanine nucleotide exchange factor 8 (GEF8) activity in Arabidopsis. Funct. Plant Biol. 2017, 44, 1029–1038. [Google Scholar] [CrossRef]
- Cools, T.L.; Vriens, K.; Struyfs, C.; Verbandt, S.; Ramada, M.H.S.; Brand, G.D.; Bloch, C.; Koch, B.; Traven, A.; Drijfhout, J.W.; et al. The antifungal plant defensin HsAFP1 is a phosphatidic acid-interacting peptide inducing membrane permeabilization. Front. Microbiol. 2017, 8, 2295. [Google Scholar] [CrossRef] [Green Version]
- Im, Y.J.; Davis, A.J.; Perera, I.Y.; Johannes, E.; Allen, N.S.; Boss, W.F. The N-terminal membrane occupation and recognition nexus domain of Arabidopsis phosphatidylinositol phosphate kinase 1 regulates enzyme activity. J. Biol. Chem. 2007, 282, 5443–5452. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J.; Eriksson, S.K.; Harryson, P.; Pierog, S.; Colby, T.; Bartels, D.; Röhrig, H. The lysine-rich motif of intrinsically disordered stress protein CDeT11-24 from Craterostigma plantagineum is responsible for phosphatidic acid binding and protection of enzymes from damaging effects caused by desiccation. J. Exp. Bot. 2012, 63, 4919–4929. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Shen, L.; Guo, J.; Jing, W.; Qu, Y.; Li, W.; Bi, R.; Xuan, W.; Zhang, Q.; Zhang, W. Phosphatidic acid directly regulates PINOID-dependent phosphorylation and activation of the PIN-FORMED2 auxin efflux transporter in response to salt stress. Plant Cell 2019, 31, 250–271. [Google Scholar] [CrossRef] [Green Version]
- Julkowska, M.M.; McLoughlin, F.; Galvan-Ampudia, C.S.; Rankenberg, J.M.; Kawa, D.; Klimecka, M.; Haring, M.A.; Munnik, T.; Kooijman, E.E.; Testerink, C. Identification and functional characterization of the Arabidopsis Snf1-related protein kinase SnRK2.4 phosphatidic acid-binding domain. Plant Cell Environ. 2015, 38, 614–624. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Tian, Q.; Yang, L.; Zhang, H.; Shi, Y.; Shen, Y.; Zhou, Z.; Wu, Q.; Zhang, Q.; Zhang, W. Phosphatidic acid directly binds with rice potassium channel OsAKT2 to inhibit its activity. Plant J. 2020, 102, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Yang, L.; Zhang, W. Multiple basic amino acid residues contribute to phosphatidic acid-mediated inhibition of rice potassium channel OsAKT2. Plant Signal. Behav. 2020, 15, 1789818. [Google Scholar] [CrossRef] [PubMed]
- Yperman, K.; Papageorgiou, A.C.; Merceron, R.; De Munck, S.; Bloch, Y.; Eeckhout, D.; Jiang, Q.; Tack, P.; Grigoryan, R.; Evangelidis, T.; et al. Distinct EH domains of the endocytic TPLATE complex confer lipid and protein binding. Nat. Commun. 2021, 12, 3050. [Google Scholar] [CrossRef] [PubMed]
- Testerink, C.; Larsen, P.B.; van der Does, D.; van Himbergen, J.A.J.; Munnik, T. Phosphatidic acid binds to and inhibits the activity of Arabidopsis CTR1. J. Exp. Bot. 2007, 58, 3905–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleskot, R.; Pejchar, P.; Žárský, V.; Staiger, C.J.; Potocký, M. Structural insights into the inhibition of actin-capping protein by interactions with phosphatidic acid and phosphatidylinositol (4,5)-bisphosphate. PLoS Comput. Biol. 2012, 8, e1002765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, S.; Dalal, V.; Mishra, G. Identification of novel phosphatidic acid binding domain on sphingosine kinase 1 of Arabidopsis thaliana. Plant Physiol. Biochem. 2018, 128, 178–184. [Google Scholar] [CrossRef]
- Zimmerberg, J.; Kozlov, M.M. How proteins produce cellular membrane curvature. Nat. Rev. Mol. Cell Biol. 2005, 7, 9–19. [Google Scholar] [CrossRef]
