
Root Membrane Ubiquitinome under Short-Term Osmotic Stress


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Differentially Accumulated Proteins in Response to Mannitol Treatment
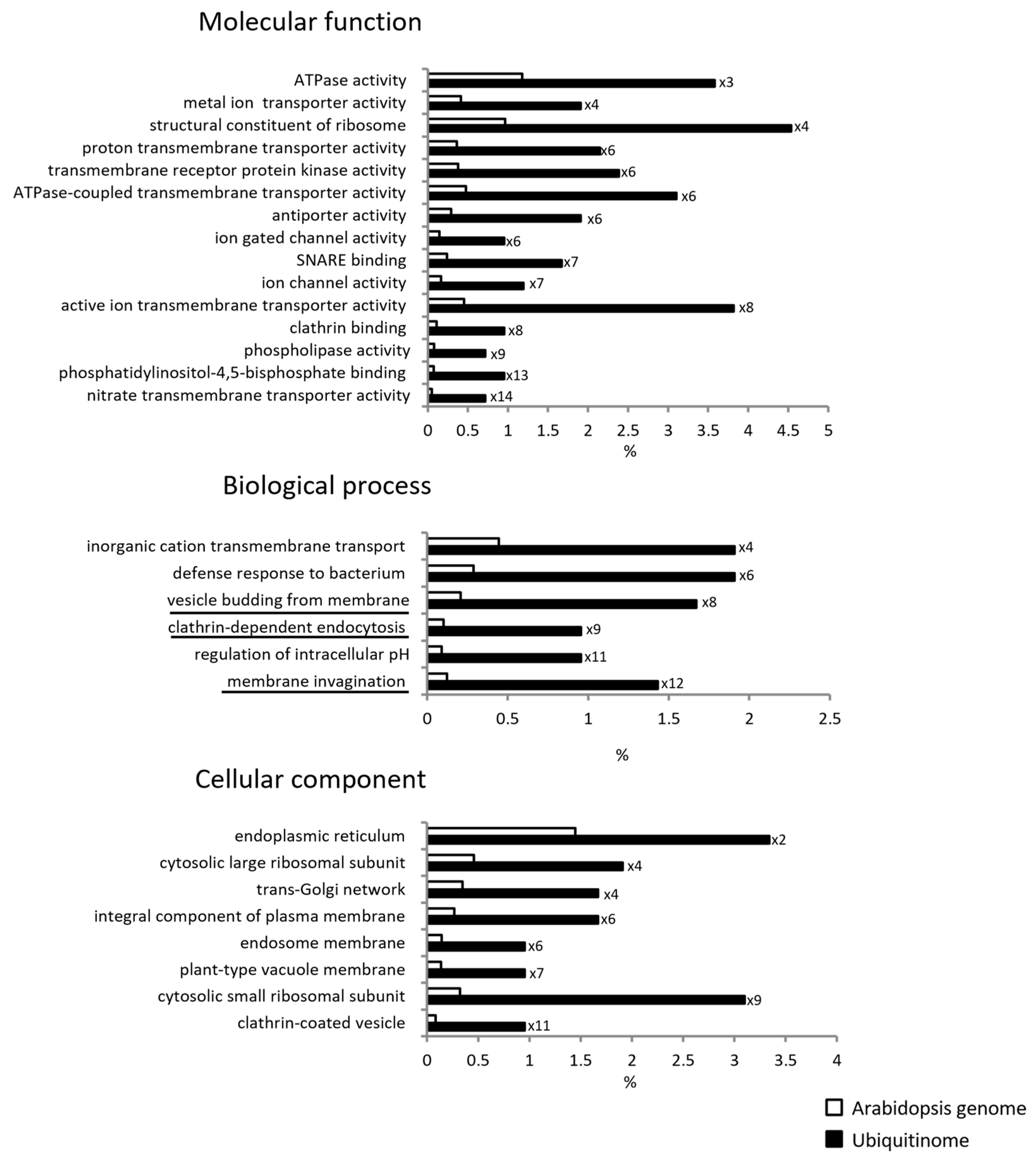
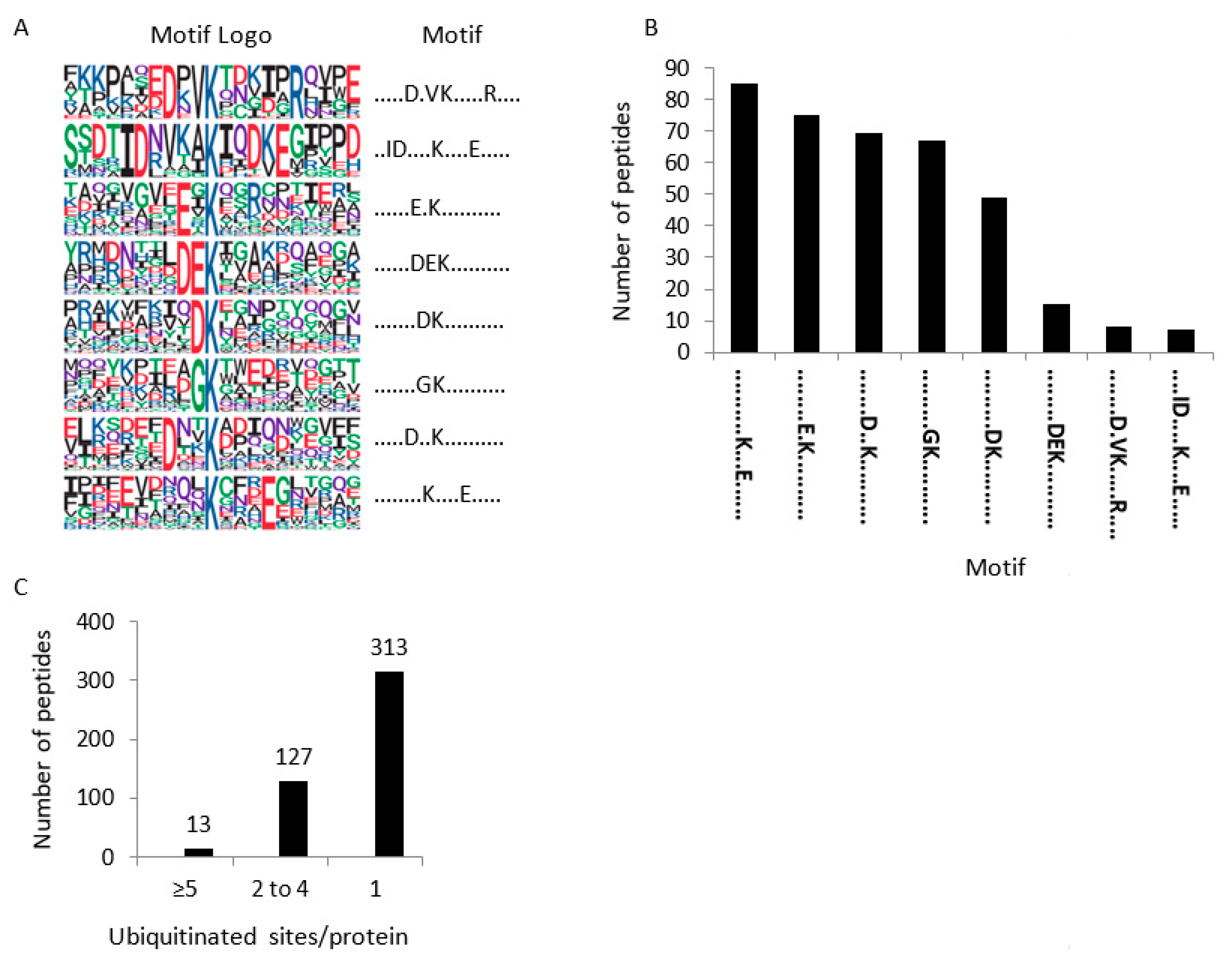
2.2. Characterization of the Root Membrane Ubiquitinome
2.3. Differentially Accumulated Ubiquitinated Proteins in Response to Mannitol Treatment
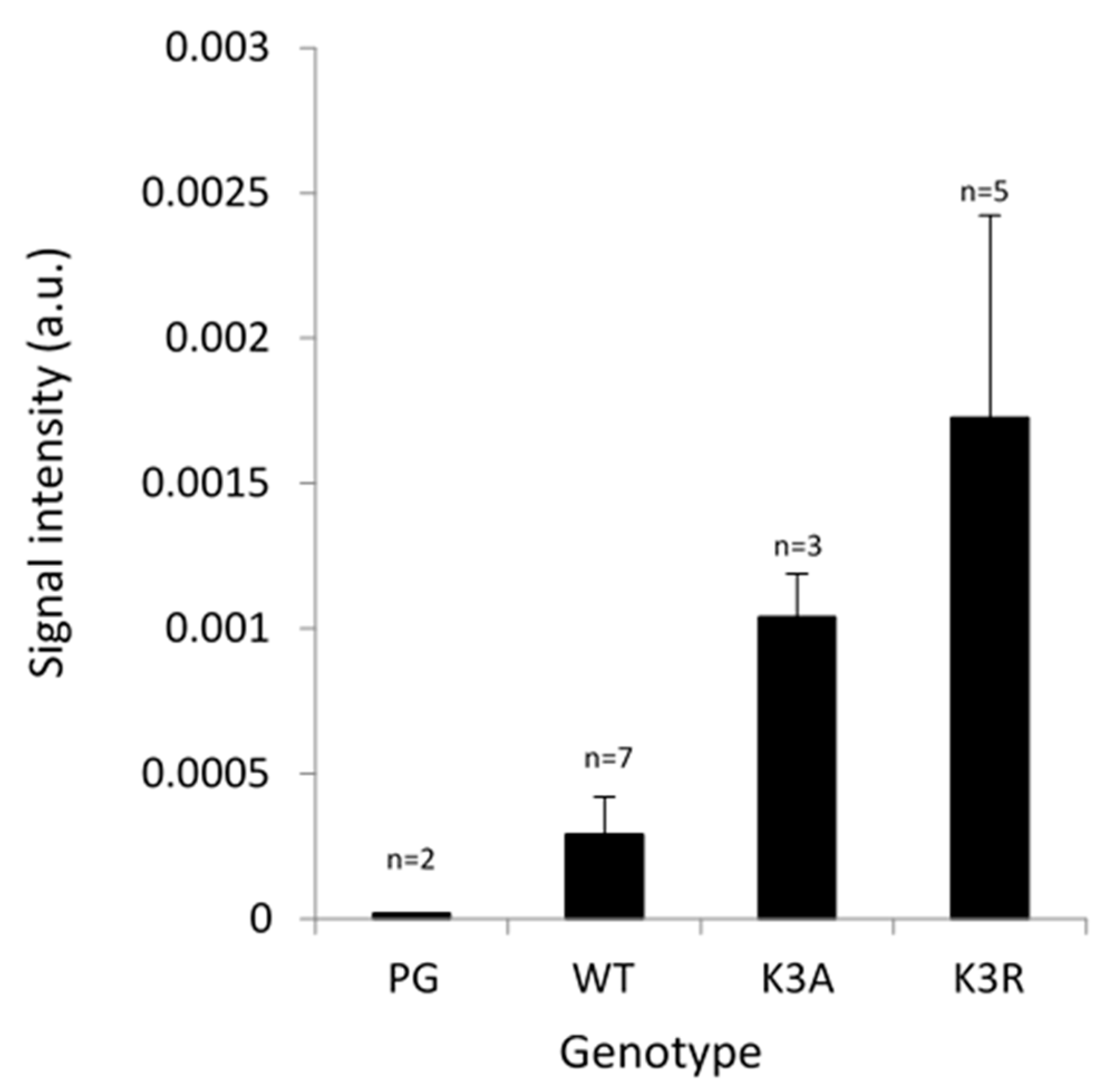
2.4. Ubiquitination of PIP Aquaporins
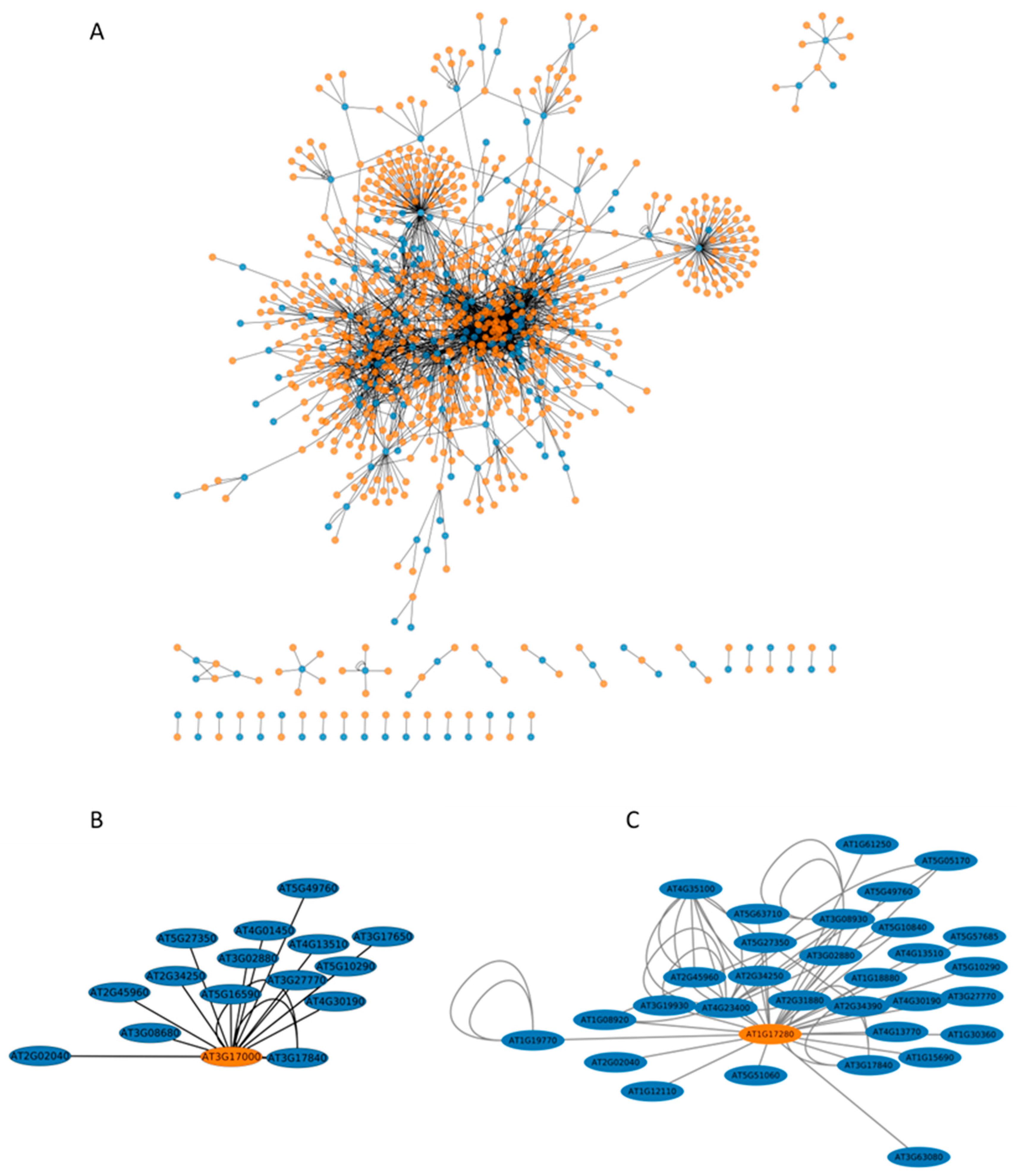
2.5. The Interactome of Ubiquitinated Proteins
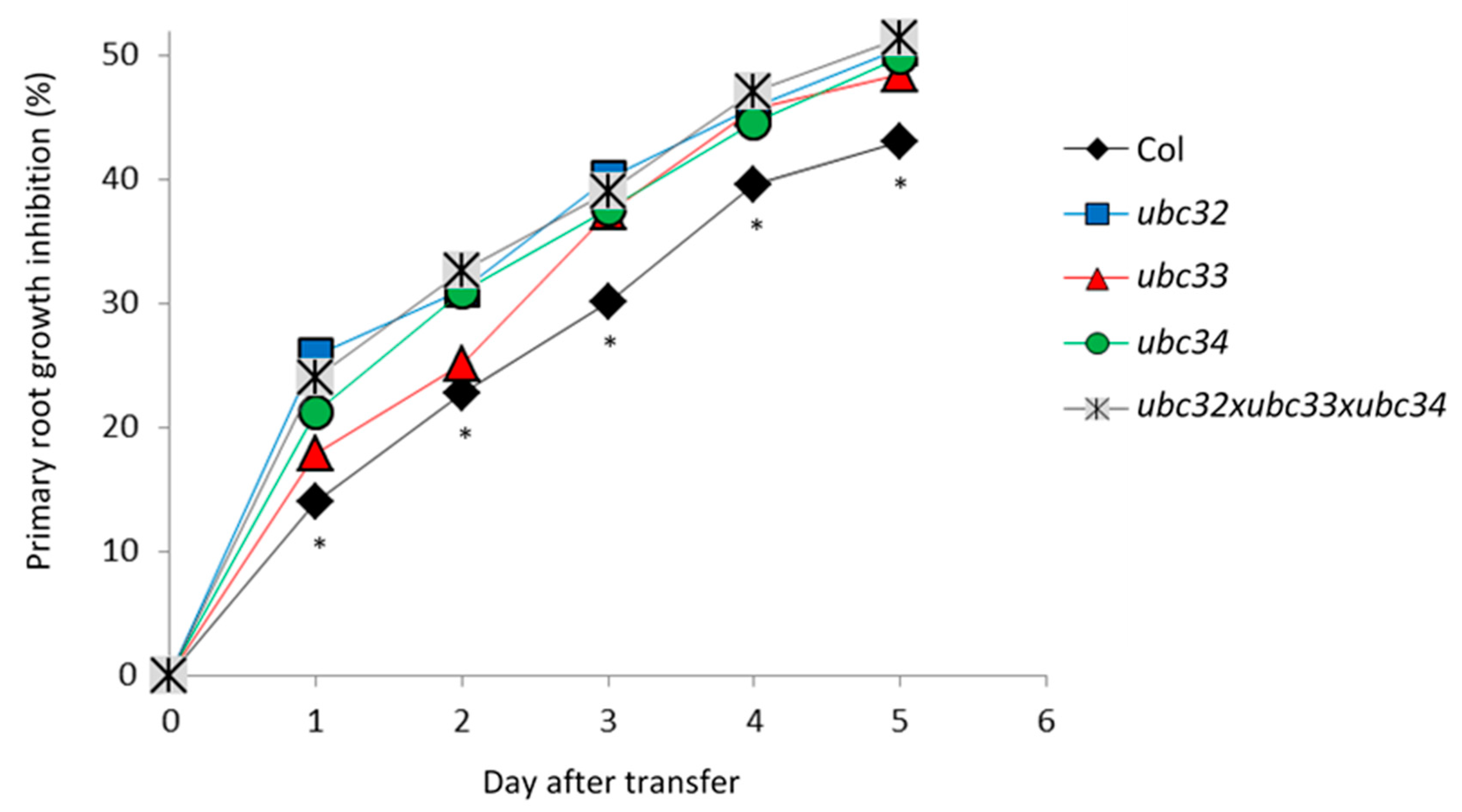
2.6. The Osmotic Phenotype of Ubc32, Ubc33 and Ubc34 Mutants
3. Discussion
3.1. A Resource of Ubiquitinated Membrane Proteins
3.2. The Role of Ubiquitination in Response to Short-Term Osmotic Treatment
3.3. UBC32 and UBC34 Contribute to Primary Root Growth under Osmotic Stress
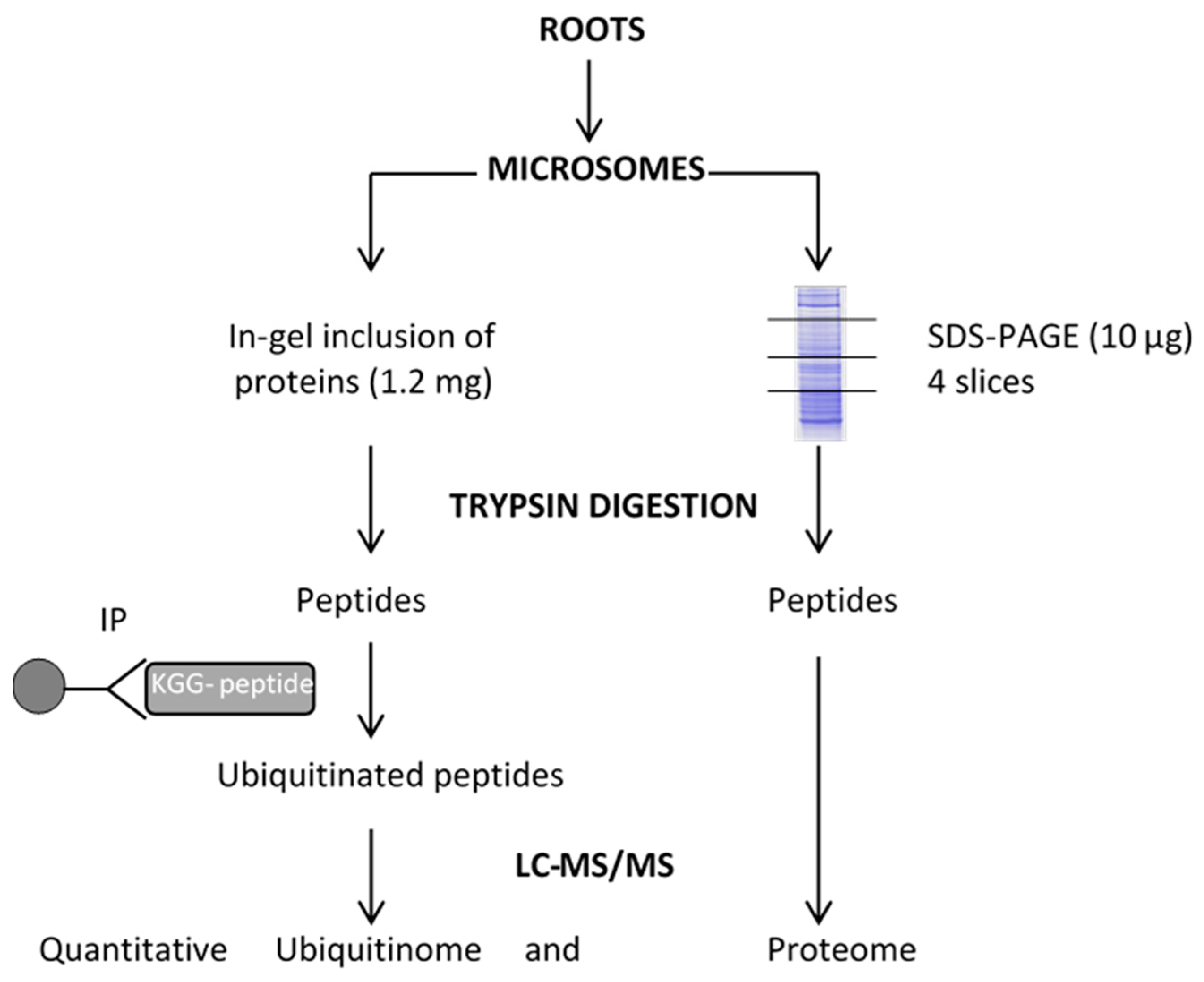
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Microsome Extraction
4.3. Protein Digestion
4.4. Enrichment of Ubiquitinated Peptides
4.5. LC-MS/MS Analysis
4.6. Identification and Quantification of Whole Proteome and Ubiquitinome
4.7. Bioinformatic Analyses
4.8. Ectopic Expression of PIP2;1 in Suspension Cells
4.9. Root Architecture Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ray, D.K.; Gerber, J.S.; MacDonald, G.K.; West, P.C. Climate variation explains a third of global crop yield variability. Nat. Commun. 2015, 6, 5989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, D.; Riezman, H. Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science 2007, 315, 201–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, C.J.; Millar, A.H. Protein turnover in plant biology. Nat. Plants 2015, 1, 15017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callis, J. The Ubiquitination Machinery of the Ubiquitin System. Arab. Book 2014, 12, e0174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, C.K.; Sadanandom, A. Ubiquitin chain topology in plant cell signaling: A new facet to an evergreen story. Front. Plant Sci. 2014, 5, 122. [Google Scholar] [CrossRef] [Green Version]
- Pickart, C.M.; Eddins, M.J. Ubiquitin: Structures, functions, mechanisms. Biochim. Biophys. Acta-Mol. Cell Res. 2004, 1695, 55–72. [Google Scholar] [CrossRef] [Green Version]
- Komander, D.; Rape, M. The Ubiquitin Code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Romero-Barrios, N.; Monachello, D.; Dolde, U.; Wong, A.; San Clemente, H.; Cayrel, A.; Johnson, A.; Lurin, C.; Vert, G. Advanced Cataloging of Lysine-63 Polyubiquitin Networks by Genomic, Interactome, and Sensor-Based Proteomic Analyses. Plant Cell 2020, 32, 123–138. [Google Scholar] [CrossRef]
