
Phytomedicines to Target Hepatitis B Virus DNA Replication: Current Limitations and Future Approaches


 , , , and
, , , and 
Abstract
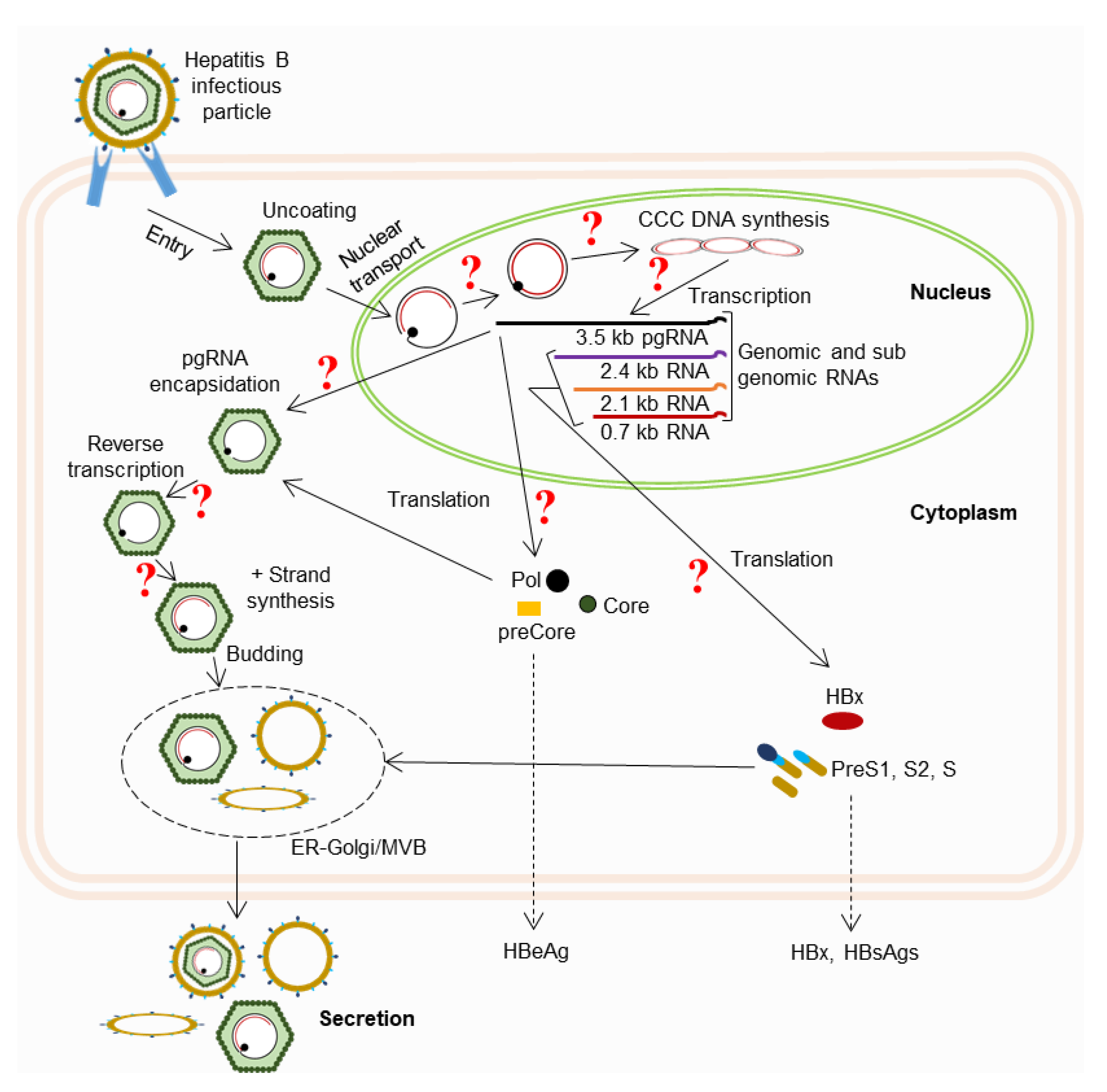
:1. Introduction
2. Phytomedicines and Phytochemicals Regulating HBV DNA Replication
2.1. Terpenoid Compounds
2.2. Flavonoid Compounds
2.3. Phenolic and Polyphenolic Compounds
2.4. Enyne Compounds
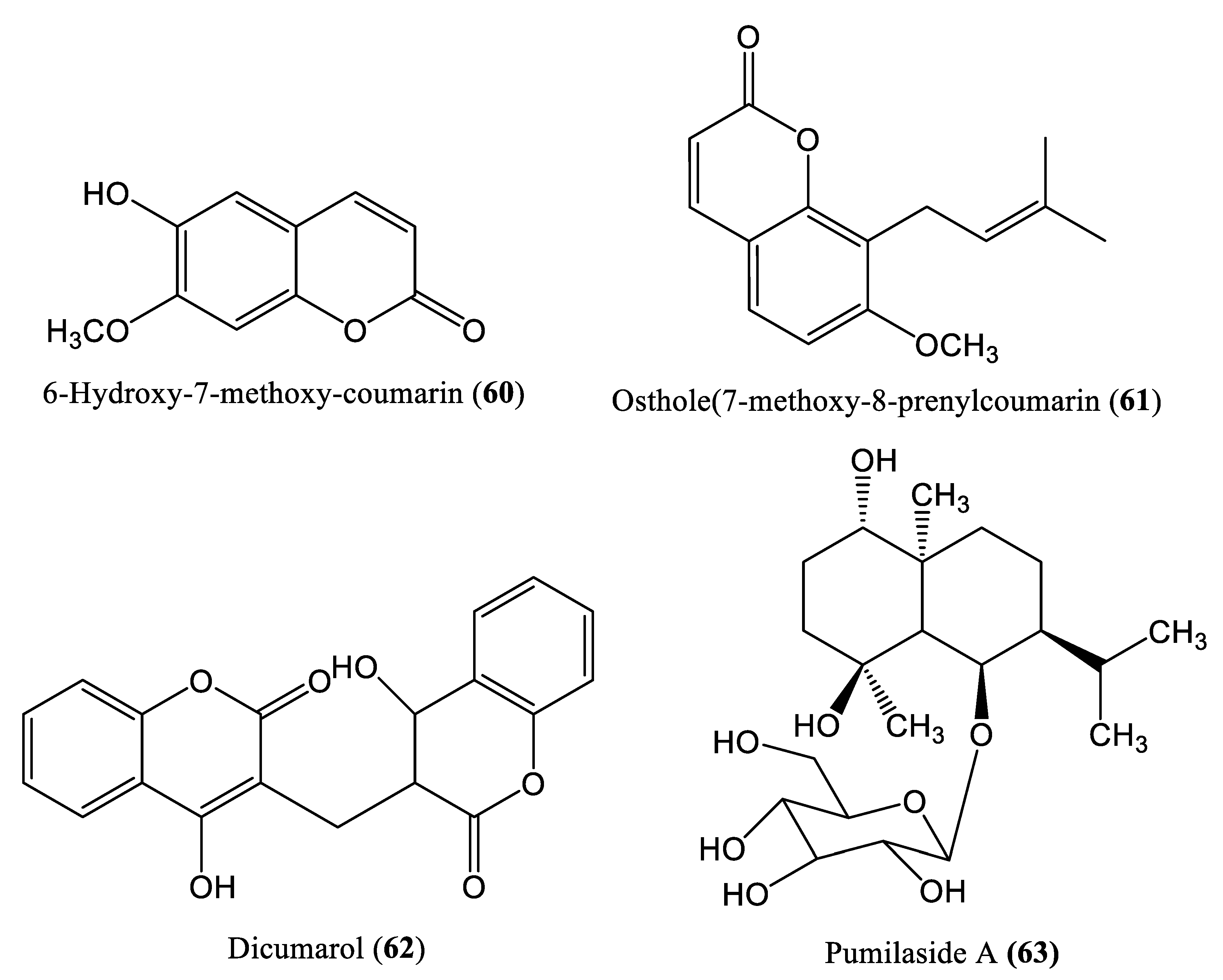
2.5. Lactone Compounds
2.6. Lignan Compounds
2.7. Xanthone Compounds
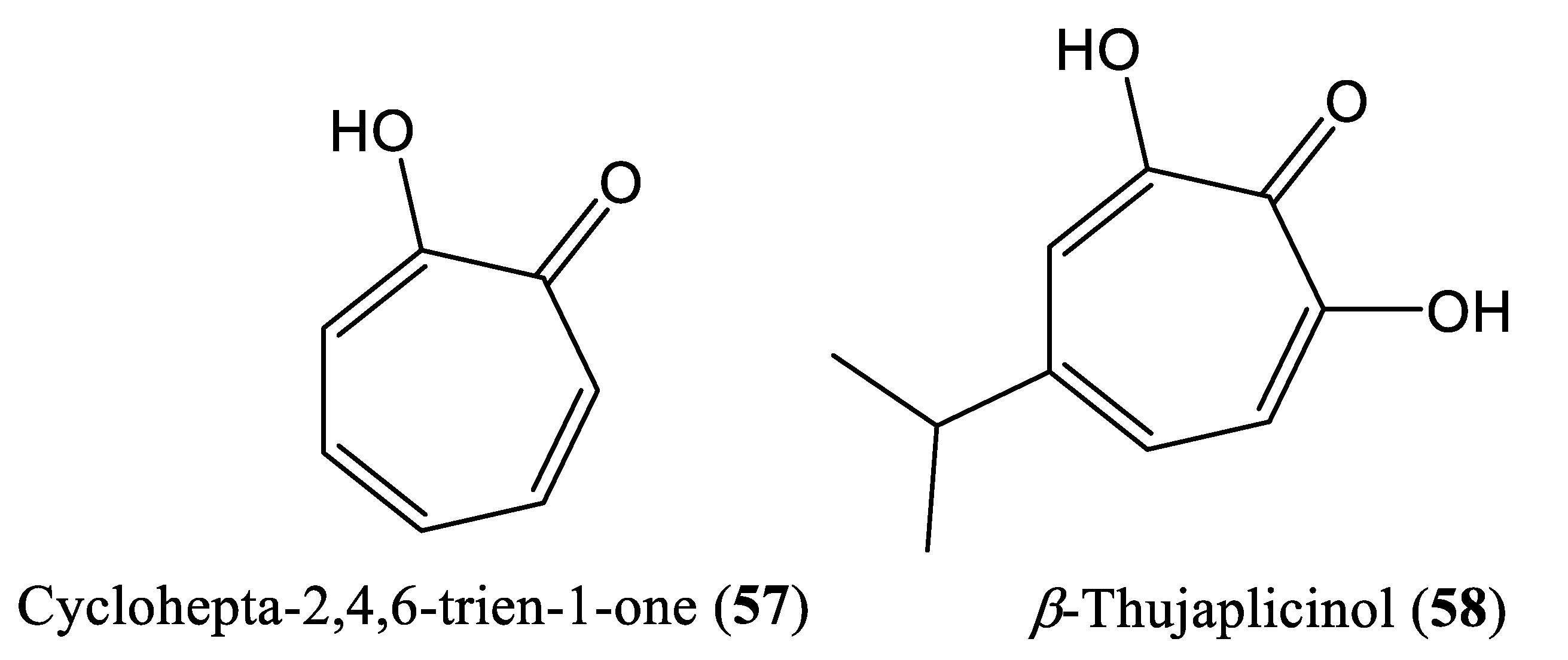
2.8. Tropolone Compounds
2.9. Polysaccharide Compounds
2.10. Others
3. Limitations of Previous Studies
4. Future Approaches
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Toy, M.; Hutton, D.; McCulloch, K.; Romero, N.; Revill, P.A.; Penicaud, M.C.; So, S.; Cowie, B.C. The price tag of a potential cure for chronic hepatitis B infection: A cost threshold analysis for USA, China and Australia. Liver Int. 2022, 42, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.-L.; Li, B.; Zhou, Y.-S.; Zhang, X.; Li, L.; Song, B.; Liu, P.; Yuan, Y.; Zhao, Z.-P.; Jiao, J.; et al. Chinese Military Medical Experts Group in Sierra, L., Prevalence and associated knowledge of hepatitis B infection among healthcare workers in Freetown, Sierra Leone. BMC Infect. Dis. 2018, 18, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guvenir, M.; Arikan, A. Hepatitis B Virus: From Diagnosis to Treatment. Pol. J. Microbiol. 2020, 69, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.G.; Akter, S.; Ohsaki, E.; Ueda, K. Impact of the Interaction of Hepatitis B Virus with Mitochondria and Associated Proteins. Viruses 2020, 12, 175. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.J. Hepatitis B: The virus and disease. Hepatology 2009, 49 (Suppl. 5), S13–S21. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.G.; Suwanmanee, Y.; Du, K.; Ueda, K. Analysis of the Physicochemical Properties, Replication and Pathophysiology of a Massively Glycosylated Hepatitis B Virus HBsAg Escape Mutant. Viruses 2021, 13, 2328. [Google Scholar] [CrossRef]
- Warner, N.; Locarnini, S. Mechanisms of hepatitis B virus resistance development. Intervirology 2014, 57, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-M.; Yau, T.O.; Yu, J. Management of chronic hepatitis B infection: Current treatment guidelines, challenges, and new developments. World J. Gastroenterol. 2014, 20, 6262–6278. [Google Scholar] [CrossRef]
- Wang, G.-F.; Shi, L.-P.; Zuo, J.-P. Anti-hepatitis B virus drugs in clinical and preclinical development. Virol. Sin. 2008, 23, 137–145. [Google Scholar] [CrossRef]
