
Antitumor Effect of Regorafenib on MicroRNA Expression in Hepatocellular Carcinoma Cell Lines

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
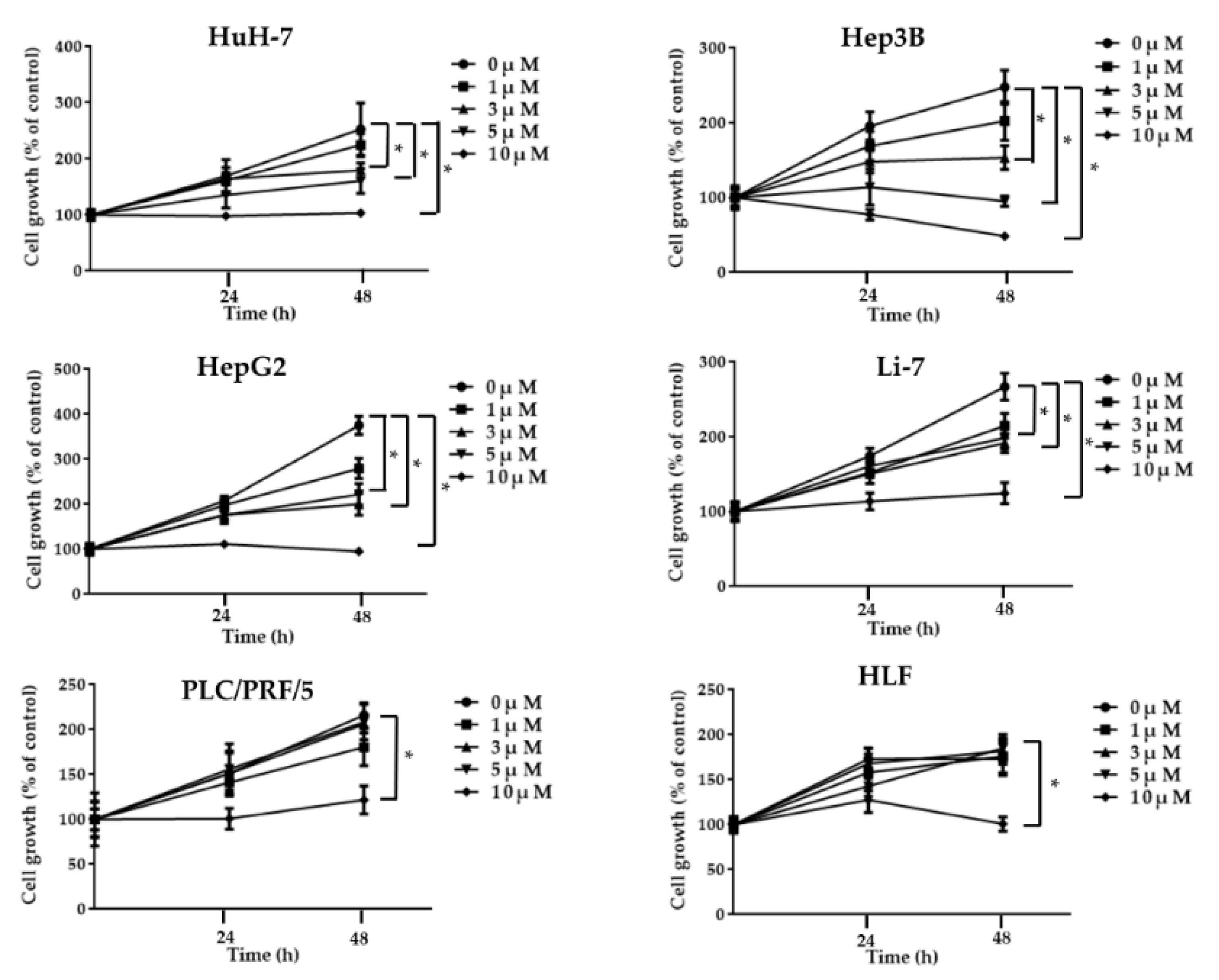
2.1. Regorafenib Suppresses Cell Proliferation of Human HCC Cells
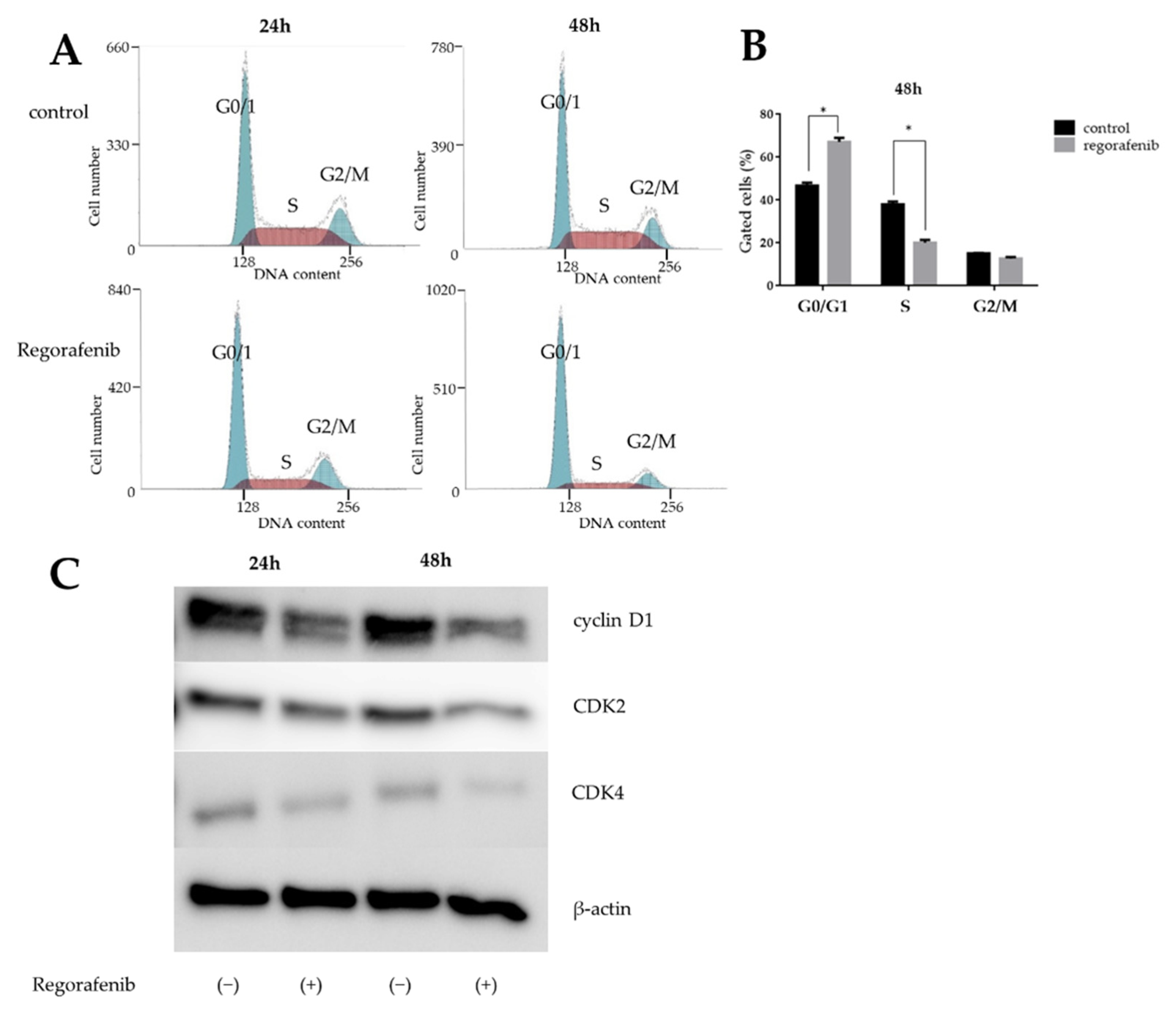
2.2. Regorafenib Induces Cell Cycle Arrest in the G0/G1 Phase and Affects Cell Cycle-Related Proteins
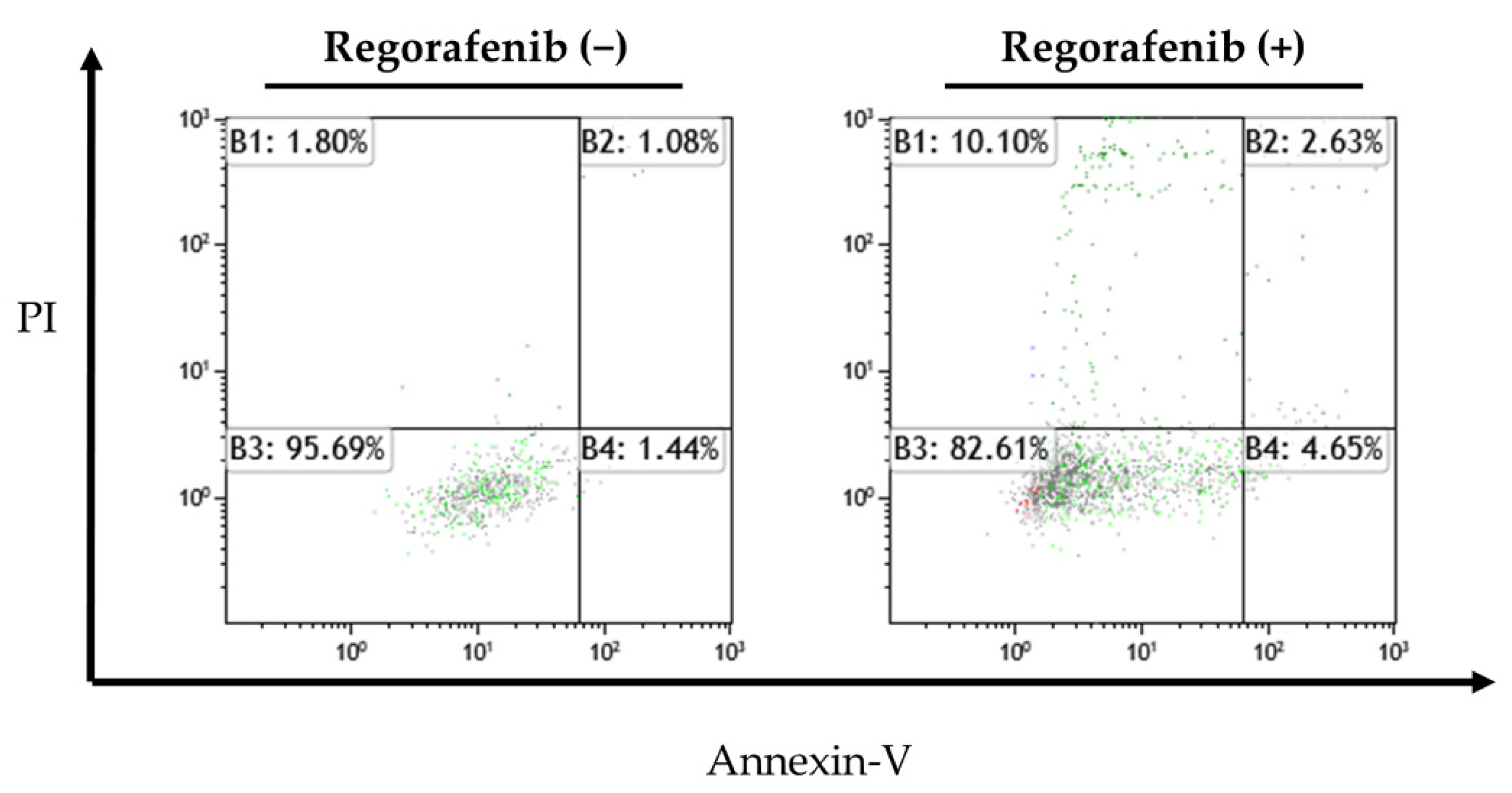
2.3. Apoptosis-Inducing Effect of Regorafenib in HCC Cells
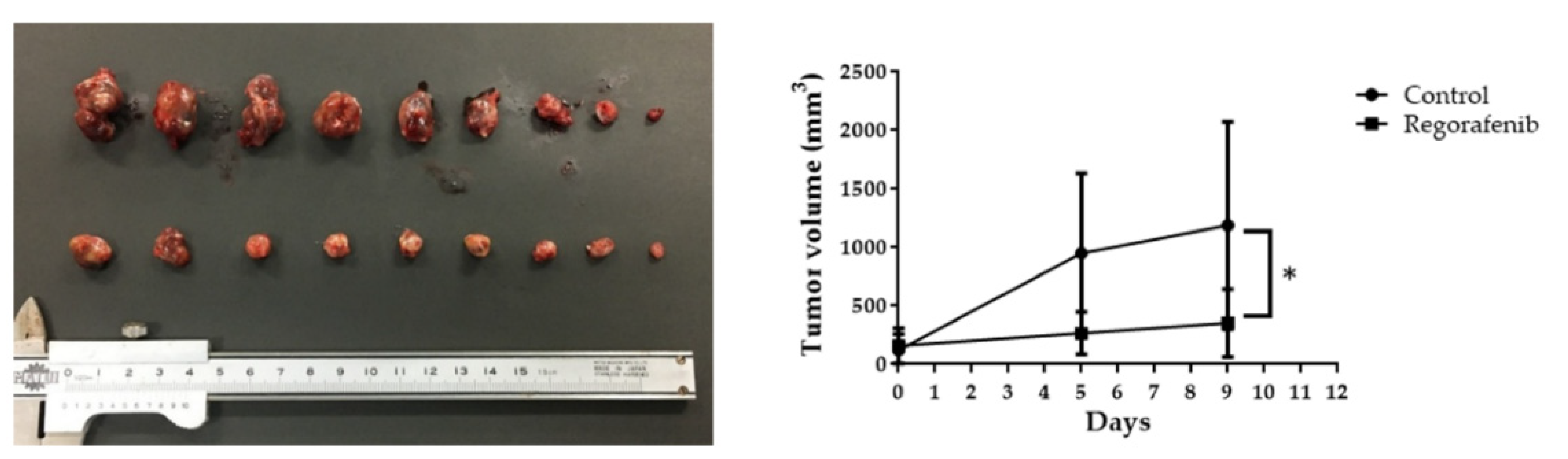
2.4. Regorafenib Suppresses HCC Tumor Growth In Vivo
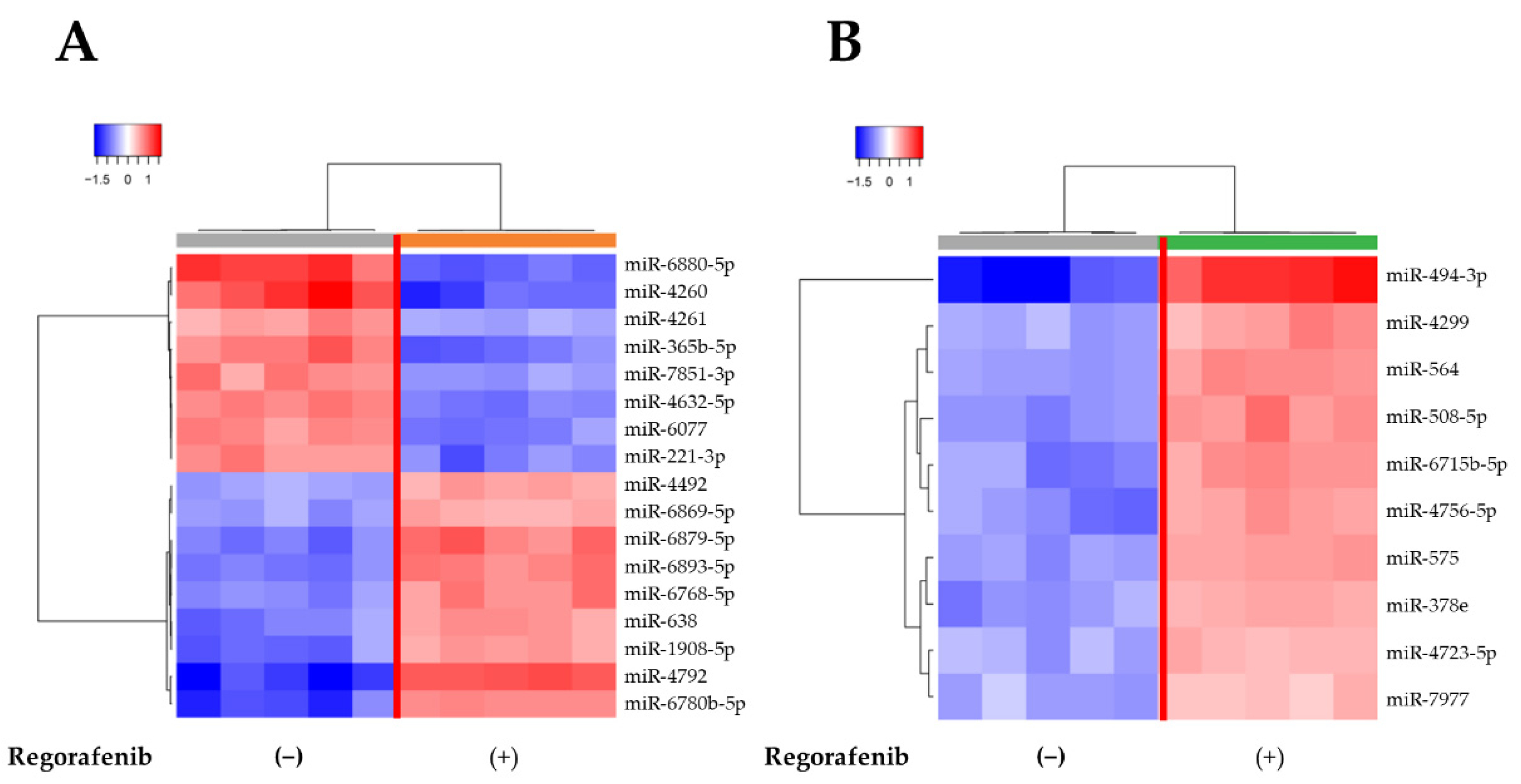
2.5. MiRNA Expression Signatures Are Different in Regorafenib-Treated and -Untreated HuH-7 Cells
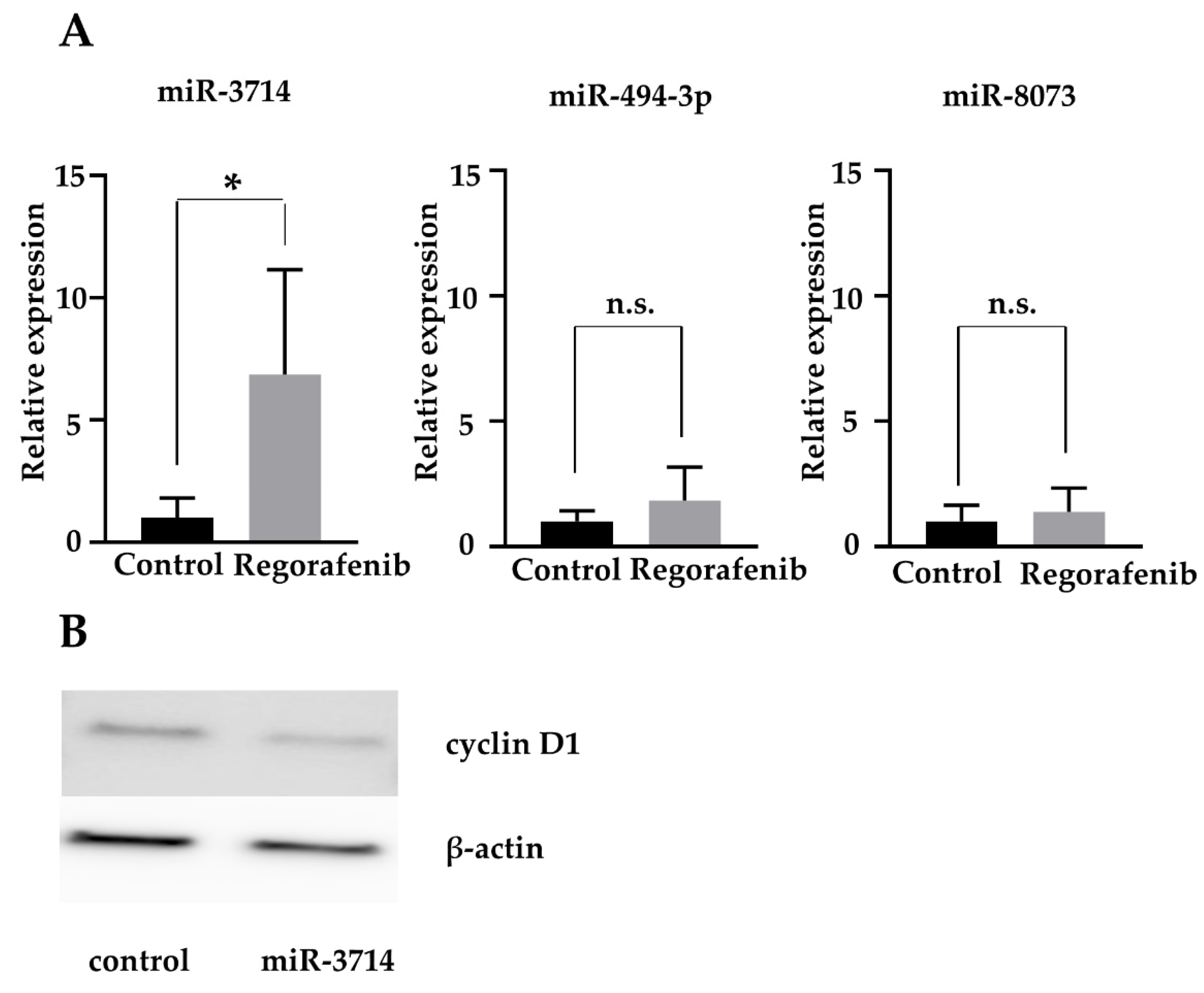
2.6. MiR-3714 Suppresses Cyclin D1 Expression in HCC Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Lines and Culture
4.3. Cell Proliferation Assay
4.4. Cell Cycle and Apoptosis Analysis
4.5. Western Blot Analysis
4.6. Analysis of the Therapeutic Effect of Regorafenib In Vivo
4.7. Microarray Analysis of MiRNAs
4.8. Quantitative Polymerase Chain Reaction (qPCR) Analysis of MiRNAs
4.9. Transfection of MiRNA Mimic
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [Green Version]
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet 2018, 391, 1301–1314. [Google Scholar] [CrossRef]
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet 2012, 379, 1245–1255. [Google Scholar] [CrossRef]
- Altekruse, S.F.; Henley, S.J.; Cucinelli, J.E.; McGlynn, K.A. Changing Hepatocellular Carcinoma Incidence and Liver Cancer Mortality Rates in the United States. Am. J. Gastroenterol. 2014, 109, 542–553. [Google Scholar] [CrossRef] [Green Version]
- Tovoli, F.; Granito, A.; de Lorenzo, S.; Bolondi, L. Regorafenib for the treatment of hepatocellular carcinoma. Drugs Today 2018, 54, 5–13. [Google Scholar] [CrossRef]
- Bruix, J.; Qin, S.; Merle, P.; Granito, A.; Huang, Y.-H.; Bodoky, G.; Pracht, M.; Yokosuka, O.; Rosmorduc, O.; Breder, V.; et al. Regorafenib for patients with hepatocellular carcinoma who progressed on sorafenib treatment (RESORCE): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 389, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Rimassa, L.; Pressiani, T.; Personeni, N.; Santoro, A. Regorafenib for the treatment of unresectable hepatocellular carcinoma. Expert Rev. Anticancer Ther. 2017, 17, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Montal, R.; Sia, D.; Finn, R.S. Molecular therapies and precision medicine for hepatocellular carcinoma. Nat. Rev. Clin. Oncol. 2018, 15, 599–616. [Google Scholar] [CrossRef]