- Stone, S.L. Role of the Ubiquitin Proteasome System in Plant Response to Abiotic Stress. In International Review of Cell and Molecular Biology; Galluzzi, L., Ed.; Academic Press Ltd.: Cambridge, MA, USA; Elsevier Science Ltd.: London, UK, 2018; Volume 343, pp. 65–110. [Google Scholar]
- Turek, I.; Tischer, N.; Lassig, R.; Trujillo, M. Multi-tiered pairing selectivity between E2 ubiquitin-conjugating enzymes and E3 ligases. J. Biol. Chem. 2018, 293, 16324–16336. [Google Scholar] [CrossRef] [Green Version]
- Grubb, L.E.; Derbyshire, P.; Dunning, K.E.; Zipfel, C.; Menke, F.L.H.; Monaghan, J. Large-scale identification of ubiquitination sites on membrane-associated proteins in Arabidopsis thaliana seedlings. Plant Physiol. 2021, 185, 1483–1488. [Google Scholar] [CrossRef]
- Guo, J.H.; Liu, J.X.; Wei, Q.; Wang, R.M.; Yang, W.Y.; Ma, Y.Y.; Chen, G.J.; Yu, Y.X. Proteomes and Ubiquitylomes Analysis Reveals the Involvement of Ubiquitination in Protein Degradation in Petunias. Plant Physiol. 2017, 173, 668–687. [Google Scholar] [CrossRef] [PubMed]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma Membrane H+-ATPase Regulation in the Center of Plant Physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Shea, J.P.; Chou, M.F.; Quader, S.A.; Ryan, J.K.; Church, G.M.; Schwartz, D. pLogo: A probabilistic approach to visualizing sequence motifs. Nat. Methods 2013, 10, 1211. [Google Scholar] [CrossRef]
- Braun, P.; Carvunis, A.R.; Charloteaux, B.; Dreze, M.; Ecker, J.R.; Hill, D.E.; Roth, F.P.; Vidal, M.; Galli, M.; Balumuri, P.; et al. Evidence for network evolution in an Arabidopsis Interactome map. Science 2011, 333, 601–607. [Google Scholar]
- Jones, A.M.; Xuan, Y.H.; Xu, M.; Wang, R.S.; Ho, C.H.; Lalonde, S.; You, C.H.; Sardi, M.I.; Parsa, S.A.; Smith-Valle, E.; et al. Border control–A membrane-linked interactome of Arabidopsis. Sci. 2014, 344, 711–716. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, R.; Wu, Y.; Wei, S.; Wang, Q.; Zheng, Y.; Ran, X.; Shang, X.; Yu, F.; Yang, X.; et al. ERAD-related E2 and E3 enzymes modulate the drought response by regulating the stability of PIP2 aquaporins. Plant Cell 2021, 33, 2883–2898. [Google Scholar] [CrossRef]