- Woo, A.S.J.; Kwok, R.; Ahmed, T. Alpha-interferon treatment in hepatitis B. Ann. Transl. Med. 2017, 5, 159. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhu, Y.; Feng, S.; Ishida, Y.; Chiu, T.-P.; Saito, T.; Wang, S.; Ann, D.K.; Ou, J.-H.J. Macrophages activated by hepatitis B virus have distinct metabolic profiles and suppress the virus via IL-1β to downregulate PPARα and FOXO3. Cell Rep. 2022, 38, 110284. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.P.; Easterbrook, P.J.; McMahon, B.J. Epidemiology of Hepatitis B Virus Infection and Impact of Vaccination on Disease. Clin. Liver Dis. 2016, 20, 607–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrodomenico, M.; Muselli, M.; Provvidenti, L.; Scatigna, M.; Bianchi, S.; Fabiani, L. Long-term immune protection against HBV: Associated factors and determinants. Hum. Vaccin. Immunother. 2021, 17, 2268–2272. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.G.; Mahmud, M.M.; Nazir, K.H.M.N.H.; Ueda, K. PreS1 Mutations Alter the Large HBsAg Antigenicity of a Hepatitis B Virus Strain Isolated in Bangladesh. Int. J. Mol. Sci. 2020, 21, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.G.; Mahmud, M.M.; Rahman, M.A.; Akter, S.; Nazir, K.; Saha, S.; Wada, M.; Ohsaki, E.; Honda, T.; Ueda, K. Complete Genome Sequence of a Precore-Defective Hepatitis B Virus Genotype D2 Strain Isolated in Bangladesh. Microbiol. Resour. Announc. 2020, 9, e00083-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.G.; Ueda, K. A meta-analysis on genetic variability of RT/HBsAg overlapping region of hepatitis B virus (HBV) isolates of Bangladesh. Infect. Agents Cancer 2019, 14, 33. [Google Scholar] [CrossRef] [Green Version]
- Ko, K.; Takahashi, K.; Nagashima, S.; Yamamoto, C.; Ork, V.; Sugiyama, A.; Akita, T.; Ohisa, M.; Chuon, C.; Hossain, M.S.; et al. Existence of hepatitis B virus surface protein mutations and other variants: Demand for hepatitis B infection control in Cambodia. BMC Infect. Dis. 2020, 20, 305. [Google Scholar] [CrossRef] [Green Version]
- Rapti, I.; Hadziyannis, S. Risk for hepatocellular carcinoma in the course of chronic hepatitis B virus infection and the protective effect of therapy with nucleos(t)ide analogues. World J. Hepatol. 2015, 7, 1064–1073. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Wei, M.; Zhang, C.; Xu, T.; Liu, L.; Xu, Z. Potential resistant mutations within HBV reverse transcriptase sequences in nucleos(t)ide analogues-experienced patients with hepatitis B virus infection. Sci. Rep. 2019, 9, 8078. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.Y.; Lee, C.H. Epidemiology and prevention of hepatitis B virus infection. Korean J. Hepatol. 2011, 17, 87–95. [Google Scholar] [CrossRef]
- Chang, M.S.; Nguyen, M.H. Epidemiology of hepatitis B and the role of vaccination. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Nsibirwa, S.; Anguzu, G.; Kamukama, S.; Ocama, P.; Nankya-Mutyoba, J. Herbal medicine use among patients with viral and non-viral Hepatitis in Uganda: Prevalence, patterns and related factors. BMC Complementary Med. Ther. 2020, 20, 169. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S. Introductory chapter: Terpenes and terpenoids. In Terpenes and Terpenoids; Books on Demand: Schleswig-Holstein, Germany, 2018. [Google Scholar]