- Wilhelm, S.M.; Dumas, J.; Adnane, L.; Lynch, M.; Carter, C.A.; Schütz, G.; Thierauch, K.-H.; Zopf, D. Regorafenib (BAY 73-4506): A new oral multikinase inhibitor of angiogenic, stromal and oncogenic receptor tyrosine kinases with potent preclinical antitumor activity. Int. J. Cancer 2011, 129, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Abou-Elkacem, L.; Arns, S.; Brix, G.; Gremse, F.; Zopf, D.; Kiessling, F.; Lederle, W. Regorafenib Inhibits Growth, Angiogenesis, and Metastasis in a Highly Aggressive, Orthotopic Colon Cancer Model. Mol. Cancer Ther. 2013, 12, 1322–1331. [Google Scholar] [CrossRef] [Green Version]
- Grothey, A.; van Cutsem, E.; Sobrero, A.; Siena, S.; Falcone, A.; Ychou, M.; Humblet, Y.; Bouché, O.; Mineur, L.; Barone, C.; et al. Regorafenib monotherapy for previously treated metastatic colorectal cancer (CORRECT): An international, multicentre, randomised, placebo-controlled, phase 3 trial. Lancet 2013, 381, 303–312. [Google Scholar] [CrossRef]
- Demetri, G.D.; Reichardt, P.; Kang, Y.-K.; Blay, J.-Y.; Rutkowski, P.; Gelderblom, H.; Hohenberger, P.; Leahy, M.; von Mehren, M.; Joensuu, H.; et al. Efficacy and safety of regorafenib for advanced gastrointestinal stromal tumours after failure of imatinib and sunitinib (GRID): An international, multicentre, randomised, placebo-controlled, phase 3 trial. Lancet 2013, 381, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Wu, L.-W.; Zhang, Z.-Y.; Chen, M.-L.; Li, Y.-L.; Zhang, C. The anti-tumor effect of regorafenib in lung squamous cell carcinoma in vitro. Biochem. Biophys. Res. Commun. 2018, 503, 1123–1129. [Google Scholar] [CrossRef]
- Chiang, C.H.; Chung, J.G.; Hsu, F.T. Regorefenib induces extrinsic/intrinsic apoptosis and inhibits MAPK/NF-κB-modulated tumor progression in bladder cancer In Vitro and In Vivo. Environ. Toxicol. 2019, 34, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.-J.; Liu, Y.-C.; Hsu, F.-T. Protein Kinase B and Extracellular Signal-Regulated Kinase Inactivation is Associated with Regorafenib-Induced Inhibition of Osteosarcoma Progression In Vitro and In Vivo. J. Clin. Med. 2019, 8, 900. [Google Scholar] [CrossRef] [Green Version]
- Lai, S.C.; Su, Y.T.; Chi, C.C.; Kuo, Y.C.; Lee, K.F.; Wu, Y.C.; Lan, P.C.; Yang, M.H.; Chang, T.S.; Huang, Y.-H. DNMT3b/OCT4 expression confers sorafenib resistance and poor prognosis of hepatocellular carcinoma through IL-6/STAT3 regulation. Exp. Clin. Cancer Res. 2019, 38, 474. [Google Scholar] [CrossRef] [Green Version]
- Murakami, S.; Ninomiya, W.; Sakamoto, E.; Shibata, T.; Akiyama, H.; Tashiro, F. SRY and OCT4 Are Required for the Acquisition of Cancer Stem Cell-Like Properties and Are Potential Differentiation Therapy Targets. Stem Cells 2015, 33, 2652–2663. [Google Scholar] [CrossRef]