- Maurel, C.; Boursiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Santoni, V.; Verdoucq, L.; Sommerer, N.; Vinh, J.; Pflieger, D.; Maurel, C. Methylation of aquaporins in plant plasma membrane. Biochem. J. 2006, 400, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lalonde, S.; Obrdlik, P.; Vatani, A.N.; Parsa, S.A.; Vilarino, C.; Revuelta, J.L.; Frommer, W.B.; Rhee, S.Y. Uncovering Arabidopsis membrane protein interactome enriched in transporters using mating-based split ubiquitin assays and classification models. Frontiers Plant Sci. 2012, 3, 124. [Google Scholar] [CrossRef] [Green Version]
- Ahn, M.Y.; Oh, T.R.; Seo, D.H.; Kim, J.H.; Cho, N.H.; Kim, W.T. Arabidopsis group XIV ubiquitin-conjugating enzymes AtUBC32, AtUBC33, and AtUBC34 play negative roles in drought stress response. J. Plant Physiol. 2018, 230, 73–79. [Google Scholar] [CrossRef]
- Kim, D.Y.; Scalf, M.; Smith, L.M.; Vierstra, R.D. Advanced proteomic analyses yield a deep catalog of ubiquitylation targets in Arabidopsis. Plant Cell 2013, 25, 1523–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzano, C.; Abraham, Z.; Lopez-Torrejon, G.; Del Pozo, J.C. Identification of ubiquitinated proteins in Arabidopsis. Plant Mol. Biol. 2008, 68, 145–158. [Google Scholar] [CrossRef]
- Maor, R.; Jones, A.; Nuhse, T.S.; Studholme, D.J.; Peck, S.C.; Shirasu, K. Multidimensional protein identification technology (MudPIT) analysis of ubiquitinated proteins in plants. Mol. Cell. Proteom. 2007, 6, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svozil, J.; Hirsch-Hoffmann, M.; Dudler, R.; Gruissem, W.; Baerenfaller, K. Protein Abundance Changes and Ubiquitylation Targets Identified after Inhibition of the Proteasome with Syringolin A. Mol. Cell. Proteom. 2014, 13, 1523–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, A.; Stes, E.; Cybulski, N.; Van Bel, M.; Inigo, S.; Durand, A.N.; Timmerman, E.; Heyman, J.; Pauwels, L.; De Veylder, L.; et al. It’s Time for Some “Site”-Seeing: Novel Tools to Monitor the Ubiquitin Landscape in Arabidopsis thaliana. Plant Cell 2016, 28, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Xu, J.; Liu, X.Y.; Liang, W.X.; Xin, M.M.; Du, J.K.; Hu, Z.R.; Peng, H.R.; Guo, W.L.; Ni, Z.F.; et al. Identification of HSP90C as a substrate of E3 ligase TaSAP5 through ubiquitylome profiling. Plant Sci. 2019, 287, 110170. [Google Scholar] [CrossRef]
- Fuglsang, A.T.; Kristensen, A.; Cuin, T.A.; Schulze, W.X.; Persson, J.; Thuesen, K.H.; Ytting, C.K.; Oehlenschlaeger, C.B.; Mahmood, K.; Sondergaard, T.E.; et al. Receptor kinase-mediated control of primary active proton pumping at the plasma membrane. Plant J. 2014, 80, 951–964. [Google Scholar] [CrossRef]