- Yao, H.; Wang, G.; Guo, L.; Wang, X. Phosphatidic acid interacts with a MYB transcription factor and regulates its nuclear localization and function in Arabidopsis. Plant Cell 2013, 25, 5030–5042. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-C.; Guo, L.; Wang, X. Phosphatidic acid binds to cytosolic glyceraldehyde-3-phosphate dehydrogenase and promotes its cleavage in Arabidopsis. J. Biol. Chem. 2013, 288, 11834–11844. [Google Scholar] [CrossRef] [Green Version]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific α-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef]
- Roy Choudhury, S.; Pandey, S. Phosphatidic acid binding inhibits RGS1 activity to affect specific signaling pathways in Arabidopsis. Plant J. 2017, 90, 466–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-C.; Nusinow, D.A.; Sorkin, M.L.; Pruneda-Paz, J.; Wang, X. Interaction and regulation between lipid mediator phosphatidic acid and circadian clock regulators. Plant Cell 2019, 31, 399–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Y.; Song, P.; Hu, Y.; Jin, X.; Jia, Q.; Zhang, X.; Chen, L.; Zhang, Q. Regulation of stomatal movement by cortical microtubule organization in response to darkness and ABA signaling in Arabidopsis. Plant Growth Regul. 2018, 84, 467–479. [Google Scholar] [CrossRef]
- Shen, L.; Zhuang, B.; Wu, Q.; Zhang, H.; Nie, J.; Jing, W.; Yang, L.; Zhang, W. Phosphatidic acid promotes the activation and plasma membrane localization of MKK7 and MKK9 in response to salt stress. Plant Sci. 2019, 287, 110190. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, D.M.; Yu, W.W.; Shi, L.L.; Zhang, Y.; Lai, Y.X.; Huang, L.P.; Qi, H.; Chen, Q.F.; Yao, N.; et al. Phosphatidic acid modulates MPK3- and MPK6-mediated hypoxia signaling in Arabidopsis. Plant Cell 2021, 34, 889–909. [Google Scholar] [CrossRef]
- Barbosa, I.C.R.; Shikata, H.; Zourelidou, M.; Heilmann, M.; Heilmann, I.; Schwechheimer, C. Phospholipid composition and a polybasic motif determine D6 PROTEIN KINASE polar association with the plasma membrane and tropic responses. Development 2016, 143, 4687–4700. [Google Scholar] [CrossRef] [Green Version]
- Klimecka, M.; Bucholc, M.; Maszkowska, J.; Krzywińska, E.; Goch, G.; Lichocka, M.; Szczegielniak, J.; Dobrowolska, G. Regulation of ABA-non-activated SNF1-related protein kinase 2 signaling pathways by phosphatidic acid. Int. J. Mol. Sci. 2020, 21, 4984. [Google Scholar] [CrossRef]
- Chen, X.; Li, L.; Xu, B.; Zhao, S.; Lu, P.; He, Y.; Ye, T.; Feng, Y.-Q.; Wu, Y. Phosphatidylinositol-specific phospholipase C2 functions in auxin-modulated root development. Plant Cell Environ. 2018, 42, 1441–1457. [Google Scholar] [CrossRef]
- Peters, C.; Kim, S.-C.; Devaiah, S.; LI, M.; Wang, X. Non-specific phospholipase C5 and diacylglycerol promote lateral root development under mild salt stress in Arabidopsis. Plant Cell Environ. 2014, 37, 2002–2013. [Google Scholar] [CrossRef]
- Tan, S.; Zhang, X.; Kong, W.; Yang, X.-L.; Molnár, G.; Vondráková, Z.; Filepová, R.; Petrášek, J.; Friml, J.; Xue, H.-W. The lipid code-dependent phosphoswitch PDK1–D6PK activates PIN-mediated auxin efflux in Arabidopsis. Nat. Plants 2020, 6, 556–569. [Google Scholar] [CrossRef]