- Chen, K.-F.; Yeh, P.-Y.; Yeh, K.-H.; Lu, Y.-S.; Huang, S.-Y.; Cheng, A.-L. Down-regulation of Phospho-Akt Is a Major Molecular Determinant of Bortezomib-Induced Apoptosis in Hepatocellular Carcinoma Cells. Cancer Res. 2008, 68, 6698–6707. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.F.; Yu, H.C.; Liu, T.H.; Lee, S.S.; Chen, P.J.; Cheng, A.L. Synergistic interactions between sorafenib and bortezomib in hepato-cellular carcinoma involve PP2A-dependent Akt inactivation. J. Hepatol. 2010, 52, 88–95. [Google Scholar] [CrossRef]
- Ogasawara, S.; Mihara, Y.; Kondo, R.; Kusano, H.; Akiba, J.; Yano, H. Antiproliferative Effect of Lenvatinib on Human Liver Cancer Cell Lines In Vitro and In Vivo. Anticancer Res. 2019, 39, 5973–5982. [Google Scholar] [CrossRef]
- Matsuki, M.; Hoshi, T.; Yamamoto, Y.; Ikemori-Kawada, M.; Minoshima, Y.; Funahashi, Y.; Matsui, J. Lenvatinib inhibits angiogenesis and tumor fibroblast growth factor signaling pathways in human hepatocellular carcinoma models. Cancer Med. 2018, 7, 2641–2653. [Google Scholar] [CrossRef] [PubMed]
- Futami, T.; Okada, H.; Kihara, R.; Kawase, T.; Nakayama, A.; Suzuki, T.; Kameda, M.; Shindoh, N.; Terasaka, T.; Hirano, M.; et al. ASP5878, a Novel Inhibitor of FGFR1, 2, 3, and 4, Inhibits the Growth of FGF19-Expressing Hepatocellular Carcinoma. Mol. Cancer Ther. 2016, 16, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myojin, Y.; Kodama, T.; Maesaka, K.; Motooka, D.; Sato, Y.; Tanaka, S.; Abe, Y.; Ohkawa, K.; Mita, E.; Hayashi, Y.; et al. ST6GAL1 Is a Novel Serum Biomarker for Lenvatinib-Susceptible FGF19-Driven Hepatocellular Carcinoma. Clin. Cancer Res. 2021, 27, 1150–1161. [Google Scholar] [CrossRef] [PubMed]
- Qie, S.; Diehl, J.A. Cyclin D1, cancer progression, and opportunities in cancer treatment. J. Mol. Med. 2016, 94, 1313–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morishita, A.; Masaki, T. miRNA in hepatocellular carcinoma. Hepatol. Res. 2015, 45, 128–141. [Google Scholar] [CrossRef]
- Shi, D.-M.; Bian, X.-Y.; Qin, C.-D.; Wu, W.-Z. miR-106b-5p promotes stem cell-like properties of hepatocellular carcinoma cells by targeting PTEN via PI3K/Akt pathway. OncoTargets Ther. 2018, 11, 571–585. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Song, G.; Yao, L.; Liu, Y.; Liu, M.; Li, S.; Tang, H. miR-3928v is induced by HBx via NF-kappaB/EGR1 and contributes to hepatocellular carcinoma malignancy by down-regulating VDAC3. J. Exp. Clin. Cancer Res. 2018, 37, 14. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, Y.; Li, X.; Liang, X.; Wang, L.; Song, J.; Zhang, X.; Zhang, C.; Niu, J.; Zhang, P.; et al. microRNA-874 suppresses tumor proliferation and metastasis in hepatocellular carcinoma by targeting the DOR/EGFR/ERK pathway. Cell Death Dis. 2018, 9, 130. [Google Scholar] [CrossRef]
