
Embedded Human Periodontal Ligament Stem Cells Spheroids Enhance Cementogenic Differentiation via Plasminogen Activator Inhibitor 1


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
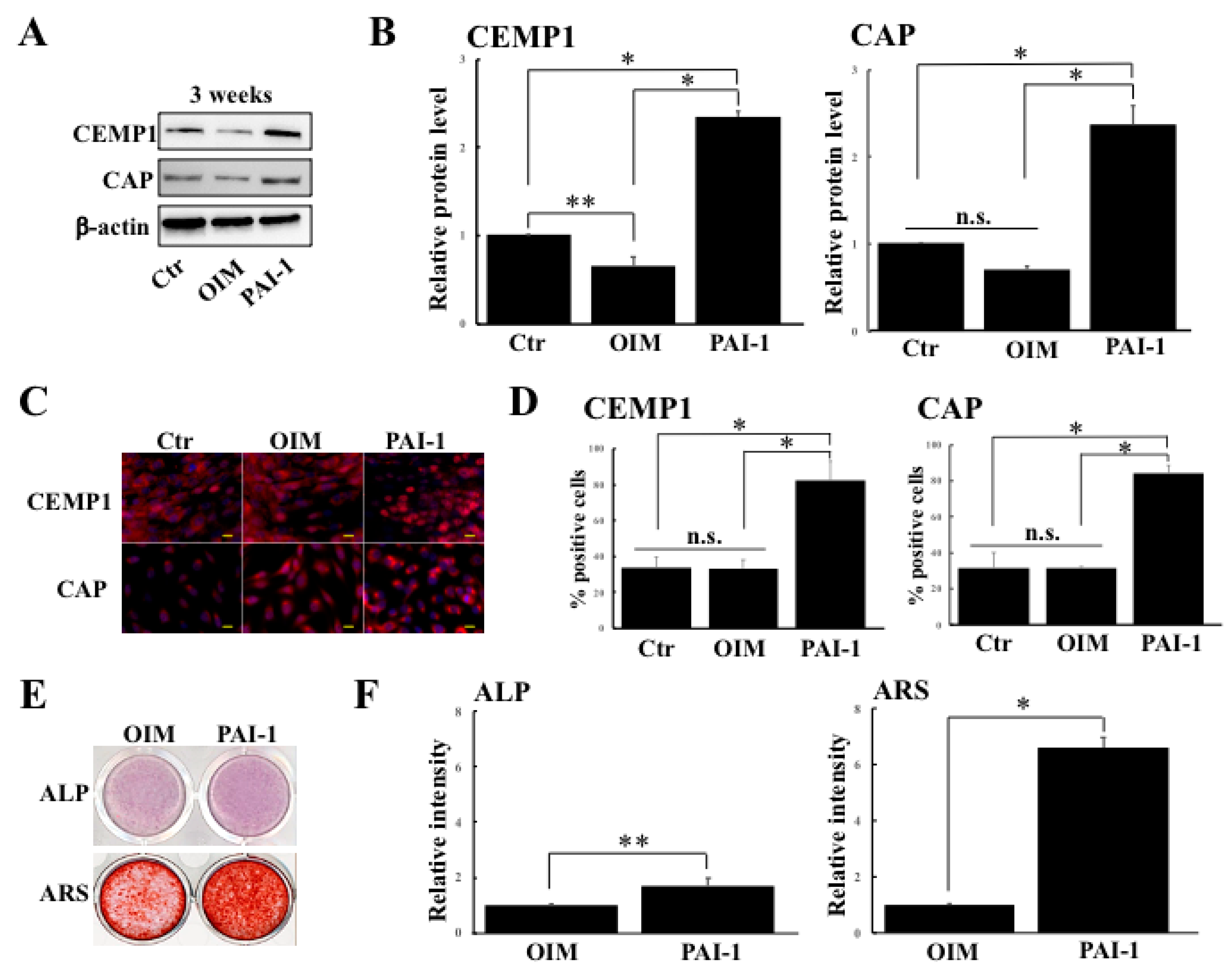
2.1. rhPAI-1 Enhances Cementogenic Differentiation of HPLSCs in 2D Culture
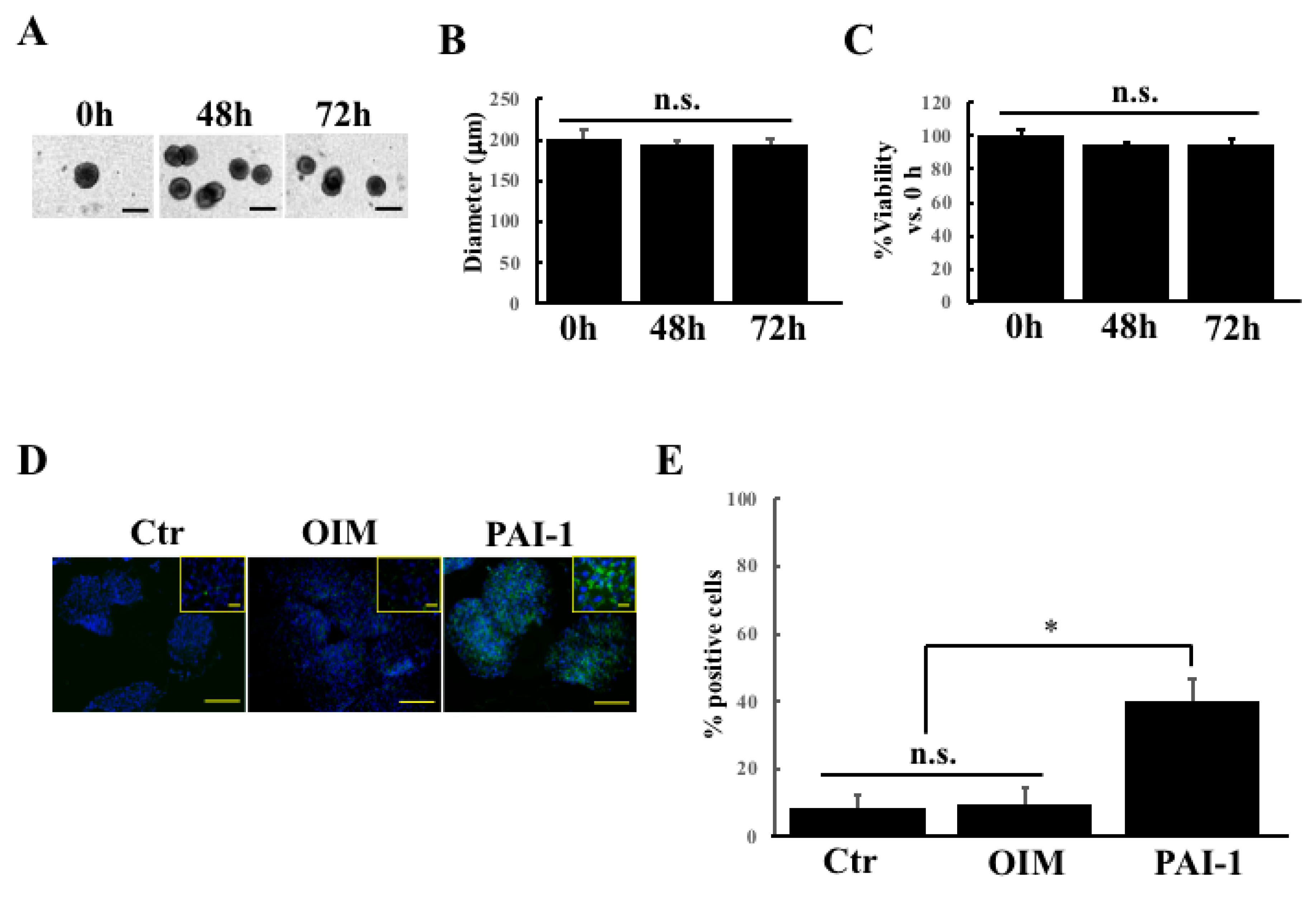
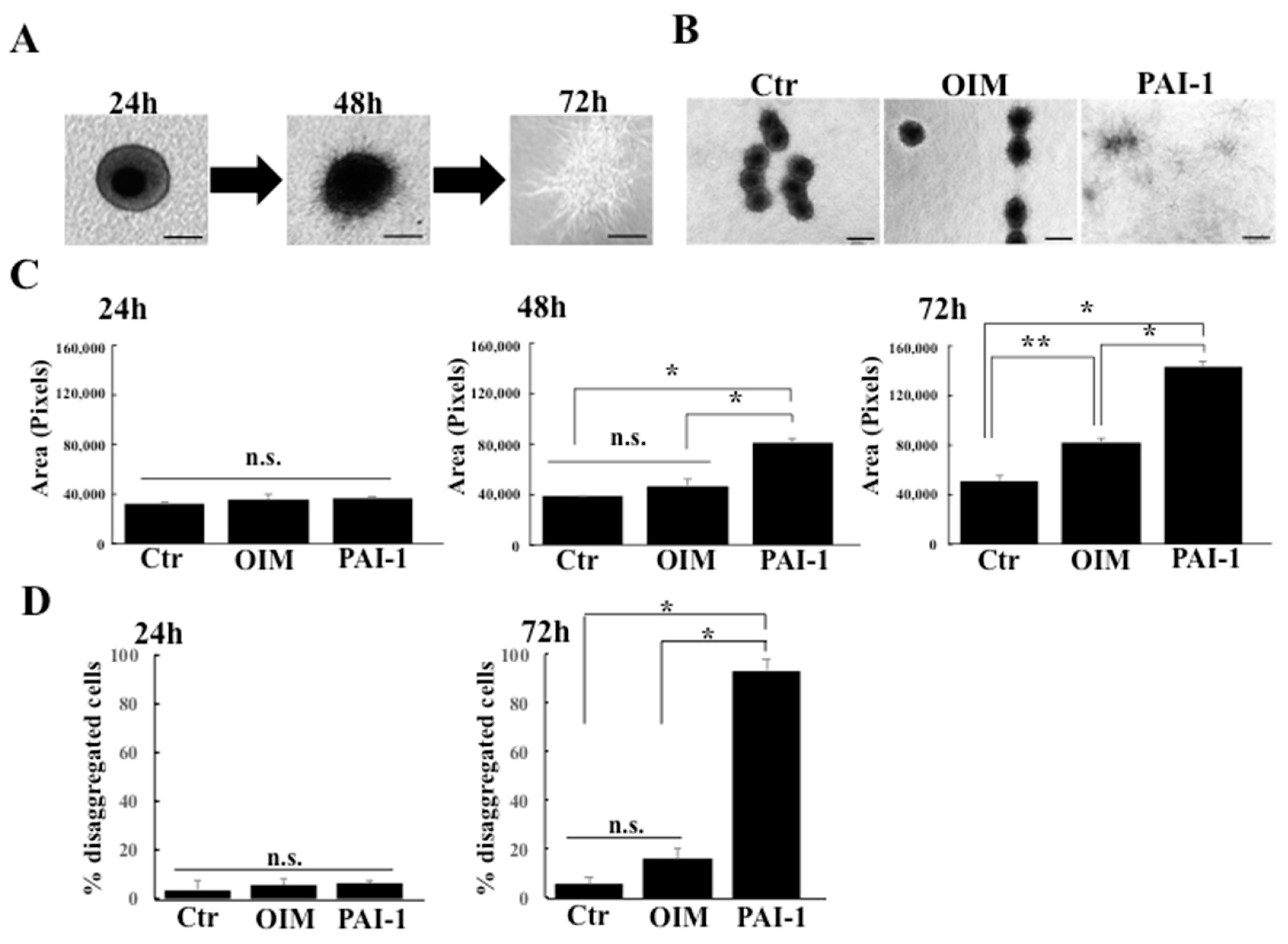
2.2. rhPAI-1 Induces Cementoblastic Spheroids Embedded in Collagen Gel
2.3. rhPAI-1 Promotes Interplay between HPLSC Spheroids and Collagenous ECM
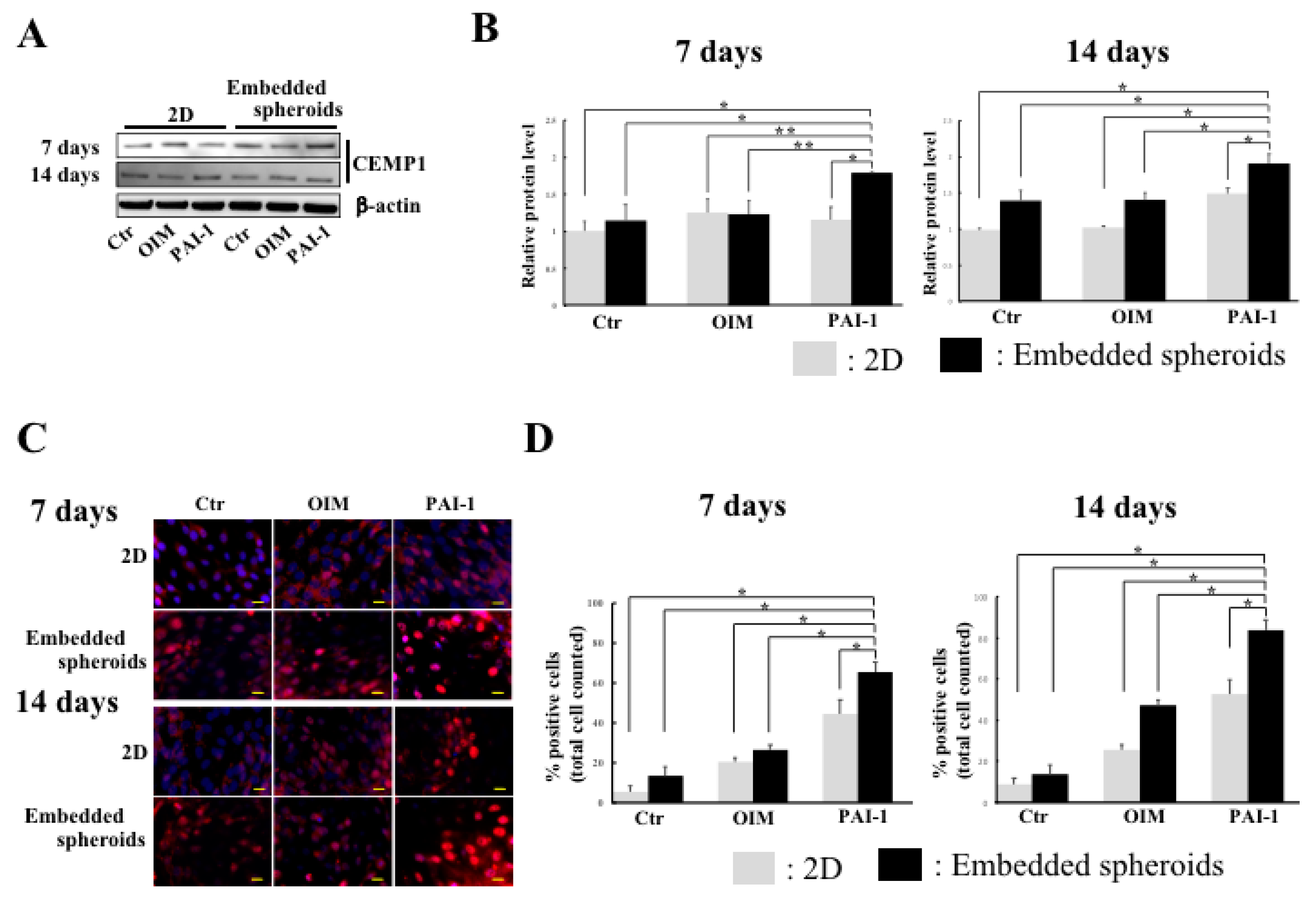
2.4. CEMP1 Expression Is Elevated in Embedded HPLSC Spheroids Treated with rhPAI-1
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Collagen-Embedded HPLSC Spheroids
4.3. Western Blot Analysis
4.4. Immunocytochemistry Analysis
4.5. Alkaline Phosphatase (ALP) and Alizarin Red S (ARS) Staining
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nanci, A.; Bosshardt, D.D. Structure of periodontal tissues in health and disease. Periodontology 2000, 2006, 11–28. [Google Scholar] [CrossRef]
- Hwang, Y.J.; Kolettis, N.; Yang, M.; Gillard, E.R.; Sanchez, E.; Sun, C.H.; Tromberg, B.J.; Krasieva, T.B.; Lyubovitsky, J.G. Multiphoton imaging of actin filament formation and mitochondrial energetics of human ACBT gliomas. Photochem. Photobiol. 