
Bis-Indole Alkaloids Isolated from the Sponge Spongosorites calcicola Disrupt Cell Membranes of MRSA


Abstract
:1. Introduction
2. Results
2.1. MIC and MBC of the Bis-Indoles against Target Pathogens
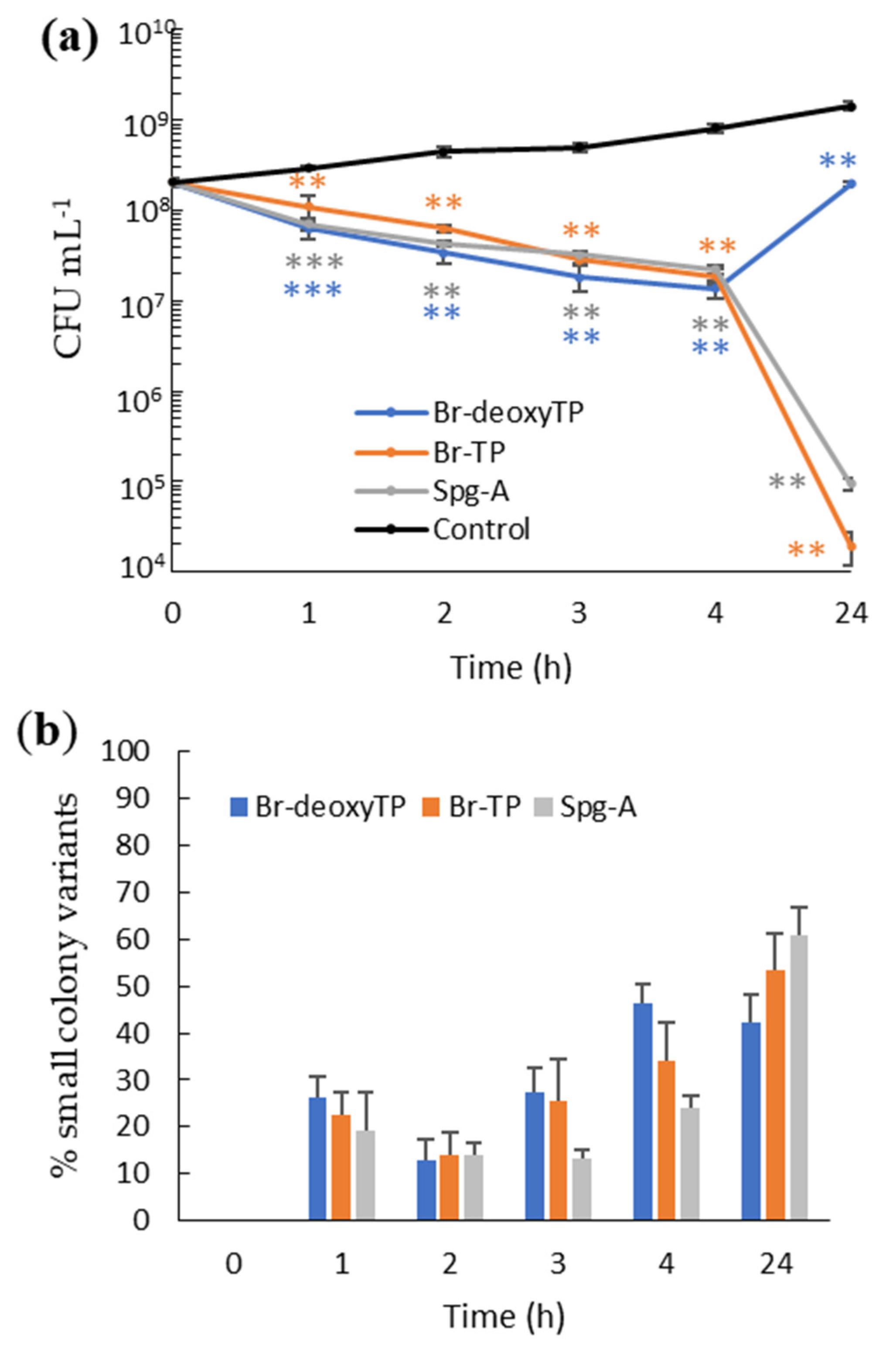
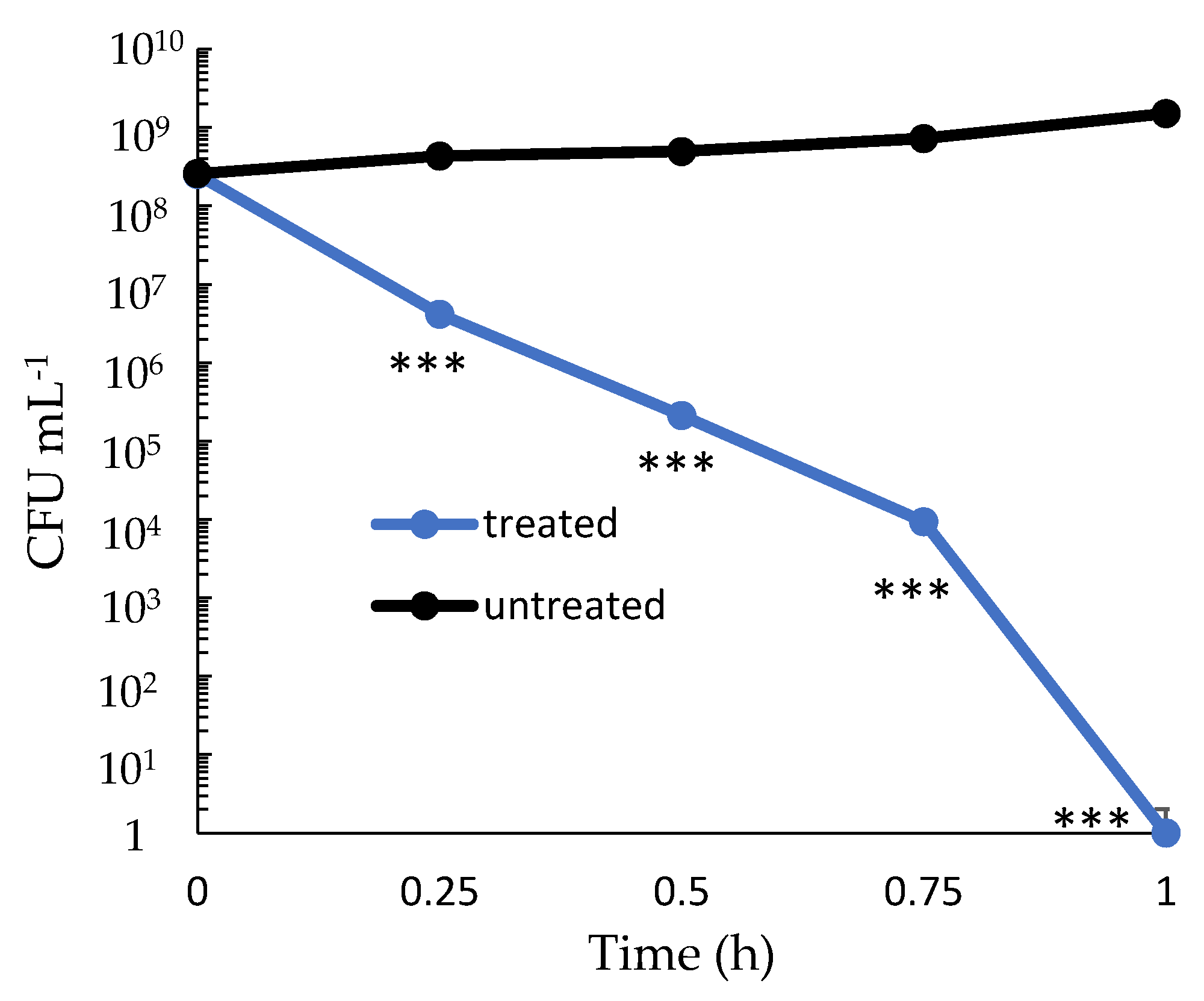
