
Roles of E3 Ubiquitin Ligases in Plant Responses to Abiotic Stresses
by
 ,
,
Shuang Wang
1,†, 
Xiaoyan Lv
2,†,
Jialin Zhang
1,
Daniel Chen
3,
Sixue Chen
4,
Guoquan Fan
5,
Chunquan Ma
1,* and
Yuguang Wang
1,* 1
Engineering Research Center of Agricultural Microbiology Technology, Ministry of Education, Heilongjiang University, Harbin 150080, China
2
School of Life Science and Technology, Harbin Institute of Technology, Harbin 150080, China
3
Judy Genshaft Honors College and College of Arts and Sciences, University of South Florida, Tampa, FL 33620, USA
4
Plant Molecular and Cellular Biology Program, Department of Biology, Genetics Institude, University of Florida, Gainesville, FL 32610, USA
5
Industrial Crops Institute, Heilongjiang Academy of Agricultural Sciences, Harbin 150086, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2022, 23(4), 2308; https://doi.org/10.3390/ijms23042308
Submission received: 20 January 2022
/
Revised: 13 February 2022
/
Accepted: 16 February 2022
/
Published: 19 February 2022
(This article belongs to the Special Issue Molecular Aspects of Plant Salinity Stress and Tolerance)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Plants are frequently exposed to a variety of abiotic stresses, such as those caused by salt, drought, cold, and heat. All of these stressors can induce changes in the proteoforms, which make up the proteome of an organism. Of the many different proteoforms, protein ubiquitination has attracted a lot of attention because it is widely involved in the process of protein degradation; thus regulates many plants molecular processes, such as hormone signal transduction, to resist external stresses. Ubiquitin ligases are crucial in substrate recognition during this ubiquitin modification process. In this review, the molecular mechanisms of plant responses to abiotic stresses from the perspective of ubiquitin ligases have been described. This information is critical for a better understanding of plant molecular responses to abiotic stresses.
1. Ubiquitination Modification
Proteins are not only structural molecules but also action molecules in all life forms. The lifespans of protein molecules in a cell range from less than a minute to many days [1]; thus, protein production and degradation play a fundamental role in all cells during plant growth, development, and responses to environmental changes. By regulating the abundance of key proteins plants are able to modulate signaling events, ensuring that the proper response is initiated when required and only for the appropriate length of time [2]. Protein degradation requires the degradation of signals—N-degrons/C-degrons—including not only adjacent sequence motifs but also internal lysine residues modified by polyubiquitin. All 20 amino acids function as destabilizing N-terminal residues, which complicates the N-degron pathways [1].
The endoplasmic reticulum-associated protein degradation (ERAD) pathway [3,4], ubiquitin proteasome system (UPS) that degrades ubiquitinated proteins via 26S proteasome [5], and the lysosome-mediated intracellular degradation pathway [6] are all related to ubiquitination modification. Ubiquitination is a multistep process governed by ubiquitin-activating enzymes (E1s), ubiquitin-conjugating enzymes (E2s), and ubiquitin-ligase enzymes (E3s) that successively ligate ubiquitin to substrate proteins [7]. Due to conservation, the constantly high expression levels of ubiquitin in different tissues and organs allow it to be utilized as a housekeeping gene marker for gene expression analysis in plants [8]. This also highlights the prevalence of ubiquitinated proteins in the proteome as a major proteoform [9,10].
A single ubiquitin can be attached to one (monoubiquitination) or multiple (multimonoubiquitination) lysine residues within the substrate protein. Alternatively, repetition of the conjugation process can generate a polyubiquitin chain on a single lysine of the substrate (polyubiquitination) [2]. After the enrichment of ubiquitinated peptides [11,12,13], the lysine sites modified by ubiquitin can be identified by mass spectrometry and immunological methods [14,15]. In addition, mass spectrometry has been used to achieve the quantification of ubiquitin levels [16,17]. However, the quantitative study of ubiquitin in plant cells has not been reported. Different ubiquitin pools were quantitatively analyzed in HEK293 and MEF cell lines, mouse brains, and human frontal cortexes through differential affinity chromatography combined with protein standard absolute quantification (PSAQ) mass spectrometry technology (Ub-PSAQ). The results showed that the proportional distribution of ubiquitin, ‘free’ ubiquitin, ubiquitin chains, and monoubiquitinated-modified conjugates were different in different cell types [15]. In addition, under the treatment of proteasome inhibitor MG-132, the proportions of the ubiquitin pools changed [12]. Studies in different model cells have shown that the redistribution mechanism of ubiquitin pools may be different, including de novo synthesis of ubiquitin and the transformation of multi-ubiquitin chains to mono-ubiquitin chains [15].
Ubiquitin has seven lysine residues (K6, K11, K27, K29, K33, K48, and K63) that provide sites for the formation of different isopeptide chain linkages. In addition, the free amino group of the N-terminal methionine (Met1) of ubiquitin can also be modified by other free ubiquitin molecules in tandem, which further increases the diversity and complexity of ubiquitin chains. A polyubiquitin chain can be homogeneous when the same lysine residue is used to build the polymer, or of mixed topology when different lysine residues are used to create the ubiquitin–ubiquitin linkage [18]. The Lys48-linked chains are the predominant linkage type in cells [19]. Pproteins modified by K48 will be degraded; however, Lys63 chains are non-degradable modification chains. Additionally, there are some atypical ubiquitin modifications on different sites of amino acid residues (Met1, Lys6, Lys11, Lys27, Lys29, and Lys33) [20]. Ubiquitin is also modified by small chemically-distinct post-translational modifications (PTMs), such as phosphorylation and acetylation. Six out of the seven lysine residues in ubiquitin can become acetylated [21]. There are multiple sites on ubiquitin that can be phosphorylated, such as Thr7, Thr12, Thr14, Ser20, Thr22, Thr55, Thr67, Tyr59, and Ser59. Moreover, multiple ubiquitin lysine residues (Lys6, Lys11, Lys27, Lys48, and Lys63) can be targeted for SUMOylation [22]. Compared to other PTMs, ubiquitination is a relatively complicated modification in the cells.
2. Classification and Functions of E3 Ubiquitin Ligases
Plants engage various regulations at the levels of transcription, translation, and post-translational modifications to mediate stress perception, signaling, and responses [2]. PTMs are at the heart of many cellular signaling events [23]. Protein ubiquitination is one of the most prevalent PTMs as it regulates a plethora of cellular processes in distinct manners [24] and orchestrates a spectrum of different cellular processes, including substrate degradation, protein localization, and enzyme activation or inactivation. Ubiquitin ligases regulate protein abundance to ensure that the stress response is initiated only when required, maintained at an appropriate intensity and eliminated once it is no longer needed [25].
Ubiquitination begins with the activation of ubiquitin by E1 followed by the transfer of ubiquitin to E2, whose active-site cysteine forms a thioester bond with the C-terminal carboxyl group of ubiquitin. Substrate-recruiting E3 interacts with the E2–ubiquitin (E2–Ub) intermediate, allowing for the transfer of ubiquitin to the target [18,26]. The specific combination of E2 and E3 enzymes recruited to a substrate dictates the chain linkage type [27]. At the end of the three-enzyme cascade, E3 exhibits strict control over both the efficiency and substrate specificity of the ubiquitination reaction [26].
Ubiquitin ligase E3 has varying isoforms, which can be divided into the following categories according to its different catalytic domains: RING (really interesting new gene), HECT (homology to E6-associated carboxyl terminus), and U-box [2,18]. At the same time, HECT, U-box, and RING all belong to a single subunit of E3, while the multi-subunit refers to an enzyme with F-box sequence characteristics [28]. RING E3 is one of the key types of ubiquitin ligase. The RING-finger domain exists not only in the single subunit RING ubiquitin ligase and RBR ubiquitin ligase (ring between ring) [29,30] but also in the multi-subunit cullin-RING E3 [31]. The U-box domain is similar to the RING domain in that it is essentially a modified RING-finger domain [32]. The U-box domain is a domain of about 70 amino acid residues in both lower and higher organisms [33]. The corresponding proline residue of S. cerevisiae UFD2 proline 924 is completely conserved in the U-box proteins of mammals and other organisms [34]. Hatakeyama, S. et al. demonstrated that conserved proline is essential for the function of U-box proteins because mutation of the U-box domain (P1140A) abolishes the E3 activity of UFD2a [34]. Therefore, U-box ligase E3 is considered as the third ubiquitin ligase E3 isoform besides RING and HECT. As with RING ligases, U-box ligases also do not form E3–Ub intermediates [33]. There are 2, 21, and 77 U-box proteins in yeast, humans, and rice, respectively [34,35,36]. An increasing number of U-box proteins indicates not only their importance in governing cellular processes that are specific to plants but also the wide range of functional involvement that these proteins could have as part of regulated plant growth and development [32]. RBR is a single subunit ubiquitin ligase containing two RING domains, one of which contains cysteine residues, which can bind to ubiquitin to form an E3–Ub intermediate [37].
As a key enzyme in ubiquitination modification, ubiquitin ligase plays a role in determining substrate specificity. E3 can recognize substrates through specific sequences, such as the APC/C complex recognition D-box motif [38], and the KEN-box and HECT family recognition PY motifs of substrates [39]. E3 can also recognize substrates through adaptors. For example, the NEDD4 family ubiquitin ligase SMURF1 uses the adaptor Smad7 to bind TGFβ and mediate the ubiquitination degradation of TGFβ [40].
3. Ubiquitin Ligases in Plant Abiotic Stresses
As sessile organisms, plants have evolved a variety of complex adaptive mechanisms to cope with adverse environmental conditions, including the maintenance of ion homeostasis, accumulation of antioxidant enzymes, and synthesis of compatible products [41,42,43]. Furthermore, stress-related cis-acting elements or transcription factors (TFs) as well as stress response genes can be activated in plant stress responses and adaptations. Abiotic stressors, such as soil salinity, drought, or extreme temperature variations, impair crop productivity and are therefore the main causes of reduced crop yield [44,45].
3.1. Salt Stress
Human population increase, dysfunctional drainage, and irrigation-aggravated soil salinization [46] cause there to be a high concentration of Na+ in the soil, leading to saline–alkaline land. This hyperosmotic condition hinders the absorption of water by plants and nutrients in the soil, which results in a significant decline in crop productivity [47,48]. Salt stress has both osmotic and ionic or ion-toxicity effects on cells [49]. Proteins critical for salt-stress signaling, ion and water transport, redox homeostasis, and metabolism are regulated to bring about ionic and water homeostasis and cellular stability under salt stress [49].