2011, 87, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.P.; Hanna, W.; El-Hamidi, H.; Celli, J.P. Longitudinal measurement of extracellular matrix rigidity in 3D tumor models using particle-tracking microrheology. J. Vis. Exp. 2014, 88, e51302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobita, M.; Mizuno, H. Periodontal disease and periodontal tissue regeneration. Curr. Stem Cell Res. Ther. 2010, 5, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Aldahmash, A.; Zaher, W.; Al-Nbaheen, M.; Kassem, M. Human stromal (mesenchymal) stem cells: Basic biology and current clinical use for tissue regeneration. Ann. Saudi. Med. 2012, 32, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Hang, K.; Ying, L.; Bai, J.; Wang, Z.; Kuang, Z.; Xue, D.; Pan, Z. Knockdown of SERPINB2 enhances the osteogenic differentiation of human bone marrow mesenchymal stem cells via activation of the Wnt/b-catenin signalling pathway. Stem. Cell Res. Ther. 2021, 12, 525. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Jin, H.; Kim, J.Y.; Lim, K.T.; Choung, H.W.; Park, J.Y.; Chung, J.H.; Choung, P.H. Hypoxia promotes CEMP1 expression and induces cementoblastic differentiation of human dental stem cells in an HIF-1-dependent manner. Tissue Eng. Part A 2014, 20, 410–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Choung, H.W.; Lim, K.T.; Jin, B.; Jin, C.; Chung, J.H.; Choung, P.H. Recombinant human plasminogen activator inhibitor-1 promotes cementogenic differentiation of human periodontal ligament stem cells. Tissue Eng. Part A 2015, 21, 2817–2828. [Google Scholar] [CrossRef] [Green Version]