- Kohno, T.; Morishita, A.; Iwama, H.; Fujita, K.; Tani, J.; Takuma, K.; Nakahara, M.; Oura, K.; Tadokoro, T.; Nomura, T.; et al. Comprehensive analysis of circulating microRNAs as predictive biomarkers for sorafenib therapy outcome in hepatocellular carcinoma. Oncol. Lett. 2020, 20, 1727–1733. [Google Scholar] [CrossRef]
- Teufel, M.; Seidel, H.; Köchert, K.; Meinhardt, G.; Finn, R.S.; Llovet, J.M.; Bruix, J. Biomarkers Associated with Response to Regorafenib in Patients with Hepatocellular Carcinoma. Gastroenterology 2019, 156, 1731–1741. [Google Scholar] [CrossRef] [Green Version]
- Weidle, U.H.; Schmid, D.; Birzele, F.; Brinkmann, U. MicroRNAs Involved in Metastasis of Hepatocellular Carcinoma: Target Candidates, Functionality and Efficacy in Animal Models and Prognostic Relevance. Cancer Genom. Proteom. 2020, 17, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, A.; Takayama, A.; Arai, T.; Kawauchi, J.; Sudo, H. MicroRNA-8073: Tumor suppressor and potential therapeutic treatment. PLoS ONE 2018, 13, e0209750. [Google Scholar] [CrossRef] [PubMed]
- Faversani, A.; Amatori, S.; Augello, C.; Colombo, F.; Porretti, L.; Fanelli, M.; Ferrero, S.; Palleschi, A.; Pelicci, P.G.; Belloni, E.; et al. miR-494-3p is a novel tumor driver of lung carcinogenesis. Oncotarget 2017, 8, 7231–7247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-M.; Wang, B.-Y.; Lee, C.-H.; Lee, H.-T.; Li, J.-J.; Hong, G.-C.; Hung, Y.-C.; Chien, P.-J.; Chang, C.-Y.; Hsu, L.-S.; et al. Hinokitiol up-regulates miR-494-3p to suppress BMI1 expression and inhibits self-renewal of breast cancer stem/progenitor cells. Oncotarget 2017, 8, 76057–76068. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Huang, Z.-P.; Liu, J.; Qiu, Y.; Tao, Y.-P.; Wang, M.-C.; Yao, H.; Hou, K.-Z.; Gu, F.-M.; Xu, X.-F. MiR-494-3p promotes PI3K/AKT pathway hyperactivation and human hepatocellular carcinoma progression by targeting PTEN. Sci. Rep. 2018, 8, 10461. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lim, W.; Bazer, F.W.; Whang, K.-Y.; Song, G. Quercetin inhibits proliferation of endometriosis regulating cyclin D1 and its target microRNAs in vitro and in vivo. J. Nutr. Biochem. 2019, 63, 87–100. [Google Scholar] [CrossRef]
- Ghanbarian, M.; Afgar, A.; Yadegarazari, R.; Najafi, R.; Teimoori-Toolabi, L. Through oxaliplatin resistance induction in colorectal cancer cells, increasing ABCB1 level accompanies decreasing level of miR-302c-5p, miR-3664-5p and miR-129-5p. Biomed. Pharmacother. 2018, 108, 1070–1080. [Google Scholar] [CrossRef]