- Vu, L.D.; Gevaert, K.; De Smet, I. Protein Language: Post-Translational Modifications Talking to Each Other. Trends Plant Sci. 2018, 23, 1068–1080. [Google Scholar] [CrossRef]
- He, D.L.; Li, M.; Damaris, R.N.; Bu, C.; Xue, J.Y.; Yang, P.F. Quantitative ubiquitylomics approach for characterizing the dynamic change and extensive modulation of ubiquitylation in rice seed germination. Plant J. 2020, 101, 1430–1447. [Google Scholar] [CrossRef]
- Wang, Y.F.; Chao, Q.; Li, Z.; Lu, T.-C.; Zheng, H.-Y.; Zhao, C.-F.; Shen, Z.; Li, X.-H.; Wang, B.-C. Large-scale Identification and Time-course Quantification of Ubiquitylation Events During Maize Seedling De-etiolation. Genom. Proteom. Bioinform. 2019, 17, 603–622. [Google Scholar] [CrossRef]
- Xie, X.; Kang, H.X.; Liu, W.D.; Wang, G.L. Comprehensive Profiling of the Rice Ubiquitome Reveals the Significance of Lysine Ubiquitination in Young Leaves. J. Proteome Res. 2015, 14, 2017–2025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, L.R.; Shi, C.N.; Tian, Q.Z.; Lv, G.G.; Wang, Y.; Cui, D.Q.; Chen, F. Comprehensive profiling of lysine ubiquitome reveals diverse functions of lysine ubiquitination in common wheat. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.; Vert, G. Unraveling K63 Polyubiquitination Networks by Sensor-Based Proteomics. Plant Physiol. 2016, 171, 1808–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Li, C.F.; Ruan, D.N.; Powers, S.; Thompson, P.A.; Frohman, M.A.; Chan, C.H. The DNA Damage Transducer RNF8 Facilitates Cancer Chemoresistance and Progression through Twist Activation. Mol. Cell 2016, 63, 1021–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, C.; Held, J.M.; Gibson, B.W.; Benz, C.C. ErbB2 Trafficking and Degradation Associated with K48 and K63 Polyubiquitination. Cancer Res. 2010, 70, 3709–3717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deruyffelaere, C.; Bouchez, I.; Morin, H.; Guillot, A.; Miquel, M.; Froissard, M.; Chardot, T.; D’Andrea, S. Ubiquitin-Mediated Proteasomal Degradation of Oleosins is Involved in Oil Body Mobilization During Post-Germinative Seedling Growth in Arabidopsis. Plant Cell Physiol. 2015, 56, 1374–1387. [Google Scholar] [CrossRef] [Green Version]
- Valencia, J.P.; Goodman, K.; Otegui, M.S. Endocytosis and Endosomal Trafficking in Plants. Annu. Rev. Plant Biol. 2016, 67, 309–335. [Google Scholar] [CrossRef]
- Romero-Barrios, N.; Vert, G. Proteasome-independent functions of lysine-63 polyubiquitination in plants. New Phytol. 2018, 217, 995–1011. [Google Scholar] [CrossRef] [Green Version]
