
Improvement and Re-Evolution of Tetraploid Wheat for Global Environmental Challenge and Diversity Consumption Demand

 , , , and
, , , and
Abstract
:1. Introduction
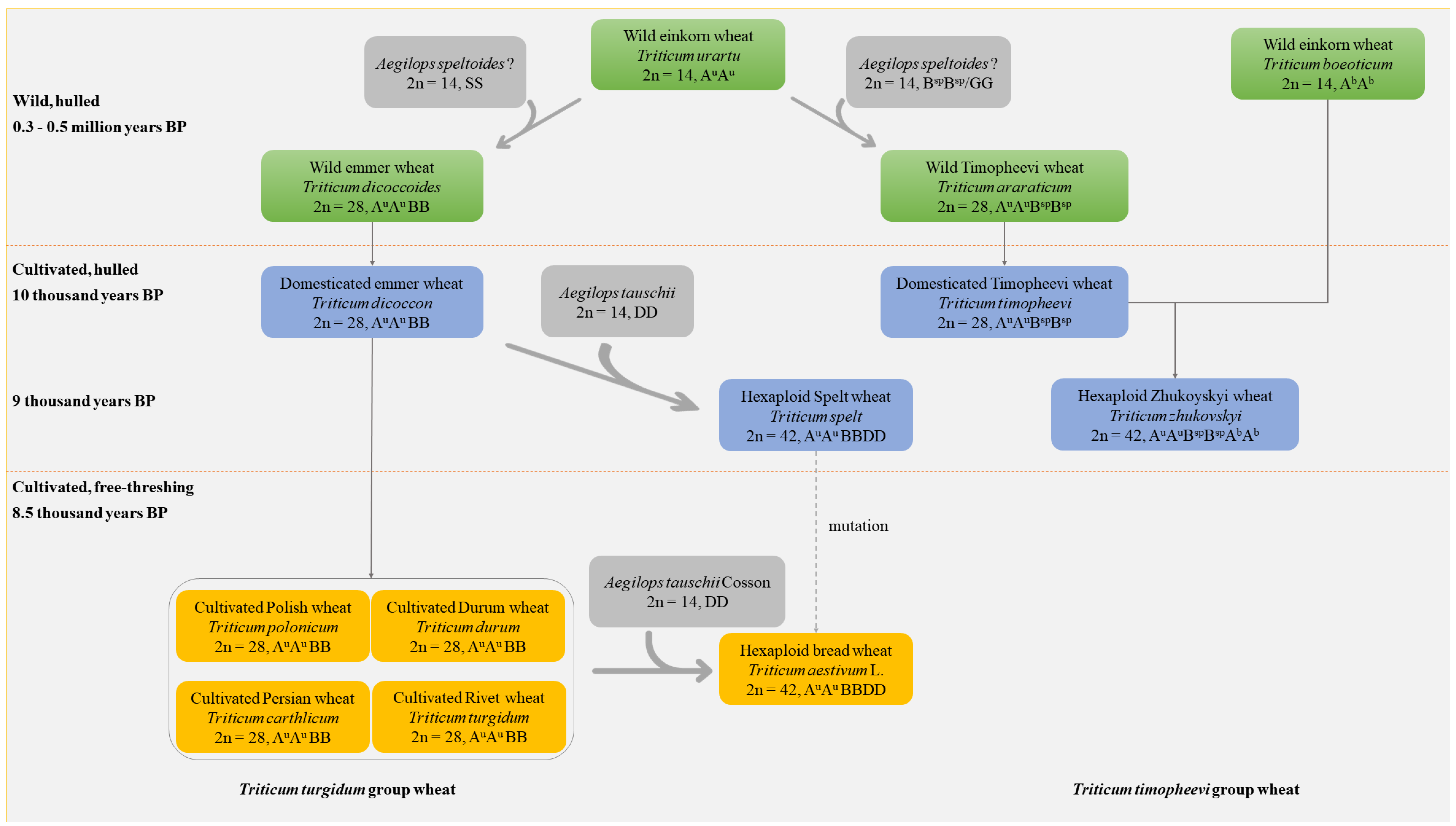
2. Origin, Domestication, and Evolution of Tetraploid Wheat
2.1. Origin Process of Tetraploid Wheat
2.2. Domestication and Evolution Process
2.3. Variations of Major Traits during Domestication and Evolution
2.3.1. Changes from Brittle to Non-Brittle Rachis
2.3.2. Variations from Non-Free Threshing to Free Threshing
2.3.3. Other Qualitative Domestication Traits Involved
2.4. Importance and Breeding Challenge of Durum Wheat
3. Improvement and Re-Evolution of Durum Wheat
3.1. Strategies and Approaches
3.2. Major Achievements
3.2.1. Crop Yield Potential
3.2.2. Grain Quality
3.2.3. Biotic Resistance
3.2.4. Abiotic Tolerance
3.2.5. Wheat-Related Disorders in Humans
4. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sousa, D.T.; Ribeiro, M.; Sabença, C.; Igrejas, G. The 10,000-Year Success Story of Wheat! Foods 2021, 10, 2124. [Google Scholar] [CrossRef] [PubMed]
- Kenzhebayeva, S.; Abekova, A.; Atabayeva, S.; Yernazarova, G.; Omirbekova, N.; Zhang, G.; Turasheva, S.; Asrandina, S.; Sarsu, F.; Wang, Y. Mutant lines of spring wheat with increased iron, zinc, and micronutrients in grains and enhanced bioavailability for human health. BioMed Res. Int. 2019, 2019, 9692053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.-P.; Zhang, Y.-Q.; Tong, Y.-P.; Xue, Y.-F.; Liu, D.-Y.; Zhang, W.; Deng, Y.; Meng, Q.-F.; Yue, S.-C.; Yan, P. Harvesting more grain zinc of wheat for human health. Sci. Rep. 2017, 7, 7016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Faltermaier, A.; Waters, D.; Becker, T.; Arendt, E.; Gastl, M. Common wheat (Triticum aestivum L.) and its use as a brewing cereal—A review. J. Inst. Brew. 2014, 120, 1–15. [Google Scholar] [CrossRef]
- Jouanin, A.; Gilissen, L.J.; Boyd, L.A.; Cockram, J.; Leigh, F.J.; Wallington, E.J.; Van den Broeck, H.C.; Van der Meer, I.M.; Schaart, J.G.; Visser, R.G. Food processing and breeding strategies for coeliac-safe and healthy wheat products. Food Res. Int. 2018, 110, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, T.; Halford, N. Food security: The challenge of increasing wheat yield and the importance of not compromising food safety. Ann. Appl. Biol. 2014, 164, 354–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Guo, J. How does the self-sufficiency rate affect international price volatility transmissions in the wheat sector? Evidence from wheat-exporting countries. Humanit. Soc. Sci. Commun. 2020, 7, 113. [Google Scholar] [CrossRef]
- Tripathi, A.D.; Mishra, R.; Maurya, K.K.; Singh, R.B.; Wilson, D.W. Estimates for world population and global food availability for global health. In The Role of Functional Food Security in Global Health; Elsevier: Amsterdam, The Netherlands, 2019; pp. 3–24. [Google Scholar]
- International Wheat Genome Sequencing Consortium (IWGSC). Wheat—A Key Crop for Food Security. Available online: http://www.wheatgenome.org/News/Media-resources/Fact-Sheets-Infographics/Wheat-A-Key-Crop-for-Food-Security (accessed on 18 April 2018).