3.1.1. E3s Participate in the SOS Pathway and MAPK Cascade
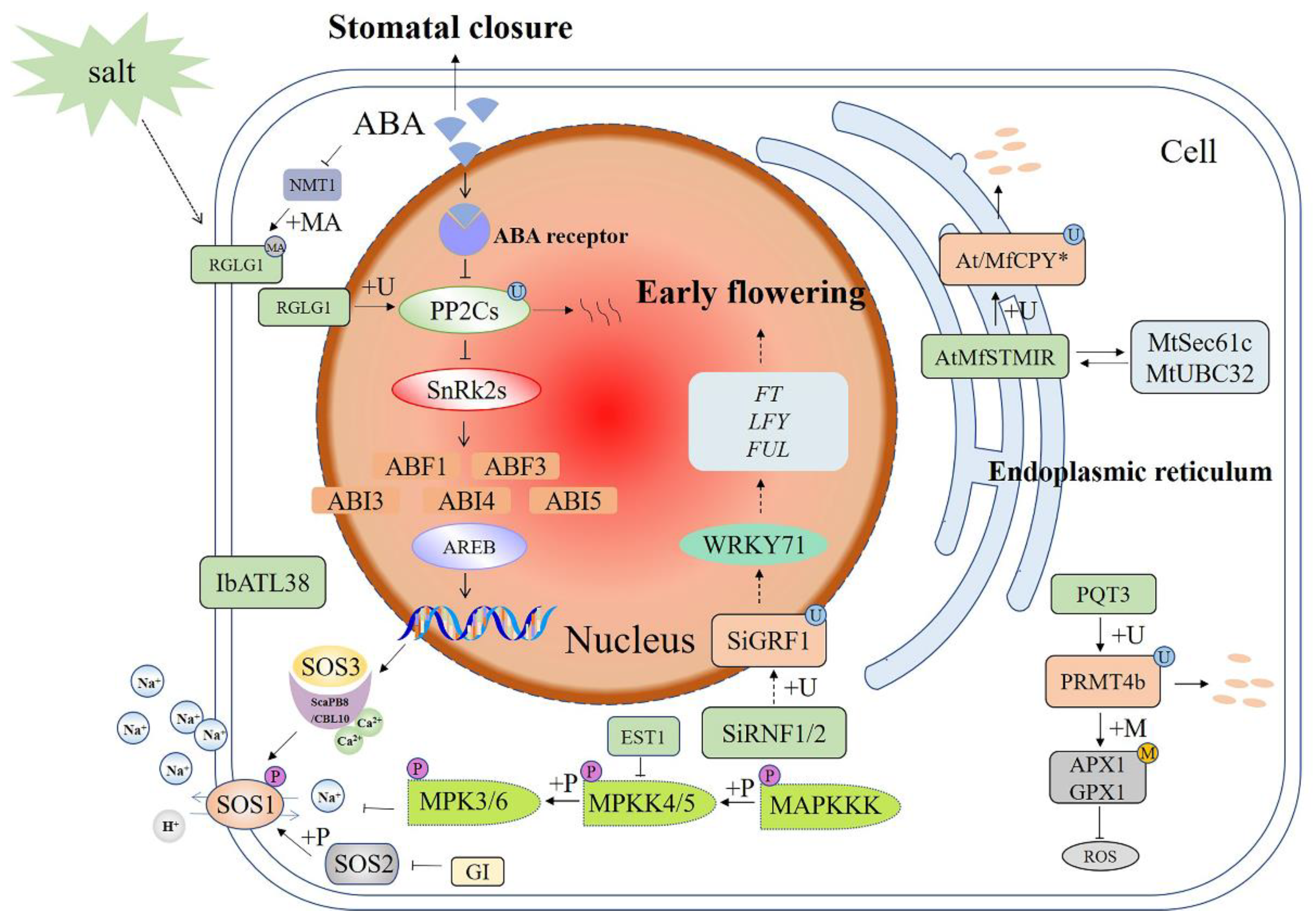
The salt overly sensitive (SOS) pathway is a well known signaling module that controls cellular ion homoeostasis (Figure 1) [50,51]. During salt stress, extracellular Na+ induces a transient rise in the cytosolic Ca2+ concentration, which triggers the SOS pathway [52]. A calcium-derived signal activates the SOS pathway by binding to the SOS3 and ScaPB8/CBL10 calcium-binding proteins, which activate the SOS2 protein kinase to regulate the SOS1, a plasma membrane (PM) Na+/H+ antiporter. SOS2 is a key regulator in the SOS pathway, relaying the signal downstream through changes in protein phosphorylation [52,53]. Interestingly, a flowering time regulator, GIGANTEA (GI) can prevent SOS2 from phosphorylating SOS1 in the absence of salt stress [54]. This mechanism may allow plants to maintain regular growth and development programs under adverse circumstances. In addition to the SOS pathway, the mitogen-activated protein kinase (MAPK) cascade functions during biotic and abiotic stress responses by receiving extracellular signals and activating the expression of downstream target genes [55]. A MAPK cascade is minimally composed of distinct combinations of at least three protein kinases: a MAPKKK (MAP3K/ MEKK/MKKK), a MAPKK (MKK/MEK), and a MAPK (MPK), which form a cascade of activation via transphosphorylation in a sequential manner [56,57].
The SOS and MAPK pathways are closely regulated by protein ubiquitination. EST1 (Ever shorter Telomeres 1) encodes an F-box protein, which, as a subunit of SCF E3 ubiquitin ligase, negatively regulates Arabidopsis in response to salt stress [58]. The salt-tolerant phenotype of the est1 mutant is also dependent on the function of SOS1, which is downstream of EST1. However, there is no direct interaction between SOS1 and EST1. On the contrary, EST1 directly interacts with MKK4 and negatively regulates its protein level, which may lead to decreased activity of the MKK4-MPK6 cascade reaction, resulting in decreased activity of the Na+/H+ antiporter. Compared with the wild type (WT), the est1 mutant showed higher PM Na+/H+ antiporter activity, resulting in lower intracellular Na+ concentration, which led to lower Na+ accumulation during salt stress, and thus higher salt tolerance [58]. Another example was E3 ubiquitin ligase IbATL38 (Ipomoea batatas Arabidopsis Toxicos en Levadura 38), which is localized to the nucleus and cell membrane. The expression pattern of IbATL38 was different under different stress treatments and the expression was induced under salt and ABA treatment. Under salt stress, A. thaliana plants overexpressing IbATL38 showed a trend of improved growth and increased expression of stress response genes such as AtSOS1, AtSOS2, AtSOS3, AtRD29A, and AtKIN2. The results suggested that IbATL38 was involved in plant responses to salt stress as a positive regulatory factor [59].
3.1.2. E3s Participate in the ABA Signaling Pathway
Under normal growth conditions, the E3 ubiquitin ligase RGLG (RING domain ligase) is localized in the PM [60]. The myristoyltransferase NMT1 is decreased under ABA or salt stress conditions. This in turn inhibits the myristoylation modification of RGLG, resulting in the migration of RGLG to the nucleus. This change of subcellular localization enables its interaction with PP2C in the nucleus, leading to degradation of ubiquitinated PP2C and thus turning on the ABA pathway [61] (Figure 1).
3.1.3. E3s Participate in the Flowering Pathway
In plants, 14-3-3 proteins are recognized as mediators of signal transduction and function in both plant development and stress response [62]. A foxtail millet 14-3-3 protein SiGRF1 (Setaria italica growtn-regulating factor 1) is involved in flowering under salt stress and ubiquitinated by E3 ubiquitin ligase SiRNF1/2 (Setaria italica RING finger protein 1/2). The SiGRF1 gene may regulate the initiation date of flowering in plants exposed to salt stress by up-regulating the transcription level of WRKY71 to promote Flowering Locus T (FT) and LEAFY (LFY) expression to act against the inhibition by DELLAs, etc. Additionally, SiGRF1 helps to avoid salt stress by accelerating plant flowering time [63].
3.1.4. E3s Participate in ROS Homeostasis
A high concentration of reactive oxygen species (ROS) causes a direct impact upon biological membranes, disrupts macromolecules, promotes cell senescence, and induces irreversible cellular damage [64]. Two eminent types of defensive responses are enzymatic response and non-enzymatic response [65]. In Arabidopsis, protein arginine methyltransferase 4b (PRMT4b) can methylate histones on the chromatin of an A. thaliana ascorbate peroxidase 1 (AtAPX1) gene and a glutathione peroxidase 1 (AtGPX1) gene, consequently increasing their expression and leading to enhanced stress resistance [66]. On the other hand, PRMT4b interacts with the E3 ubiquitin ligase paraquat tolerance 3 (PQT3), resulting in the degradation of PRMT4b through the 26S proteasome. Therefore, PQT3 is regarded as a negative regulator through indirect regulation of antioxidant enzymes [67]. OsPQT3 is a homologous gene of AtPQT3, which also plays a negative regulatory role in plant resistance to paraquat and salt stress. Compared with the WT, the Ospqt3 mutant had higher APX, GPX, and SOD activity, as well as less ROS accumulation [64]. The study on the salt tolerance of rice seedlings and vegetative growth showed that the survival rate, seed setting rate, and tillering rate of the mutant were higher than those of the WT after salt stress treatment, indicating that the mutant was more resistant to salt stress [68].
3.1.5. E3s Participate in the ERAD Pathway
Previous studies in A. thaliana have found that the E3 ubiquitin ligase MfSTMIR (Medicago falcata salt tunicamycin-induced RING finger protein), an endoplasmic reticulum (ER) protein, ubiquitinates MfCPY*, which contains the G-to-R mutation at residue 255 [69]. MfSTMIR can interact with the E2 MtUBC32 (ubiquitin-conjugating enzyme 32) and Sec61-translocon subunit MtSec61c that can bind proteasome 19S regulatory particle, which can extract an ERAD substrate from the ER. However, MfSTMIR does not degrade Sec61c as a ubiquitination substrate, which helps reduce stress on the ER while under salt stress. Therefore, MfSTMIR can act as a positive regulator in response to salt stress [70].