- Galun, D.; Srdic-Rajic, T.; Bogdanovic, A.; Loncar, Z.; Zuvela, M. Targeted therapy and personalized medicine in hepatocellular carcinoma: Drug resistance, mechanisms, and treatment strategies. J. Hepatocell. Carcinoma 2017, 4, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Ghousein, A.; Mosca, N.; Cartier, F.; Charpentier, J.; Dupuy, J.-W.; Raymond, A.; Bioulac-Sage, P.; Grosset, C.F. miR-4510 blocks hepatocellular carcinoma development through RAF1 targeting and RAS/RAF/MEK/ERK signalling inactivation. Liver Int. 2020, 40, 240–251. [Google Scholar] [CrossRef]
- Masaki, T.; Tokuda, M.; Yoshida, S.; Nakai, S.; Morishita, A.; Uchida, N.; Funaki, T.; Kita, Y.; Funakoshi, F.; Nonomura, T.; et al. Comparison study of the expressions of myristoylated alanine-rich C kinase substrate in hepatocellular carcinoma, liver cirrhosis, chronic hepatitis, and normal liver. Int. J. Oncol. 2005, 26, 661–671. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Du, Y.; Ma, H.; Liang, Q.; Zhu, X.; Tian, J. Preclinical comparison of regorafenib and sorafenib efficacy for hepatocellular carcinoma using multimodality molecular imaging. Cancer Lett. 2019, 453, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, Y.; Yu, Y.; Pang, J.; Woodfield, S.E.; Tao, L.; Guan, S.; Zhang, H.; Bieerkehazhi, S.; Shi, Y.; et al. Small molecule inhibitor regorafenib inhibits RET signaling in neuroblastoma cells and effectively suppresses tumor growth In Vivo. Oncotarget 2017, 8, 104090–104103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Incalci, M.; Colombo, T.; Ubezio, P.; Nicoletti, I.; Giavazzi, R.; Erba, E.; Ferrarese, L.; Meco, D.; Riccardi, R.; Sessa, C.; et al. The combination of yondelis and cisplatin is synergistic against human tumor xenografts. Eur. J. Cancer 2003, 39, 1920–1926. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated | Down Regulated | ||||||
|---|---|---|---|---|---|---|---|
| miRNA | FC | p Value | FDR | miRNA | FC | p Value | FDR |
| (Treated/ Untreated) | (Treated/ Untreated) | ||||||
| miR-6891-5p | 13.915 | 0.000005 | 0.000080 | miR-206 | 0.131 | 0.000025 | 0.000194 |
| miR-3162-5p | 12.868 | 0.000074 | 0.000400 | miR-1292-5p | 0.158 | 0.000007 | 0.000092 |
| miR-3687 | 11.359 | 0.000001 | 0.000037 | miR-4287 | 0.163 | 0.000035 | 0.000230 |
| miR-3679-5p | 9.705 | 0.000013 | 0.000126 | miR-4633-5p | 0.202 | 0.000002 | 0.000056 |
| miR-4476 | 9.241 | 0.000014 | 0.000128 | miR-4260 | 0.209 | 0.000000 | 0.000022 |
| miR-4257 | 7.270 | 0.000001 | 0.000037 | miR-4254 | 0.210 | 0.000004 | 0.000066 |
| miR-6816-5p | 7.039 | 0.000021 | 0.000171 | miR-7843-5p | 0.217 | 0.000015 | 0.000138 |
| miR-6124 | 7.017 | 0.000007 | 0.000092 | miR-6776-3p | 0.219 | 0.000004 | 0.000065 |
| miR-4725-3p | 6.513 | 0.000012 | 0.000125 | miR-6880-5p | 0.223 | 0.000000 | 0.000003 |
| miR-150-3p | 6.180 | 0.000058 | 0.000339 | miR-106b-3p | 0.232 | 0.000016 | 0.000146 |
| miR-3616-3p | 6.046 | 0.000002 | 0.000044 | miR-4319 | 0.240 | 0.000009 | 0.000109 |