- Chang, X.; Riemann, M.; Liu, Q.; Nick, P. Actin as Deathly Switch? How Auxin Can Suppress Cell-Death Related Defence. PLoS ONE 2015, 10, e0125498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altúzar-Molina, A.R.; Muñoz-Sánchez, J.A.; Vázquez-Flota, F.; Monforte-González, M.; Racagni-Di Palma, G.; Hernández-Sotomayor, S.M.T. Phospholipidic signaling and vanillin production in response to salicylic acid and methyl jasmonate in Capsicum chinense J. cells. Plant Physiol. Biochem. 2011, 49, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Kalachova, T.; Iakovenko, O.; Kretinin, S.; Kravets, V. Involvement of phospholipase D and NADPH-oxidase in salicylic acid signaling cascade. Plant Physiol. Biochem. 2013, 66, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Rodas-Junco, B.A.; Muñoz-Sánchez, J.A.; Vázquez-Flota, F.; Hernández-Sotomayor, S.M.T. Salicylic-acid elicited phospholipase D responses in Capsicum chinense cell cultures. Plant Physiol. Biochem. 2015, 90, 32–37. [Google Scholar] [CrossRef]
- Krinke, O.; Flemr, M.; Vergnolle, C.; Collin, S.; Renou, J.-P.; Taconnat, L.; Yu, A.; Burketová, L.; Valentová, O.; Zachowski, A.; et al. Phospholipase D Activation Is an Early Component of the Salicylic Acid Signaling Pathway in Arabidopsis Cell Suspensions. Plant Physiol. 2009, 150, 424. [Google Scholar] [CrossRef] [Green Version]
- Rainteau, D.; Humbert, L.; Delage, E.; Vergnolle, C.; Cantrel, C.; Maubert, M.-A.; Lanfranchi, S.; Maldiney, R.; Collin, S.; Wolf, C.; et al. Acyl chains of phospholipase D transphosphatidylation products in Arabidopsis cells: A study using multiple reaction monitoring mass spectrometry. PLoS ONE 2012, 7, e41985. [Google Scholar] [CrossRef] [Green Version]
- Kalachova, T.; Puga-Freitas, R.; Kravets, V.; Soubigou-Taconnat, L.; Repellin, A.; Balzergue, S.; Zachowski, A.; Ruelland, E. The inhibition of basal phosphoinositide-dependent phospholipase C activity in Arabidopsis suspension cells by abscisic or salicylic acid acts as a signalling hub accounting for an important overlap in transcriptome remodelling induced by these hormones. Env. Exp. Bot. 2016, 123, 37–49. [Google Scholar] [CrossRef]
- Ruelland, E.; Pokotylo, I.; Djafi, N.; Cantrel, C.; Repellin, A.; Zachowski, A. Salicylic acid modulates levels of phosphoinositide dependent-phospholipase C substrates and products to remodel the Arabidopsis suspension cell transcriptome. Front. Plant Sci 2014, 5, 608. [Google Scholar] [CrossRef] [Green Version]
- Rodas-Junco, B.A.; Cab-Guillen, Y.; Muñoz-Sanchez, J.A.; Vázquez-Flota, F.; Monforte-Gonzalez, M.; Hérnandez-Sotomayor, S.M.T. Salicylic acid induces vanillin synthesis through the phospholipid signaling pathway in Capsicum chinense cell cultures. Plant Signal. Behav. 2013, 8, e26752. [Google Scholar] [CrossRef] [Green Version]
- Wimalasekera, R.; Pejchar, P.; Holk, A.; Martinec, J.; Scherer, G.F.E. Plant phosphatidylcholine-hydrolyzing phospholipases C NPC3 and NPC4 with roles in root development and brassinolide signaling in Arabidopsis thaliana. Mol. Plant 2010, 3, 610–625. [Google Scholar] [CrossRef]
- Pokotylo, I.V.; Kretynin, S.V.; Khripach, V.A.; Ruelland, E.; Blume, Y.B.; Kravets, V.S. Influence of 24-epibrassinolide on lipid signalling and metabolism in Brassica napus. Plant Growth Regul. 2014, 73, 9–17. [Google Scholar] [CrossRef]