- Zhao, Y.L.; Cai, G.M.; Hong, X.; Shan, L.M.; Xiao, X.H. Anti-hepatitis B virus activities of triterpenoid saponin compound from Potentilla anserine L. Phytomed. Int. J. Phytother. Phytopharm. 2008, 15, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Li, Z.; Gong, X.; Fu, X.; Xiao, X.; He, M.; Huang, B.; Xu, Z. Total saponins extracted from Abrus cantoniensis Hance suppress hepatitis B virus replication in vitro and in rAAV8-1.3HBV transfected mice. J. Ethnopharmacol. 2020, 249, 112366. [Google Scholar] [CrossRef]
- Li, Y.Q.; Wang, S.F. Anti-hepatitis B activities of ganoderic acid from Ganoderma lucidum. Biotechnol. Lett. 2006, 28, 837–841. [Google Scholar] [CrossRef]
- Zhou, W.B.; Zeng, G.Z.; Xu, H.M.; He, W.J.; Tan, N.H. Astataricusones A-D and astataricusol A, five new anti-HBV shionane-type triterpenes from Aster tataricus L. f. Mol. Basel Switz. 2013, 18, 14585–14596. [Google Scholar] [CrossRef]
- Guo, R.H.; Geng, C.A.; Huang, X.Y.; Ma, Y.B.; Zhang, Q.; Wang, L.J.; Zhang, X.M.; Zhang, R.P.; Chen, J.J. Synthesis of hemslecin A derivatives: A new class of hepatitis B virus inhibitors. Bioorganic. Med. Chem. Lett. 2013, 23, 1201–1205. [Google Scholar] [CrossRef]
- Wang, L.-J.; Geng, C.-A.; Ma, Y.-B.; Luo, J.; Huang, X.-Y.; Chen, H.; Zhou, N.-J.; Zhang, X.-M.; Chen, J.-J. Design, synthesis, and molecular hybrids of caudatin and cinnamic acids as novel anti-hepatitis B virus agents. Eur. J. Med. Chem. 2012, 54, 352–365. [Google Scholar] [CrossRef]
- Wang, L.J.; Geng, C.A.; Ma, Y.B.; Huang, X.Y.; Luo, J.; Chen, H.; Zhang, X.M.; Chen, J.J. Synthesis, biological evaluation and structure-activity relationships of glycyrrhetinic acid derivatives as novel anti-hepatitis B virus agents. Bioorganic. Med. Chem. Lett. 2012, 22, 3473–3479. [Google Scholar] [CrossRef]
- Tseng, Y.P.; Kuo, Y.H.; Hu, C.P.; Jeng, K.S.; Janmanchi, D.; Lin, C.H.; Chou, C.K.; Yeh, S.F. The role of helioxanthin in inhibiting human hepatitis B viral replication and gene expression by interfering with the host transcriptional machinery of viral promoters. Antivir. Res. 2008, 77, 206–214. [Google Scholar] [CrossRef]
- Huang, Q.; Zhang, S.; Huang, R.; Wei, L.; Chen, Y.; Lv, S.; Liang, C.; Tan, S.; Liang, S.; Zhuo, L.; et al. Isolation and identification of an anti-hepatitis B virus compound from Hydrocotyle sibthorpioides Lam. J. Ethnopharmacol. 2013, 150, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Huang, R.; Wei, L.; Chen, Y.; Lv, S.; Liang, C.; Zhang, X.; Yin, F.; Li, H.; Zhuo, L.; et al. Antiviral activity of methyl helicterate isolated from Helicteres angustifolia (Sterculiaceae) against hepatitis B virus. Antivir. Res. 2013, 100, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Oriakhi, K.; Uadia, P.O.; Shaheen, F.; Jahan, H.; Ibeji, C.U.; Iqbal, C.M. Isolation, characterization, and hepatoprotective properties of betulinic acid and ricinine from Tetracarpidium conophorum seeds (Euphorbiaceae). J. Food Biochem. 