- Enghiad, A.; Ufer, D.; Countryman, A.M.; Thilmany, D.D. An overview of global wheat market fundamentals in an era of climate concerns. Int. J. Agron. 2017, 2017, 3931897. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Farooq, M.; Al-Sadi, A.M.; Nawaz, A.; Jabran, K.; Siddique, K.H. Impact of climate change on biology and management of wheat pests. Crop Protect. 2020, 137, 105304. [Google Scholar] [CrossRef]
- Harkness, C.; Semenov, M.A.; Areal, F.; Senapati, N.; Trnka, M.; Balek, J.; Bishop, J. Adverse weather conditions for UK wheat production under climate change. Agric. For. Meteorol. 2020, 282, 107862. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Ochoa, I.M.; Asseng, S.; Kassie, B.T.; Xiong, W.; Robertson, R.; Pequeno, D.N.L.; Sonder, K.; Reynolds, M.; Babar, M.A.; Milan, A.M. Climate change impact on Mexico wheat production. Agric. For. Meteorol. 2018, 263, 373–387. [Google Scholar] [CrossRef]
- Miedaner, T.; Juroszek, P. Climate change will influence disease resistance breeding in wheat in Northwestern Europe. Theor. Appl. Genet. 2021, 134, 1771–1785. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Li Liu, D.; Wang, B.; Feng, P.; Bai, H.; Tang, J. Climate change impact on yields and water use of wheat and maize in the North China Plain under future climate change scenarios. Agric. Water Manag. 2020, 238, 106238. [Google Scholar] [CrossRef]
- Yen, C.; Yang, J. Biosystematics of Triticeae: Volume I. Triticum-Aegilops Complex; Springer Nature: Cham, Switzerland, 2020. [Google Scholar]
- Kihara, H. Discovery of the DD–analyzer, one of the analyzers of Triticum vulgare. Agric. Hortic. 1944, 19, 889–890. [Google Scholar]
- McFadden, E.S.; Sears, E.R. The origin of Triticum spelta and its free-threshing hexaploid relatives. J. Hered. 1946, 37, 81–89. [Google Scholar] [CrossRef]
- Peng, J.; Sun, D.; Nevo, E. Wild emmer wheat, Triticum dicoccoides, occupies a pivotal position in wheat domestication process. Aust. J. Crop Sci. 2011, 5, 1127–1143. [Google Scholar]
- Dubcovsky, J.; Dvorak, J. Genome plasticity a key factor in the success of polyploid wheat under domestication. Science 2007, 316, 1862–1866. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Liu, D.; Yang, W.; Kishii, M.; Mao, L. Synthetic hexaploid wheat: Yesterday, today, and tomorrow. Engineering 2018, 4, 552–558. [Google Scholar] [CrossRef]
- Agriculture and Agri-Food Canada. Canada: Outlook for Principal Field Crops. Available online: https://www.agr.gc.ca/eng/canadas-agriculture-sectors/crops/reports-and-statistics-data-for-canadian-principal-field-crops/canada-outlook-for-principal-field-crops-2021-04-20/?id=1619114343630 (accessed on 20 April 2021).
- Tidiane Sall, A.; Chiari, T.; Legesse, W.; Seid-Ahmed, K.; Ortiz, R.; Van Ginkel, M.; Bassi, F.M. Durum wheat (Triticum durum Desf.): Origin, cultivation and potential expansion in Sub-Saharan Africa. Agronomy 2019, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Xynias, I.N.; Mylonas, I.; Korpetis, E.G.; Ninou, E.; Tsaballa, A.; Avdikos, I.D.; Mavromatis, A.G. Durum wheat breeding in the Mediterranean region: Current status and future prospects. Agronomy 2020, 10, 432. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, Y. Evolution of polyploid Triticum wheats under cultivation: The role of domestication, natural hybridization and allopolyploid speciation in their diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, M.; Zhang, L.; Ning, S.; Huang, L.; Yuan, Z.; Wu, B.; Yan, Z.; Dai, S.; Jiang, B.; Zheng, Y. The Resurgence of Introgression Breeding, as Exemplified in Wheat Improvement. Front. Plant Sci. 2020, 11, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, C.F.; Kiszonas, A.M.; Murray, J.; Boehm, J.D.; Ibba, M.I.; Zhang, M.; Cai, X. Re-evolution of durum wheat by introducing the Hardness and Glu-D1 loci. Front. Sustain. Food Syst. 2019, 3, 103. [Google Scholar] [CrossRef]
- Harlan, J.R.; Zohary, D. Distribution of wild wheats and barley. Science 1966, 153, 1074–1080. [Google Scholar] [CrossRef]
- Abbo, S.; Lev-Yadun, S.; Gopher, A. Agricultural origins: Centers and noncenters; a Near Eastern reappraisal. Crit. Rev. Plant Sci. 2010, 29, 317–328. [Google Scholar] [CrossRef]
- Rahman, S.; Islam, S.; Yu, Z.; She, M.; Nevo, E.; Ma, W. Current progress in understanding and recovering the wheat genes lost in evolution and domestication. Int. J. Mol. Sci. 2020, 21, 5836. [Google Scholar] [CrossRef]
- Dvořák, J.; Terlizzi, P.d.; Zhang, H.-B.; Resta, P. The evolution of polyploid wheats: Identification of the A genome donor species. Genome 1993, 36, 21–31. [Google Scholar] [CrossRef]
- Ling, H.-Q.; Zhao, S.; Liu, D.; Wang, J.; Sun, H.; Zhang, C.; Fan, H.; Li, D.; Dong, L.; Tao, Y. Draft genome of the wheat A-genome progenitor Triticum urartu. Nature 2013, 496, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Haider, N. The origin of the B-genome of bread wheat (Triticum aestivum L.). Russ. J. Genet. 2013, 49, 263–274. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Johnson, B.; Dhaliwal, H. Reproductive isolation of Triticum boeoticum and Triticum urartu and the origin of the tetraploid wheats. Am. J. Bot. 1976, 63, 1088–1094. [Google Scholar] [CrossRef]
- Heun, M.; Schäfer-Pregl, R.; Klawan, D.; Castagna, R.; Accerbi, M.; Borghi, B.; Salamini, F. Site of einkorn wheat domestication identified by DNA fingerprinting. Science 1997, 278, 1312–1314. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.; Sears, E.R. The wild gene resources of wheat. Sci. Am. 1981, 244, 102–113. [Google Scholar] [CrossRef]
- Watanabe, N. Breeding opportunities for early, free-threshing and semi-dwarf Triticum monococcum L. Euphytica 2017, 213, 201. [Google Scholar] [CrossRef]
- Feldman, M.; Levy, A. Allopolyploidy—A shaping force in the evolution of wheat genomes. Cytogenet. Genome Res. 2005, 109, 250–258. [Google Scholar] [CrossRef]
- Pont, C.; Leroy, T.; Seidel, M.; Tondelli, A.; Duchemin, W.; Armisen, D.; Lang, D.; Bustos-Korts, D.; Goué, N.; Balfourier, F. Tracing the ancestry of modern bread wheats. Nat. Genet. 2019, 51, 905–911. [Google Scholar] [CrossRef]
- Daud, H.M.; Gustafson, J. Molecular evidence for Triticum speltoides as a B-genome progenitor of wheat (Triticum aestivum). Genome 1996, 39, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Baum, B.; Edwards, T.; Johnson, D. Phylogenetic relationships among diploid Aegilops species inferred from 5S rDNA units. Mol. Phylogen. Evol. 2009, 53, 34–44. [Google Scholar] [CrossRef]
- Golovnina, K.; Glushkov, S.; Blinov, A.; Mayorov, V.; Adkison, L.; Goncharov, N. Molecular phylogeny of the genus Triticum L. Plant Syst. Evol. 2007, 264, 195–216. [Google Scholar] [CrossRef]
- Li, L.-F.; Zhang, Z.-B.; Wang, Z.-H.; Li, N.; Sha, Y.; Wang, X.-F.; Ding, N.; Li, Y.; Zhao, J.; Wu, Y.; et al. Genome sequences of the five Sitopsis species of Aegilops and the origin of polyploid wheat B-subgenome. Mol. Plant 2022. [Google Scholar] [CrossRef]
- Sears, E. The cytology and genetics of the wheats and their relatives. Adv. Genet. 1948, 2, 239–270. [Google Scholar]
- Huang, S.; Sirikhachornkit, A.; Su, X.; Faris, J.; Gill, B.; Haselkorn, R.; Gornicki, P. Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of the Triticum/Aegilops complex and the evolutionary history of polyploid wheat. Proc. Natl. Acad. Sci. USA 2002, 99, 8133–8138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281–301. [Google Scholar] [CrossRef]
- Wagenaar, E. Studies on the genome constitution of Triticum timopheevi Zhuk. II. The T. timopheevi complex and its origin. Evolution 1966, 20, 150–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charmet, G. Wheat domestication: Lessons for the future. Comptes Rendus Biol. 2011, 334, 212–220. [Google Scholar] [CrossRef]