| miR-4294 | 5.425 | 0.000089 | 0.000462 | miR-4695-3p | 0.244 | 0.000004 | 0.000065 |
| miR-4792 | 5.366 | 0.000000 | 0.000003 | miR-4758-3p | 0.247 | 0.000012 | 0.000125 |
| miR-6870-5p | 5.156 | 0.000002 | 0.000042 | miR-874-5p | 0.257 | 0.000027 | 0.000200 |
| miR-885-3p | 4.943 | 0.000051 | 0.000304 | miR-6801-3p | 0.261 | 0.000042 | 0.000264 |
| miR-4749-5p | 4.816 | 0.000001 | 0.000034 | miR-887-3p | 0.268 | 0.000034 | 0.000229 |
| miR-365a-5p | 4.812 | 0.000006 | 0.000082 | miR-7114-5p | 0.272 | 0.000026 | 0.000199 |
| miR-1275 | 4.804 | 0.000004 | 0.000066 | miR-4664-5p | 0.283 | 0.000009 | 0.000109 |
| miR-1229-5p | 4.657 | 0.000089 | 0.000462 | miR-5195-3p | 0.287 | 0.000012 | 0.000125 |
| miR-6721-5p | 4.403 | 0.000003 | 0.000064 | miR-365b-5p | 0.298 | 0.000000 | 0.000011 |
| miR-652-5p | 4.156 | 0.000011 | 0.000125 | miR-3177-3p | 0.315 | 0.000002 | 0.000044 |
| miR-1247-5p | 4.036 | 0.000069 | 0.000377 | miR-345-5p | 0.316 | 0.000010 | 0.000111 |
| miR-6511b-5p | 4.019 | 0.000075 | 0.000400 | miR-4687-3p | 0.318 | 0.000005 | 0.000080 |
| miR-6789-5p | 3.907 | 0.000015 | 0.000138 | miR-7155-5p | 0.322 | 0.000033 | 0.000223 |
| miR-6830-5p | 3.768 | 0.000003 | 0.000061 | miR-4632-5p | 0.325 | 0.000000 | 0.000001 |
| miR-6780b-5p | 3.707 | 0.000000 | 0.000018 | miR-6777-5p | 0.325 | 0.000001 | 0.000034 |
| miR-4327 | 3.460 | 0.000038 | 0.000247 | miR-671-3p | 0.328 | 0.000001 | 0.000034 |
| miR-6879-5p | 3.338 | 0.000000 | 0.000011 | miR-1229-3p | 0.331 | 0.000009 | 0.000111 |
| miR-6893-5p | 3.201 | 0.000000 | 0.000003 | miR-5739 | 0.333 | 0.000003 | 0.000061 |
| miR-1915-3p | 3.136 | 0.000032 | 0.000223 | miR-6735-5p | 0.335 | 0.000005 | 0.000080 |
| miR-3188 | 3.020 | 0.000039 | 0.000250 | miR-6077 | 0.335 | 0.000000 | 0.000004 |
| miR-1908-5p | 2.890 | 0.000000 | 0.000018 | miR-1913 | 0.339 | 0.000021 | 0.000171 |
| miR-6768-5p | 2.825 | 0.000000 | 0.000012 | miR-221-3p | 0.349 | 0.000000 | 0.000023 |
| miR-4530 | 2.806 | 0.000003 | 0.000061 | miR-187-5p | 0.350 | 0.000001 | 0.000023 |
| miR-3918 | 2.805 | 0.000020 | 0.000171 | miR-6816-3p | 0.363 | 0.000012 | 0.000125 |
| miR-638 | 2.776 | 0.000000 | 0.000015 | miR-4443 | 0.370 | 0.000004 | 0.000066 |
| miR-4665-5p | 2.769 | 0.000091 | 0.000465 | miR-7851-3p | 0.374 | 0.000000 | 0.000008 |
| miR-663a | 2.752 | 0.000010 | 0.000111 | miR-6741-5p | 0.378 | 0.000004 | 0.000066 |
| miR-6798-5p | 2.677 | 0.000031 | 0.000223 | miR-4323 | 0.379 | 0.000031 | 0.000223 |
| miR-6850-5p | 2.652 | 0.000014 | 0.000128 | miR-4786-3p | 0.381 | 0.000067 | 0.000376 |
| miR-6875-5p | 2.651 | 0.000005 | 0.000069 | miR-4786-5p | 0.384 | 0.000098 | 0.000494 |
| miR-4467 | 2.630 | 0.000003 | 0.000061 | miR-320b | 0.387 | 0.000017 | 0.000152 |
| miR-6858-5p | 2.472 | 0.000002 | 0.000044 | miR-5002-3p | 0.387 | 0.000009 | 0.000109 |