2021, 45, e13288. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, P.; Zhang, S.; Zhao, L.; Dong, J.; Jie, S.; Pang, R.; Li, S. Tea polyphenols exerts anti-hepatitis B virus effects in a stably HBV-transfected cell line. J. Huazhong Univ. Sci. Technology. Med. Sci. 2009, 29, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-M. Anti-infective potential of catechins and their derivatives against viral hepatitis. Clin. Exp. Vaccine Res. 2018, 7, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.W.; Geng, C.A.; Jiang, F.Q.; Ma, Y.B.; He, K.; Zhou, N.J.; Zhang, X.M.; Zhou, J.; Chen, J.J. Chemical constituents of Swertia yunnanensis and their anti-hepatitis B virus activity. Fitoterapia 2013, 89, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.J.; Tsai, Y.C.; Chiang, S.Y.; Wang, G.J.; Kuo, Y.C.; Chang, Y.C.; Wu, Y.Y.; Wu, Y.C. Anti-viral effect of a compound isolated from Liriope platyphylla against hepatitis B virus in vitro. Virus Res. 2014, 192, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.W.; Geng, C.A.; Ma, Y.B.; Zhang, X.M.; Zhou, J.; Tao, Y.D.; Chen, J.J. Chemical constituents of Swertia mussotii and their anti-hepatitis B virus activity. Fitoterapia 2015, 102, 15–22. [Google Scholar] [CrossRef]
- Zembower, D.E.; Lin, Y.M.; Flavin, M.T.; Chen, F.C.; Korba, B.E. Robustaflavone, a potential non-nucleoside anti-hepatitis B agent. Antivir. Res. 1998, 39, 81–88. [Google Scholar] [CrossRef]
- Huang, R.L.; Chen, C.C.; Huang, H.L.; Chang, C.G.; Chen, C.F.; Chang, C.; Hsieh, M.T. Anti-hepatitis B virus effects of wogonin isolated from Scutellaria baicalensis. Planta Med. 2000, 66, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Huang, H.; Feng, M.; Zhou, W.; Shi, X.; Zhou, P. In Vitro and in vivo anti-hepatitis B virus activities of a plant extract from Geranium carolinianum L. Antivir. Res. 2008, 79, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.Q.; Huang, Z.M.; Yang, X.B.; Liu, H.Z.; Wu, G.X. In Vivo and in vitro anti-hepatitis B virus activity of total phenolics from Oenanthe javanica. J. Ethnopharmacol. 2008, 118, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kim, Y.H.; Lee, K.R. Isolation of quinic acid derivatives and flavonoids from the aerial parts of Lactuca indica L. and their hepatoprotective activity in vitro. Bioorganic Med. Chem. Lett. 2007, 17, 6739–6743. [Google Scholar] [CrossRef]