- Derevyanchuk, M.; Kretynin, S.; Iakovenko, O.; Litvinovskaya, R.; Zhabinskii, V.; Martinec, J.; Blume, Y.; Khripach, V.; Kravets, V. Effect of 24-epibrassinolide on Brassica napus alternative respiratory pathway, guard cells movements and phospholipid signaling under salt stress. Steroids 2017, 117, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Derevyanchuk, M.; Kretynin, S.; Kolesnikov, Y.; Litvinovskaya, R.; Martinec, J.; Khripach, V.; Kravets, V. Seed germination, respiratory processes and phosphatidic acid accumulation in Arabidopsis diacylglycerol kinase knockouts – The effect of brassinosteroid, brassinazole and salinity. Steroids 2019, 147, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Kumar, R.; Baek, D.; Hyun, T.-K.; Chung, W.S.; Yun, D.-J.; Kim, J.-Y. Arabidopsis thaliana RECEPTOR DEAD KINASE1 functions as a positive regulator in plant responses to ABA. Mol. Plant 2017, 10, 223–243. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Wu, K.; Lin, F.; Qu, Y.; Liu, X.; Zhang, Q. Phosphatidic acid integrates calcium signaling and microtubule dynamics into regulating ABA-induced stomatal closure in Arabidopsis. Planta 2014, 239, 565–575. [Google Scholar] [CrossRef]
- Uwase, G.; Enrico, T.; Chelimo, D.; Keyser, B.; Johnson, R. Measuring gene expression in bombarded barley aleurone layers with increased throughput. J. Vis. Exp. 2018, 133, 56728. [Google Scholar] [CrossRef]
- Pandey, S.; Wang, R.-S.; Wilson, L.; Li, S.; Zhao, Z.; Gookin, T.E.; Assmann, S.M.; Albert, R. Boolean modeling of transcriptome data reveals novel modes of heterotrimeric G-protein action. Mol. Syst. Biol. 2010, 6, 372. [Google Scholar] [CrossRef]
- Bates, G.W.; Rosenthal, D.M.; Sun, J.; Chattopadhyay, M.; Peffer, E.; Yang, J.; Ort, D.R.; Jones, A.M. A comparative study of the Arabidopsis thaliana guard-cell transcriptome and its modulation by sucrose. PLoS ONE 2012, 7, e49641. [Google Scholar] [CrossRef]
- Cenzano, A.; Cantoro, R.; Racagni, G.; De Los Santos-Briones, C.; Hernández-Sotomayor, T.; Abdala, G. Phospholipid and phospholipase changes by jasmonic acid during stolon to tuber transition of potato. Plant Growth Regul. 2008, 56, 307. [Google Scholar] [CrossRef]
- Profotová, B.; Burketová, L.; Novotná, Z.; Martinec, J.; Valentová, O. Involvement of phospholipases C and D in early response to SAR and ISR inducers in Brassica napus plants. Plant Physiol. Biochem. 2006, 44, 143–151. [Google Scholar] [CrossRef]
- Muñoz-Sánchez, J.A.; Altúzar-Molina, A.; Hérnandez-Sotomayor, S.M.T. Phospholipase signaling is modified differentially by phytoregulators in Capsicum chinense J. cells. Plant Signal. Behav. 2012, 7, 1103–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Liu, X.; Yin, X.; Dong, T.; Yu, M.; Wu, Y. Rice non-specific phospholipase C6 is involved in mesocotyl elongation. Plant Cell Physiol. 2021, 62, 985–1000. [Google Scholar] [CrossRef] [PubMed]
- Amini, A.; Glévarec, G.; Andreu, F.; Reverdiau, P.; Rideau, M.; Crèche, J. Effects of phosphatidic acid on cytokinin signal transduction in periwinkle cells. J. Plant Growth Regul. 2008, 27, 394–399. [Google Scholar] [CrossRef]
- Kravets, V.S.; Kolesnikov, Y.S.; Kretynin, S.V.; Getman, I.A.; Romanov, G.A. Rapid activation of specific phospholipase(s) D by cytokinin in Amaranthus assay system. Physiol. Plant 2010, 138, 249–255. [Google Scholar] [CrossRef]
- Qu, Y.; An, Z.; Zhuang, B.; Jing, W.; Zhang, Q.; Zhang, W. Copper amine oxidase and phospholipase D act independently in abscisic acid (ABA)-induced stomatal closure in Vicia faba and Arabidopsis. J. Plant Res. 2014, 127, 533–544. [Google Scholar] [CrossRef]
- Echevarría-Machado, I.; Ramos-Díaz, A.; Brito-Argáez, L.; Racagni-Di Palma, G.; Loyola-Vargas, V.M.; Hernández-Sotomayor, S.M.T. Polyamines modify the components of phospholipids-based signal transduction pathway in Coffea arabica L. cells. Plant Physiol. Biochem. 2005, 43, 874–881. [Google Scholar] [CrossRef]