3.2. Drought Stress
3.2.1. E3s Participate in DREB2A-Mediated Stress Signaling
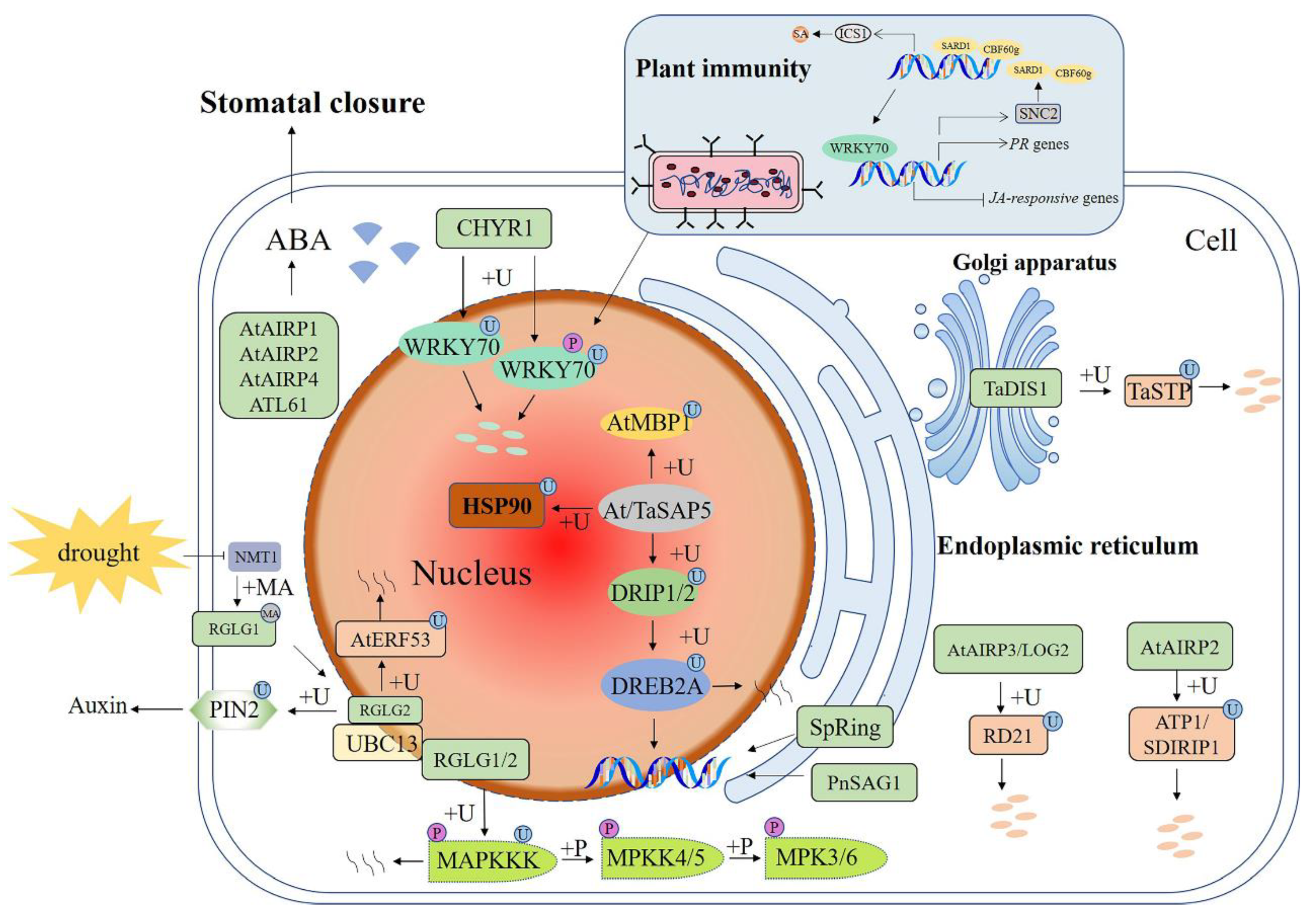
Drought and salt have overlapping signals and both result in hyperosmotic stress and accumulation of phytohormone abscisic acid (ABA) [41] (Figure 1 and Figure 2). Dehydration-responsive element-binding protein 2A (DREB2A) is a key transcriptional activator that induces transcription of many drought-response genes [71]. In wheat and Arabidopsis, DREB2A interacting protein 1 (DRIP1) and DRIP2 ubiquitinate DREB2A, resulting in its degradation by the 26S proteasome, which keeps it at a very low level. Under drought stress, TaSAP5 (Triticum aestivum stress-associated protein) acts as an E3 ubiquitin ligase to mediate DRIP [72] and HSP90C (chloroplast heat shock protein 90) ubiquitination [73] and degradation. Thereby, ubiquitination of DREB2A by DRIPs is decreased, leading to sufficient accumulation of DREB2A to initiate the expression of downstream genes. Thus, it can be concluded that SAP5 plays a role as a positive regulator in plant drought responses (Figure 2).
3.2.2. E3s Participate in MAPK Cascades
Another example of the interplay between drought and salt stress is E3 ubiquitin ligase RGLG (Figure 1 and Figure 2). Under drought stress, the modification of RGLG was also affected, resulting in a shift from the PM to the nucleus. It was found that RGLG interacts with a drought-inducible TF ethylene response factor53 (AtERF53) and mediates its degradation [60]. Meanwhile, RGLG can also interact with ubiquitin conjugating enzyme UBC13 to mediate ubiquitination of PM-located auxin carrier protein pin-formed 2 (PIN2) to control its turnover, thus affecting auxin transport. Mutations in RGLG1/2 caused an arrest of endocytosis of PIN2 and decreased auxin levels, eventually leading to the branching of root hairs [74,75]. Further studies demonstrated that RGLG could respond to drought stress by participating in the MAPK pathway, and RGLG ubiquitinated MAPKKK18 to promote its degradation, thus playing a negative regulatory role in plant drought tolerance [76].
3.2.3. E3s Participate in the ABA Signaling Pathway
SpRing is a RING-H2 ubiquitin ligase found in tomatoes and is specifically located in the ER. ABA, drought, and salt stress could induce the expression of SpRing. Under salt stress, the chlorophyll content of the SpRing-silenced plants decreased and produced more malondialdehyde and H2O2. In SpRing overexpression Arabidopsis plants, NCED3 (Nine-Cis-Epoxycarotenoid Dioxygenase 3), RD29A (Responsive to Deciccation29A), and Rab18 exhibited increased expression levels [77]. In Arabidopsis, RING ubiquitin ligase AtAIRP1 (A. thaliana ABA-insensitive RING 1) and AtAIRP2 are positive regulators in ABA-dependent responses to drought stress. AtAIRP1 is rapidly induced by drought and ABA. It positively regulates ABA-promoted stomatal closure, which may reduce transpirational water loss in response to dehydration stress [78,79]. AtAIRP2 down-regulates ATP1/SDIRIP1 (AtAIRP2 target protein 1/ SDIR1-interacting protein 1) through UPS during Arabidopsis seed germination [80]. AtAIRP3/LOG2 (loss of glutamine dumper 2) was up-regulated by high salinity, drought, and ABA treatments. Under ABA treatment, the mean stomatal diameter of leaves of both WT and the atairp3/log2 mutant decreased, but the extent of the reduction in the mutant was not as great as that in the WT. RD21 (responsive to desiccation 21) was initially isolated as a drought-induced cysteine proteinase [81]. The RD21 protein was ubiquitinated in vitro by AtAIRP3/LOG2 and degraded by 26S proteasome. Thus, AtAIRP3/LOG2 is a positive regulator of the ABA-mediated drought and salt stress tolerance [82]. AtAIRP4 acts as a positive regulator of ABA-mediated drought avoidance and a negative regulator of salt tolerance. The atairp4 mutant showed reduced sensitivity of root elongation and stomatal closure to ABA, whereas plants with overexpressed AtAIRP4 were hypersensitive to salt and osmotic stresses during seed germination. The transcriptional abundances of ABA-responsive genes in overexpression plants were higher than those in the WT and atairp4 plants [83].
PEG, ABA, and NaCl treatments significantly induced the expression of TaDIS1 (T. aestivum drought-induced SINA protein 1) in wheat. Overexpression of TaDIS1 in Arabidopsis reduced the tolerance to drought stress and increased sensitivity to ABA during seed germination [84]. TaDIS1 interacts with TaSTP (T. aestivum salt tolerant protein) in the Golgi apparatus and degrades TaSTP via the 26S proteasome pathway [85]. A cytoplasmic PnSAG1 is a PUB-ARM ubiquitin E3 ligase from the Antarctic moss Pohlia nutans. Its transcription was rapidly induced by ABA, salt, and drought stress. Overexpression of PnSAG1 in A. thaliana increased the sensitivity to salt and ABA during seed germination and decreased the expression levels of salt-/ABA-related genes [86]. In Arabidopsis, a C3H2C3 RING-type E3 ligase ATL61 plays an important role in drought tolerance. Point mutation of ATL61H109A, H122A (mATL61) abolished E3 ubiquitin ligase activity, and mATL61 overexpression lines exhibited similar ABA-related phenotypes as the WT plants. ATL61-overexpression plants exhibited ABA hypersensitivity and were more tolerant to drought, while the atl61 mutant plants were insensitive to ABA [87].
3.2.4. E3s Participate in Phosphorylation and Ubiquitination Crosstalk
Another RING-H2 E3 ubiquitin ligase, CHY zinc-finger and RING protein1 (CHYR1), was induced by drought stress and ABA treatment. It is located in the cytoplasm and nucleus [88]. CHYR1 positively regulates ABA-induced ROS production and stomatal closure [88]. In addition, studies have found that CHYR1 is involved in plant resistance to pathogen attack [81]. One mechanism is that CHYR1 mediates the turnover of WRKY70, which is known to be involved in plant osmotic stress and immune responses. In normal growth conditions, WRKY70 is mainly nonphosphorylated, so few phosphorylated WRKY70s will be degraded by the CHYR1-mediated 26S proteasome pathway. Upon pathogen attack, WRKY70 mainly becomes phosphorylated at Thr22 and Ser34 residues. The phosphorylated WRKY70 (WRKY70-P) is recognized by CHYR1 and gets ubiquitinated and degraded [89]. During plant immunity, the TF SARD1 (SAR deficient 1) and CBF60g (calmodulin-binding Factor 60-like g) mediate the synthesis of the plant defense hormone salicylic acid (SA) by regulating the transcription of isochorismate synthase 1 (ICS1) [90]. Overexpression of WRKY70 activates the expression of SA-responsive pathogenesis-related (PR) genes, whereas silencing of WRKY70 leads to up-regulated expression of jasmonic acid (JA)-responsive genes. WRKY70 can positively regulate the expression of SNC2 (suppressor of NPR1, constitutive 2) and downstream genes SARD1 and CBP60g. In turn SARD1 and CBP60g can also bind to the promoter of WRKY70, forming a circular structure that promotes the expression of each gene [91,92,93]. In general, this is a great example of phosphorylation and ubiquitination crosstalk in plant responses to drought and biotic stress.