- Haglund, K.; Shimokawa, N.; Szymkiewicz, I.; Dikic, I. Cbl-directed monoubiquitination of CIN85 is involved in regulation of ligand-induced degradation of EGF receptors. Proc. Natl. Acad. Sci. USA 2002, 99, 12191–12196. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Hankins, H.M.; MacDonald, C.; Erlinger, S.J.; Frazier, M.N.; Diab, N.S.; Piper, R.C.; Jackson, L.P.; MacGurn, J.A.; Graham, T.R. COPI mediates recycling of an exocytic SNARE by recognition of a ubiquitin sorting signal. eLife 2017, 6, e28342. [Google Scholar] [CrossRef]
- Clague, M.J.; Heride, C.; Urbe, S. The demographics of the ubiquitin system. Trends Cell Biol. 2015, 25, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, M.; Vialaret, J.; Li, G.-W.; Hem, S.; Prado, K.; Rossignol, M.; Maurel, C.; Santoni, V. Coordinated post-translational responses of aquaporins to abiotic and nutritional stimuli in Arabidopsis roots. Mol. Cell. Proteom. 2013, 12, 3886–3897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martiniere, A.; Fiche, J.B.; Smokvarska, M.; Mari, S.; Alcon, C.; Dumont, X.; Hematy, K.; Jaillais, Y.; Nollmann, M.; Maurel, C. Osmotic Stress Activates Two Reactive Oxygen Species Pathways with Distinct Effects on Protein Nanodomains and Diffusion. Plant Physiol. 2019, 179, 1581–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, B.; Bhatt, T.K. Genome-wide identification and expression analysis of E2 ubiquitin-conjugating enzymes in tomato. Sci. Rep. 2017, 7, 8613. [Google Scholar] [CrossRef] [Green Version]
- Zhiguo, E.; Zhang, Y.P.; Li, T.T.; Wang, L.; Zhao, H.M. Characterization of the Ubiquitin-Conjugating Enzyme Gene Family in Rice and Evaluation of Expression Profiles under Abiotic Stresses and Hormone Treatments. PLoS ONE 2015, 10, e0122621. [Google Scholar]
- Zhou, G.A.; Chang, R.Z.; Qiu, L.J. Overexpression of soybean ubiquitin-conjugating enzyme gene GmUBC2 confers enhanced drought and salt tolerance through modulating abiotic stress-responsive gene expression in Arabidopsis. Plant Mol. Biol. 2010, 72, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Liu, R.J.; Wang, Q.; Xie, Q. ERAD Tuning of the HRD1 Complex Component AtOS9 Is Modulated by an ER-Bound E2, UBC32. Mol. Plant 2017, 10, 891–894. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Zhong, Y.W.; Wu, Y.R.; Liu, L.J.; Wang, P.F.; Liu, R.J.; Cui, F.; Li, Q.L.; Yang, X.Y.; Fang, S.Y.; et al. HRD1-mediated ERAD tuning of ER-bound E2 is conserved between plants and mammals. Nat. Plants 2016, 2, 16094. [Google Scholar] [CrossRef]