- Wang, G.F.; Shi, L.P.; Ren, Y.D.; Liu, Q.F.; Liu, H.F.; Zhang, R.J.; Li, Z.; Zhu, F.H.; He, P.L.; Tang, W.; et al. Anti-hepatitis B virus activity of chlorogenic acid, quinic acid and caffeic acid in vivo and in vitro. Antivir. Res. 2009, 83, 186–190. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Y.; Ding, X.R.; Chen, S.H.; Yang, J.; Wang, X.J.; Jia, G.L.; Chen, H.S.; Bo, X.C.; Wang, S.Q. Protocatechuic aldehyde inhibits hepatitis B virus replication both in vitro and in vivo. Antivir. Res. 2007, 74, 59–64. [Google Scholar] [CrossRef]
- Geng, C.A.; Ma, Y.B.; Zhang, X.M.; Yao, S.Y.; Xue, D.Q.; Zhang, R.P.; Chen, J.J. Mulberrofuran G and isomulberrofuran G from Morus alba L.: Anti-hepatitis B virus activity and mass spectrometric fragmentation. J. Agric. Food Chem. 2012, 60, 8197–8202. [Google Scholar] [CrossRef]
- Zhao, Y.; Geng, C.A.; Chen, H.; Ma, Y.B.; Huang, X.Y.; Cao, T.W.; He, K.; Wang, H.; Zhang, X.M.; Chen, J.J. Isolation, synthesis and anti-hepatitis B virus evaluation of p-hydroxyacetophenone derivatives from Artemisia capillaris. Bioorganic Med. Chem. Lett. 2015, 25, 1509–1514. [Google Scholar] [CrossRef]
- Geng, C.-A.; Yang, T.-H.; Huang, X.-Y.; Yang, J.; Ma, Y.-B.; Li, T.-Z.; Zhang, X.-M.; Chen, J.-J. Anti-hepatitis B virus effects of the traditional Chinese herb Artemisia capillaris and its active enynes. J. Ethnopharmacol. 2018, 224, 283–289. [Google Scholar] [CrossRef]
- Labuda, I. Flavor Compounds. In Encyclopedia of Microbiology, 3rd ed.; Schaechter, M., Ed.; Academic Press: Oxford, UK, 2009; pp. 305–320. [Google Scholar]
- Chen, H.; Ma, Y.B.; Huang, X.Y.; Geng, C.A.; Zhao, Y.; Wang, L.J.; Guo, R.H.; Liang, W.J.; Zhang, X.M.; Chen, J.J. Synthesis, structure-activity relationships and biological evaluation of dehydroandrographolide and andrographolide derivatives as novel anti-hepatitis B virus agents. Bioorganic Med. Chem. Lett. 2014, 24, 2353–2359. [Google Scholar] [CrossRef]
- Romero, M.R.; Efferth, T.; Serrano, M.A.; Castaño, B.; Macias, R.I.; Briz, O.; Marin, J.J. Effect of artemisinin/artesunate as inhibitors of hepatitis B virus production in an "in vitro" replicative system. Antivir. Res. 2005, 68, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.A.; Wang, L.J.; Zhang, X.M.; Ma, Y.B.; Huang, X.Y.; Luo, J.; Guo, R.H.; Zhou, J.; Shen, Y.; Zuo, A.X.; et al. Anti-hepatitis B virus active lactones from the traditional Chinese herb: Swertia mileensis. Chem. Weinh. Der Bergstr. Ger. 2011, 17, 3893–3903. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-L.; Cao, T.-W.; Jiang, F.-Q.; Geng, C.-A.; Zhang, X.-M.; Huang, X.-Y.; Wang, L.-J.; Kang, H.; Hao, C.; Liang, W.-J.; et al. Swerpunilactones A and B, the first example of xanthone and secoiridoid heterodimers from Swertia punicea, S. hispidicalyx, and S. yunnanensis. Tetrahedron Lett. 2013, 54, 2710–2712. [Google Scholar] [CrossRef]