- Dvorak, J.; Akhunov, E.D. Tempos of gene locus deletions and duplications and their relationship to recombination rate during diploid and polyploid evolution in the Aegilops-Triticum alliance. Genetics 2005, 171, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.; Kislev, M.E. Domestication of emmer wheat and evolution of free-threshing tetraploid wheat. Isr. J. Plant Sci. 2007, 55, 207–221. [Google Scholar] [CrossRef]
- Kislev, M.E.; Nadel, D.; Carmi, I. Epipalaeolithic (19,000 BP) cereal and fruit diet at Ohalo II, Sea of Galilee, Israel. Rev. Palaeobot. Palynol. 1992, 73, 161–166. [Google Scholar] [CrossRef]
- Özkan, H.; Willcox, G.; Graner, A.; Salamini, F.; Kilian, B. Geographic distribution and domestication of wild emmer wheat (Triticum dicoccoides). Genet. Resour. Crop Evol. 2010, 58, 11–53. [Google Scholar] [CrossRef]
- Aaronsohn, A.; Schweinfurth, G. Die auffindung des wilden emmers (Triticum dicoccum) in Nordpalästina. In Altneuland Monatsschrift für die Irtschaft Erschliessung Palästinas; Kessinger Publishing: Whitefish, MT, USA, 1906; Volume 7, pp. 213–220. [Google Scholar]
- Tanno, K.-I.; Willcox, G. How fast was wild wheat domesticated? Science 2006, 311, 1886. [Google Scholar] [CrossRef] [Green Version]
- Mori, N.; Moriguchi, T.; Nakamura, C. RFLP analysis of nuclear DNA for study of phylogeny and domestication of tetraploid wheat. Genes Genet. Syst. 1997, 72, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, M. Wheat evolution: Integrating archaeological and biological evidence. In Wheat Taxonomy: The Legacy of John Percival; The Linnean Society of London: London, UK, 2001. [Google Scholar]
- Feldman, M. Origin of cultivated wheat. In The World Wheat Book; Bonjean, A.P., Angus, W.J., Eds.; Lavoisier Publishing: Paris, France, 2001; pp. 3–56. [Google Scholar]
- Salamini, F.; Özkan, H.; Brandolini, A.; Schäfer-Pregl, R.; Martin, W. Genetics and geography of wild cereal domestication in the near east. Nat. Rev. Genet. 2002, 3, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Nave, M.; Taş, M.; Raupp, J.; Tiwari, V.K.; Ozkan, H.; Poland, J.; Hale, I.; Komatsuda, T.; Distelfeld, A. The independent domestication of Timopheev’s wheat: Insights from haplotype analysis of the Brittle rachis 1 (BTR1-A) gene. Genes 2021, 12, 338. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Kondo, Y.; Ishii, T.; Kawahara, T.; Nakamura, C. Genetic diversity and origin of timopheevi wheat inferred by chloroplast DNA fingerprinting. Breed. Sci. 2009, 59, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.-C.; Yang, Z.-L.; You, F.; Kawahara, T.; Waines, J.; Dvorak, J. The structure of wild and domesticated emmer wheat populations, gene flow between them, and the site of emmer domestication. Theor. Appl. Genet. 2007, 114, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Upadhya, M.; Swaminathan, M. Genome analysis in Triticum zhukovskyi, a new hexaploid wheat. Chromosoma 1963, 14, 589–600. [Google Scholar] [CrossRef]
- Mori, N.; Ishii, T.; Ishido, T.; Hirosawa, S.; Watatani, H.; Kawahara, T.; Nesbitt, M.; Belay, G.; Takumi, S.; Ogihara, Y.; et al. Origins of domesticated emmer and common wheat inferred from chloroplast DNA fingerprinting. In Proceedings of the Tenth International Wheat Genetics Symposium, Paestum, Italy, 1–6 September 2003; pp. 25–28. [Google Scholar]
- Salamini, F.; Heun, M.; Ozkan, H.; Wunder, J. On DNA markers, phylogenetic trees and mode of origin of crops. Genome 2004, 46, 448–453. [Google Scholar]
- Özkan, H.; Brandolini, A.; Schäfer-Pregl, R.; Salamini, F. AFLP analysis of a collection of tetraploid wheats indicates the origin of emmer and hard wheat domestication in southeast Turkey. Mol. Biol. Evol. 2002, 19, 1797–1801. [Google Scholar] [CrossRef] [Green Version]
- Salamini, F.; Heun, M.; Brandolini, A.; Ozkan, H.; Wunder, J. Comment on “AFLP data and the origins of domesticated crops”. Genome 2004, 47, 615–620. [Google Scholar] [CrossRef] [Green Version]
- Özkan, H.; Brandolini, A.; Pozzi, C.; Effgen, S.; Wunder, J.; Salamini, F. A reconsideration of the domestication geography of tetraploid wheats. Theor. Appl. Genet. 2005, 110, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Gill, B.S. Multiple genetic pathways for seed shattering in the grasses. Funct. Integr. Genom. 2006, 6, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Ronin, Y.; Fahima, T.; Röder, M.S.; Li, Y.; Nevo, E.; Korol, A. Domestication quantitative trait loci in Triticum dicoccoides, the progenitor of wheat. Proc. Natl. Acad. Sci. USA 2003, 100, 2489–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balter, M. Seeking agriculture’s ancient roots. Science 2007, 316, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Fujii, Y.; Kato, N.; Ban, T.; Martinek, P. Microsatellite mapping of the genes for brittle rachis on homoeologous group 3 chromosomes in tetraploid and hexaploid wheats. J. Appl. Genet. 2006, 47, 93–98. [Google Scholar] [CrossRef]
- Chen, Q.-F.; Yen, C.; Yang, J.-L. Chromosome location of the gene for brittle rachis in the Tibetan weedrace of common wheat. Genet. Resour. Crop Evol. 1998, 45, 407–410. [Google Scholar] [CrossRef]
- Watanabe, N.; Takesada, N.; Fujii, Y.; Martinek, P. Comparative mapping of genes for brittle rachis in Triticum and Aegilops. Czech J. Genet. Plant Breed 2005, 41, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Nalam, V.J.; Vales, M.I.; Watson, C.J.; Kianian, S.F.; Riera-Lizarazu, O. Map-based analysis of genes affecting the brittle rachis character in tetraploid wheat (Triticum turgidum L.). Theor. Appl. Genet. 2006, 112, 373–381. [Google Scholar] [CrossRef]
- Pourkheirandish, M.; Hensel, G.; Kilian, B.; Senthil, N.; Chen, G.; Sameri, M.; Azhaguvel, P.; Sakuma, S.; Dhanagond, S.; Sharma, R. Evolution of the grain dispersal system in barley. Cell 2015, 162, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Avni, R.; Nave, M.; Barad, O.; Baruch, K.; Twardziok, S.O.; Gundlach, H.; Hale, I.; Mascher, M.; Spannagl, M.; Wiebe, K. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 2017, 357, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Pourkheirandish, M.; Dai, F.; Sakuma, S.; Kanamori, H.; Distelfeld, A.; Willcox, G.; Kawahara, T.; Matsumoto, T.; Kilian, B.; Komatsuda, T. On the origin of the non-brittle rachis trait of domesticated einkorn wheat. Front. Plant Sci. 2018, 8, 2031. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Scoles, G.J.; Hucl, P. The genetics of rachis fragility and glume tenacity in semi-wild wheat. Euphytica 1997, 94, 119–124. [Google Scholar] [CrossRef]
- Watanabe, N.; Sugiyama, K.; Yamagishi, Y.; Sakata, Y. Comparative telosomic mapping of homoeologous genes for brittle rachis in tetraploid and hexaploid wheats. Hereditas 2002, 137, 180–185. [Google Scholar] [CrossRef]
- Peng, J.; Liu, Z.; Liu, X.; Yan, J.; Sun, D.; Nevo, E. Evolutionary agriculture domestication of wild emmer wheat. In New Horizons in Evolution; Elsevier: Amsterdam, The Netherlands, 2021; pp. 193–255. [Google Scholar]
- Sharma, J.S.; Running, K.L.; Xu, S.S.; Zhang, Q.; Haugrud, A.R.P.; Sharma, S.; McClean, P.E.; Faris, J.D. Genetic analysis of threshability and other spike traits in the evolution of cultivated emmer to fully domesticated durum wheat. Mol. Genet. Genom. 2019, 294, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, J.; Deal, K.R.; Luo, M.-C.; You, F.M.; von Borstel, K.; Dehghani, H. The origin of spelt and free-threshing hexaploid wheat. J. Hered. 2012, 103, 426–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerber, E.; Rowland, G. Origin of the free threshing character in hexaploid wheat. Can. J. Genet. Cytol. 1974, 16, 145–154. [Google Scholar] [CrossRef]
- Chen, Q.-F.; Yen, C.; Yang, J.-L. Chromosome location of the gene for the hulled character in the Tibetan weedrace of common wheat. Genet. Resour. Crop Evol. 1999, 46, 543–546. [Google Scholar] [CrossRef]