3.3. Temperature and Cold Stress
Cold stress causes substantial loss in global agricultural productivity [94,95]. Under low temperatures, plants exhibit a variety of cold-induced physiological and biochemical responses, such as production of ROS, changes in membrane lipid composition, and changes in osmolytes [96,97,98]. Expression of C-repeat binding factor (CBF) genes is rapidly induced by cold, and their translational products directly bind to the promoters of cold-regulated genes. This activates their expression and thus protects plants from damage caused by cold stress (Figure 3) [96,99].
3.3.1. E3s Participate in the Cold Signaling Pathway
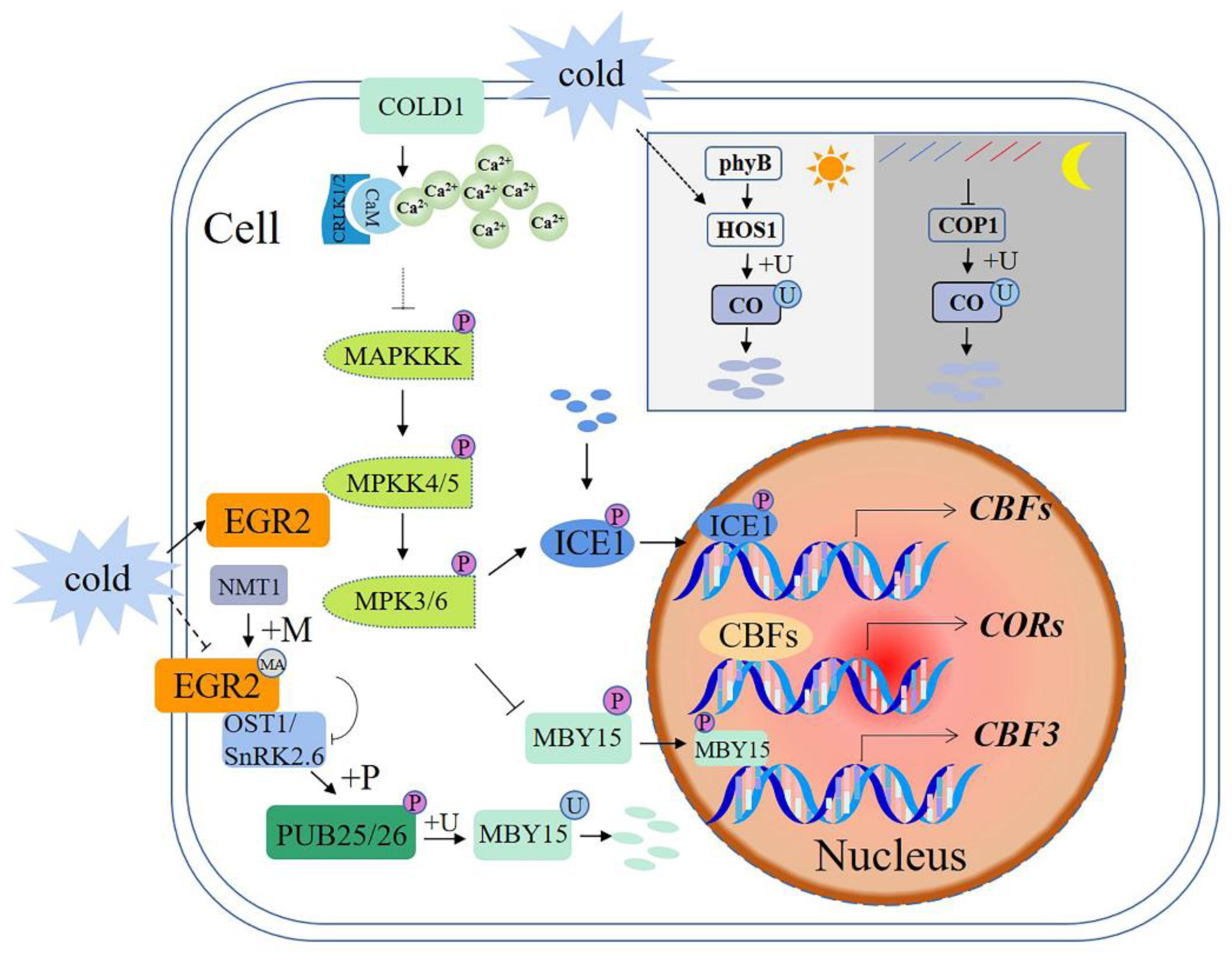
Cold stress is sensed via membrane proteins, such as a cold receptor COLD1 (chilling-tolerance divergence 1) [100], leading to a cytosolic Ca2+ spike. CPKs (calcium-dependent protein kinases) and CBLs (calcineurin B-like proteins)-CIPKs (CBL-interacting protein kinases) may mediate the Ca2+ signal to activate an MAP kinase cascade [49,101,102]. MPK3 and MPK6 mediate the phosphorylation and destabilization of a TF ICE1 [103,104], and MPK6 phosphorylates MYB15 and reduces its transcriptional activation of the CBF3 gene [105]. ICE1 (CBF expression 1) interacts with MYB15 and also directly binds to the CBF3 promoter to negatively regulate its expression, thereby modulating plant freezing tolerance [106,107].
Under cold stress, E3 ubiquitin ligases PUB25 and PUB26 can directly mediate the ubiquitination of MYB15 and lead to its degradation [108]. In addition, open stomata 1 (OST1)/SnRK2.6 is activated by cold stress and phosphorylates PUB25 and PUB26. Phosphorylation of PUB25 and PUB26 enhances MYB15 degradation in plants [105]. Therefore, PUB25 and PUB26 are located upstream of the MYB15 and downstream of OST1 to positively regulate CBF expression and plant cold responses [108,109].
3.3.2. E3s Participate in Phosphorylation and Ubiquitination Crosstalk
N-myristoylation is not only important for protein—membrane interactions but it is also important for protein—protein interactions [110,111]. Under normal conditions, a PM-localized EGR2 (clade-E growth-regulating 2) phosphatase is modified by NMT1 and interacts with OST1 to inhibit OST1 activity. At low temperatures, the interaction between EGR2 and NMT1 is weakened, resulting in the inhibition of the myristic acylation of EGR2, which partially contributes to increased OST1 activity in response to low-temperature stress [112].
3.3.3. E3s Participate in the Flowering Pathway
Light and temperature are two important environmental determinants of flowering time. Constans (CO) is a central activator of photoperiodic flowering [113]. CO protein is stable during light but is degraded rapidly in darkness. Dark-induced CO protein degradation is mediated by a ubiquitin—proteasome system, with phyB signaling through HOS1 (high expression of osmotically responsive gene 1), promoting CO degradation in the morning under long daylight conditions. At night, blue and far-red signals inhibit the activity of COP1 (constitutive photomorphogenic 1), leading to an increase in CO abundance. In darkness, the CO protein is rapidly degraded through COP1-mediated ubiquitination system. At low temperatures, cold-activated HOS1 induces CO degradation, resulting in delayed flowering. In summary, CO acts as a molecular hub that integrates light and cold stress signals into photoperiodic flowering [114,115].
3.4. Temperature and Heat Stress
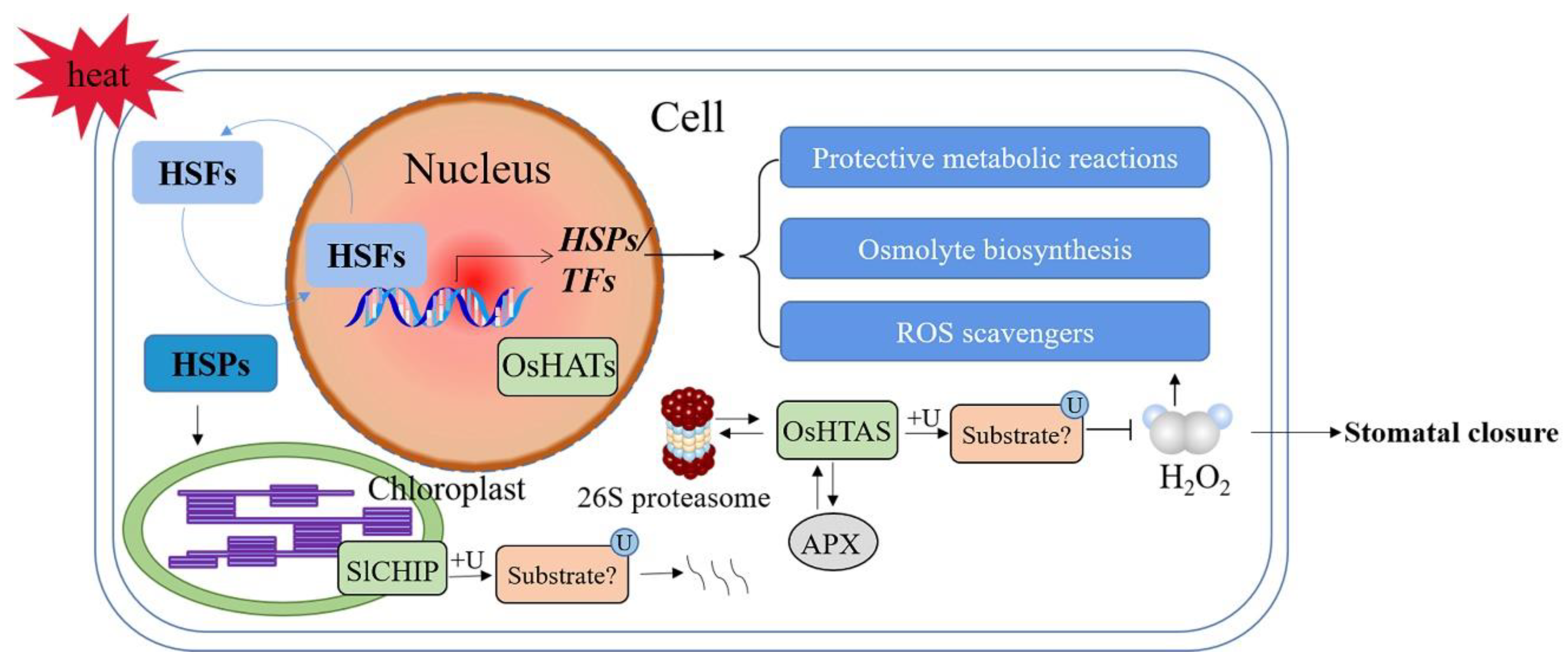
Heat stress affects plant seed germination, photosynthesis, respiration, water transpiration, and membrane stability [116]. As the terminal components of high temperature signal transduction, heat shock TFs (HSFs) trigger the transcription of heat-responsive genes encoding heat shock proteins (HSPs) and other heat-protective proteins. These include molecules such as ROS scavengers, enzymes involved in the biosynthesis of protective metabolites and osmolytes, apoptotic regulators, and other TFs [117,118,119]. Under stress conditions, HSPs interact with key heat-responsive proteins (e.g., HSFs) to form complexes, preventing them from denaturation. HSFs are then transported into the nuclei and form active trimers for transcription activation (Figure 4) [120,121]. In addition, HSPs act as molecular chaperons that target ubiquitin-mediated degradation of misfolded or damaged proteins through autophagy and/or 26S proteasome systems [122,123,124].