- Gully, K.; Pelletier, S.; Guillou, M.-C.; Ferrand, M.; Aligon, S.; Pokotylo, I.; Perrin, A.; Vergne, E.; Fagard, M.; Ruelland, E.; et al. The SCOOP12 peptide regulates defense response and root elongation in Arabidopsis thaliana. J. Exp. Bot. 2019, 70, 1349–1365. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Xuan, Y.; Xu, M.; Wang, R.-S.; Ho, C.-H.; Lalonde, S.; You, C.H.; Sardi, M.I.; Parsa, S.A.; Smith-Valle, E.; et al. Border control—a membrane-linked interactome of Arabidopsis. Science 2014, 344, 711–716. [Google Scholar] [CrossRef]
- Scuffi, D.; Nietzel, T. Hydrogen sulfide increases production of NADPH oxidase-dependent hydrogen peroxide and phospholipase D-derived phosphatidic acid in guard cell signaling. Plant Physiol. 2018, 176, 2532–2542. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, P.; de Graaf, B.H.J.; Zhang, H.; Jiao, H.; Tang, C.; Zhang, S.; Wu, J. Phosphatidic acid counteracts S-RNase signaling in pollen by stabilizing the actin cytoskeleton. Plant Cell 2018, 30, 1023–1039. [Google Scholar] [CrossRef]
- Chen, Z.-F.; Ru, J.-N.; Sun, G.-Z.; Du, Y.; Chen, J.; Zhou, Y.-B.; Chen, M.; Ma, Y.-Z.; Xu, Z.-S.; Zhang, X.-H. Genomic-wide analysis of the PLC family and detection of GmPI-PLC7 responses to drought and salt stresses in soybean. Front. Plant Sci. 2021, 12, 631470. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; MacLean, D.; Jikumaru, Y.; Hill, L.; Yamaguchi, S.; Kamiya, Y.; Jones, J.D.G. The microRNA miR393 re-directs secondary metabolite biosynthesis away from camalexin and towards glucosinolates. Plant J. 2011, 67, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Goda, H.; Sasaki, E.; Akiyama, K.; Maruyama-Nakashita, A.; Nakabayashi, K.; Li, W.; Ogawa, M.; Yamauchi, Y.; Preston, J.; Aoki, K.; et al. The AtGenExpress hormone and chemical treatment data set: Experimental design, data evaluation, model data analysis and data access. Plant J. Cell Mol. Biol. 2008, 55, 526–542. [Google Scholar] [CrossRef]
- Van Zanten, M.; Basten Snoek, L.; Van Eck-Stouten, E.; Proveniers, M.C.G.; Torii, K.U.; Voesenek, L.A.C.J.; Peeters, A.J.M.; Millenaar, F.F. Ethylene-induced hyponastic growth in Arabidopsis thaliana is controlled by ERECTA. Plant J. 2010, 61, 83–95. [Google Scholar] [CrossRef]
- Argyros, R.D.; Mathews, D.E.; Chiang, Y.-H.; Palmer, C.M.; Thibault, D.M.; Etheridge, N.; Argyros, D.A.; Mason, M.G.; Kieber, J.J.; Schaller, G.E. Type B Response Regulators of Arabidopsis play key roles in cytokinin signaling and plant development. Plant Cell 2008, 20, 2102–2116. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Singh, M.; Laxmi, A. Multiple interactions between glucose and brassinosteroid signal transduction pathways in Arabidopsis are uncovered by whole-genome transcriptional profiling. Plant Physiol. 2015, 168, 1091–1105. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.; Hilbert, B.; Dueckershoff, K.; Roitsch, T.; Krischke, M.; Mueller, M.J.; Berger, S. General detoxification and stress responses are mediated by oxidized lipids through TGA transcription factors in Arab. Plant Cell 2008, 20, 768–785. [Google Scholar] [CrossRef] [Green Version]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B.; Sussman, M.R. A peptide hormone and its receptor protein kinase regulate plant cell expansion. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [Green Version]