- Fujii, S.; Maeda, H.; Wada, N.; Tomokiyo, A.; Saito, M.; Akamine, A. Investigating a clonal human periodontal ligament progenitor/stem cell line in vitro and in vivo. J. Cell Physiol. 2008, 215, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, P.; Yu, Q.T.; Tran, T.; Bhatti, F.U.; Zhu, X.; Huang, G.T. Cementogenic genes in human periodontal ligament stem cells are downregulated in response to osteogenic stimulation while upregulated by vitamin C treatment. Cell Tissue Res. 2017, 368, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I. Adult mesenchymal stem cells: When, where, and how. Stem. Cells Int. 2015, 2015, 628767. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shen, H.; Zheng, W.; Tang, L.; Yang, Z.; Gao, Y.; Yang, Q.; Wang, C.; Duan, Y.; Jin, Y. Characterization of stem cells from alveolar periodontal ligament. Tissue Eng. Part A 2011, 17, 1015–1026. [Google Scholar] [CrossRef]
- Yasunaga, M.; Kajiya, H.; Toshimitsu, T.; Nakashima, H.; Tamaoki, S.; Ishikawa, H.; Maeda, H.; Ohno, J. The early autophagic pathway contributes to osteogenic differentiation of human periodontal ligament stem cells. J. Hard Tissue Biol. 2019, 28, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Arzate, H.; Jiménez-García, L.F.; Alvarez-Pérez, M.A.; Landa, A.; Bar-Kana, I.; Pitaru, S. Immunolocalization of a human cementoblastoma-conditioned medium-derived protein. J. Dent. Res. 2002, 81, 541–546. [Google Scholar] [CrossRef]