Under heat shock, the misfolded proteins increased rapidly, and the transcription level of U-box E3 ubiquitin ligase gene SlCHIP (Solanum lycopersicum carboxyl terminus of the HSC70-interacting proteins) in tomatoes increased at the early stage (Figure 4). Under heat stress, the photosynthetic capacity of Slchip-silenced tomato plants decreased. It is well known that photosynthesis is very sensitive to heat stress due to damage to photosynthetic organs. The response of chloroplasts to heat stress is crucial to reduce the damage and improve survival rate under high temperatures [125], suggesting that CHIP plays an important role in protecting chloroplasts from heat stress. In addition, silenced tomato plants also showed higher electrolyte permeability. These results indicated that tomato SlCHIP plays a key role in heat stress responses, mostly by targeting degradation of misfolded proteins produced during heat stress [126].
RING-H2-type E3 ubiquitin ligases (OsHTAS) localized to the nucleus and the cytoplasm are highly expressed in mesophyll cells. OsHTAS can interact with APX and 26S proteasome, thereby effectively regulating the accumulation of H2O2 and functions in the leaves to enhance heat tolerance, e.g., through modulating H2O2-induced stomatal closure [127]. E3 ubiquitin ligases tend to recognize misfolded proteins with the assistance of HSPs [128], and the presence of chaperones in OsHTAS complexes is not clear. Additionally, the complete sets of OsHTAS substrates in the nucleus and the cytoplasm, and in stomatal guard cells, need to be characterized using proteomics tools in the future.
4. Conclusions
Plants are sessile organisms. Environmental stresses cause perturbation of cellular metabolism and oxidative damage to cellular structures and molecules. Cell signaling in response to salt, drought, cold, and the stress hormone ABA largely relies on the SnRK family of protein kinases in plants. This mainly involves the activation of SnRK2 kinases to mediate several rapid responses, including gene expression regulation, stomatal closure, protein PTMs, changes in metabolism related to stress resistance, and plant growth modulation [41,129,130,131]. MAPKs have also been implicated in ABA signaling [132] and cold responses [46,90,91,92]. Of the different stresses, there are common themes, e.g., ROS and redox regulation, biosynthesis of protective metabolites/osmolytes, kinase regulation, protein phosphorylation, and ubiquitination. For example, hormone-induced protein ubiquitination plays a crucial role in determining the half-life of key regulators in plant stress responses. Plants utilize the UPS system to alter intracellular protein abundance, which is vital in responding to and resisting the environmental stressors. Activation of stress signaling may also involve ubiquitin-dependent degradation of negative regulators and/or accumulation of positive regulators [2]. Clearly, changes in the expression of the ubiquitin enzymes can alter plant responses to abiotic stressors [2]. PTM crosstalks, especially with ubiquitination are clearly demonstrated in plant responses to salt, drought, and cold [86,95,96]. Although ubiquitin ligases in plant responses to salt, drought, cold, and heat stressors may involve different pathways (e.g., ABA signaling, MAPK cascade, and ROS homeostasis), their direct substrates under the different conditions and in single cell types (e.g., guard cells) are largely unknown. For example, how SiGRF1 interacts with the aforementioned GI function is an interesting question to be addressed in the future. Future research on large-scale proteomic discovery of the E3 ligase substrates in a spatial and temporal manner, characterizing the novel targets, elucidating PTM crosstalks, and constructing molecular networks in plant stress responses will fill critical knowledge gaps and improve understanding of the molecular mechanisms underlying plant resistance to abiotic stressors. The improved knowledge will have translational potential in improving crop resilience through molecular breeding, and thereby contributing to global food security.
Author Contributions
Writing and editing, S.W. and X.L.; data search and collation, J.Z., D.C., S.C. and G.F.; conception and modification, Y.W. and C.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China Project (32172055; 32072122), Natural Science Foundation of Heilongjiang Province (YQ2020037C), China Postdoctoral Science Foundation (2020M670944), Science Foundation for Distinguished Young Scholars of Heilongjiang University, Initiation Fund for Postdoctoral Research in Heilongjiang Province, Youth Innovative Talents Training Program of Heilongjiang Regular Universities.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Haiying Li at Heilongjiang University for her suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Varshavsky, A. N-degron and C-degron pathways of protein degradation. Proc. Natl. Acad. Sci. USA 2019, 116, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, S.L. Role of the Ubiquitin Proteasome System in Plant Response to Abiotic Stress. Int. Rev. Cell Mol. Biol. 2019, 343, 65–110. [Google Scholar] [PubMed]
- Lopata, A.; Kniss, A.; Löhr, F.; Rogov, V.V.; Dötsch, V. Ubiquitination in the ERAD Process. Int. J. Mol. Sci. 2020, 21, 5369. [Google Scholar] [CrossRef] [PubMed]
- Betegon, M.; Brodsky, J.L. Unlocking the door for ERAD. Nat. Cell Biol. 2020, 22, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Coll-Martínez, B.; Crosas, B. How the 26S Proteasome Degrades Ubiquitinated Proteins in the Cell. Biomolecules 2019, 9, 395. [Google Scholar] [CrossRef] [Green Version]
- Grumati, P.; Dikic, I. Ubiquitin signaling and autophagy. J. Biol. Chem. 2018, 293, 5404–5413. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Luo, Z.Q. Post-translational regulation of ubiquitin signaling. J. Cell Biol. 2019, 218, 1776–1786. [Google Scholar] [CrossRef]
- Brunner, A.M.; Yakovlev, I.A.; Strauss, S.H. Validating internal controls for quantitative plant gene expression studies. BMC Plant Biol. 2004, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.M.; Kelleher, N.L. Proteoforms as the next proteomics currency. Science 2018, 359, 1106–1107. [Google Scholar] [CrossRef]
- Smith, L.M.; Agar, J.N.; Chamot-Rooke, J.; Danis, P.O.; Ge, Y.; Loo, J.A.; Paša-Tolić, L.; Tsybin, Y.O.; Kelleher, N.L. The Human Proteoform Project: Defining the human proteome. Sci. Adv. 2021, 7, eabk0734. [Google Scholar] [CrossRef]
- Zhang, Y.; Fonslow, B.R.; Shan, B.; Baek, M.C.; Yates, J.R., 3rd. Protein analysis by shotgun/bottom-up proteomics. Chem. Rev. 2013, 113, 2343–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjerpe, R.; Aillet, F.; Lopitz-Otsoa, F.; Lang, V.; England, P.; Rodriguez, M.S. Efficient protection and isolation of ubiquitylated proteins using tandem ubiquitin-binding entities. EMBO Rep. 2009, 10, 1250–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Paige, J.S.; Jaffrey, S.R. Global analysis of lysine ubiquitination by ubiquitin remnant immunoaffinity profiling. Nat. Biotechnol. 2010, 28, 868–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.; Schwartz, D.; Elias, J.E.; Thoreen, C.C.; Cheng, D.; Marsischky, G.; Roelofs, J.; Finley, D.; Gygi, S.P. A proteomics approach to understanding protein ubiquitination. Nat. Biotechnol. 2003, 21, 921–926. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, S.E.; Riley, B.E.; Shaler, T.A.; Trevino, R.S.; Becker, C.H.; Schulman, H.; Kopito, R.R. Protein standard absolute quantification (PSAQ) method for the measurement of cellular ubiquitin pools. Nat. Methods 2011, 8, 691–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtake, F.; Tsuchiya, H.; Tanaka, K.; Saeki, Y. Methods to measure ubiquitin chain length and linkage. Methods Enzymol. 2019, 618, 105–133. [Google Scholar]
- Ryu, K.Y.; Baker, R.T.; Kopito, R.R. Ubiquitin-specific protease 2 as a tool for quantification of total ubiquitin levels in biological specimens. Anal. Biochem. 2006, 353, 153–155. [Google Scholar] [CrossRef]
- Stone, S.L. The role of ubiquitin and the 26S proteasome in plant abiotic stress signaling. Front. Plant Sci. 2014, 5, 135. [Google Scholar] [CrossRef]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef]
- Swatek, K.N.; Usher, J.L.; Kueck, A.F.; Gladkova, C.; Mevissen, T.E.T.; Pruneda, J.N.; Skern, T.; Komander, D. Insights into ubiquitin chain architecture using Ub-clipping. Nature 2019, 572, 533–537. [Google Scholar] [CrossRef]
- Choudhary, C.; Kumar, C.; Gnad, F.; Nielsen, M.L.; Rehman, M.; Walther, T.C.; Olsen, J.V.; Mann, M. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 2009, 325, 834–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swatek, K.N.; Komander, D. Ubiquitin modifications. Cell Res. 2016, 26, 399–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, L.D.; Gevaert, K.; De Smet, I. Protein Language: Post-Translational Modifications Talking to Each Other. Trends Plant Sci. 2018, 23, 1068–1080. [Google Scholar] [CrossRef] [PubMed]
- Mevissen, T.E.T.; Komander, D. Mechanisms of Deubiquitinase Specificity and Regulation. Annu. Rev. Biochem. 2017, 86, 159–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Serio, R.J.; Schofield, A.; Liu, H.; Rasmussen, S.R.; Hofius, D.; Stone, S.L. Arabidopsis RING-type E3 ubiquitin ligase XBAT35.2 promotes proteasome-dependent degradation of ACD11 to attenuate abiotic stress tolerance. Plant J. 2020, 104, 1712–1723. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Shabek, N. Ubiquitin Ligases: Structure, Function, and Regulation. Annu. Rev. Biochem. 2017, 86, 129–157. [Google Scholar] [CrossRef]
- Clague, M.J.; Urbé, S. Ubiquitin: Same molecule, different degradation pathways. Cell 2010, 143, 682–685. [Google Scholar] [CrossRef] [Green Version]
- Michel, M.A.; Komander, D.; Elliott, P.R. Enzymatic Assembly of Ubiquitin Chains. Methods Mol. Biol. 2018, 1844, 73–84. [Google Scholar]
- Smit, J.J.; Sixma, T.K. RBR E3-ligases at work. EMBO Rep. 2014, 15, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Deshaies, R.J.; Joazeiro, C.A. RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 2009, 78, 399–434. [Google Scholar] [CrossRef]