2.2. Dose-Dependency and Time-Kill Kinetics
2.3. Bis-Indoles Induce Transient Small Colony Variants in MRSA
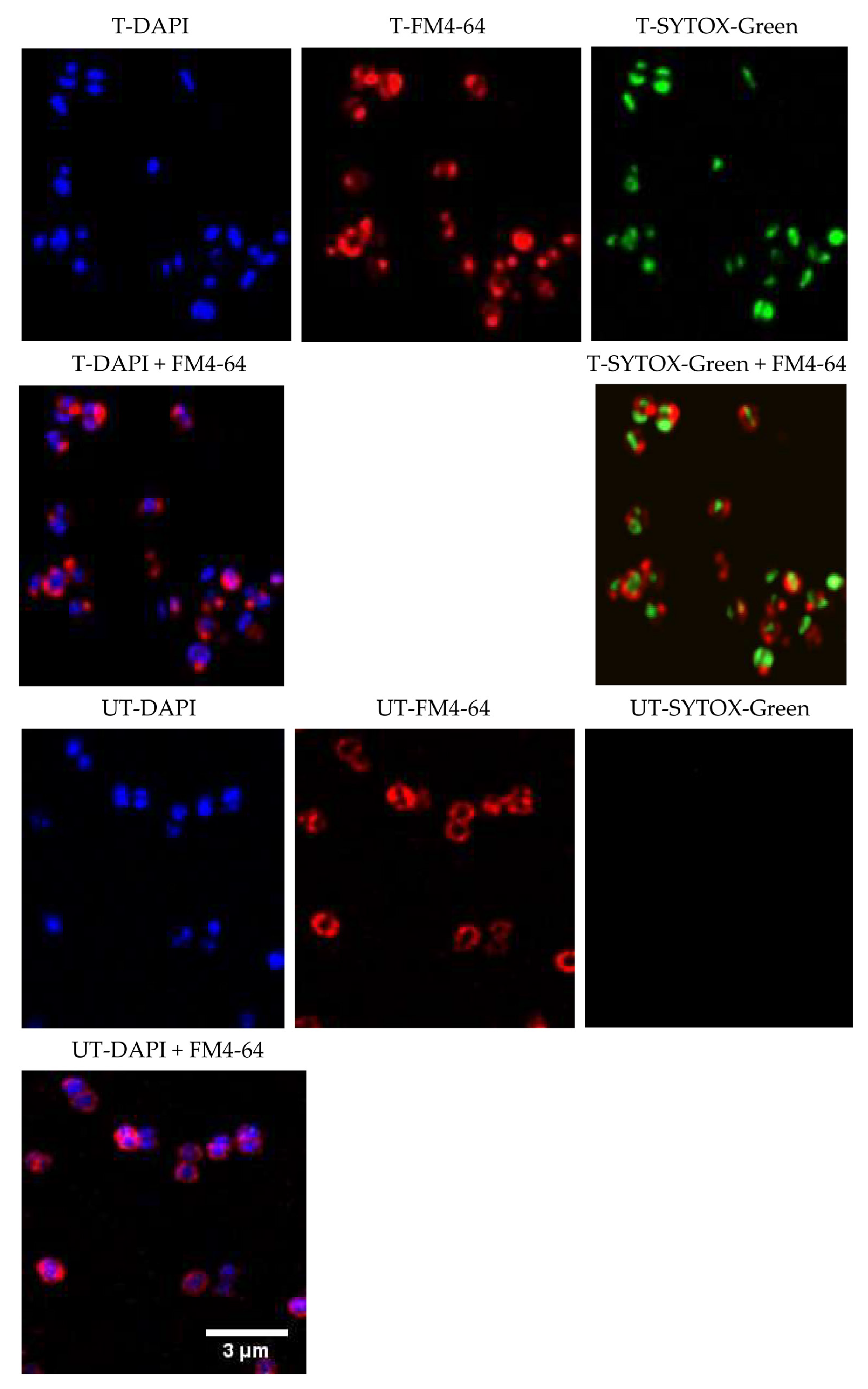
2.4. Mechanism of Action
2.5. Synergistic Effect of Bis-Indole Alkaloids with Aminoglycosides