- Nalam, V.J.; Vales, M.I.; Watson, C.J.; Johnson, E.B.; Riera-Lizarazu, O. Map-based analysis of genetic loci on chromosome 2D that affect glume tenacity and threshability, components of the free-threshing habit in common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 116, 135–145. [Google Scholar] [CrossRef]
- Faris, J.D.; Gill, B.S. Genomic targeting and high-resolution mapping of the domestication gene Q in wheat. Genome 2002, 45, 706–718. [Google Scholar] [CrossRef]
- Simons, K.J.; Fellers, J.P.; Trick, H.N.; Zhang, Z.; Tai, Y.-S.; Gill, B.S.; Faris, J.D. Molecular characterization of the major wheat domestication gene Q. Genetics 2006, 172, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Sun, G.; Kong, X.; Jia, M.; Wang, K.; Ye, X.; Zhou, Y.; Geng, S.; Mao, L.; Li, A. The soft glumes of common wheat are sterile-lemmas as determined by the domestication gene Q. Crop J. 2019, 7, 113–117. [Google Scholar] [CrossRef]
- Faris, J.D.; Zhang, Z.; Chao, S. Map-based analysis of the tenacious glume gene Tg-B1 of wild emmer and its role in wheat domestication. Gene 2014, 542, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Faris, J.D.; Zhang, Q.; Chao, S.; Zhang, Z.; Xu, S.S. Analysis of agronomic and domestication traits in a durum × cultivated emmer wheat population using a high-density single nucleotide polymorphism-based linkage map. Theor. Appl. Genet. 2014, 127, 2333–2348. [Google Scholar] [CrossRef] [PubMed]
- Jantasuriyarat, C.; Vales, M.; Watson, C.; Riera-Lizarazu, O. Identification and mapping of genetic loci affecting the free-threshing habit and spike compactness in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 108, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, K.; Tang, H.; Gong, Q.; Du, L.; Pei, X.; Ye, X. CRISPR/Cas9 editing of wheat TaQ genes alters spike morphogenesis and grain threshability. J. Genet. Genom. 2020, 47, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Debernardi, J.M.; Lin, H.; Chuck, G.; Faris, J.D.; Dubcovsky, J. microRNA172 plays a crucial role in wheat spike morphogenesis and grain threshability. Development 2017, 144, 1966–1975. [Google Scholar] [PubMed] [Green Version]
- Greenwood, J.R.; Finnegan, E.J.; Watanabe, N.; Trevaskis, B.; Swain, S.M. New alleles of the wheat domestication gene Q reveal multiple roles in growth and reproductive development. Development 2017, 144, 1959–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Belcram, H.; Gornicki, P.; Charles, M.; Just, J.; Huneau, C.; Magdelenat, G.; Couloux, A.; Samain, S.; Gill, B.S.; et al. Duplication and partitioning in evolution and function of homoeologous Q loci governing domestication characters in polyploid wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 18737–18742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muramatsu, M. Dosage effect of the spelta gene q of hexaploid wheat. Genetics 1963, 48, 469. [Google Scholar] [CrossRef] [PubMed]
- Faris, J.; Simons, K.J.; Zhang, Z.; Gill, B. The wheat super domestication gene Q. Front. Wheat Biosci. 2005, 129–148. [Google Scholar]
- Vaughan, D.; Balazs, E.; Heslop-Harrison, J. From crop domestication to super-domestication. Ann. Bot. 2007, 100, 893–901. [Google Scholar] [CrossRef]
- Xie, Q.; Li, N.; Yang, Y.; Lv, Y.; Yao, H.; Wei, R.; Sparkes, D.L.; Ma, Z. Pleiotropic effects of the wheat domestication gene Q on yield and grain morphology. Planta 2018, 247, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Gegas, V.C.; Nazari, A.; Griffiths, S.; Simmonds, J.; Fish, L.; Orford, S.; Sayers, L.; Doonan, J.H.; Snape, J.W. A genetic framework for grain size and shape variation in wheat. Plant Cell 2010, 22, 1046–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiger, D.; Elias, E.; Cantrell, R. Evaluation of lines derived from wild emmer chromosome substitutions: I. Quality traits. Crop Sci. 1996, 36, 223–227. [Google Scholar] [CrossRef]
- Campbell, B.; Baenziger, P.S.; Gill, K.; Eskridge, K.M.; Budak, H.; Erayman, M.; Dweikat, I.M.; Yen, Y. Identification of QTLs and environmental interactions associated with agronomic traits on chromosome 3A of wheat. Crop Sci. 2003, 43, 1493–1505. [Google Scholar] [CrossRef]
- Kato, K.; Miura, H.; Sawada, S. Mapping QTLs controlling grain yield and its components on chromosome 5A of wheat. Theor. Appl. Genet. 2000, 101, 1114–1121. [Google Scholar] [CrossRef]
- Liu, J.; Tang, H.; Qu, X.; Liu, H.; Li, C.; Tu, Y.; Li, S.; Habib, A.; Mu, Y.; Dai, S. A novel, major, and validated QTL for the effective tiller number located on chromosome arm 1BL in bread wheat. Plant Mol. Biol. 2020, 104, 173–185. [Google Scholar] [CrossRef]
- Spielmeyer, W.; Richards, R. Comparative mapping of wheat chromosome 1AS which contains the tiller inhibition gene (tin) with rice chromosome 5S. Theor. Appl. Genet. 2004, 109, 1303–1310. [Google Scholar] [CrossRef]
- Cockram, J.; Jones, H.; Leigh, F.J.; O’Sullivan, D.; Powell, W.; Laurie, D.A.; Greenland, A.J. Control of flowering time in temperate cereals: Genes, domestication, and sustainable productivity. J. Exp. Bot. 2007, 58, 1231–1244. [Google Scholar] [CrossRef]
- Würschum, T.; Langer, S.M.; Longin, C.F.H.; Tucker, M.R.; Leiser, W.L. A modern Green Revolution gene for reduced height in wheat. Plant J. 2017, 92, 892–903. [Google Scholar] [CrossRef] [Green Version]
- Pearce, S.; Saville, R.; Vaughan, S.P.; Chandler, P.M.; Wilhelm, E.P.; Sparks, C.A.; Al-Kaff, N.; Korolev, A.; Boulton, M.I.; Phillips, A.L. Molecular characterization of Rht-1 dwarfing genes in hexaploid wheat. Plant Physiol. 2011, 157, 1820–1831. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P. The genes of the Green Revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Chai, S.; Yao, Q.; Zhang, X.; Xiao, X.; Fan, X.; Zeng, J.; Sha, L.; Kang, H.; Zhang, H.; Li, J. The semi-dwarfing gene Rht-dp from dwarf polish wheat (Triticum polonicum L.) is the “Green Revolution” gene Rht-B1b. BMC Genom. 2021, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-Y.; Lin, L.-J.; Song, Z.-J.; Yuan, J.-Y.; Zhong, M.-Y.; Zhang, H.-Q.; Fan, X.; Sha, L.-N.; Wang, Y.; Xu, L.-L. Identification, fine mapping and characterization of Rht-dp, a recessive wheat dwarfing (reduced height) gene derived from Triticum polonicum. Genes Genom. 2012, 34, 509–515. [Google Scholar] [CrossRef]
- Zaharieva, M.; Monneveux, P. Cultivated einkorn wheat (Triticum monococcum L. subsp. monococcum): The long life of a founder crop of agriculture. Genet. Resour. Crop Evol. 2014, 61, 677–706. [Google Scholar] [CrossRef]
- Arzani, A. Emmer (Triticum turgidum ssp. dicoccum) flour and bread. In Flour and Breads and Their Fortification in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2019; pp. 89–98. [Google Scholar]
- Cooper, R. Re-discovering ancient wheat varieties as functional foods. J. Tradit. Complementary Med. 2015, 5, 138–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magallanes-López, A.M.; Ammar, K.; Morales-Dorantes, A.; González-Santoyo, H.; Crossa, J.; Guzmán, C. Grain quality traits of commercial durum wheat varieties and their relationships with drought stress and glutenins composition. J. Cereal Sci. 2017, 75, 1–9. [Google Scholar] [CrossRef]
- Elias, E. Durum wheat products. In Durum Wheat Quality in the Mediterranean Region; Options Méditerranéennes: Série A. Séminaires Méditerranéens, n. 22; International Center for Advanced Mediterranean Agronomic Studies (CIHEAM): Saragosa, Spain, 1995; Volume 40, pp. 23–31. [Google Scholar]
- International Pasta Organisation (IPO). Pasta, World Consumption Boom Since The Lockdown Started 1 Consumer Out of 4 Ate More, 25% Export Increase in 6 Months. Available online: https://internationalpasta.org/news/pasta-world-consumption-boom-since-the-lockdown-started-1-consumer-out-of-4-ate-more-25-export-increase-in-6-months/ (accessed on 16 October 2020).