- Enchev, R.I.; Schulman, B.A.; Peter, M. Protein neddylation: Beyond cullin-RING ligases. Nat. Rev. Mol. Cell Biol. 2015, 16, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Yee, D.; Goring, D.R. The diversity of plant U-box E3 ubiquitin ligases: From upstream activators to downstream target substrates. J. Exp. Bot. 2009, 60, 1109–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, S.; Nakayama, K.I. U-box proteins as a new family of ubiquitin ligases. Biochem. Biophys. Res. Commun. 2003, 302, 635–645. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Yada, M.; Matsumoto, M.; Ishida, N.; Nakayama, K.I. U box proteins as a new family of ubiquitin-protein ligases. J. Biol. Chem. 2001, 276, 33111–33120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koegl, M.; Hoppe, T.; Schlenker, S.; Ulrich, H.D.; Mayer, T.U.; Jentsch, S. A novel ubiquitination factor, E4, is involved in multiubiquitin chain assembly. Cell 1999, 96, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.R.; Park, C.H.; Venu, R.C.; Gough, J.; Wang, G.L. Classification, expression pattern, and E3 ligase activity assay of rice U-box-containing proteins. Mol. Plant 2008, 1, 800–815. [Google Scholar] [CrossRef] [Green Version]
- Spratt, D.E.; Walden, H.; Shaw, G.S. RBR E3 ubiquitin ligases: New structures, new insights, new questions. Biochem. J. 2014, 458, 421–437. [Google Scholar] [CrossRef] [Green Version]
- Lindon, C.; Pines, J. Ordered proteolysis in anaphase inactivates Plk1 to contribute to proper mitotic exit in human cells. J. Cell Biol. 2004, 164, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Rotin, D.; Kumar, S. Physiological functions of the HECT family of ubiquitin ligases. Nat. Rev. Mol. Cell Biol. 2009, 10, 398–409. [Google Scholar] [CrossRef]
- Kavsak, P.; Rasmussen, R.K.; Causing, C.G.; Bonni, S.; Zhu, H.; Thomsen, G.H.; Wrana, J.L. Smad7 binds to Smurf2 to form an E3 ubiquitin ligase that targets the TGF beta receptor for degradation. Mol. Cell 2000, 6, 1365–1375. [Google Scholar] [CrossRef]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bossoreille de Ribou, S.; Douam, F.; Hamant, O.; Frohlich, M.W.; Negrutiu, I. Plant science and agricultural productivity: Why are we hitting the yield ceiling? Plant Sci. 2013, 210, 159–176. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants: Growth, antioxidant activity and phenolic compounds. Plant Physiol. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef]
- Roper, S.D.; Chaudhari, N. Taste buds: Cells, signals and synapses. Nat. Rev. Neurosci. 2017, 18, 485–497. [Google Scholar] [CrossRef]
- Luo, X.; Wang, B.; Gao, S.; Zhang, F.; Terzaghi, W.; Dai, M. Genome-wide association study dissects the genetic bases of salt tolerance in maize seedlings. J. Integr. Plant Biol. 2019, 61, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.; Pardo, J.M.; Batelli, G.; Van Oosten, M.J.; Bressan, R.A.; Li, X. The Salt Overly Sensitive (SOS) pathway: Established and emerging roles. Mol. Plant 2013, 6, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Quintero, F.J.; Ohta, M.; Shi, H.; Zhu, J.K.; Pardo, J.M. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 9061–9066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishitani, M.; Liu, J.; Halfter, U.; Kim, C.S.; Shi, W.; Zhu, J.K. SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding. Plant Cell 2000, 12, 1667–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Ishitani, M.; Halfter, U.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000, 97, 3730–3734. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Ali, Z.; Park, H.J.; Park, S.J.; Cha, J.Y.; Perez-Hormaeche, J.; Quintero, F.J.; Shin, G.; Kim, M.R.; Qiang, Z.; et al. Release of SOS2 kinase from sequestration with GIGANTEA determines salt tolerance in Arabidopsis. Nat. Commun. 2013, 4, 1352. [Google Scholar] [CrossRef] [Green Version]
- De Zelicourt, A.; Colcombet, J.; Hirt, H. The Role of MAPK Modules and ABA during Abiotic Stress Signaling. Trends Plant Sci. 2016, 21, 677–685. [Google Scholar] [CrossRef]
- Colcombet, J.; Hirt, H. Arabidopsis MAPKs: A complex signalling network involved in multiple biological processes. Biochem. J. 2008, 413, 217–226. [Google Scholar] [CrossRef]
- Ichimura, K.; Mizoguchi, T.; Yoshida, R.; Yuasa, T.; Shinozaki, K. Various abiotic stresses rapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6. Plant J. 2000, 24, 655–665. [Google Scholar] [CrossRef]
- Liu, J.; Lin, Q.F.; Qi, S.L.; Feng, X.J.; Han, H.L.; Xu, T.; Hua, X.J. The F-box protein EST1 modulates salt tolerance in Arabidopsis by regulating plasma membrane Na(+)/H(+) antiport activity. J. Plant Physiol. 2020, 251, 153217. [Google Scholar] [CrossRef]
- Du, B.; Nie, N.; Sun, S.; Hu, Y.; Bai, Y.; He, S.; Zhao, N.; Liu, Q.; Zhai, H. A novel sweetpotato RING-H2 type E3 ubiquitin ligase gene IbATL38 enhances salt tolerance in transgenic Arabidopsis. Plant Sci. 2021, 304, 110802. [Google Scholar] [CrossRef]
- Cheng, M.C.; Hsieh, E.J.; Chen, J.H.; Chen, H.Y.; Lin, T.P. Arabidopsis RGLG2, functioning as a RING E3 ligase, interacts with AtERF53 and negatively regulates the plant drought stress response. Plant Physiol. 2012, 158, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Belda-Palazon, B.; Julian, J.; Coego, A.; Wu, Q.; Zhang, X.; Batistic, O.; Alquraishi, S.A.; Kudla, J.; An, C.; Rodriguez, P.L. ABA inhibits myristoylation and induces shuttling of the RGLG1 E3 ligase to promote nuclear degradation of PP2CA. Plant J. 2019, 98, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.R.; Salinas, J.; Collinge, D.B. 14-3-3 proteins and the response to abiotic and biotic stress. Plant Mol. Biol. 2002, 50, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jiang, C.; Kang, L.; Zhang, H.; Song, Y.; Zou, Z.; Zheng, W. Over-Expression of a 14-3-3 Protein From Foxtail Millet Improves Plant Tolerance to Salinity Stress in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. The Response of Salinity Stress-Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef] [PubMed]
- Clavel, M.; Pélissier, T.; Descombin, J.; Jean, V.; Picart, C.; Charbonel, C.; Saez-Vásquez, J.; Bousquet-Antonelli, C.; Deragon, J.M. Parallel action of AtDRB2 and RdDM in the control of transposable element expression. BMC Plant Biol. 2015, 15, 70. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Cai, X.T.; Du, J.; Zhao, T.L.; Wang, P.F.; Zhao, P.X.; Liu, R.; Xie, Q.; Cao, X.F.; Xiang, C.B. PARAQUAT TOLERANCE3 Is an E3 Ligase That Switches off Activated Oxidative Response by Targeting Histone-Modifying PROTEIN METHYLTRANSFERASE4b. PLoS Genet. 2016, 12, e1006332. [Google Scholar] [CrossRef] [Green Version]
- Alfatih, A.; Wu, J.; Jan, S.U.; Zhang, Z.S.; Xia, J.Q.; Xiang, C.B. Loss of rice PARAQUAT TOLERANCE 3 confers enhanced resistance to abiotic stresses and increases grain yield in field. Plant Cell Environ. 2020, 43, 2743–2754. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kawanabe, M.; Hayashi, Y.; Endo, T.; Nishikawa, S. A vacuolar carboxypeptidase mutant of Arabidopsis thaliana is degraded by the ERAD pathway independently of its N-glycan. Biochem. Biophys. Res. Commun. 2010, 393, 384–389. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, H.; Duan, M.; Zhu, F.; Wen, J.; Dong, J.; Wang, T. Medicago falcata MfSTMIR, an E3 ligase of endoplasmic reticulum-associated degradation, is involved in salt stress response. Plant J. 2019, 98, 680–696. [Google Scholar] [CrossRef] [Green Version]
- Mizoi, J.; Kanazawa, N.; Kidokoro, S.; Takahashi, F.; Qin, F.; Morimoto, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Heat-induced inhibition of phosphorylation of the stress-protective transcription factor DREB2A promotes thermotolerance of Arabidopsis thaliana. J. Biol. Chem. 2019, 294, 902–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Yin, Y.; Liu, X.; Tong, S.; Xing, J.; Zhang, Y.; Pudake, R.N.; Izquierdo, E.M.; Peng, H.; Xin, M.; et al. The E3 Ligase TaSAP5 Alters Drought Stress Responses by Promoting the Degradation of DRIP Proteins. Plant Physiol. 2017, 175, 1878–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Xu, J.; Liu, X.; Liang, W.; Xin, M.; Du, J.; Hu, Z.; Peng, H.; Guo, W.; Ni, Z.; et al. Identification of HSP90C as a substrate of E3 ligase TaSAP5 through ubiquitylome profiling. Plant Sci. 2019, 287, 110170. [Google Scholar] [CrossRef] [PubMed]
- Pan, I.C.; Schmidt, W. Functional implications of K63-linked ubiquitination in the iron deficiency response of Arabidopsis roots. Front. Plant Sci. 2014, 4, 542. [Google Scholar] [CrossRef] [Green Version]
- Leitner, J.; Retzer, K.; Korbei, B.; Luschnig, C. Dynamics in PIN2 auxin carrier ubiquitylation in gravity-responding Arabidopsis roots. Plant Signal. Behav. 2012, 7, 1271–1273. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Kang, L.; Li, Y.; Wu, C.; Zheng, C.; Liu, P.; Huang, J. RING finger protein RGLG1 and RGLG2 negatively modulate MAPKKK18 mediated drought stress tolerance in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 484–493. [Google Scholar] [CrossRef]
- Qi, S.; Lin, Q.; Zhu, H.; Gao, F.; Zhang, W.; Hua, X. The RING Finger E3 Ligase SpRing is a Positive Regulator of Salt Stress Signaling in Salt-Tolerant Wild Tomato Species. Plant Cell Physiol. 2016, 57, 528–539. [Google Scholar] [CrossRef] [Green Version]
- Ryu, M.Y.; Cho, S.K.; Kim, W.T. The Arabidopsis C3H2C3-type RING E3 ubiquitin ligase AtAIRP1 is a positive regulator of an abscisic acid-dependent response to drought stress. Plant Physiol. 2010, 154, 1983–1997. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.K.; Ryu, M.Y.; Seo, D.H.; Kang, B.G.; Kim, W.T. The Arabidopsis RING E3 ubiquitin ligase AtAIRP2 plays combinatory roles with AtAIRP1 in abscisic acid-mediated drought stress responses. Plant Physiol. 2011, 157, 2240–2257. [Google Scholar] [CrossRef] [Green Version]
- Oh, T.R.; Kim, J.H.; Cho, S.K.; Ryu, M.Y.; Yang, S.W.; Kim, W.T. AtAIRP2 E3 Ligase Affects ABA and High-Salinity Responses by Stimulating Its ATP1/SDIRIP1 Substrate Turnover. Plant Physiol. 2017, 174, 2515–2531. [Google Scholar] [CrossRef] [Green Version]
- Koizumi, M.; Yamaguchi-Shinozaki, K.; Tsuji, H.; Shinozaki, K. Structure and expression of two genes that encode distinct drought-inducible cysteine proteinases in Arabidopsis thaliana. Gene 1993, 129, 175–182. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, W.T. The Arabidopsis RING E3 ubiquitin ligase AtAIRP3/LOG2 participates in positive regulation of high-salt and drought stress responses. Plant Physiol. 2013, 162, 1733–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Liu, Q.; Liu, Z.; Yang, H.; Wang, J.; Li, X.; Yang, Y. Arabidopsis C3HC4-RING finger E3 ubiquitin ligase AtAIRP4 positively regulates stress-responsive abscisic acid signaling. J. Integr. Plant Biol. 2016, 58, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, L.; Zhang, L.; Lv, Q.; Zhao, Y.; Li, X. Isolation and identification of wheat gene TaDIS1 encoding a RING finger domain protein, which negatively regulates drought stress tolerance in transgenic Arabidopsis. Plant Sci. 2018, 275, 49–59. [Google Scholar] [CrossRef]