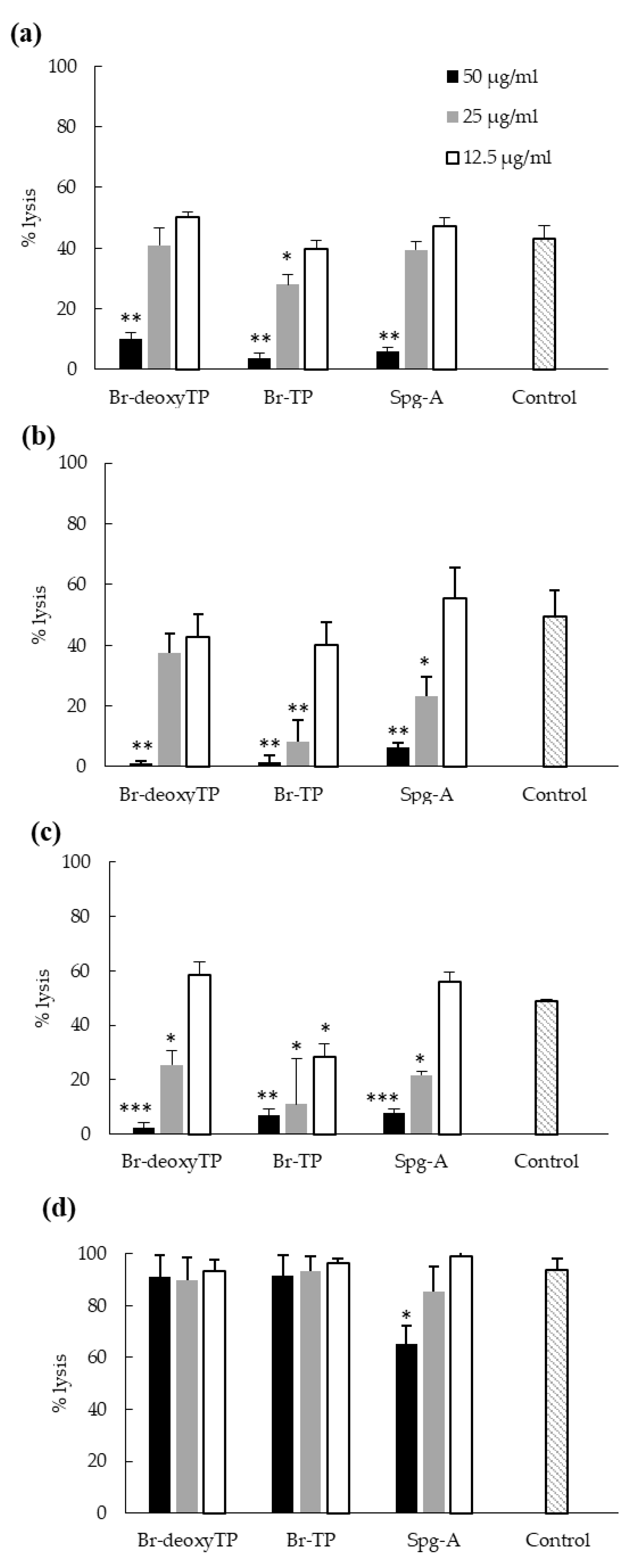
2.6. Inhibition of Bacterial Cytotoxicity toward HeLa Cells
3. Discussion
4. Materials and Methods
4.1. Reagents and Bioactive Molecules
4.2. Bacterial and Mammalian Culture
4.3. Determination of MIC and MBC
4.4. Time-Kill Assay
4.5. Synergistic Effect of the Bis-Indoles in Combination with Traditional Antibiotics
4.6. Cytological Profiling and Fluorescence Microscopy
4.7. Inhibition of Bacterial Cytotoxicity toward HeLa Cells
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 5 May 2021).