- Cui, F.; Liu, L.J.; Li, Q.L.; Yang, C.W.; Xie, Q. UBC32 Mediated Oxidative Tolerance in Arabidopsis. J. Genet. Genom. 2012, 39, 415–417. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, Z.P.; Xu, Y.; Yu, M.; Ren, Y.; Zhang, S.Z.; Yang, G.D.; Huang, J.G.; Yan, K.; Zheng, C.C.; et al. Regulation of the stability and ABA import activity of NRT1.2/NPF4.6 by CEPR2-mediated phosphorylation in Arabidopsis. Mol. Plant 2021, 14, 633–646. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, Y.J.; Ding, D.; Zhou, Y.; Ding, L.P.; Wei, J.H.; Wang, H.Z. Endoplasmic reticulum-localized UBC34 interaction with lignin repressors MYB221 and MYB156 regulates the transactivity of the transcription factors in Populus tomentosa. BMC Plant Biol. 2019, 19, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 222, 680–865. [Google Scholar] [CrossRef] [PubMed]
- Balliau, T.; Blein-Nicolas, M.; Zivy, M. Evaluation of Optimized Tube-Gel Methods of Sample Preparation for Large-Scale Plant Proteomics. Proteomes 2018, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Rofidal, V.; Hem, S.; Gil, J.; Nosarzewska, J.; Berger, N.; Demolombe, V.; Bouzayen, M.; Azhar, B.J.; Shakeel, S.N.; et al. Targeted Proteomics Allows Quantification of Ethylene Receptors and Reveals SIETR3 Accumulation in Never-Ripe Tomatoes. Front. Plant Sci. 2019, 10, 1054. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.Y.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGI | Description | Ubi-Peptide Sequence | K-Ub | Ubi- Peptide Ratio | Protein Ratio | UBC 32 | UBC 34 |
|---|---|---|---|---|---|---|---|
| proteins with decreased ubiquitination | |||||||
| AT1G01580.1 | FRO2 | IEAFITRDNDAGDEAKAGK | 528 | DISP | INV | ||
| AT1G08090.1 | NRT2;1 | ATLEKAGEVAKDKFGK | 259 | DISP | 1.92 (p = 0.03) | ||
| AT1G13480.1 | Protein of unknown function | LDSELTSLGKSIEIGK | 211 | DISP | INV | ||
| AT1G32450.1 | NRT1;5 | SacCLEIYNKDTMoxKK | 9 or 13 or 14 | DISP | INV | ||
| AT1G48210.2 | Protein kinase superfamily protein | LSEDKVKQCVDAR | 300 | DISP | 1.22 (p = 0.03) | ||
| AT1G58030.1 | CAT2 | DGLLPSIFSDINKR | 369 | DISP | INV | ||
| AT2G17440.1 | PIRL5, ras group-related | DITEKGAQAVVQYMNDLVEAR | 484 | DISP | 1.51 (p = 0.01) | ||
| AT2G23200.1 | receptor-like protein kinase | SKGTIDEILDPSLIGQIETNSLKK | 710 | DISP | INV | ||
| AT2G31610.1 | 40S ribosomal protein S3-1 | TQNVLGEKGRR | 62 | 0.33 (p = 0.01) | INV | ||
| AT3G01290.1 | HIR2 | AEGEAESKYLSGLGIAR | 196 | DISP | 1.23 (p = 0.02) | ||
| AT3G04840.1 | 40S ribosomal protein S3a-1 | NVGKTLVSR | 45 | DISP | INV | ||
| AT3G17410.1 | CARK1 | LSEDKVKQCVDAR | 301 | DISP | 1.27 (p = 0.03) | ||