- Zhang, M.; Deng, Y.; Zhang, H.B.; Su, X.L.; Chen, H.L.; Yu, T.; Guo, P. Two new coumarins from Herpetospermum caudigerum. Chem. Pharm. Bull. 2008, 56, 192–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.J.; Liu, Q.C.; Zhang, T.; Liu, X.; Wang, L.Q.; Yuan, H.L. Preparation of herpetolide A nanosuspension lyophilized powder and evaluation of its anti-hepatitis B virus activity. China J. Chin. Mater. Med. 2020, 45, 1076–1081. [Google Scholar]
- Xu, H.Y.; Ren, J.H.; Su, Y.; Ren, F.; Zhou, Y.J.; Jiang, H.; Cheng, S.T.; Zhang, C.R.; Chen, J. Anti-hepatitis B virus activity of swertisin isolated from Iris tectorum Maxim. J. Ethnopharmacol. 2020, 257, 112787. [Google Scholar] [CrossRef] [PubMed]
- Bekhit, A.E.D.; Bekhit, A.A. Chapter 7-natural antiviral compounds. In Studies in Natural Products Chemistry; Attaur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 42, pp. 195–228. [Google Scholar]
- Li, J.; Meng, A.P.; Guan, X.L.; Li, J.; Wu, Q.; Deng, S.P.; Su, X.J.; Yang, R.Y. Anti-hepatitis B virus lignans from the root of Streblus asper. Bioorganic Med. Chem. Lett. 2013, 23, 2238–2244. [Google Scholar] [CrossRef]
- Li, J.; Huang, Y.; Guan, X.L.; Li, J.; Deng, S.P.; Wu, Q.; Zhang, Y.J.; Su, X.J.; Yang, R.Y. Anti-hepatitis B virus constituents from the stem bark of Streblus asper. Phytochemistry 2012, 82, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.J.; Geng, C.A.; Huang, X.Y.; Ma, Y.B.; Zhang, X.M.; Wang, J.L.; Chen, J.J. Anti-hepatitis B virus active constituents from Swertia chirayita. Fitoterapia 2015, 100, 27–34. [Google Scholar] [CrossRef]
- Ares, A.M.; Bernal, J.L.; Nozal, M.J.; Bernal, J. Chapter 6-analysis of herbal bioactives. In Aromatic Herbs in Food; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 201–232. [Google Scholar]
- Tutik Sri, W.; Adita Ayu, P.; Aty, W.; Hak, H.; Chie, A.-U.; Achmad Fuad, H. Antiviral Activity of Indonesian Medicinal Plants against Hepatitis B Virus. Pharmacogn. J. 2020, 12, 157. [Google Scholar] [CrossRef]
- Liu, X.; Ma, C.; Liu, Z.; Kang, W. Natural Products: Review for Their Effects of Anti-HBV. BioMed Res. Int. 2020, 2020, 3972390. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.W.; Geng, C.A.; Ma, Y.B.; He, K.; Wang, H.L.; Zhou, N.J.; Zhang, X.M.; Tao, Y.D.; Chen, J.J. Xanthones with anti-hepatitis B virus activity from Swertia mussotii. Planta Med. 2013, 79, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.W.; Geng, C.A.; Ma, Y.B.; He, K.; Zhou, N.J.; Zhou, J.; Zhang, X.M.; Chen, J.J. Chemical constituents of Swertia delavayi and their anti-hepatitis B virus activity. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2015, 40, 897–902. [Google Scholar]
- Cox, R.J.; Al-Fahad, A. Chemical mechanisms involved during the biosynthesis of tropolones. Curr. Opin. Chem. Biol. 2013, 17, 532–536. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, X.; Cao, F.; Huang, A.; Tavis, J.E. β-Thujaplicinol inhibits hepatitis B virus replication by blocking the viral ribonuclease H activity. Antivir. Res. 2013, 99, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Mou, J.F.; Lin, X.Z.; Su, H.L.; Lu, H.L.; Liu, Q.B.; Liang, B.; Chen, X.; Liang, C.Q.; Zhou, X.L. Anti-hepatitis B virus activity and hepatoprotective effect of des(rhamnosyl) verbascoside from Lindernia ruellioides in vitro. Phytother. Res. PTR 2021, 35, 4555–4566. [Google Scholar] [CrossRef]