- Alvarez-Pérez, M.A.; Narayanan, S.; Zeichner-David, M.; Rodríguez Carmona, B.; Arzate, H. Molecular cloning, expression and immunolocalization of a novel human cementum-derived protein (CP-23). Bone 2006, 38, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Barkana, I.; Narayanan, A.S.; Grosskop, A.; Savion, N.; Pitaru, S. Cementum attachment protein enriches putative cementoblastic populations on root surfaces in vitro. J. Demt. Res. 2000, 79, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Xu, Y.; Qi, Y.; Wang, X.; Patel, D.K.; Dutta, S.D.; Chen, R.; Lim, K.T. Evaluation of osteogenic/ cementogenic modulating potential of PAI-1 transfected media for stem cells. IEEE Trans. Nanobiosci. 2020, 19, 446–456. [Google Scholar] [CrossRef]
- Liu, R.M. Oxidative stress, plasminogen activator inhibitor 1, and lung fibrosis. Antioxid. Redox. Signal 2008, 10, 303–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, D.E.; De Taeye, B.M.; Eren, M. PAI-1 antagonists: Predictable indications and unconventional applications. Curr. Drug Targets 2007, 8, 962–970. [Google Scholar] [CrossRef]
- Lundgren, C.H.; Brown, S.L.; Nordt, T.K.; Sobel, B.E.; Fujii, S. Elaboration of type-1 plasminogen activator inhibitor from adipocytes. A potential pathogenetic link between obesity and cardiovascular disease. Circulation 1996, 93, 106–110. [Google Scholar] [CrossRef]
- Stefansson, S.; McMahon, G.A.; Petitclerc, E.; Lawrence, D.A. Plasminogen activator inhibitor-1 in tumor growth, angiogenesis and vascular remodeling. Curr. Pharm. Des. 2003, 9, 1545–1564. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, J.D.; Sappino, A.P.; Belin, D. The plasminogen activator/plasmin system. J. Clin. Investig. 