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, P.; Raskina, O.; Ma, X.; Nevo, E. Wheat evolution, domestication, and improvement. In Making of a Wheat Plant; John Wiley & Sons (Wiley): Hoboken, NJ, USA, 2009; pp. 5–30. [Google Scholar]
- Padmanaban, S.; Zhang, P.; Hare, R.A.; Sutherland, M.W.; Martin, A. Pentaploid wheat hybrids: Applications, characterisation, and challenges. Front. Plant Sci. 2017, 8, 358. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.B.; Somers, D.J. Genome-wide reduction of genetic diversity in wheat breeding. Crop Sci. 2009, 49, 161–168. [Google Scholar] [CrossRef]
- Haudry, A.; Cenci, A.; Ravel, C.; Bataillon, T.; Brunel, D.; Poncet, C.; Hochu, I.; Poirier, S.; Santoni, S.; Glémin, S. Grinding up wheat: A massive loss of nucleotide diversity since domestication. Mol. Biol. Evol. 2007, 24, 1506–1517. [Google Scholar] [CrossRef] [Green Version]
- Nevo, E. Evolution of wild emmer wheat and crop improvement. J. Syst Evol. 2014, 52, 673–696. [Google Scholar] [CrossRef]
- Xie, W.; Nevo, E. Wild emmer: Genetic resources, gene mapping and potential for wheat improvement. Euphytica 2008, 164, 603–614. [Google Scholar] [CrossRef]
- El Haddad, N.; Kabbaj, H.; Zaïm, M.; El Hassouni, K.; Tidiane Sall, A.; Azouz, M.; Ortiz, R.; Baum, M.; Amri, A.; Gamba, F. Crop wild relatives in durum wheat breeding: Drift or thrift? Crop Sci. 2021, 61, 37–54. [Google Scholar] [CrossRef]
- Ogbonnaya, F.; Halloran, G.; Lagudah, E. D genome of wheat—60 years on from Kihara, Sears and McFadden. Front. Wheat BioSci. 2005, 11, 205–220. [Google Scholar]
- Mastrangelo, A.M.; Cattivelli, L. What Makes Bread and Durum Wheat Different? Trends Plant Sci. 2021, 26, 677–684. [Google Scholar] [CrossRef]
- Li, A.; Liu, D.; Wu, J.; Zhao, X.; Hao, M.; Geng, S.; Yan, J.; Jiang, X.; Zhang, L.; Wu, J. mRNA and small RNA transcriptomes reveal insights into dynamic homoeolog regulation of allopolyploid heterosis in nascent hexaploid wheat. Plant Cell 2014, 26, 1878–1900. [Google Scholar] [CrossRef] [Green Version]
- Sharma, H.; Waines, J.; Foster, K. Variability in Primitive and Wild Wheats for Useful Genetic Characters 1. Crop Sci. 1981, 21, 555–559. [Google Scholar] [CrossRef]
- Monneveux, P.; Zaharieva, M.; Rekika, D.; Royo, C.; Nachit, M.; Fonzo, N.; Araus, J. The utilisation of Triticum and Aegilops species for the improvement of durum wheat. In Durum Wheat Improvement in the Mediterranean Region: New Challenges; Options Méditerranéennes: Série A. Séminaires Méditerranéens, n. 40; International Center for Advanced Mediterranean Agronomic Studies (CIHEAM): Saragosa, Spain, 2000; pp. 71–81. [Google Scholar]
- Mohammadi, M.; Mirlohi, A.; Majidi, M.M.; Kartalaei, E.S. Emmer wheat as a source for trait improvement in durum wheat: A study of general and specific combining ability. Euphytica 2021, 217, 64. [Google Scholar] [CrossRef]
- Giorgi, B. A homoeologous pairing mutant isolated in Triticum durum cv. Cappelli. Mutat. Breed. Newsl. 1978, 11, 4–5. [Google Scholar]
- Sears, E.R. Genetics society of canada award of excellence lecture an induced mutant with homoeologous pairing in common wheat. Can. J. Genet. Cytol. 1977, 19, 585–593. [Google Scholar] [CrossRef]
- Joppa, L.; Williams, N. Langdon durum disomic substitution lines and aneuploid analysis in tetraploid wheat. Genome 1988, 30, 222–228. [Google Scholar] [CrossRef]
- Joppa, L.; Williams, N. A Disomic-5D-Nullisomic-5B Substitution Line of Durum Wheat 1. Crop Sci. 1979, 19, 509–511. [Google Scholar] [CrossRef]
- Joppa, L. Development of D-genome Disomic Substitution lines of Durum wheat (Triticum turgidum L.). In Fourth International Wheat Genetics Symposium Cytogenetics; University of Missouri-Columbia: Columbia, MI, USA, 1973; pp. 685–690. [Google Scholar]
- Gupta, P.; Langridge, P.; Mir, R. Marker-assisted wheat breeding: Present status and future possibilities. Mol. Breed. 2010, 26, 145–161. [Google Scholar] [CrossRef]
- Varshney, R.K.; Bohra, A.; Yu, J.; Graner, A.; Zhang, Q.; Sorrells, M.E. Designing future crops: Genomics-assisted breeding comes of age. Trends Plant Sci. 2021, 26, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yang, F.; Zhang, J.; Liu, H.; Rahman, S.; Islam, S.; Ma, W.; She, M. Application of CRISPR/Cas9 in Crop Quality Improvement. Int. J. Mol. Sci. 2021, 22, 4206. [Google Scholar] [CrossRef]
- Habash, D.; Kehel, Z.; Nachit, M. Genomic approaches for designing durum wheat ready for climate change with a focus on drought. J. Exp. Bot. 2009, 60, 2805–2815. [Google Scholar] [CrossRef]
- Siddiqui, H.A.; Harvey-Samuel, T.; Mansoor, S. Gene drive: A faster route to plant improvement. Trends Plant Sci. 2021, 26, 1204–1206. [Google Scholar] [CrossRef]
- McClung, A.M.; Cantrell, R.; Quick, J.; Gregory, R. Influence of the Rht1 Semidwarf Gene on Yield, Yield Components, and Grain Protein in Durum Wheat 1. Crop Sci. 1986, 26, 1095–1099. [Google Scholar] [CrossRef]
- Duan, S.; Zhao, Z.; Qiao, Y.; Cui, C.; Morgunov, A.; Condon, A.G.; Chen, L.; Hu, Y.-G. GAR dwarf gene Rht14 reduced plant height and affected agronomic traits in durum wheat (Triticum durum). Field Crop Res. 2020, 248, 107721. [Google Scholar] [CrossRef]
- Zhao, Z.; Duan, S.; Hao, J.; Cui, C.; Yang, Y.; Condon, A.G.; Chen, F.; Hu, Y.-G.; Chen, L. The dwarf gene Rht15 improved lodging resistance but differentially affected agronomic and quality traits in durum wheat. Field Crop Res. 2021, 263, 108058. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, C.; Yan, G.; Liu, C. Improving yield and quality traits of durum wheat by introgressing chromosome segments from hexaploid wheat. Genet. Mol. Res. 2013, 12, 6120–6129. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liu, Q.; Wang, Q.; Yang, N.; Li, J.; Wan, H.; Liu, Z.; Yang, S.; Wang, Y.; Zhang, J. Characterization of the Durum Wheat-Aegilops tauschii 4D (4B) Disomic Substitution Line YL-443 With Superior Characteristics of High Yielding and Stripe Rust Resistance. Front. Plant Sci. 2021, 12, 745290. [Google Scholar] [CrossRef] [PubMed]
- Sestili, F.; Pagliarello, R.; Zega, A.; Saletti, R.; Pucci, A.; Botticella, E.; Masci, S.; Tundo, S.; Moscetti, I.; Foti, S. Enhancing grain size in durum wheat using RNAi to knockdown GW2 genes. Theor. Appl. Genet. 2019, 132, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Gazza, L.; Zanella, L.; Pogna, N. Development of durum wheat (Triticum turgidum ssp. durum) lines with soft kernel texture by chromosome engineering. In Proceedings of the 11th International Wheat Genetics Symposium, Brisbane, Australia, 24–29 August 2008; Sydney University Press: Sydney, Australia, 2008. [Google Scholar]
- Morris, C.F.; Simeone, M.C.; King, G.; Lafiandra, D. Transfer of soft kernel texture from Triticum aestivum to durum wheat, Triticum turgidum ssp. durum. Crop Sci. 2011, 51, 114–122. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Q.; Li, X.; Xiao, X.; Sun, F.; Wang, C.; Hu, W.; Feng, Z.; Chang, J.; Chen, M. Coexpression of the high molecular weight glutenin subunit 1Ax1 and puroindoline improves dough mixing properties in durum wheat (Triticum turgidum L. ssp. durum). PLoS ONE 2012, 7, e50057. [Google Scholar]