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Government of the United Kingdom: London, UK, 2016.
- Weiner-Lastinger, L.M.; Abner, S.; Edwards, J.R.; Kallen, A.J.; Karlsson, M.; Magill, S.S.; Pollock, D.; See, I.; Soe, M.M.; Walters, M.S.; et al. Antimicrobial-resistant pathogens associated with adult healthcare-associated infections: Summary of data reported to the National Healthcare Safety Network, 2015–2017. Infect. Control. Hosp. Epidemiol. 2019, 41, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourtis, A.P.; Hatfield, K.; Baggs, J.; Mu, Y.; See, I.; Epson, E.; Nadle, J.; Kainer, M.A.; Dumyati, G.; Petit, S.; et al. Vital signs:epidemiology and recent trends in methicillin-resistant and in methicillin-susceptible Staphylococcus aureus bloodstream infections—United States. Morb. Mortal. Wkly. Rep. 2019, 68, 214–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papkou, A.; Hedge, J.; Kapel, N.; Young, B.; MacLean, R.C. Efflux pump activity potentiates the evolution of antibiotic resistance across S. aureus isolates. Nat. Commun. 2020, 11, 3970. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic resistance and the MRSA problem. Microbiol. Spectr. 2019, 7, 18. [Google Scholar] [CrossRef]
- Watkins, R.R.; Holubar, M.; David, M.Z. Antimicrobial resistance in methicillin-resistant Staphylococcus aureus to newer antimicrobial agents. Antimicrob. Agents Chemother. 2019, 63, e01216–e01219. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Dai, J.; Han, R.; Xu, Y.; Li, N.; Wang, J.; Dan, W. Recent progress of antibacterial natural products: Future antibiotic candidates. Bioorganic Chem. 2020, 101, 103922. [Google Scholar] [CrossRef]
- Motelica, L.; Ficai, D.; Oprea, O.; Ficai, A.; Trusca, R.-D.; Andronescu, E.; Holban, A. Biodegradable alginate films with ZnO nanoparticles and Citronella essential oil—A novel antimicrobial structure. Pharmaceutics 2021, 13, 1020. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Hussein, D.E.; Algammal, A.M.; George, T.T.; Jeandet, P.; Al-Snafi, A.E.; Tiwari, A.; Pagnossa, J.P.; Lima, C.M.; Thorat, N.D.; et al. Application of natural antimicrobials in food preservation: Recent views. Food Control 2021, 126, 108066. [Google Scholar] [CrossRef]
- Han, B.-N.; Hong, L.-L.; Gu, B.-B.; Sun, Y.-T.; Wang, J.; Liu, J.-T.; Lin, H.-W. Natural products from sponges. In Symbiotic Microbiomes of Coral Reefs Sponges and Corals; Li, Z., Ed.; Springer: Berlin, Germany, 2019; pp. 329–463. [Google Scholar]
- Varijakzhan, D.; Loh, J.-Y.; Yap, W.-S.; Yusoff, K.; Seboussi, R.; Lim, S.-H.; Lai, K.-S.; Chong, C.-M. Bioactive compounds from marine sponges: Fundamentals and applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Keyzers, R.A.; Davies-Coleman, M.T. Anti-inflammatory metabolites from marine sponges. Chem. Soc. Rev. 2005, 34, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.-B.; Mar, W.; Kim, S.; Kim, J.-Y.; Lee, T.-H.; Kim, J.-G.; Shin, D.; Sim, C.J.; Shin, J. Antimicrobial activity and cytotoxicity of bis(indole) alkaloids from the sponge Spongosorites sp. Biol. Pharm. Bull. 2006, 29, 570–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-G.; Huang, H.; Jiang, B. Progress in studies of novel marine bis(indole) alkaloids. Curr. Org. Chem. 2004, 8, 1691–1720. [Google Scholar] [CrossRef]