- Mashiguchi, K.; Sakaki, E.; Shimada, Y.; Nagae, M.; Ueno, K.; Nakano, T.; Yoneyama, K.; Suzuki, Y.; Asami, T. Feedback-regulation of strigolactone biosynthetic genes and strigolactone-regulated genes in Arabidopsis. Biosci. Biotechnol. Biochem. 2009, 73, 2460–2465. [Google Scholar] [CrossRef] [Green Version]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.E.; Rajashekar, C.B.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses. Role of phospholipase D alpha in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [Green Version]
- Noack, L.C.; Bayle, V.; Armengot, L.; Rozier, F.d.r.; Mamode-Cassim, A.; Stevens, F.D.; Caillaud, M.-C.c.; Munnik, T.; Mongrand, S.b.; Pleskot, R.; et al. A nanodomain-anchored scaffolding complex is required for the function and localization of phosphatidylinositol 4-kinase alpha in plants. Plant Cell 2021, 34, 302–332. [Google Scholar] [CrossRef] [PubMed]
- Mamode Cassim, A.; Gouguet, P.; Gronnier, J.; Laurent, N.; Germain, V.; Grison, M.; Boutté, Y.; Gerbeau-Pissot, P.; Simon-Plas, F.; Mongrand, S. Plant lipids: Key players of plasma membrane organization and function. Prog. Lipid Res. 2019, 73, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hu, M.; Yuan, D.; Yun, Z.; Gao, Z.; Su, Z.; Zhang, Z. Melatonin alleviates pericarp browning in litchi fruit by regulating membrane lipid and energy metabolisms. Postharvest Biol. Technol. 2020, 160, 111066. [Google Scholar] [CrossRef]
- Wong, A.; Donaldson, L.; Portes, M.T.; Eppinger, J.; Feijó, J.A.; Gehring, C. Arabidopsis DIACYLGLYCEROL KINASE4 is involved in nitric oxide-dependent pollen tube guidance and fertilization. Development 2020, 147. [Google Scholar] [CrossRef]
- Yao, S.; Peng, S.; Wang, X. Phospholipase Dε interacts with autophagy-related-protein 8 and promotes autophagy in Arabidopsis response to nitrogen deficiency. Plant J. 2022. [Google Scholar] [CrossRef]
- Holland, P.; Knævelsrud, H.; Søreng, K.; Mathai, B.J.; Lystad, A.H.; Pankiv, S.; Bjørndal, G.T.; Schultz, S.W.; Lobert, V.H.; Chan, R.B.; et al. HS1BP3 negatively regulates autophagy by modulation of phosphatidic acid levels. Nat. Commun. 2016, 7, 13889. [Google Scholar] [CrossRef]
- Su, Y.; Li, M.; Guo, L.; Wang, X. Different effects of phospholipase Dζ2 and non-specific phospholipase C4 on lipid remodeling and root hair growth in Arabidopsis response to phosphate deficiency. Plant J. Cell Mol. Biol. 2018, 94, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Duran, H.G.S.; van Haarst, J.C.; Schijlen, E.G.W.M.; Ruyter-Spira, C.; Medema, M.H.; Dong, L.; Bouwmeester, H.J. The role of strigolactones in P deficiency induced transcriptional changes in tomato roots. BMC Plant Biol. 2021, 21, 349. [Google Scholar] [CrossRef]
- Takáč, T.; Šamajová, O.; Vadovič, P.; Pechan, T.; Šamaj, J. Shot-gun proteomic analysis on roots of Arabidopsis pldα1 mutants suggesting the involvement of PLDα1 in mitochondrial protein import, vesicular trafficking and glucosinolate biosynthesis. Int. J. Mol. Sci. 2018, 20, 82. [Google Scholar] [CrossRef] [Green Version]
- Takáč, T.; Pechan, T.; Šamajová, O.; Šamaj, J. Proteomic analysis of Arabidopsis pldα1 mutants revealed an important role of phospholipase D alpha 1 in chloroplast biogenesis. Front. Plant Sci. 2019, 10, 89. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PA Target | PA-Binding Site | Effect on Activity | Role in Signaling | Lipid Signaling Pathway | Notes (PA Binding Specificity) | Refs. |
|---|---|---|---|---|---|---|
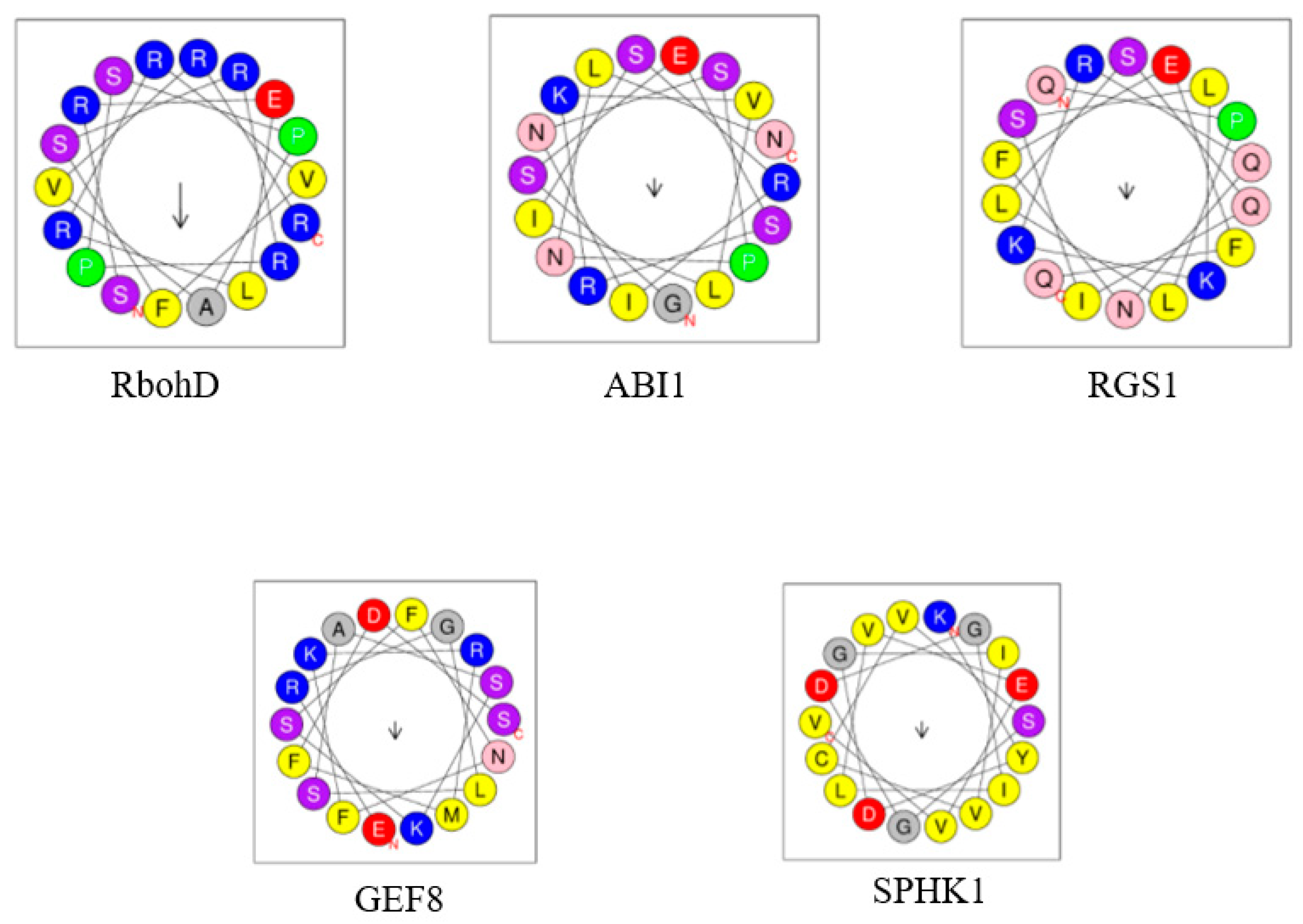