- Sissons, M.; Pleming, D.; Sestili, F.; Lafiandra, D. Effect of Glu-D1 gene introgression and amylose content on breadmaking potential of blends of durum and hexaploid wheat. Cereal Chem. 2019, 96, 193–206. [Google Scholar] [CrossRef]
- Ceoloni, C.; Ciaffi, M.; Lafiandra, D.; Giorgi, B. Chromosome engineering as a means of transferring 1D storage protein genes from common to durum wheat. In Proceedings of the Eighth International Wheat Genetics Symposium, Beijing, China, 20–25 July 1993; pp. 159–163. [Google Scholar]
- Ceoloni, C.; Biagetti, M.; Ciaffi, M.; Forte, P.; Pasquini, M. Wheat chromosome engineering at the 4x level: The potential of different alien gene transfers into durum wheat. Euphytica 1996, 89, 87–97. [Google Scholar] [CrossRef]
- Vitellozzi, F.; Ciaffi, M.; Dominici, L.; Ceoloni, C. Isolation of a chromosomally engineered durum wheat line carrying the common wheat Glu-D1d allele. Agronomie 1997, 17, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Gennaro, A.; Forte, P.; Panichi, D.; Lafiandra, D.; Pagnotta, M.A.; D’Egidio, M.G.; Ceoloni, C. Stacking small segments of the 1D chromosome of bread wheat containing major gluten quality genes into durum wheat: Transfer strategy and breeding prospects. Mol. Breed. 2012, 30, 149–167. [Google Scholar] [CrossRef]
- Lukaszewski, A. Registration of six germplasms of bread wheat having variations of cytogenetically engineered wheat-rye translocation 1RS. 1BL.(Registrations Of Germplasm). Crop Sci. 2003, 43, 1137–1139. [Google Scholar] [CrossRef]
- Kovacs, M.; Howes, N.; Clarke, J.; Leisle, D. Quality Characteristics of Durum Wheat Lines Deriving High Protein From a Triticum dicoccoides (6b) Substitution. J. Cereal Sci. 1998, 27, 47–51. [Google Scholar] [CrossRef]
- Yu, S.; Li, M.; Dubcovsky, J.; Tian, L. Mutant combinations of lycopene ε-cyclase and β-carotene hydroxylase 2 homoeologs increased β-carotene accumulation in endosperm of tetraploid wheat (Triticum turgidum L.) grains. Plant Biotechnol. J. 2021, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Zhou, Y.; Wight, C.; Pu, Z.; Qi, P.; Jiang, Q.; Deng, M.; Wang, Z.; Wei, Y. Conferring resistance to pre-harvest sprouting in durum wheat by a QTL identified in Triticum spelta. Euphytica 2017, 213, 19. [Google Scholar] [CrossRef]
- Ceoloni, C.; Forte, P.; Gennaro, A.; Micali, S.; Carozza, R.; Bitti, A. Recent developments in durum wheat chromosome engineering. Cytogenet. Genome Res. 2005, 109, 328–334. [Google Scholar] [CrossRef]
- Ceoloni, C.; Kuzmanović, L.; Ruggeri, R.; Rossini, F.; Forte, P.; Cuccurullo, A.; Bitti, A. Harnessing genetic diversity of wild gene pools to enhance wheat crop production and sustainability: Challenges and opportunities. Diversity 2017, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Zaim, M.; El Hassouni, K.; Gamba, F.; Filali-Maltouf, A.; Belkadi, B.; Sourour, A.; Amri, A.; Nachit, M.; Taghouti, M.; Bassi, F.M. Wide crosses of durum wheat (Triticum durum Desf.) reveal good disease resistance, yield stability, and industrial quality across Mediterranean sites. Field Crop Res. 2017, 214, 219–227. [Google Scholar] [CrossRef]
- Rinaldo, A.; Gilbert, B.; Boni, R.; Krattinger, S.G.; Singh, D.; Park, R.F.; Lagudah, E.; Ayliffe, M. The Lr34 adult plant rust resistance gene provides seedling resistance in durum wheat without senescence. Plant Biotechnol. J. 2017, 15, 894–905. [Google Scholar] [CrossRef] [Green Version]
- Kerber, E.; Dyck, P. Inheritance of stem rust resistance transferred from diploid wheat (Triticum monococcum) to tetraploid and hexaploid wheat and chromosome location of the gene involved. Can. J. Genet. Cytol. 1973, 15, 397–409. [Google Scholar] [CrossRef]
- Ma, H.; Singh, R.; Mujeeb-Kazi, A. Resistance to stripe rust in durum wheats, A-genome diploids, and their amphiploids. Euphytica 1997, 94, 279–286. [Google Scholar] [CrossRef]
- Plamenov, D.; Belchev, I.; Kiryakova, V.; Spetsov, P. Fungal resistance of Triticum durum-T. monococcum ssp. aegilopoides amphiploid. J. Plant Dis. Prot. 2009, 116, 60–62. [Google Scholar] [CrossRef]
- Megyeri, M.; Mikó, P.; Molnár, I.; Kovács, G. Development of synthetic amphiploids based on different Triticum turgidum × T. monococcum crosses to improve the adaptability of cereals. Acta Agron. Hung. 2015, 59, 267–274. [Google Scholar] [CrossRef]
- Prat, N.; Guilbert, C.; Prah, U.; Wachter, E.; Steiner, B.; Langin, T.; Robert, O.; Buerstmayr, H. QTL mapping of Fusarium head blight resistance in three related durum wheat populations. Theor. Appl. Genet. 2017, 130, 13–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Leng, Y.; Chao, S.; Xu, S.S.; Zhong, S. Molecular mapping of QTL for Fusarium head blight resistance introgressed into durum wheat. Theor. Appl. Genet. 2018, 131, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Giancaspro, A.; Giove, S.L.; Zito, D.; Blanco, A.; Gadaleta, A. Mapping QTLs for Fusarium head blight resistance in an interspecific wheat population. Front. Plant Sci. 2016, 7, 1381. [Google Scholar] [CrossRef] [PubMed]
- Giancaspro, A.; Lionetti, V.; Giove, S.L.; Zito, D.; Fabri, E.; Reem, N.; Zabotina, O.A.; De Angelis, E.; Monaci, L.; Bellincampi, D. Cell wall features transferred from common into durum wheat to improve Fusarium Head Blight resistance. Plant Sci. 2018, 274, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Rai, K.M.; Pirseyedi, S.; Elias, E.M.; Xu, S.; Dill-Macky, R.; Kianian, S.F. Epigenetic regulation of gene expression improves Fusarium head blight resistance in durum wheat. Sci. Rep. 2020, 10, 17610. [Google Scholar] [CrossRef] [PubMed]
- Siham, O.; Abderraouf, H.; Saadia, L.; Nasserlhaq, N. Phenotypic characterization of recombinant inbred durum wheat lines segregating for resistance to Hessian fly (Mayetiola destructor (Say),) in Morocco. Rev. Nat. Technol. 2017, 9, 18. [Google Scholar]
- Rampino, P.; Mita, G.; Pataleo, S.; De Pascali, M.; Di Fonzo, N.; Perrotta, C. Acquisition of thermotolerance and HSP gene expression in durum wheat (Triticum durum Desf.) cultivars. Environ. Exp. Bot. 2009, 66, 257–264. [Google Scholar] [CrossRef]
- Khateeb, W.A.; Muhaidat, R.; Alahmed, S.; Al Zoubi, M.S.; Al-Batayneh, K.M.; El-Oqlah, A.; Gamar, M.A.; Hussein, E.; Aljabali, A.A.; Alkaraki, A.K. Heat shock proteins gene expression and physiological responses in durum wheat (Triticum durum) under salt stress. Physiol. Mol. Biol. Plants 2020, 26, 1599–1608. [Google Scholar] [CrossRef]
- Bacher, H.; Zhu, F.; Gao, T.; Liu, K.; Dhatt, B.K.; Awada, T.; Zhang, C.; Distelfeld, A.; Yu, H.; Peleg, Z. Wild emmer introgression alters root-to-shoot growth dynamics in durum wheat in response to water stress. Plant Physiol. 2021, 187, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Ayadi, M.; Brini, F.; Masmoudi, K. Overexpression of a wheat aquaporin gene, TdPIP2; 1, enhances salt and drought tolerance in transgenic durum wheat cv. Maali. Int. J. Mol. Sci. 2019, 20, 2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzedine, F.; Gherroucha, H.; Baka, M. Improvement of salt tolerance in durum wheat by ascorbic acid application. J. Stress Physiol. Biochem 2011, 7, 27–37. [Google Scholar]