- Lv, Q.; Zhang, L.; Zan, T.; Li, L.; Li, X. Wheat RING E3 ubiquitin ligase TaDIS1 degrade TaSTP via the 26S proteasome pathway. Plant Sci. 2020, 296, 110494. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, S.; Liu, H.; Chen, K.; Zhang, P. PnSAG1, an E3 ubiquitin ligase of the Antarctic moss Pohlia nutans, enhanced sensitivity to salt stress and ABA. Plant Physiol. Biochem. 2019, 141, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Wang, T.; Shi, W.; Li, S.; Liu, Z.; Wang, J.; Yang, Y. E3 ubiquitin ligase ATL61 acts as a positive regulator in abscisic acid mediated drought response in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 528, 292–298. [Google Scholar] [CrossRef]
- Ding, S.; Zhang, B.; Qin, F. Arabidopsis RZFP34/CHYR1, a Ubiquitin E3 Ligase, Regulates Stomatal Movement and Drought Tolerance via SnRK2.6-Mediated Phosphorylation. Plant Cell 2015, 27, 3228–3244. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, B.; Lou, S.; Bi, H.; Tang, H.; Tong, S.; Song, Y.; Chen, N.; Zhang, H.; Jiang, Y.; et al. CHYR1 ubiquitinates the phosphorylated WRKY70 for degradation to balance immunity in Arabidopsis thaliana. New Phytol. 2021, 230, 1095–1109. [Google Scholar] [CrossRef]
- Sun, T.; Liang, W.; Zhang, Y.; Li, X. Negative regulation of resistance protein-mediated immunity by master transcription factors SARD1 and CBP60g. J. Integr. Plant Biol. 2018, 60, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, Y.; Fang, B.; Gannon, P.; Ding, P.; Li, X.; Zhang, Y. Arabidopsis snc2-1D activates receptor-like protein-mediated immunity transduced through WRKY70. Plant Cell 2010, 22, 3153–3163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Ding, Y.; Tian, H.; Wang, S.; Zhang, Y. WRKY54 and WRKY70 positively regulate SARD1 and CBP60g expression in plant immunity. Plant Signal. Behav. 2021, 16, 1932142. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, Y.; Ding, P.; Johnson, K.; Li, X.; Zhang, Y. The ankyrin-repeat transmembrane protein BDA1 functions downstream of the receptor-like protein SNC2 to regulate plant immunity. Plant Physiol. 2012, 159, 1857–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, M.R.; Knight, H. Low-temperature perception leading to gene expression and cold tolerance in higher plants. New Phytol. 2012, 195, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Shi, Y.; Yang, S. Molecular Regulation of Plant Responses to Environmental Temperatures. Mol. Plant 2020, 13, 544–564. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. PLANT COLD ACCLIMATION: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar]
- Browse, J.; Xin, Z. Temperature sensing and cold acclimation. Curr. Opin. Plant Biol. 2001, 4, 241–246. [Google Scholar] [CrossRef]
- Stockinger, E.J.; Gilmour, S.J.; Thomashow, M.F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 1997, 94, 1035–1040. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Yang, S. COLD1: A cold sensor in rice. Sci. China Life Sci. 2015, 58, 409–410. [Google Scholar] [CrossRef] [Green Version]
- Atif, R.M.; Shahid, L.; Waqas, M.; Ali, B.; Rashid, M.A.R.; Azeem, F.; Nawaz, M.A.; Wani, S.H.; Chung, G. Insights on Calcium-Dependent Protein Kinases (CPKs) Signaling for Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 5298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyal, S.K.; Mahiwal, S.; Nambiar, D.M.; Pandey, G.K. CBL-CIPK module-mediated phosphoregulation: Facts and hypothesis. Biochem. J. 2020, 477, 853–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Ding, Y.; Shi, Y.; Zhang, X.; Zhang, S.; Gong, Z.; Yang, S. MPK3- and MPK6-Mediated ICE1 Phosphorylation Negatively Regulates ICE1 Stability and Freezing Tolerance in Arabidopsis. Dev. Cell 2017, 43, 630–642.e4. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, P.; Si, T.; Hsu, C.C.; Wang, L.; Zayed, O.; Yu, Z.; Zhu, Y.; Dong, J.; Tao, W.A.; et al. MAP Kinase Cascades Regulate the Cold Response by Modulating ICE1 Protein Stability. Dev. Cell 2017, 43, 618–629.e615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kim, H.S.; Bahk, S.; An, J.; Yoo, Y.; Kim, J.Y.; Chung, W.S. Phosphorylation of the transcriptional repressor MYB15 by mitogen-activated protein kinase 6 is required for freezing tolerance in Arabidopsis. Nucleic Acids Res. 2017, 45, 6613–6627. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, M.; Hao, Y.; Kapoor, A.; Dong, C.H.; Fujii, H.; Zheng, X.; Zhu, J.K. A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance. J. Biol. Chem. 2006, 281, 37636–37645. [Google Scholar] [CrossRef] [Green Version]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.H.; Hong, X.; Agarwal, M.; Zhu, J.K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ding, Y.; Li, Z.; Shi, Y.; Wang, J.; Hua, J.; Gong, Z.; Zhou, J.M.; Yang, S. PUB25 and PUB26 Promote Plant Freezing Tolerance by Degrading the Cold Signaling Negative Regulator MYB15. Dev. Cell 2019, 51, 222–235.e225. [Google Scholar] [CrossRef]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, S.; Aderem, A. The myristoyl-electrostatic switch: A modulator of reversible protein-membrane interactions. Trends Biochem. Sci. 1995, 20, 272–276. [Google Scholar] [CrossRef]
- Wright, M.H.; Heal, W.P.; Mann, D.J.; Tate, E.W. Protein myristoylation in health and disease. J. Chem. Biol. 2010, 3, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Lv, J.; Shi, Y.; Gao, J.; Hua, J.; Song, C.; Gong, Z.; Yang, S. EGR2 phosphatase regulates OST1 kinase activity and freezing tolerance in Arabidopsis. EMBO J. 2019, 38, e99819. [Google Scholar] [CrossRef]
- Yang, S.; Weers, B.D.; Morishige, D.T.; Mullet, J.E. CONSTANS is a photoperiod regulated activator of flowering in sorghum. BMC Plant Biol. 2014, 14, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.H.; Seo, P.J.; Park, C.M. The E3 ubiquitin ligase HOS1 regulates Arabidopsis flowering by mediating CONSTANS degradation under cold stress. J. Biol. Chem. 2012, 287, 43277–43287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazaro, A.; Valverde, F.; Piñeiro, M.; Jarillo, J.A. The Arabidopsis E3 ubiquitin ligase HOS1 negatively regulates CONSTANS abundance in the photoperiodic control of flowering. Plant Cell 2012, 24, 982–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshad, M.S.; Farooq, M.; Asch, F.; Krishna, J.S.V.; Prasad, P.V.V.; Siddique, K.H.M. Thermal stress impacts reproductive development and grain yield in rice. Plant Physiol. Biochem. 2017, 115, 57–72. [Google Scholar] [CrossRef]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef]
- Busch, W.; Wunderlich, M.; Schöffl, F. Identification of novel heat shock factor-dependent genes and biochemical pathways in Arabidopsis thaliana. Plant J. 2005, 41, 1–14. [Google Scholar] [CrossRef]
- Fortunati, A.; Piconese, S.; Tassone, P.; Ferrari, S.; Migliaccio, F. A new mutant of Arabidopsis disturbed in its roots, right-handed slanting, and gravitropism defines a gene that encodes a heat-shock factor. J. Exp. Bot. 2008, 59, 1363–1374. [Google Scholar] [CrossRef] [Green Version]
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat shock factors: Integrators of cell stress, development and lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef]
- Gomez-Pastor, R.; Burchfiel, E.T.; Thiele, D.J. Regulation of heat shock transcription factors and their roles in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Amm, I.; Sommer, T.; Wolf, D.H. Protein quality control and elimination of protein waste: The role of the ubiquitin-proteasome system. Biochim. Biophys. Acta 2014, 1843, 182–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, S.; Cuervo, A.M. The coming of age of chaperone-mediated autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Von Koskull-Döring, P.; Scharf, K.D.; Nover, L. The diversity of plant heat stress transcription factors. Trends Plant Sci. 2007, 12, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Feng, L.; Li, J.; He, Z. Genetic and epigenetic control of plant heat responses. Front. Plant Sci. 2015, 6, 267. [Google Scholar] [CrossRef]
- Zhang, Y.; Lai, X.; Yang, S.; Ren, H.; Yuan, J.; Jin, H.; Shi, C.; Lai, Z.; Xia, G. Functional analysis of tomato CHIP ubiquitin E3 ligase in heat tolerance. Sci. Rep. 2021, 11, 1713. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, C.; Wei, C.; Liu, X.; Wang, M.; Yu, F.; Xie, Q.; Tu, J. The RING Finger Ubiquitin E3 Ligase OsHTAS Enhances Heat Tolerance by Promoting H2O2-Induced Stomatal Closure in Rice. Plant Physiol. 2016, 170, 429–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xia, G.; Zhu, Q. Conserved and Unique Roles of Chaperone-Dependent E3 Ubiquitin Ligase CHIP in Plants. Front. Plant Sci. 2021, 12, 699756. [Google Scholar] [CrossRef]
- Jung, C.; Nguyen, N.H.; Cheong, J.J. Transcriptional Regulation of Protein Phosphatase 2C Genes to Modulate Abscisic Acid Signaling. Int. J. Mol. Sci. 2020, 21, 9517. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, N. Abscisic Acid and abiotic stress signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danquah, A.; de Zelicourt, A.; Colcombet, J.; Hirt, H. The role of ABA and MAPK signaling pathways in plant abiotic stress responses. Biotechnol. Adv. 2014, 32, 40–52. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram of E3 ubiquitin ligases in different signaling pathways involved in plant responses to salt stress. The salt-stress pathways crosstalk with the ABA and MAPK pathways. E3 functions by ubiquitination of downstream target proteins. +P—phosphorylation; +U—ubiquitination; +MA—N-myristoylation; +M—methylation; ABA—abscisic acid; SOS—salt overly sensitive; GI—GIGANTEA; IbATL38—Ipomoea batatas Arabidopsis Toxicos en Levadura 38; SiGRF1—Setaria italica growtn-regulating factor1; EST1—Ever shorter Telomeres 1; SiRNF1/2—Setaria italica RING finger protein 1/2; RGLG—RING domain ligase; FT—Flowering Locus T; LFY—LEAFY; PRMT4b—protein arginine methyltransferase 4b; AtAPX1—Arabidopsis thaliana ascorbate peroxidase 1; GPX1—glutathione peroxidase 1; PQT3—paraquat tolerance 3; MfSTMIR—Medicago falcata salt tunicamycin-induced RING finger protein; MtUBC32—ubiquitin-conjugating enzyme 32; ROS—reactive oxygen species; MAPK—mitogen-activated protein kinase. A solid arrow shows a promoting effect or positive regulation; a dotted arrow shows that the specific mechanism of action is unclear; a horizontal line shows inhibition or negative regulation; a double arrow shows interacting proteins.