| AtRbohD (At5g47910), AtRbohF (At1g64060), respiratory burst oxidase homolog protein D and F | Arg149, Arg150, between EF-hands and N-terminus (in cytosolic region), in the vicinity of phosphorylation motif (AtRbohD); residues 105 to 341 (AtRbohF) | Stimulation at PM (AtRbohD) | ABA signaling, ROS generation, ROS-dependent NO production, stomatal closure | AtPLDα1 | di18:1PA, di18:2PA, 16:0–18:1 PA, 16:0–18:2PA, 18:0–18:2 PA), but not to di16:0PA, di18:0PA; AtRbohD has higher affinity to PA than AtRbohF | [60] |
| AtSPHK1 (At4g21540), AtSPHK2 (At4g21534), sphingosine kinases | VSGDGI motif (AtSPHK1) | Stimulation by promoting substrate binding | ABA signaling, AtPLDα1 activation (for amplification of ABA response), stomatal closure, seed germination, root elongation | AtPLDα1 | Strong binding to 18:1/18:1PA, 16:0/18:1PA, 16:0/18:2PA | [71,72] |
| AtRopGEF8 (At3g24620), guanine nucleotide exchange factor 8 | Lys13, Lys18 in AtGEF8 PRONE domain and N-terminus | Stimulation of AtGEF8 activity towards small GTPase ROP7 | ABA signaling activation | di18:2PA, di18:1PA, 16:0–18:1PA, 16:0–18:2PA, 16:0–18:1PA, 18:0–18:2PA | [61] | |
| AtRGS1 (At3g26090), regulator of G-protein signaling | Lys259 in RGS domain | Inhibition of GTPase-accelerating protein activity of AtRGS1 | ABA signaling activation, inhibition of seed germination, root growth and stomatal responses | - | - | [73] |
| AtMAP65-1 (At5g55230), a microtubule-associated protein | Lys and Arg from 53KRK55 and 61KSR63 sequences, Ser-428 in 428SK429 sequence, out of microtubule binding or phosphorylation regions | Promotion of AtMAP65-1-induced microtubule-bundling activity | Salt stress signaling enhances tubulin polymerization involving AtMAP65-1 increases salt tolerance by microtubule stabilization | AtPLDα1 | 16:0–18:1PA, 16:0–18:2PA, 18:0–18:1PA, 18:0–18:2PA, but not di18:0PA | [74] |
| AtMPK6 (At2g43790), a mitogen-activated protein kinase 6 | - | Stimulation | Salt stress signaling, salt tolerance by AtMPK6-dependent phosphorylation of AtSOS1 | AtPLDα1 | Significantly to di18:1PA, di18:2PA, 16:0–18:1PA, 16:0–18:2 PA, 18:0–18:2PA | [75] |
| AtMKK7 (At1g18350), AtMKK9 (At1g73500), a mitogen-activated protein kinase kinases | - | Activation, translocation to PM | Salt stress signaling, salt tolerance, maximization of the signal transduction efficiency | AtPLDα1 | Strongly to di18:1PA, 16:0–18:1PA, 16:0–18:2PA, 18:0–18:2PA | [76] |
| AtMPK3 (At3g45640)/ AtMPK6 (At2g43790), a mitogen-activated protein kinases | - | Stimulation, interaction with and phosphorylation of hypoxia-related transcription factor RAP2/12 | Hypoxia stress signaling, translocation to nucleus and stimulation of hypoxia-related transcription factor RAP2/12, feedback inhibition of PA production, tolerance to submergence | AtPLDα1, AtPLDδ | - | [77] |
| AtPINOID (At2g34650), a protein kinase | Lys119 to Lys121 | Stimulation, tethering to PM | Salt stress signaling, salt tolerance, enhanced AtPINOID-dependent AtPIN2 phosphorylation and PM localization, activation of PIN2-mediated auxin efflux and redistribution | AtPLDα1, AtPLDδ | di18:1PA, di18:2PA, 16:0–18:1PA, 16:0–18:2PA, 18:0–18:1PA, 18:0-18:2PA | [65] |
| AtSnRK2.4 (At1g10940) | - | Phosphorylation of SnRK2 targets, affects its interaction with PA and subcellular localization | - | - | 16:0/18:1 PA | [78] |