1991, 88, 1067–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordstrom, S.M.; Carleton, S.M.; Carson, W.L.; Eren, M.; Phillips, C.L.; Vaughan, D.E. Transgenic over-expression of plasminogen activator inhibitor-1 results in age-dependent and gender-specific increases in bone strength and mineralization. Bone 2007, 41, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.S.; Tseng, C.S.; Linju Yen, B.; Dai, L.G.; Hsieh, P.S.; Hsu, S.H. Solid freeform-fabricated scaffolds designed to carry multicellular mesenchymal stem cell spheroids for cartilage regeneration. Eur. Cell Mater. 2013, 26, 179–194. [Google Scholar] [CrossRef]
- Hutmacher, D.W. Biomaterials offer cancer research the third dimension. Nat. Mater. 2010, 9, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Teoh, S.H.; Hui, J.H.; Fisk, N.M.; Choolani, M.; Chan, J.K. The potential of human fetal mesenchymal stem cells for off-the-shelf bone tissue engineering application. Biomaterials 2012, 33, 2656–2672. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrazzo, P.; Pizzuti, V.; Zia, S.; Sargenti, A.; Gazzola, D.; Roda, B.; Bonsi, L.; Alviano, F. Microfluidic tools for enhanced characterization of therapeutic stem cells and prediction of their potential antimicrobial secretome. Antibiotics 2021, 10, 750. [Google Scholar] [CrossRef]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H. The third dimension bridges the gap between cell culture and live tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef]