- Luo, M.; Dubcovsky, J.; Goyal, S.; Dvořák, J. Engineering of interstitial foreign chromosome segments containing the K+/Na+ selectivity gene Kna1 by sequential homoeologous recombination in durum wheat. Theor. Appl. Genet. 1996, 93, 1180–1184. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Ryan, P.R.; Yan, Z.; Delhaize, E. Introgression of a 4D chromosomal fragment into durum wheat confers aluminium tolerance. Ann. Bot. 2014, 114, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Zhang, P.; Ryan, P.R.; Rathjen, T.M.; Yan, Z.; Delhaize, E. Introgression of genes from bread wheat enhances the aluminium tolerance of durum wheat. Theor. Appl. Genet. 2016, 129, 729–739. [Google Scholar] [CrossRef]
- Munns, R.; Hare, R.; James, R.; Rebetzke, G. Genetic variation for improving the salt tolerance of durum wheat. Aust. J. Agric. Res. 2000, 51, 69–74. [Google Scholar] [CrossRef]
- Mohammadi, M.; Mirlohi, A.; Majidi, M.M.; Rabbani, A. Exploring the breeding potential of Iranian emmer wheats to increase durum wheat tolerance to drought. Plant Genet. Resour. 2021, 19, 363–374. [Google Scholar] [CrossRef]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.V.; Giménez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef]
- Camerlengo, F.; Frittelli, A.; Sparks, C.; Doherty, A.; Martignago, D.; Larré, C.; Lupi, R.; Sestili, F.; Masci, S. CRISPR-Cas9 multiplex editing of the α-amylase/trypsin inhibitor genes to reduce allergen proteins in durum wheat. Front. Sustain. Food Syst. 2020, 4, 104. [Google Scholar] [CrossRef]
- Velde, K.V.D.; Thomas, S.G.; Heyse, F.; Kaspar, R.; Straeten, D.V.D.; Rohde, A. N-terminal truncated RHT-1 proteins generated by translational reinitiation cause semi-dwarfing of wheat Green Revolution alleles. Mol. Plant 2021, 14, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Mefleh, M.; Conte, P.; Fadda, C.; Giunta, F.; Piga, A.; Hassoun, G.; Motzo, R. From ancient to old and modern durum wheat varieties: Interaction among cultivar traits, management, and technological quality. J. Sci. Food Agric. 2019, 99, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.F. Puroindolines: The molecular genetic basis of wheat grain hardness. Plant Mol. Biol. 2002, 48, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Bhave, M.; Morris, C.F. Molecular genetics of puroindolines and related genes: Allelic diversity in wheat and other grasses. Plant Mol. Biol. 2008, 66, 205–219. [Google Scholar] [CrossRef]
- Li, W.; Huang, L.; Gill, B.S. Recurrent Deletions of Puroindoline Genes at the Grain Hardness Locus in Four Independent Lineages of Polyploid Wheat. Plant Physiol. 2007, 146, 200–212. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.F. Bread-baking quality and the effects of Glu-D1 gene introgressions in durum wheat (Triticum turgidum ssp. durum). Cereal Chem. 2021, 98, 1151–1158. [Google Scholar] [CrossRef]
- Colasuonno, P.; Marcotuli, I.; Blanco, A.; Maccaferri, M.; Condorelli, G.E.; Tuberosa, R.; Parada, R.; de Camargo, A.C.; Schwember, A.R.; Gadaleta, A. Carotenoid pigment content in durum wheat (Triticum turgidum L. var durum): An overview of quantitative trait loci and candidate genes. Front. Plant Sci. 2019, 10, 1347. [Google Scholar] [CrossRef] [Green Version]
- Patil, R.; Oak, M.; Deshpande, A.; Tamhankar, S. Development of a robust marker for Psy-1 homoeologs and its application in improvement of yellow pigment content in durum wheat. Mol. Breed. 2018, 38, 136. [Google Scholar] [CrossRef]
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgensen, L.N.; Hovmøller, M.S.; Huerta-Espino, J. Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef] [Green Version]
- David, B.G. Hessian Fly (Diptera: Cecidomyiidae) Injury and Loss of Winter Wheat Grain Yield and Quality. J. Econ. Entomol. 1999, 92, 1190–1197. [Google Scholar]
- Raman, H.; Ryan, P.R.; Raman, R.; Stodart, B.J.; Zhang, K.; Martin, P.; Wood, R.; Sasaki, T.; Yamamoto, Y.; Mackay, M. Analysis of TaALMT1 traces the transmission of aluminum resistance in cultivated common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2008, 116, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.-C.; Dvořák, J. Molecular mapping of an aluminum tolerance locus on chromosome 4D of Chinese Spring wheat. Euphytica 1996, 91, 31–35. [Google Scholar] [CrossRef]
- Cabanillas, B. Gluten-related disorders: Celiac disease, wheat allergy, and nonceliac gluten sensitivity. Crit. Rev. Food Sci. Nutr. 2020, 60, 2606–2621. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Bhatia, S.; Chunduri, V.; Kaur, S.; Sharma, S.; Kapoor, P.; Kumari, A.; Garg, M. Pathogenesis of celiac disease and other gluten related disorders in wheat and strategies for mitigating them. Front. Nutr. 2020, 7, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherf, K.A. Immunoreactive cereal proteins in wheat allergy, non-celiac gluten/wheat sensitivity (NCGS) and celiac disease. Curr. Opin. Food Sci. 2019, 25, 35–41. [Google Scholar] [CrossRef]
- Sabença, C.; Ribeiro, M.; Sousa, T.d.; Poeta, P.; Bagulho, A.S.; Igrejas, G. Wheat/Gluten-Related Disorders and Gluten-Free Diet Misconceptions: A Review. Foods 2021, 10, 1765. [Google Scholar] [CrossRef] [PubMed]
- García-Molina, M.D.; Giménez, M.J.; Sánchez-León, S.; Barro, F. Gluten free wheat: Are we there? Nutrients 2019, 11, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barro, F.; Iehisa, J.C.; Giménez, M.J.; García-Molina, M.D.; Ozuna, C.V.; Comino, I.; Sousa, C.; Gil-Humanes, J. Targeting of prolamins by RNAi in bread wheat: Effectiveness of seven silencing-fragment combinations for obtaining lines devoid of coeliac disease epitopes from highly immunogenic gliadins. Plant Biotechnol. J. 2016, 14, 986–996. [Google Scholar] [CrossRef] [Green Version]
- Gil-Humanes, J.; Pistón, F.; Tollefsen, S.; Sollid, L.M.; Barro, F. Effective shutdown in the expression of celiac disease-related wheat gliadin T-cell epitopes by RNA interference. Proc. Natl. Acad. Sci. USA 2010, 107, 17023–17028. [Google Scholar] [CrossRef] [Green Version]
- Henggeler, J.C.; Veríssimo, M.; Ramos, F. Non-coeliac gluten sensitivity: A review of the literature. Trends Food Sci. Technol. 2017, 66, 84–92. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Conti, A.; Pravettoni, V.; Bonomi, S.; Iametti, S.; Fortunato, D.; Scibilia, J.; Bindslev-Jensen, C.; Ballmer-Weber, B. Wheat IgE-mediated food allergy in European patients: α-amylase inhibitors, lipid transfer proteins and low-molecular-weight glutenins. Int. Arch. Allergy Immunol. 2007, 144, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Plamenov, D.; Belchev, I.; Spetsov, P. Anther culture response of Triticum durum × T. monococcum ssp. aegilopoides amphiploid. Cereal Res Commun. 2009, 37, 255–259. [Google Scholar] [CrossRef]
- Li, H.; Liu, X.; Zhang, M.; Feng, Z.; Zhang, L. Development and identification of new synthetic T. turgidum–T. monococcum amphiploids. Plant Genet. Resour. 2018, 16, 555–563. [Google Scholar] [CrossRef]
- Gul Kazi, A.; Rasheed, A.; Bux, H.; Napar, A.A.; Ali, A.; Mujeeb-Kazi, A. Cytological, Phenological and Molecular Characterization of B (S)-Genome Synthetic Hexaploids (2n = 6x = 42; AABBSS). Cereal Res. Commun. 2015, 43, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Rafique, K.; Rasheed, A.; Kazi, A.G.; Bux, H.; Naz, F.; Mahmood, T.; Mujeeb-Kazi, A. Powdery mildew resistance in some new wheat amphiploids (2n = 6x = 42) derived from A- and S-genome diploid progenitors. Plant Genet. Resour. 2012, 10, 165–170. [Google Scholar] [CrossRef]