- Hwang, J.; Kim, D.; Park, J.S.; Park, H.J.; Shin, J.; Lee, S.K. Park Photoprotective activity of topsentin, A bis(Indole) alkaloid from the marine sponge Spongosorites genitrix, by regulation of COX-2 and Mir-4485 expression in UVB-irradiated human keratinocyte cells. Mar. Drugs 2020, 18, 87. [Google Scholar] [CrossRef] [Green Version]
- Zoraghi, R.; Worrall, L.; See, R.H.; Strangman, W.; Popplewell, W.; Gong, H.; Samaai, T.; Swayze, R.D.; Kaur, S.; Vuckovic, M.; et al. Methicillin-resistant Staphylococcus aureus (MRSA) pyruvate kinase as a target for bis-indole alkaloids with antibacterial activities*. J. Biol. Chem. 2011, 286, 44716–44725. [Google Scholar] [CrossRef] [Green Version]
- Jennings, L.K.; Khan, N.M.D.; Kaur, N.; Rodrigues, D.; Morrow, C.; Boyd, A.; Thomas, O.P. Brominated bisindole alkaloids from the celtic sea sponge Spongosorites calcicola. Molecules 2019, 24, 3890. [Google Scholar] [CrossRef] [Green Version]
- Opperman, T.J.; Kwasny, S.M.; Li, J.B.; Lewis, M.A.; Aiello, D.; Williams, J.D.; Peet, N.P.; Moir, D.T.; Bowlin, T.L.; Long, E.C. DNA targeting as a likely mechanism underlying the antibacterial activity of synthetic bis-indole antibiotics. Antimicrob. Agents Chemother. 2016, 60, 7067–7076. [Google Scholar] [CrossRef] [Green Version]
- Rehberg, N.; Sommer, G.A.; Drießen, D.; Kruppa, M.; Adeniyi, E.T.; Chen, S.; Wang, L.; Wolf, K.; Tasch, B.O.A.; Ioerger, T.R.; et al. Nature-inspired (di)azine-bridged bisindole alkaloids with potent antibacterial in vitro and in vivo efficacy against methicillin-resistant Staphylococcus aureus. J. Med. Chem. 2020, 63, 12623–12641. [Google Scholar] [CrossRef]
- Melter, O.; Radojevič, B. Small colony variants of Staphylococcus aureus—review. Folia Microbiol. 2010, 55, 548–558. [Google Scholar] [CrossRef]
- McNamara, P.J.; Proctor, R.A. Staphylococcus aureus small colony variants, electron transport and persistent infections. Int. J. Antimicrob. Agents 2000, 14, 117–122. [Google Scholar] [CrossRef]
- Nonejuie, P.; Burkart, M.; Pogliano, K.; Pogliano, J. Bacterial cytological profiling rapidly identifies the cellular pathways targeted by antibacterial molecules. Proc. Natl. Acad. Sci. USA 2013, 110, 16169–16174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghooi, R.; Thatte, S.; Ghooi, R.; Thatte, S. Inhibition of cell wall synthesis—is this the mechanism of action of penicillins? Med. Hypotheses 1995, 44, 127–131. [Google Scholar] [CrossRef]
- Malanovic, N.; Lohner, K. Gram-positive bacterial cell envelopes: The impact on the activity of antimicrobial peptides. Biochim. et Biophys. Acta (BBA)-Biomembr. 2015, 1858, 936–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, J.-W.; Hung, Y.-J.; Yang, C.-H.; Chen, Y.-C. The antimicrobial activity of gramicidin A is associated with hydroxyl radical formation. PLoS ONE 2015, 10, e0117065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartash, R.; Nori, P. Beta-lactam combination therapy for the treatment of Staphylococcus aureus and Enterococcus species bacteremia: A summary and appraisal of the evidence. Int. J. Infect. Dis. 2017, 63, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Caspar, Y.; Jeanty, M.; Blu, J.; Burchak, O.; Le Pihive, E.; Maigre, L.; Schneider, M.; Jolivalt, C.; Paris, J.