| OsAKT2 (Os05g35410), a potassium channel | Arg190/Lys191, Arg644/Arg645, Arg755/Arg756 in ANK domain, cytoplasmic domain, S4 voltage sensor | Inhibition | Seedlings growth under short-day conditions | - | Strongly to 16:0–18:2PA, much weaker to di16:0PA, di18:0PA, di18:1PA, di18:2PA, 16:0–18:1PA, 18:0–18:1PA, 18:0–18:2 PA | [67,79] |
| OsGID1 (Os05g33730), a gibberellin receptor | Arg79, Arg82 | Tethering to nucleus | Gibberellin signaling, promotion of OsSLR1 (a suppressor DELLA protein) degradation | OsPLDα6 | [80] |
| Bioactive Peptide | Receptor | Co-Expressed PA-Metabolizing Enzyme | Platform | Database Dataset | Statistical Notes (Score) |
|---|---|---|---|---|---|
| PSK | PSKR1 | PLDζ1 | AT_mRNASeq_ARABI | anatomy | 0.9 |
| PSK | PSKR1 | PLDγ1 | AT_AFFY_ATH1 | perturbation | 0.69 |
| PSK | PSKR1 | PLDγ1 | AT_AFFY_ATH1 | sample | 0.69 |
| CEP1 | CEPR1 | PI-PLC3 | AT_mRNASeq_ARABI | anatomy | 0.89 |
| CLE25 | BAM2 | NPC6 | AT_AFFY_ATH1 | sample | 0.68 |
| PIP1/2 | RLK7 | DGK5 | AT_mRNASeq_ARABI | perturbation | 0.85 |
| PIP1/2 | RLK7 | PLDγ1 | AT_mRNASeq_ARABI | sample | 0.79 |
| PEP | PEPR1 | DGK1 | AT_AFFY_ATH1 | sample | 0.56 |
| PEP | PEPR2 | DGK5 | AT_mRNASeq_ARABI | sample | 0.64 |
| EPF2 | TMM | NPC2 | AT_AFFY_ATH1 | development | 0.9 |
| LURE1 | MIDS1 | PI-PLC6 | AT_mRNASeq_ARABI | perturbation | 0.61 |
| LURE1 | MIK2 | PLDγ1 | AT_mRNASeq_ARABI | perturbation | 0.82 |
| LURE1 | PRK8 | PLDα2 | AT_AFFY_ATH1 | sample | 0.82 |
| LURE1 | PRK3 | DGK4 | AT_mRNASeq_ARABI | perturbation | 0.74 |
| LURE1 | PRK5 | DGK4 | AT_mRNASeq_ARABI | perturbation | 0.74 |
| GRIM REAPER | PRK5 | DGK4 | AT_mRNASeq_ARABI | perturbation | 0.74 |
| ESF1 | SSP | DGK4 | AT_AFFY_ATH1 | sample | 0.75 |
| RALF1 | BAK1 | PLDγ1 | AT_AFFY_ATH1 | perturbation | 0.73 |
| RALF1 | BAK1 | PLDγ1 | AT_mRNASeq_ARABI | sample | 0.8 |
| RALF1 | BAK1 | PLDγ1 | AT_mRNASeq_ARABI | anatomy | 0.85 |
| Ralf4/19 | BUPS1 | DGK6 | AT_AFFY_AG | sample | 0.9 |
| Ralf4/19 | BUPS1 | PI-PLC6 | AT_mRNASeq_ARABI | sample | 0.92 |
| Ralf4/19 | BUPS2 | PI-PLC6 | AT_mRNASeq_ARABI | sample | 0.95 |
| Ralf1/17/23/ 32/33 | FERONIA | NPC1 | AT_mRNASeq_ARABI | sample | 0.7 |
| Ralf1/17/23/ 32/33 | FERONIA | PLDα1 | AT_mRNASeq_ARABI | anatomy | 0.88 |
| Ralf1/17/23/ 32/33 | FERONIA | PLDα1 | AT_mRNASeq_ARABI | sample | 0.79 |
| Ralf1/17/23/ 32/33 | FERONIA | PLDα1 | AT_AFFY_ATH1 | sample | 0.5 |
| Ralf4/19/34 | ANXUR2 | PLDα2 | AT_AFFY_AGRONOMICS | sample | 0.83 |
| CLE45 | SKM1 | PLDε | AT_mRNASeq_ARABI | anatomy | 0.61 |
| CLE45 | SKM1 | PI-PLC5 | AT_mRNASeq_ARABI | anatomy | 0.57 |
| CLE42 | PXL2 | PLDδ | AT_mRNASeq_ARABI | perturbation | 0.71 |
| TPD1 | EMS1 | PLDε | AT_AFFY_AGRONOMICS | perturbation | 0.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolesnikov, Y.; Kretynin, S.; Bukhonska, Y.; Pokotylo, I.; Ruelland, E.; Martinec, J.; Kravets, V. Phosphatidic Acid in Plant Hormonal Signaling: From Target Proteins to Membrane Conformations. Int. J. Mol. Sci. 2022, 23, 3227. https://doi.org/10.3390/ijms23063227
Kolesnikov Y, Kretynin S, Bukhonska Y, Pokotylo I, Ruelland E, Martinec J, Kravets V. Phosphatidic Acid in Plant Hormonal Signaling: From Target Proteins to Membrane Conformations. International Journal of Molecular Sciences. 2022; 23(6):3227. https://doi.org/10.3390/ijms23063227
Chicago/Turabian StyleKolesnikov, Yaroslav, Serhii Kretynin, Yaroslava Bukhonska, Igor Pokotylo, Eric Ruelland, Jan Martinec, and Volodymyr Kravets. 2022. "Phosphatidic Acid in Plant Hormonal Signaling: From Target Proteins to Membrane Conformations" International Journal of Molecular Sciences 23, no. 6: 3227. https://doi.org/10.3390/ijms23063227