| miR-4459 | 2.470 | 0.000004 | 0.000066 | miR-128-1-5p | 0.388 | 0.000076 | 0.000408 |
| miR-6088 | 2.453 | 0.000034 | 0.000230 | miR-3176 | 0.389 | 0.000001 | 0.000027 |
| miR-149-3p | 2.346 | 0.000039 | 0.000250 | miR-4293 | 0.400 | 0.000035 | 0.000230 |
| miR-4651 | 2.283 | 0.000013 | 0.000126 | miR-6766-3p | 0.401 | 0.000043 | 0.000265 |
| miR-6869-5p | 2.271 | 0.000000 | 0.000003 | miR-320e | 0.404 | 0.000024 | 0.000187 |
| miR-4492 | 2.260 | 0.000000 | 0.000002 | miR-885-5p | 0.405 | 0.000030 | 0.000217 |
| miR-3665 | 2.221 | 0.000013 | 0.000126 | miR-6716-5p | 0.410 | 0.000008 | 0.000103 |
| miR-1202 | 2.189 | 0.000031 | 0.000223 | miR-3135b | 0.412 | 0.000022 | 0.000175 |
| miR-6729-5p | 2.151 | 0.000007 | 0.000089 | miR-4502 | 0.423 | 0.000003 | 0.000064 |
| miR-7111-5p | 2.147 | 0.000013 | 0.000126 | miR-3186-3p | 0.425 | 0.000089 | 0.000462 |
| miR-1237-5p | 2.130 | 0.000004 | 0.000066 | miR-4261 | 0.430 | 0.000000 | 0.000013 |
| miR-5787 | 2.087 | 0.000002 | 0.000050 | miR-675-5p | 0.430 | 0.000018 | 0.000154 |
| miR-6724-5p | 2.058 | 0.000001 | 0.000037 | miR-939-5p | 0.437 | 0.000018 | 0.000154 |
| miR-4756-5p | 2.040 | 0.000069 | 0.000377 | miR-320c | 0.439 | 0.000030 | 0.000217 |
| miR-6889-5p | 2.032 | 0.000060 | 0.000342 | miR-8052 | 0.446 | 0.000006 | 0.000082 |
| miR-762 | 2.024 | 0.000064 | 0.000365 | miR-1228-3p | 0.448 | 0.000019 | 0.000162 |
| miR-4508 | 2.017 | 0.000001 | 0.000025 | miR-320a | 0.450 | 0.000012 | 0.000125 |
| miR-320d | 0.450 | 0.000046 | 0.000283 | ||||
| miR-27b-3p | 0.451 | 0.000089 | 0.000462 | ||||
| miR-3154 | 0.458 | 0.000037 | 0.000239 | ||||
| miR-6782-5p | 0.463 | 0.000003 | 0.000061 | ||||
| miR-4322 | 0.473 | 0.000026 | 0.000197 | ||||
| miR-6859-3p | 0.474 | 0.000091 | 0.000465 | ||||
| miR-6515-3p | 0.476 | 0.000013 | 0.000126 | ||||
| miR-210-5p | 0.477 | 0.000032 | 0.000223 | ||||
| miR-4634 | 0.482 | 0.000027 | 0.000204 | ||||
| miR-296-5p | 0.487 | 0.000035 | 0.000230 | ||||
| miR-4717-3p | 0.491 | 0.000018 | 0.000156 | ||||
| miR-4433a-5p | 0.494 | 0.000012 | 0.000125 | ||||
| miR-4498 | 0.495 | 0.000001 | 0.000037 |
| Upregulated | Down Regulated | ||||||
|---|---|---|---|---|---|---|---|
| miRNA | FC | p Value | FDR | miRNA | FC | p Value | FDR |
| (Treated/ Untreated) | (Treated/ Untreated) | ||||||
| miR-494-3p | 6.335 | 0.000000 | 0.000014 | miR-4429 | 0.391 | 0.000014 | 0.000104 |
| miR-933 | 5.926 | 0.000001 | 0.000023 | miR-6511a-5p | 0.408 | 0.000002 | 0.000031 |
| miR-4502 | 4.810 | 0.000002 | 0.000030 | miR-1247-3p | 0.441 | 0.000012 | 0.000090 |
| miR-2392 | 4.805 | 0.000016 | 0.000112 | miR-5572 | 0.449 | 0.000085 | 0.000351 |
| miR-1273c | 4.457 | 0.000006 | 0.000063 | miR-3180-3p | 0.455 | 0.000011 | 0.000089 |