- Ishii, H. A quantitative study of morphological relationships among natural tetraploid wheats and their putative ancestors. Sci Bull. Coll. Agric. Univ. Ryukyus 1972, 19, 75–85. [Google Scholar]
- Wang, K.; Shi, L.; Liang, X.; Zhao, P.; Wang, W.; Liu, J.; Chang, Y.; Hiei, Y.; Yanagihara, C.; Du, L.; et al. The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 2022. [Google Scholar] [CrossRef]
- Luo, J.; Li, S.; Xu, J.; Yan, L.; Ma, Y.; Xia, L. Pyramiding favorable alleles in an elite wheat variety in one generation by CRISPR-Cas9-mediated multiplex gene editing. Mol. Plant 2021, 14, 847–850. [Google Scholar] [CrossRef]


{kind=link}
| Ploidy | Categories | Botanical Name | Synonyms | Phenotype |
|---|---|---|---|---|
| Tetraploid 2n = 28 | T. timopheevi Zhuk. AuAuBspBsp/ AuAuGG | var. araraticum | T. araraticum Jakubz. | wild, brittle rachis, hulled |
| concv. timopheevi | T. timopheevi Zhuk. | domesticated, hulled | ||
| T. turgidum L. AuAuBB | var. dicoccoides | T. dicoccoides Koern. | wild, hulled, brittle rachis | |
| concv. dicoccon/dicoccum | T. dicoccon Schrank. | domesticated, hulled, semi-brittle rachis | ||
| concv. durum | T. durum Desf. | cultivated, free-threshing, tough rachis | ||
| concv. turgidum | T. turgidum L. | cultivated, free-threshing, tough rachis | ||
| concv. polonicum | T. polonicum L. | cultivated, free-threshing, tough rachis | ||
| concv. carthlicum | T. carthlicum Nevski. | cultivated, free-threshing, tough rachis |
| Trait | Genes/QTLs | Chromosome | Approaches | Origin | Variations | References | |
|---|---|---|---|---|---|---|---|
| Yield component | plant height | Rht1, Rht14, Rht15 | 4B, 4A, 6A | pentaploid hybridization, homologous hybridization | hexaploid wheat Norin10, durum wheat Castelporziano or Durox | reduced height; increased harvest index or pleiotropic effect | [145,146,147] |
| multiple traits | / | / | pentaploid hybridization | hexaploid wheat CSCR6 | pleiotropic effect | [148] | |
| / | 4D | pentaploid hybridization | Ae. tauschii accession AT23 | big spike, significantly increased number of spikelets and florets per spike; enhanced YR resistance | [149] | ||
| grain size | GW2-A1, GW2-B1 | 6AS, 6BS | RNA interference | durum wheat Svevo | increased kernel size | [150] | |
| Grain quality | soft kernel | Pina, Pinb | 5D | ph1c-mediated homoeologous recombination | Langdon 5D(5B) substitution line, durum wheat ph1c line Cappelli M | soft grain, vitreous kernels, high GPC, and good gluten quality | [151] |
| ph1b-mediated homoeologous recombination, homologous recombination | hexaploid wheat CS, Langdon-CS 5D(5B) substitution line | soft grain | [152] | ||||
| Pina, HMW glutenin subunit 1A × 1 | 1D, 1A | gene editing | durum wheat | soft kernel and better breadmaking quality | [153] | ||
| flour properties | Glu-D1 | 1D | ph1-mediated homoeologous recombination | multiple germplasms | improved bread-making quality | [28,154,155,156,157,158,159] | |
| grain protein content | Gpc-B1 | 6B | homologous recombination | Langdon-T. dicoccoides (6B) substitution line | increased protein level | [160] | |
| yellow pigment concentration | LCYe, HYD2 | Targeting Induced Local Lesions in Genomes (TILLING) | durum wheat | significantly increased β-carotene accumulation | [161] | ||
| pre-harvest sprouting | Qphs.sicau-3B.1 | 3B | pentaploid hybridization | hexaploid wheat T. spelta CSCR6 | high resistance to PHS | [162] | |
| Biotic resistance | leaf rust, powdery mildew, tan spot | Pm13, Lr19, YPC | 3B, 7A | ph1 mediated homoeologous recombination | Ae. Longissima, Agropyron | improved resistance to PM, LR, and YPC | [156,163,164] |
| multiple | / | inter/intra-specific hybridization | T. araraticum, T. dicoccoides, Ae. speltoides | enhanced resistance to LR and tan spot; increased grain yield and GPC | [165] | ||
| Lr34/Yr18/Sr57/Pm38/Ltn1 | / | gene editing | bread wheat | robust seedling resistance to LR, YR, and PM | [166] | ||
| stem rust | Sr22 | 7A | interspecific hybridization | T. monococcum L. cv. RL 5244 | differential resistance depends on ploidy level | [167] | |
| stripe rust | / | / | interspecific hybridization | A-genome diploids | enhanced resistance to YR, LR, and PM | [168,169,170] | |
| Yr28 | 4D | pentaploid hybridization | Ae. tauschii accession AT23 | enhanced resistance to YR | [149] | ||
| fusarium head blight | Fhb1 | 3BS, 4AL, 4BS, 5AL, 6AS | homologous recombination | hexaploid wheat Sumai-3 | improved resistance to FHB | [171] | |
| Qfhb.ndwp-5A, Qfhb.ndwp-7A | 5A,7A | homologous recombination | hexaploid wheat PI 277012 | [172] | |||
| / | 2AS, 2BS, 3AL, 4BL | pentaploid hybridization | hexaploid wheat Sumai-3 | [173] | |||
| / | / | pentaploid hybridization | hexaploid wheat Sumai-3 | [174] | |||
| / | / | mutation by treating with DNA methylation inhibitor (5-methyl-azacytidine) | / | [175] | |||
| Hessian fly | / | / | / | durum wheat resistant lines | enhanced resistance to Hessian fly, superior agro-phenological traits | [176] | |
| Abiotic resistance | thermotolerance | heat shock proteins | / | gene modify (overexpression) | / | improved thermotolerance | [177,178] |
| water | / | / | intraspecific hybridization | wild emmer | enhanced adaptation to water stress | [179] | |
| drought, salt | TdPIP2;1 | / | gene modify (overexpression) | durum wheat | enhanced drought and salt tolerance | [180] | |
| salt | / | / | induced by ascorbic acid | / | enhanced salt tolerance | [181] | |
| Kna1 locus, TaALMT1 gene | 4D | pentaploid hybridization, ph1-mediated homoeologous hybridization, homologous hybridization | bread wheat or Langdon-CS 4D (4B) substitution line | enhanced Al3+ tolerance | [182,183,184] | ||
| / | / | intraspecific hybridization | T. emmer, T. durum, T. carthlicum, T. turgidum, T. turanicum, T. polonicum | enhanced salt tolerance | [185,186] | ||
| wheat-related disorders | coeliac disease | α-gliadin genes | / | CRISPR/Cas9 (knock out) | durum wheat | highest 69% reduction of gliadin of reduced immunoreactivity | [187] |
| onset of bakers’ asthma | WTAI-CM3, WTAI-CM16 | / | CRISPR/Cas9 (knock out) | durum wheat | reduced allergen proteins | [188] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Zhang, J.; Liu, Q.; Liu, H.; Zhou, Y.; Yang, W.; Ma, W. Improvement and Re-Evolution of Tetraploid Wheat for Global Environmental Challenge and Diversity Consumption Demand. Int. J. Mol. Sci. 2022, 23, 2206. https://doi.org/10.3390/ijms23042206
Yang F, Zhang J, Liu Q, Liu H, Zhou Y, Yang W, Ma W. Improvement and Re-Evolution of Tetraploid Wheat for Global Environmental Challenge and Diversity Consumption Demand. International Journal of Molecular Sciences. 2022; 23(4):2206. https://doi.org/10.3390/ijms23042206
Chicago/Turabian StyleYang, Fan, Jingjuan Zhang, Qier Liu, Hang Liu, Yonghong Zhou, Wuyun Yang, and Wujun Ma. 2022. "Improvement and Re-Evolution of Tetraploid Wheat for Global Environmental Challenge and Diversity Consumption Demand" International Journal of Molecular Sciences 23, no. 4: 2206. https://doi.org/10.3390/ijms23042206