Figure 1.
Schematic diagram of E3 ubiquitin ligases in different signaling pathways involved in plant responses to salt stress. The salt-stress pathways crosstalk with the ABA and MAPK pathways. E3 functions by ubiquitination of downstream target proteins. +P—phosphorylation; +U—ubiquitination; +MA—N-myristoylation; +M—methylation; ABA—abscisic acid; SOS—salt overly sensitive; GI—GIGANTEA; IbATL38—Ipomoea batatas Arabidopsis Toxicos en Levadura 38; SiGRF1—Setaria italica growtn-regulating factor1; EST1—Ever shorter Telomeres 1; SiRNF1/2—Setaria italica RING finger protein 1/2; RGLG—RING domain ligase; FT—Flowering Locus T; LFY—LEAFY; PRMT4b—protein arginine methyltransferase 4b; AtAPX1—Arabidopsis thaliana ascorbate peroxidase 1; GPX1—glutathione peroxidase 1; PQT3—paraquat tolerance 3; MfSTMIR—Medicago falcata salt tunicamycin-induced RING finger protein; MtUBC32—ubiquitin-conjugating enzyme 32; ROS—reactive oxygen species; MAPK—mitogen-activated protein kinase. A solid arrow shows a promoting effect or positive regulation; a dotted arrow shows that the specific mechanism of action is unclear; a horizontal line shows inhibition or negative regulation; a double arrow shows interacting proteins.

Figure 2.
Schematic diagram of E3 ubiquitin ligases in signaling pathways involved in plant responses to drought stress. E3 functions by ubiquitination of downstream target proteins. +P—phosphorylation; +U—ubiquitination; +MA—N-myristoylation; +M—methylation; ABA—abscisic acid; DREB2A—dehydration-responsive element-binding protein 2A; DRIP1—DREB2A interacting protein 1; TaSAP5—Triticum aestivum stress-associated protein; AtAIRP1—A. thaliana ABA-insensitive RING 1; LOG2—loss of glutamine dumper 2; HSP90C—chloroplast heat shock protein 90; RGLG—RING domain ligase; PIN2—pin-formed 2; UBC13—ubiquitin-conjugating enzyme UBC13; RD21—responsive to desiccation 21; TaDIS1—Triticum aestivum drought-induced SINA protein 1; TaSTP—Triticum aestivum salt tolerant protein; CHYR1—CHY zinc-finger and RING protein1; SARD1—SAR deficient 1; CBF60g—calmodulin-binding protein 60-like g; SA—salicylic acid; ICS1—isochorismate synthase 1; PR genes—pathogenesis-related genes; JA—jasmonic acid; SNC2—suppressor of NPR1, constitutive 2; MAPK—mitogen-activated protein kinase. A solid arrow shows a promoting effect or positive regulation; a dotted arrow shows that the specific mechanism of action is unclear; a horizontal line shows inhibition or negative regulation; a double arrow shows interacting proteins.
Figure 2.
Schematic diagram of E3 ubiquitin ligases in signaling pathways involved in plant responses to drought stress. E3 functions by ubiquitination of downstream target proteins. +P—phosphorylation; +U—ubiquitination; +MA—N-myristoylation; +M—methylation; ABA—abscisic acid; DREB2A—dehydration-responsive element-binding protein 2A; DRIP1—DREB2A interacting protein 1; TaSAP5—Triticum aestivum stress-associated protein; AtAIRP1—A. thaliana ABA-insensitive RING 1; LOG2—loss of glutamine dumper 2; HSP90C—chloroplast heat shock protein 90; RGLG—RING domain ligase; PIN2—pin-formed 2; UBC13—ubiquitin-conjugating enzyme UBC13; RD21—responsive to desiccation 21; TaDIS1—Triticum aestivum drought-induced SINA protein 1; TaSTP—Triticum aestivum salt tolerant protein; CHYR1—CHY zinc-finger and RING protein1; SARD1—SAR deficient 1; CBF60g—calmodulin-binding protein 60-like g; SA—salicylic acid; ICS1—isochorismate synthase 1; PR genes—pathogenesis-related genes; JA—jasmonic acid; SNC2—suppressor of NPR1, constitutive 2; MAPK—mitogen-activated protein kinase. A solid arrow shows a promoting effect or positive regulation; a dotted arrow shows that the specific mechanism of action is unclear; a horizontal line shows inhibition or negative regulation; a double arrow shows interacting proteins.

Figure 3.
Schematic diagram of E3 ubiquitin ligases in signaling pathways involved in plant responses to cold stress. +P—phosphorylation; +U—ubiquitination; +MA—N-myristoylation; CORs—cold-regulated genes; CBF—C-repeat binding factor; COLD1—chilling-tolerance divergence 1; CBLs—calcineurin B-like proteins; CIPKs—CBL-interacting protein kinases; ICE1—CBF expression 1; OST1—open stomata1; EGR2:—lade-E growth-regulating 2; NMT1—myristoyltransferase; CO—constans; COP1—constitutive photomorphogenic 1. A solid arrow shows a promoting effect or positive regulation; a dotted arrow shows that the specific mechanism of action is unclear; a horizontal line shows inhibition or negative regulation.
Figure 3.
Schematic diagram of E3 ubiquitin ligases in signaling pathways involved in plant responses to cold stress. +P—phosphorylation; +U—ubiquitination; +MA—N-myristoylation; CORs—cold-regulated genes; CBF—C-repeat binding factor; COLD1—chilling-tolerance divergence 1; CBLs—calcineurin B-like proteins; CIPKs—CBL-interacting protein kinases; ICE1—CBF expression 1; OST1—open stomata1; EGR2:—lade-E growth-regulating 2; NMT1—myristoyltransferase; CO—constans; COP1—constitutive photomorphogenic 1. A solid arrow shows a promoting effect or positive regulation; a dotted arrow shows that the specific mechanism of action is unclear; a horizontal line shows inhibition or negative regulation.

Figure 4.
Schematic diagram of E3 ubiquitin ligases in signaling pathways involved in plant responses to heat stress. +U—ubiquitination; ROS—reactive oxygen species; HSFs—heat shock TFs; HSPs—heat shock proteins; SlCHIP—Solanum lycopersicum carboxyl terminus of the HSC70-interacting proteins; OsHTAS—Oryza sativa heat tolerance at seedling stage; H2O2—hydrogen peroxide. A solid arrow shows a promoting effect or positive regulation; a horizontal line shows inhibition or negative regulation; a double arrow shows interacting proteins.
Figure 4.
Schematic diagram of E3 ubiquitin ligases in signaling pathways involved in plant responses to heat stress. +U—ubiquitination; ROS—reactive oxygen species; HSFs—heat shock TFs; HSPs—heat shock proteins; SlCHIP—Solanum lycopersicum carboxyl terminus of the HSC70-interacting proteins; OsHTAS—Oryza sativa heat tolerance at seedling stage; H2O2—hydrogen peroxide. A solid arrow shows a promoting effect or positive regulation; a horizontal line shows inhibition or negative regulation; a double arrow shows interacting proteins.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, S.; Lv, X.; Zhang, J.; Chen, D.; Chen, S.; Fan, G.; Ma, C.; Wang, Y. Roles of E3 Ubiquitin Ligases in Plant Responses to Abiotic Stresses. Int. J. Mol. Sci. 2022, 23, 2308. https://doi.org/10.3390/ijms23042308
AMA Style
Wang S, Lv X, Zhang J, Chen D, Chen S, Fan G, Ma C, Wang Y. Roles of E3 Ubiquitin Ligases in Plant Responses to Abiotic Stresses. International Journal of Molecular Sciences. 2022; 23(4):2308. https://doi.org/10.3390/ijms23042308
Chicago/Turabian StyleWang, Shuang, Xiaoyan Lv, Jialin Zhang, Daniel Chen, Sixue Chen, Guoquan Fan, Chunquan Ma, and Yuguang Wang. 2022. "Roles of E3 Ubiquitin Ligases in Plant Responses to Abiotic Stresses" International Journal of Molecular Sciences 23, no. 4: 2308. https://doi.org/10.3390/ijms23042308
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.