- Liang, S.; Li, X.; Ma, X.; Li, A.; Wang, Y.; Reaney, M.J.T.; Shim, Y.Y. A flaxseed heteropolysaccharide stimulates immune responses and inhibits hepatitis B virus. Int. J. Biol. Macromol. 2019, 136, 230–240. [Google Scholar] [CrossRef]
- Huang, S.X.; Mou, J.F.; Luo, Q.; Mo, Q.H.; Zhou, X.L.; Huang, X.; Xu, Q.; Tan, X.D.; Chen, X.; Liang, C.Q. Anti-Hepatitis B Virus Activity of Esculetin from Microsorium fortunei In Vitro and In Vivo. Mol. Basel Switz. 2019, 24, 3475. [Google Scholar] [CrossRef] [Green Version]
- Indrasetiawan, P.; Aoki-Utsubo, C.; Hanafi, M.; Hartati, S.; Wahyuni, T.S.; Kameoka, M.; Yano, Y.; Hotta, H.; Hayashi, Y. Antiviral Activity of Cananga odorata against Hepatitis B Virus. Kobe J. Med. Sci. 2019, 65, E71–E79. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Ingredient/ Compound | Concentrations and/or IC50 | Mechanisms | Systems | Group | Plants/Sources | References |
|---|---|---|---|---|---|---|
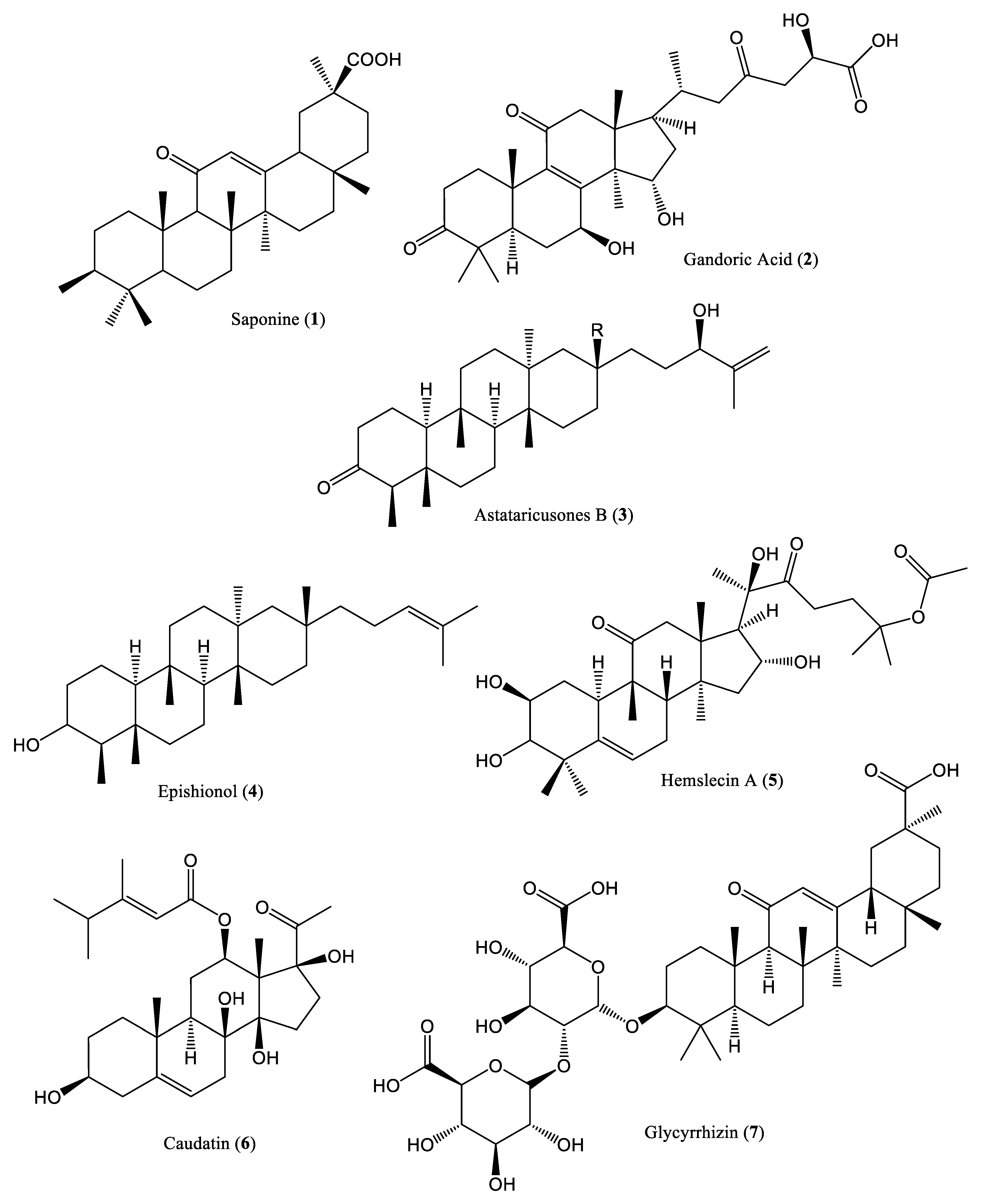
| Saponin | 40 μg/mL | Reduce DNA replication | HepG2.2.15 cells and Peking ducklings | Terpenoids | Potentilla anserina | [24] |
| Saponin | 60 µg/mL | Inhibit DNA production | HepG2.2.15 cells and C57BL/6 mice | Terpenoids | Abrus cantoniensis Hance | [25] |
| Ganoderic acid | 8 µg/mL | Inhibit HBV replication | HepG2.2.15 cells | Terpenoids | Ganoderma lucidum | [26] |
| Astataricusones B | 2.7 μM | Inhibit DNA replication | HepG 2.2.15 cells | Terpenoids | Aster tataricus | [27] |
| Epishionol | 30.7 μM | Inhibit DNA replication | HepG 2.2.15 cells | Terpenoids | Aster tataricus | [27] |
| Hemslecin A | 11.2 μM | Inhibit DNA replication | HepG2.2.15 cells | Terpenoids | Hemsleya | [28] |
| Caudatin | 40.62 mM | Inhibit DNA replication | HepG 2.2.15 cells | Terpenoids | Cynanchum auriculatum | [29] |
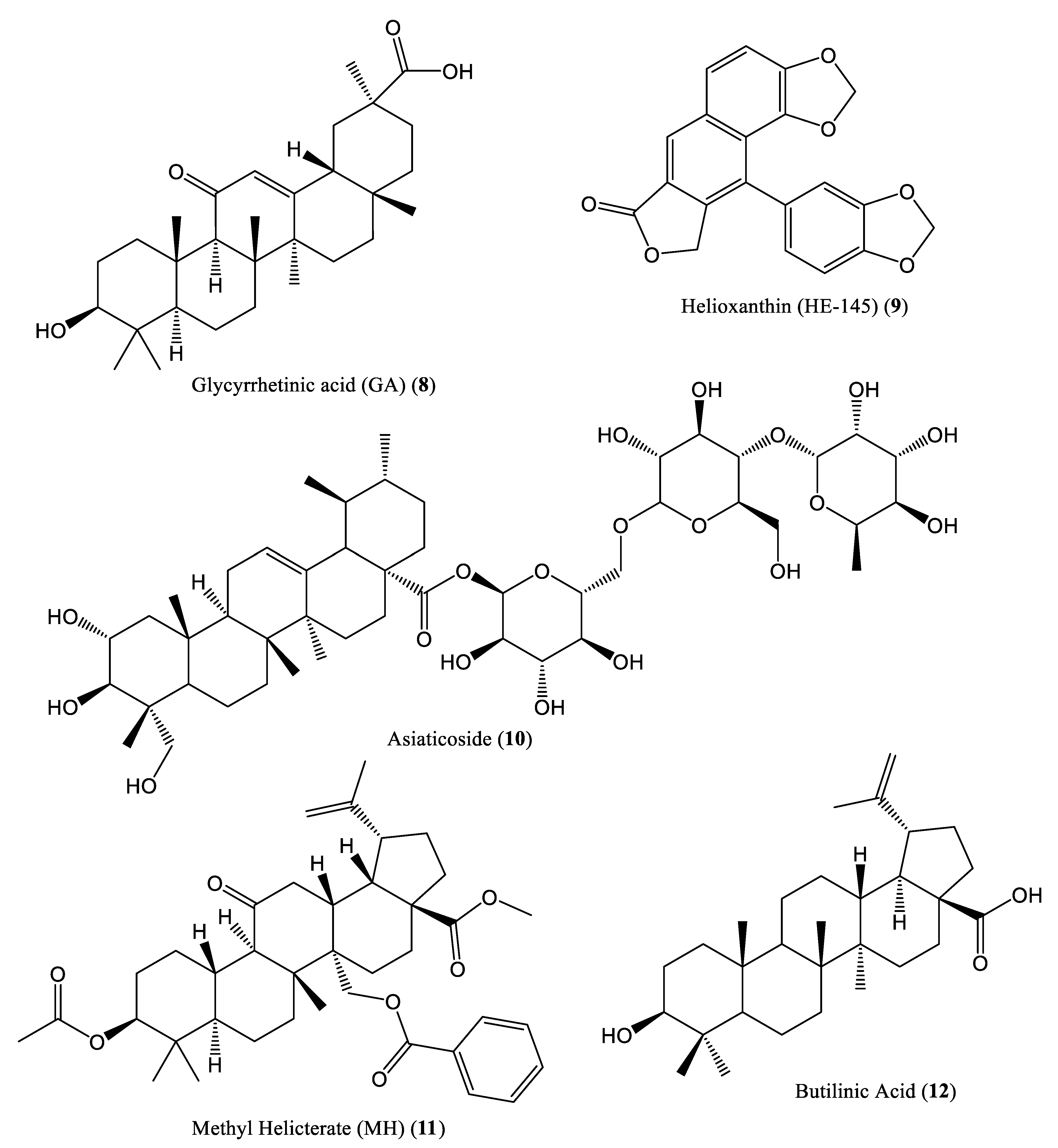
| Glycyrrhizin and glycyrrhetinic acid | 39.28 μM/L | Inhibit DNA replication | HepG 2.2.15 cells | Terpenoids | Glycyrrhizae glabra | [30] |
| Helioxanthin | 5.0 μM | Decrease DNA binding activity | HepA2 cells | Terpenoids | Taiwania cryptomerioides | [31] |
| Asiaticoside | 150.0 μM | Reduce HBV transcription, replication, and cccDNA levels | HepG2.2.15 cells and ducklings | Terpenoids | Hydrocotyle sibthorpioides | [32] |
| Methyl helicterate | 15.8 μM | Reduce cccDNA and RNA levels | HepG2.2.15 cells and ducklings | Terpenoids | Helicteres angustifolia | [33] |
| Betulinic acid | 16.7 μM | Exhibit high binding affinities for the virus | Rat models and HepG2 cells | Terpenoids | T. conophorum | [34] |