| AT3G47210.1 | Protein of unknown function | KYIISYINEQVELDSR | 62 | DISP | 1.28 (p = 0.03) | ||
| AT3G51550.1 | Feronia | VLGVGGFGKVYR | 549 | DISP | INV | ||
| AT3G53480.1 | ABC transporter ABCG37 | STLLDDGDESMoxTEKGR | 88 | DISP | INV | ||
| AT3G63080.1 | glutathione peroxidase | DSSGKEVDLSVYQGK | 25 | DISP | INV | yes | |
| AT4G08620.1 | SULTR1;1 | DFKGQTPAK | 55 | DISP | INV | ||
| AT4G33360.1 | Farnesol deshydrogenase | NVLEAVKETKTVQK | 112 or 115 or 119 | DISP | INV | ||
| AT4G37060.1 | PATATIN-like protein 5 | IDDDTLEGDASTLDLSTKSNLENLIK | 340 | DISP | INV | ||
| AT5G14040.1 | Mitochondrial phosphate carrier | FIKSEGYGGLYK | 222 | DISP | INV | ||
| AT5G56010.1 | HSP 90-3 | APFDLFDTKK | 326 or 327 | DISP | INV | ||
| AT5G65380.1 | MATE efflux family protein | VANELGAGNGKGAR | 334 | DISP | INV | ||
| proteins with increased ubiquitination | |||||||
| AT1G01580.1 | FRO2 | DNDAGDEAKAGKIK | 528 and 531 | APP | INV | ||
| AT1G02520.1 | ABC transporter ABCB11 | KQCEGPIKDGIK | 919 | APP | INV | ||
| AT1G08930.2 | ERD6 | DTIDMoxTENGGETKMSELFQR | 281 | APP | INV | ||
| AT1G08930.2 | ERD6 | DTIDMoxTENGGETKMoxSELFQR | 281 | APP | INV | ||
| AT1G11680.1 | Sterol 14-demethylase | SGKTENDMoxLQCFIESK | 253 | APP | INV | ||
| AT1G12110.1 | NRT1, NPF6.3 | KLELPADPSYLYDVDDIIAAEGSphMoxKGK | 267 or 291 or 293 | APP | INV | yes | |
| AT1G44170.3 | ALDH3H1 | LSKLLDEK | 242 | APP | INV | ||
| AT1G55450.1 | methyl transferase | AacALSDKLADAYQNAR | 6 | 2.3 (p = 0.01) | INV | ||
| AT1G59870.1 | ABC transporter ABCB11 | EVDVTKLDGEDRQK | 94 | APP | INV | ||
| AT1G61250.2 | SCAMP1, secretory carrier3 | ELQAKEAELK | 71 | APP | INV | yes | |
| AT1G61670.1 | Two-component response regulator | NELLFGLPDDVEEGKRE | 511 | APP | INV | ||
| AT2G02040.1 | NRT1, NPF 8.3 | AAVISEEESKSGDYSNSWR | 325 | APP | INV | yes | yes |
| AT2G24720.1 | glutamate receptor 2.2 | DLWKEFLK | 864 | APP | INV | ||
| AT2G32270.1 | Zinc transporter 3 | VSDGETphGESSVDSEKVQILR | 177 | APP | INV | ||
| AT2G38360.1 | prenylated RAB acceptor | SALSKPESISDAAVR | 68 | 1.72 (p = 0.02) | INV | ||
| AT2G47000.1 | ABC transporter ABCB4 | AacSESGLNGDPNILEEVSETKR | 21 | APP | INV | ||
| AT3G04840.1 | 40S ribosomal protein S3a-1 | IASEGLKHR | 62 | APP | INV | ||
| AT3G08680.2 | inactive receptor kinase | AYYFSKDEK | 407 or 410 | APP | INV | yes | |
| AT3G27770.2 | hypoxia response protein | SPLIDGDNMoxVSFEKR | 125 | APP | INV | yes | yes |
| AT3G29310.1 | BAG1 | FVQYVDDCVVKR | 230 | APP | INV | ||
| AT3G45710.1 | NRT1, NPF2.5 | DEDYHQYGLGKEAK | 272 | APP | INV | ||