-M.; Hequet, A.; et al. Novel synthetic bis-indolic derivatives with antistaphylococcal activity, including against MRSA and VISA strains. J. Antimicrob. Chemother. 2015, 70, 1727–1737. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.G.; Sanders, C.C. Selection and characterization of strains of Staphylococcus aureus displaying unusual resistance to aminoglycosides. Antimicrob. Agents Chemother. 1976, 10, 519–525. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.A.; Peters, G. Small Colony Variants in staphylococcal infections: Diagnostic and therapeutic implications. Clin. Infect. Dis. 1998, 27, 419–422. [Google Scholar] [CrossRef] [Green Version]
- Gross, M.E.; Giron, K.P.; Septimus, J.D.; Mason, E.O.; Musher, D.M. Antimicrobial activities of beta-lactam antibiotics and gentamicin against penicillin-susceptible and penicillin-resistant pneumococci. Antimicrob. Agents Chemother. 1995, 39, 1166–1168. [Google Scholar] [CrossRef] [Green Version]
- Masuda, G.; Nakamura, K.; Yajima, T.; Saku, K. Bacteriostatic and bactericidal activities of beta-lactam antibiotics enhanced by the addition of low concentrations of gentamicin. Antimicrob. Agents Chemother. 1980, 17, 334–336. [Google Scholar] [CrossRef] [Green Version]
- Frimodt-Møller, N.; Thomsen, V.F. Interaction between beta-lactam antibiotics and gentamicin against Streptococcus pneumoniae in vitro and in vivo. Acta Pathol. Microbiol. Scand. Ser. B Microbiol. 2009, 95, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-H.; Chen, W.-L. Host lipid rafts as the gates for Listeria monocytogenes infection: A Mini-Review. Front. Immunol. 2020, 11, 1666. [Google Scholar] [CrossRef] [PubMed]
- Missiakas, D.; Winstel, V. Selective host cell death by Staphylococcus aureus: A strategy for bacterial persistence. Front. Immunol. 2021, 11, 621733. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Meng, H.; Gu, D.; Li, Y.; Jia, M. Molecular mechanisms of Vibrio parahaemolyticus pathogenesis. Microbiol. Res. 2019, 222, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta 2016, 1858, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Ma, S. Recent development of membrane-active molecules as antibacterial agents. Eur. J. Med. Chem. 2019, 184, 111743. [Google Scholar] [CrossRef]
- O’Neill, E.; Pozzi, C.; Houston, P.; Smyth, D.; Humphreys, H.; Robinson, D.A.; O’Gara, J.P. Association between methicillin susceptibility and biofilm regulation in Staphylococcus aureus isolates from device-related infections. J. Clin. Microbiol. 2007, 45, 1379–1388. [Google Scholar] [CrossRef] [Green Version]
- McNamara, P.J. Genetic manipulation of Staphylococcus aureus. In Molecular Genetics; Lindsay, J., Ed.; Staphylococcus: Norfolk, UK, 2008; pp. 89–130. [Google Scholar]
- Glaser, P.; Frangeul, L.; Buchrieser, C.; Rusniok, C.; Amend, A.; Baquero, F.; Berche, P.; Bloecker, H.; Brandt, P.; Chakraborty, T.; et al. Comparative genomics of Listeria species. Science 2001, 294, 849–852. [Google Scholar] [CrossRef] [Green Version]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of V. cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- Clinical Lab Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, Approved Standard M09. 2018. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 13 January 2022).