| miR-6895-5p | 3.346 | 0.000027 | 0.000156 | miR-4448 | 0.473 | 0.000063 | 0.000276 |
| miR-4468 | 3.332 | 0.000006 | 0.000063 | miR-4322 | 0.479 | 0.000008 | 0.000073 |
| miR-508-5p | 2.788 | 0.000000 | 0.000003 | miR-4649-5p | 0.480 | 0.000005 | 0.000057 |
| miR-6715b-5p | 2.724 | 0.000000 | 0.000015 | miR-4746-3p | 0.494 | 0.000027 | 0.000156 |
| miR-4756-5p | 2.641 | 0.000000 | 0.000015 | miR-3960 | 0.496 | 0.000036 | 0.000193 |
| miR-564 | 2.583 | 0.000000 | 0.000000 | miR-6820-5p | 0.497 | 0.000007 | 0.000066 |
| miR-491-5p | 2.572 | 0.000006 | 0.000063 | miR-6789-5p | 0.499 | 0.000009 | 0.000078 |
| miR-125a-3p | 2.463 | 0.000009 | 0.000076 | ||||
| miR-6893-5p | 2.438 | 0.000009 | 0.000078 | ||||
| miR-575 | 2.431 | 0.000000 | 0.000003 | ||||
| miR-433-5p | 2.395 | 0.000002 | 0.000036 | ||||
| miR-378e | 2.392 | 0.000000 | 0.000011 | ||||
| miR-4299 | 2.379 | 0.000000 | 0.000015 | ||||
| miR-6816-3p | 2.350 | 0.000003 | 0.000040 | ||||
| miR-4531 | 2.183 | 0.000004 | 0.000049 | ||||
| miR-3714 | 2.094 | 0.000007 | 0.000066 | ||||
| miR-4723-5p | 2.072 | 0.000000 | 0.000014 | ||||
| miR-3687 | 2.071 | 0.000008 | 0.000073 | ||||
| miR-7977 | 2.014 | 0.000000 | 0.000015 | ||||
| Cell | Exosome | ||||||
|---|---|---|---|---|---|---|---|
| miRNA | Chromosomal | FC | p Value | FDR | FC | p Value | FDR |
| Location | (Treated/ Untreated) | (Treated/ Untreated) | |||||
| miR-494-3p | 14q32.31 | 0.664 | 0.000687 | 0.002067 | 6.335 | 0.000000 | 0.000014 |
| miR-3714 | 3p24.3 | 2.682 | 0.000613 | 0.001906 | 2.094 | 0.000007 | 0.000066 |
| miR-4327 | 21q22.11 | 3.460 | 0.000038 | 0.000247 | 0.519 | 0.000001 | 0.000019 |
| miR-8073 | 13q34 | 1.940 | 0.002027 | 0.004938 | 1.576 | 0.000033 | 0.000185 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takuma, K.; Fujihara, S.; Fujita, K.; Iwama, H.; Nakahara, M.; Oura, K.; Tadokoro, T.; Mimura, S.; Tani, J.; Shi, T.; et al. Antitumor Effect of Regorafenib on MicroRNA Expression in Hepatocellular Carcinoma Cell Lines. Int. J. Mol. Sci. 2022, 23, 1667. https://doi.org/10.3390/ijms23031667
Takuma K, Fujihara S, Fujita K, Iwama H, Nakahara M, Oura K, Tadokoro T, Mimura S, Tani J, Shi T, et al. Antitumor Effect of Regorafenib on MicroRNA Expression in Hepatocellular Carcinoma Cell Lines. International Journal of Molecular Sciences. 2022; 23(3):1667. https://doi.org/10.3390/ijms23031667
Chicago/Turabian StyleTakuma, Kei, Shintaro Fujihara, Koji Fujita, Hisakazu Iwama, Mai Nakahara, Kyoko Oura, Tomoko Tadokoro, Shima Mimura, Joji Tani, Tingting Shi, and et al. 2022. "Antitumor Effect of Regorafenib on MicroRNA Expression in Hepatocellular Carcinoma Cell Lines" International Journal of Molecular Sciences 23, no. 3: 1667. https://doi.org/10.3390/ijms23031667