| AT3G51550.1 | Feronia | AATKNFDESR | 534 | APP | INV | ||
| AT3G53420.2 | PIP2;1 | ASGSKSLGSphFR | 276 | 1.79 (p = 0.01) | INV | yes * | |
| AT3G53420.2 | PIP2;1 | AacKDVEAVPGEGFQTR | 3 | 1.69 (p = 0.01) | INV | yes * | |
| AT3G60330.2 | AHA7 | TQHGLETGQKPVYER | 903 | APP | 1.26 (p = 0.01) | ||
| AT3G62250.1 | 40S ribosomal protein S27a-3 | MQIFVKTLTGKTITLEVESSDTIDNVK | 11 | APP | INV | ||
| AT4G01440.1 | nodulin MtN21 EamA-like | FNEDDQEEDDDEQYKK | 354 or 355 | APP | INV | ||
| AT4G09000.1 | GRF1 | AVDKDELTVEER | 42 | APP | INV | ||
| AT4G25090.1 | RBOHG | KELSDMoxLTESLKPTR | 267 | APP | INV | ||
| AT4G30190.1 | AHA2 | AWLNLFENK | 857 | APP | 1.44 (p = 0.01) | yes | yes |
| AT4G30190.1 | AHA2 | WSEQEAAILVPGDIVSIK | 157 | APP | 1.44 (p = 0.01) | yes | yes |
| AT4G30190.1 | AHA2 | TphLHGLQPKEAVNIFPEK | 888 | APP | 1.44 (p = 0.01) | yes | yes |
| AT4G30190.1 | AHA2 | SacSLEDIKNETVDLEK | 8 | 3.00 (p = 0.02) | 1.44 (p = 0.01) | yes | yes |
| AT5G25930.1 | LRR protein kinase family | LLVYEYLEKR | 767 | APP | INV | ||
| AT5G35200.1 | clathrin assembly protein | EAPLAAGVKK | 310 | APP | INV | ||
| AT5G39510.1 | v-SNARE 11 | KILTDMoxTR | 184 | APP | INV | ||
| AT5G47910.1 | RBOHD | NKLNLPNFLK | 541 | APP | INV | ||
| AT5G59970.1 | Histone superfamily protein | DNIQGITKPAIR | 32 | 1.57 (p = 0.02) | INV | ||
| AT5G62300.2 | 40S ribosomal protein S20-1 | AacTAYQPMKPGKAGLEEPLEQIHK | 9 and 12 | APP | INV | ||
| AT5G62390.1 | BAG7 | AIAAAEAEKK | 195 | APP | INV | ||
| AT5G62390.1 | BAG7 | LEPEYPLKYLCDR | 90 | 3.02 (p = 0.03) | INV | ||
| AT5G62390.1 | BAG7 | RLEPEYPLKYLCDR | 90 | 2.09 (p = 0.01) | INV | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berger, N.; Demolombe, V.; Hem, S.; Rofidal, V.; Steinmann, L.; Krouk, G.; Crabos, A.; Nacry, P.; Verdoucq, L.; Santoni, V. Root Membrane Ubiquitinome under Short-Term Osmotic Stress. Int. J. Mol. Sci. 2022, 23, 1956. https://doi.org/10.3390/ijms23041956
Berger N, Demolombe V, Hem S, Rofidal V, Steinmann L, Krouk G, Crabos A, Nacry P, Verdoucq L, Santoni V. Root Membrane Ubiquitinome under Short-Term Osmotic Stress. International Journal of Molecular Sciences. 2022; 23(4):1956. https://doi.org/10.3390/ijms23041956
Chicago/Turabian StyleBerger, Nathalie, Vincent Demolombe, Sonia Hem, Valérie Rofidal, Laura Steinmann, Gabriel Krouk, Amandine Crabos, Philippe Nacry, Lionel Verdoucq, and Véronique Santoni. 2022. "Root Membrane Ubiquitinome under Short-Term Osmotic Stress" International Journal of Molecular Sciences 23, no. 4: 1956. https://doi.org/10.3390/ijms23041956