- Garcia, L.S. Clinical Microbiology Procedures Handbook; American Society for Microbiology Press: Washington, DC, USA, 2010. [Google Scholar]
- Levison, M.E. Pharmacodynamics of antimicrobial drugs. Infect. Dis. Clin. N. Am. 2004, 18, 451–465. [Google Scholar] [CrossRef]
- Sopirala, M.M.; Mangino, J.E.; Gebreyes, W.A.; Biller, B.; Bannerman, T.; Balada-Llasat, J.-M.; Pancholi, P. Synergy Testing by Etest, Microdilution checkerboard, and time-kill methods for pan-drug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4678–4683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.-T.; Yang, Y.-L.; Xu, Y.; Lamsa, A.; Haste, N.M.; Yang, J.Y.; Ng, J.; Gonzalez, D.; Ellermeier, C.; Straight, P.; et al. Imaging mass spectrometry of intraspecies metabolic exchange revealed the cannibalistic factors of Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2010, 107, 16286–16290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matlawska-Wasowska, K.; Finn, R.; Mustel, A.; O’Byrne, C.P.; Baird, A.W.; Coffey, E.T.; Boyd, A. The Vibrio parahaemolyticus Type III Secretion Systems manipulate host cell MAPK for critical steps in pathogenesis. BMC Microbiol. 2010, 10, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gram-Positive Bacteria | Gram-Negative Bacteria | ||||||
|---|---|---|---|---|---|---|---|
| MRSA | MSSA | LM a | VP a | ST a | EC a | ||
| Br-deoxy TP | MIC b | 25 | 25 | >50 | >50 | >50 | >50 |
| MBC b | >50 | >50 | nd c | nd | nd | nd | |
| Br-TP | MIC | 12.5 | 6.25 | 12.5 | 25 | >50 | >50 |
| MBC | >50 | 25 | >50 | 25 | nd | nd | |
| Spg-A | MIC | 12.5 | 12.5 | 25 | 25 | >50 | >50 |
| MBC | 50 | 25 | 25 | 50 | nd | nd | |
| Bacteria | MICI a (mg L−1) | MICC b (mg L−1) | FICI c | |
|---|---|---|---|---|
| Antibiotic | Bis-Indole | Antibiotic + Bis-Indole | ||
| MSSA | ||||
| Gentamicin (0.25) | Br-deoxyTP (25) | Gentamicin + Br-deoxyTP (0.03 + 3.12) | 0.24 | |
| Gentamicin (0.25) | Br-TP (6.25) | Gentamicin + Br-TP (0.06 + 1.56) | 0.48 | |
| V. parahaemolyticus | ||||
| Gentamicin (25) | Br-TP (25) | Gentamicin + Br-TP (6.25 + 1.5) | 0.31 | |
| Gentamicin (25) | Spg-A (25) | Gentamicin + Spg-A (6.25 + 3.12) | 0.37 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, N.A.; Kaur, N.; Owens, P.; Thomas, O.P.; Boyd, A. Bis-Indole Alkaloids Isolated from the Sponge Spongosorites calcicola Disrupt Cell Membranes of MRSA. Int. J. Mol. Sci. 2022, 23, 1991. https://doi.org/10.3390/ijms23041991
Khan NA, Kaur N, Owens P, Thomas OP, Boyd A. Bis-Indole Alkaloids Isolated from the Sponge Spongosorites calcicola Disrupt Cell Membranes of MRSA. International Journal of Molecular Sciences. 2022; 23(4):1991. https://doi.org/10.3390/ijms23041991
Chicago/Turabian StyleKhan, Neyaz A., Navdeep Kaur, Peter Owens, Olivier P. Thomas, and Aoife Boyd. 2022. "Bis-Indole Alkaloids Isolated from the Sponge Spongosorites calcicola Disrupt Cell Membranes of MRSA" International Journal of Molecular Sciences 23, no. 4: 1991. https://doi.org/10.3390/ijms23041991