- Martin, K.J.; Patrick, D.R.; Bissell, M.J.; Fournier, M.V. Prognostic breast cancer signature identified from 3D culture model accurately predicts clinical outcome across independent datasets. PLoS ONE 2008, 3, e2994. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Jeon, S.H.; Choung, P.H. Efficacy of periodontal stem cell transplantation in the treatment of advanced periodontitis. Cell Transpl. 2011, 20, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of osteoblast differentiation by Runx2. Adv. Exp. Med. Biol. 2010, 658, 43–49. [Google Scholar]
- Tevis, K.M.; Colson, Y.L.; Grinstaff, M.W. Embedded spheroids as models of the cancer microenvironment. Adv. Biosyst. 2017, 1, 1700083. [Google Scholar] [CrossRef]
- Gole, B.; Durán Alonso, M.B.; Dolenc, V.; Lah, T. Post-translational regulation of cathepsin B, but not of other cysteine cathepsins, contributes to increased glioblastoma cell invasiveness in vitro. Pathol. Oncol. Res. 2009, 15, 711–723. [Google Scholar] [CrossRef]
- Bell, H.S.; Wharton, S.B.; Leaver, H.A.; Whittle, I.R. Effects of N-6 essential fatty acids on glioma invasion and growth: Experimental studies with glioma spheroids in collagen gels. J. Neurosurg. 1999, 91, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Tevis, K.M.; Cecchi, R.J.; Colson, Y.L.; Grinstaff, M.W. Mimicking the tumor microenvironment to regulate macrophage phenotype and assessing chemotherapeutic efficacy in embedded cancer cell/macrophage spheroid models. Acta Biomater. 2017, 50, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vörsmann, H.; Groeber, F.; Walles, H.; Busch, S.; Beissert, S.; Walczak, H.; Kulms, D. Development of a human three-dimensional organotypic skin-melanoma spheroid model for in vitro drug testing. Cell Death Dis. 2013, 4, e719. [Google Scholar] [CrossRef] [PubMed]
- Santini, M.T.; Rainaldi, G.; Indovina, P.L. Apoptosis, cell adhesion and the extracellular matrix in the three-dimensional growth of multicellular tumor spheroids. Crit. Rev. Oncol. Hematol. 2000, 36, 75–87. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasunaga, M.; Ishikawa, H.; Tamaoki, S.; Maeda, H.; Ohno, J. Embedded Human Periodontal Ligament Stem Cells Spheroids Enhance Cementogenic Differentiation via Plasminogen Activator Inhibitor 1. Int. J. Mol. Sci. 2022, 23, 2340. https://doi.org/10.3390/ijms23042340
Yasunaga M, Ishikawa H, Tamaoki S, Maeda H, Ohno J. Embedded Human Periodontal Ligament Stem Cells Spheroids Enhance Cementogenic Differentiation via Plasminogen Activator Inhibitor 1. International Journal of Molecular Sciences. 2022; 23(4):2340. https://doi.org/10.3390/ijms23042340
Chicago/Turabian StyleYasunaga, Madoka, Hiroyuki Ishikawa, Sachio Tamaoki, Hidefumi Maeda, and Jun Ohno. 2022. "Embedded Human Periodontal Ligament Stem Cells Spheroids Enhance Cementogenic Differentiation via Plasminogen Activator Inhibitor 1" International Journal of Molecular Sciences 23, no. 4: 2340. https://doi.org/10.3390/ijms23042340