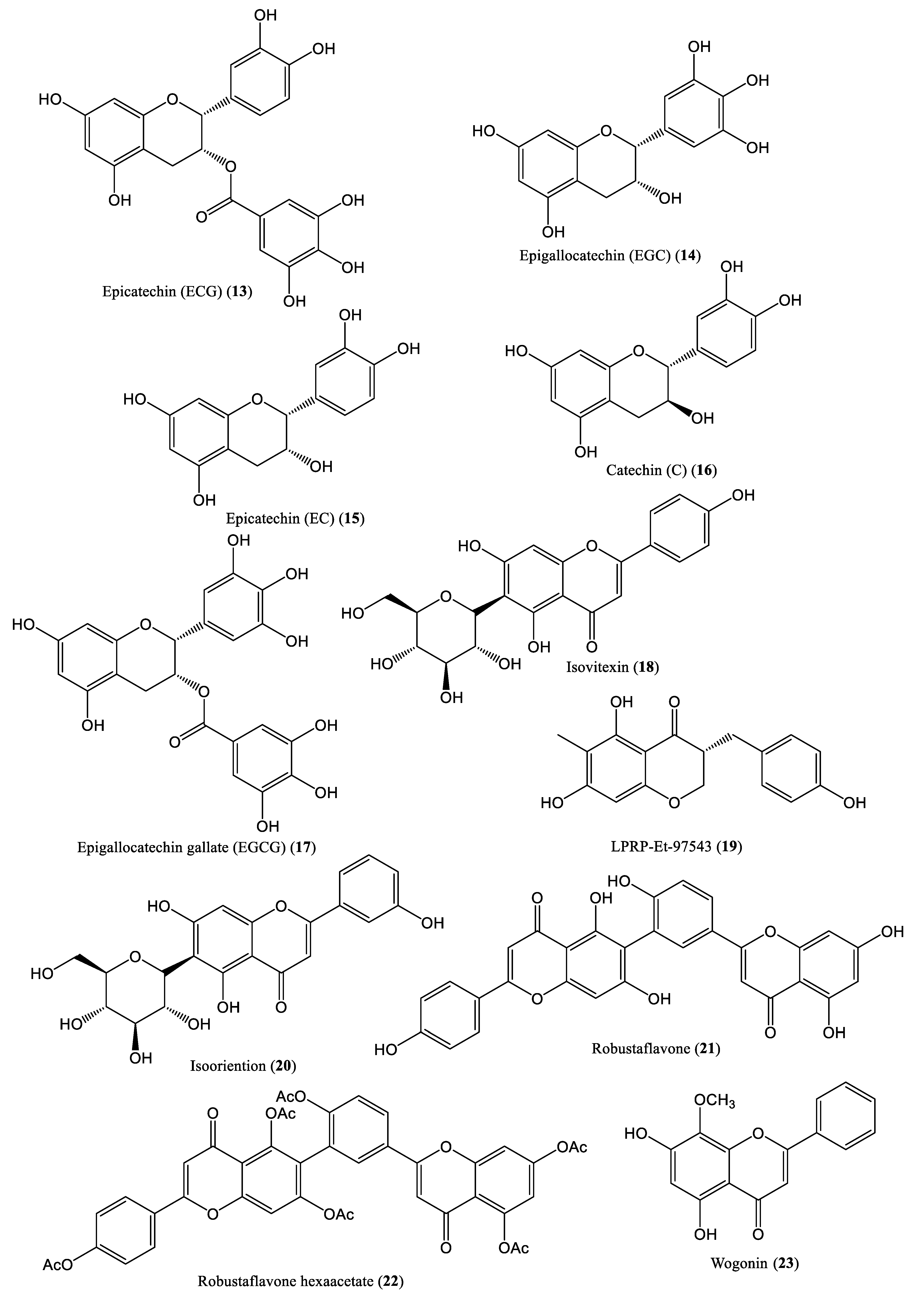
| Epigallocatechin gallate (EGCG) | 7.34 μM | Reduce cccDNA production | HepG2.117 cells | Flavonoids | Camellia sinensis | [36,37] |
| Isovitexin | 0.09 mM | Suppresses DNA replication | HepG 2.2.15 cells | Flavonoids | Swertia yunnanensis | [38] |
| LPRP-Et-97543 | 10 μg/mL | Suppress core, S, and preS promoters and DNA replication | HepG2.2.15 cells and Huh7 cells | Flavonoids | Liriope muscari | [39] |
| Isooriention | 0.02 mM | Inhibit DNA replication | HepG2.2.15 cells | Flavonoids | Swertia mussotii | [40] |
| Robustaflavone | 0.25 mM | Inhibit DNA replication | HepG2.2.15 cells | Flavonoids | Rhus succedanea | [41] |
| Wogonin | 0.73 mM | Reduce relaxed circular and linear forms of DNA | HBV-producing cell line (MS-G2) | Flavonoids | Scutellaria baicalensis | [42] |
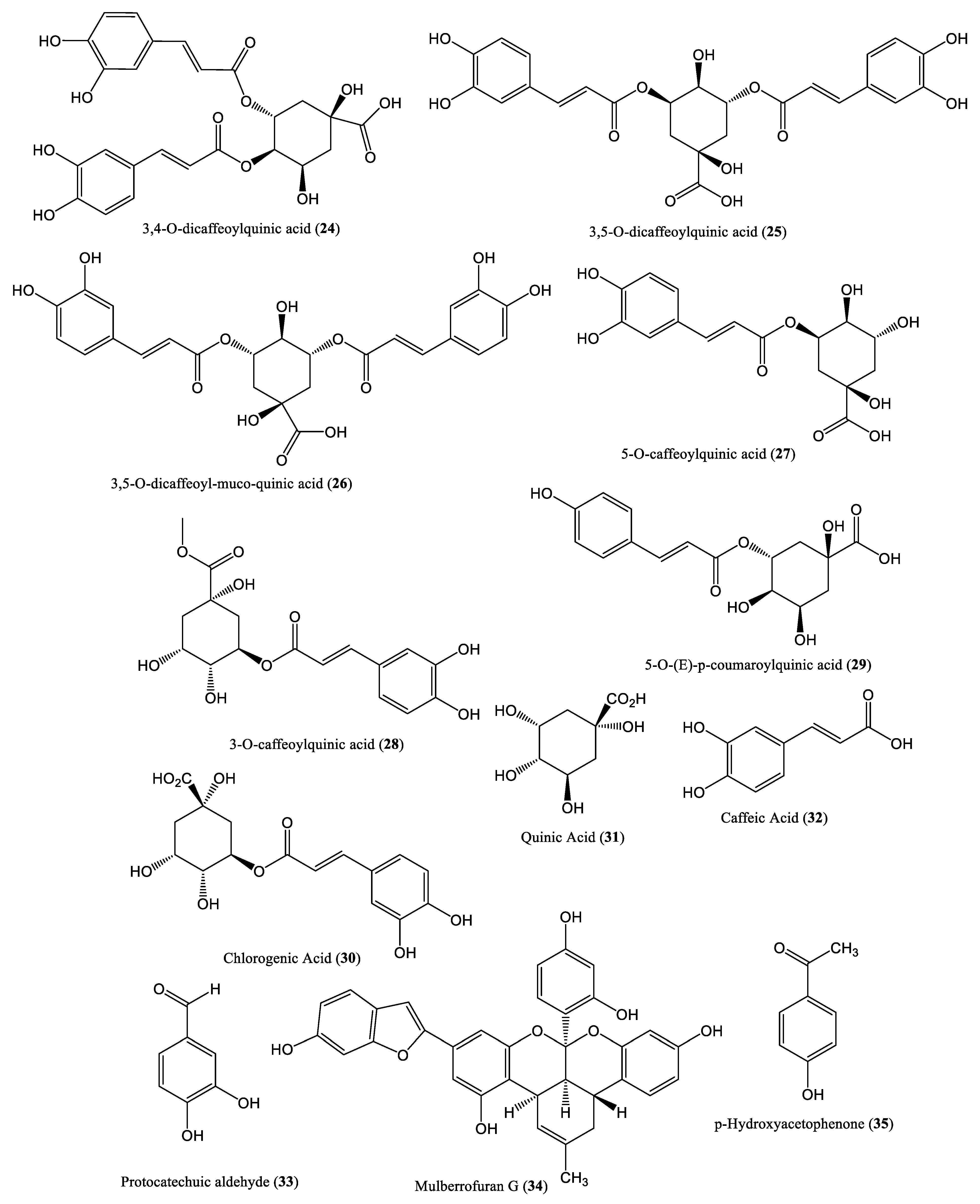
| Polyphenolic extract | 65.60 μg/mL | Reduced DNA level | HepG2.2.15 cells and ducklings | Phenolic and polyphenolic | Geranium carolinianum L. | [44] |
| Phenolic extract | 0.40 g/L | Inhibit DNA level | HepG2.2.15 cells and ducks | Phenolic and polyphenolic | Oenanthe javanica | [45] |
| Quinic acid derivatives | - | Decrease DNA levels | HepG2.2.15 cells | Phenolic and polyphenolic | Lactuca indica L. | [46] |
| Chlorogenic acid | >1000 μM | Reduce single-stranded form of HBV DNA | HepG2.2.15 cells and ducklings | Phenolic and polyphenolic | Dicotyledonous plants | [47] |
| Protocatechuic aldehyde | 100 mg/kg | Reduce DNA release | HepG2.2.15 cells and ducks | Phenolic and polyphenolic | Salvia miltiorrhiza | [48] |
| Mulberrofuran G | 3.99 μM | Inhibit DNA replication | HepG2.2.15 cells | Phenolic and polyphenolic | Morus alba L. | [49] |
| p-Hydroxyacetophenone | 306.4 μM | Inhibit DNA replication | HepG2.2.15 cells | Phenolic and polyphenolic | Artemisia morrisonensis | [50] |
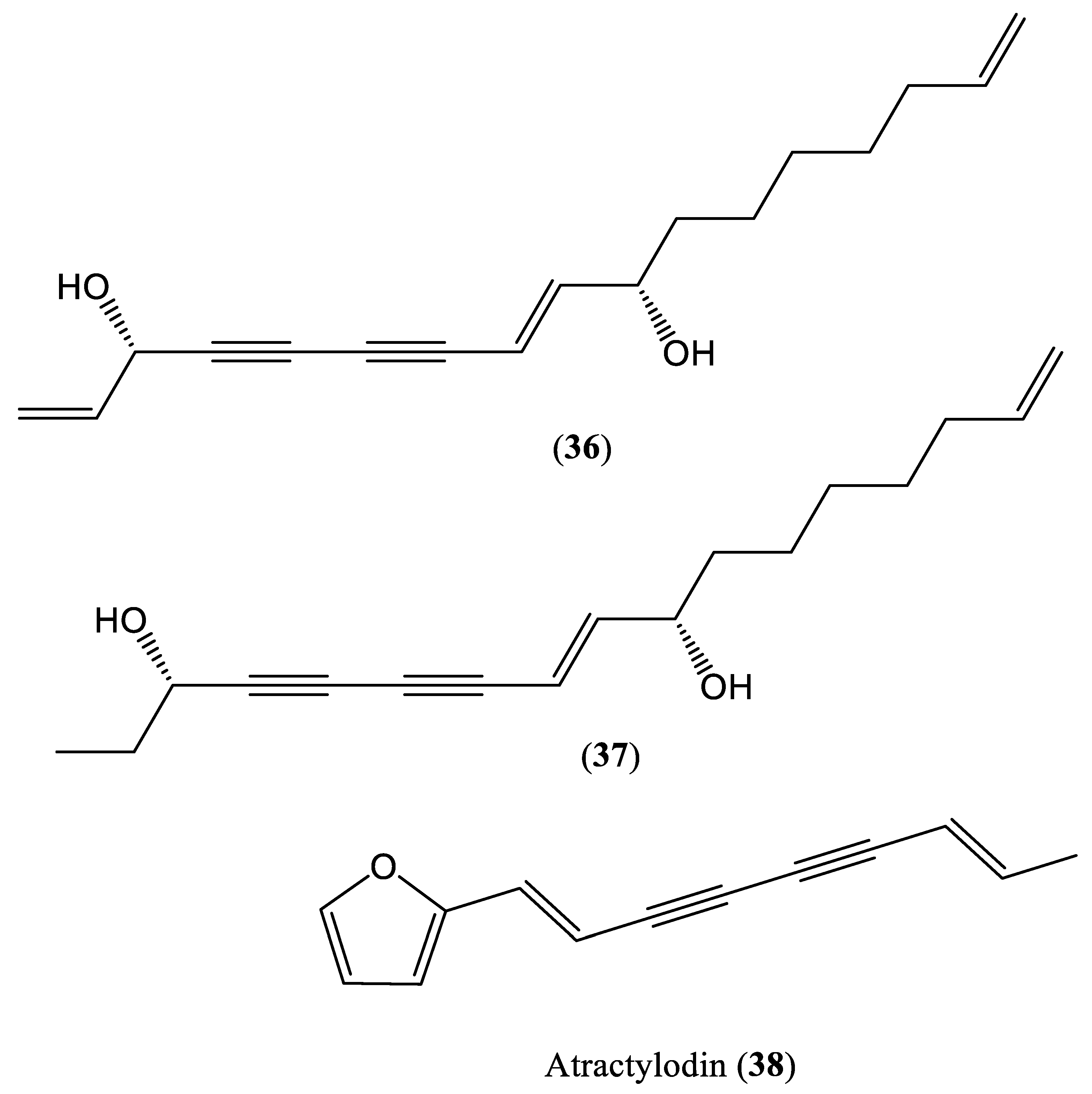
| Atractylodin | 9.8 μM | Inhibit DNA replication | HepG2.2.15 cells | Eneynes | Artemisia capillaris | [51] |
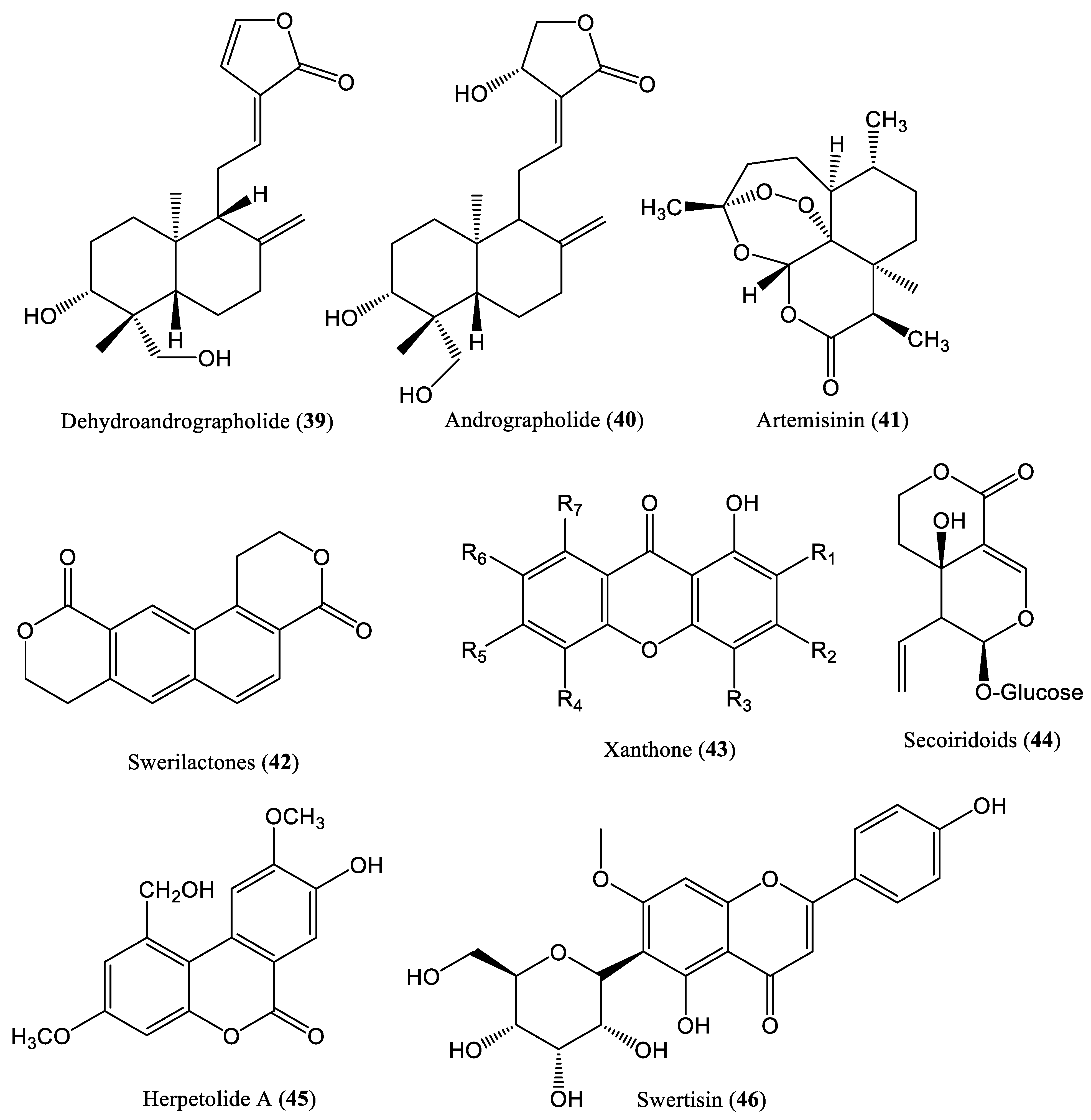
| Dehydroandrographolide and andrographolide | 54.07 μM | Inhibit DNA replication | HepG2.2.15 cells | Lactones | Andrographis paniculata | [53] |
| Artemisinin | >100 mM | Inhibit rcDNA forms | HepG2.2.15 cells | Lactones | Artemisia annua | [54] |
| Swerilactones H-K | 5.34 μM | Inhibit DNA replication | HepG2.2.15 cells | Lactones | Swertia mileensis | [55] |
| Xanthrone and secoiridoid lactone | 0.19 mM | Inhibit DNA replication | HepG2.2.15 cells | Lactones | Swertia punicea | [56] |
| Herpetolide A | 12.5 mg/kg | Decrease DNA levels | HepG2.2.15 cells | Lactones | Herpetospermum caudigerum | [58] |
| Swertisin | 125 μM | Downregulate DNA production | HepG2.2.15 cells and HBV transgenic mice | Lactones | Iris tectorum Maxim | [59] |
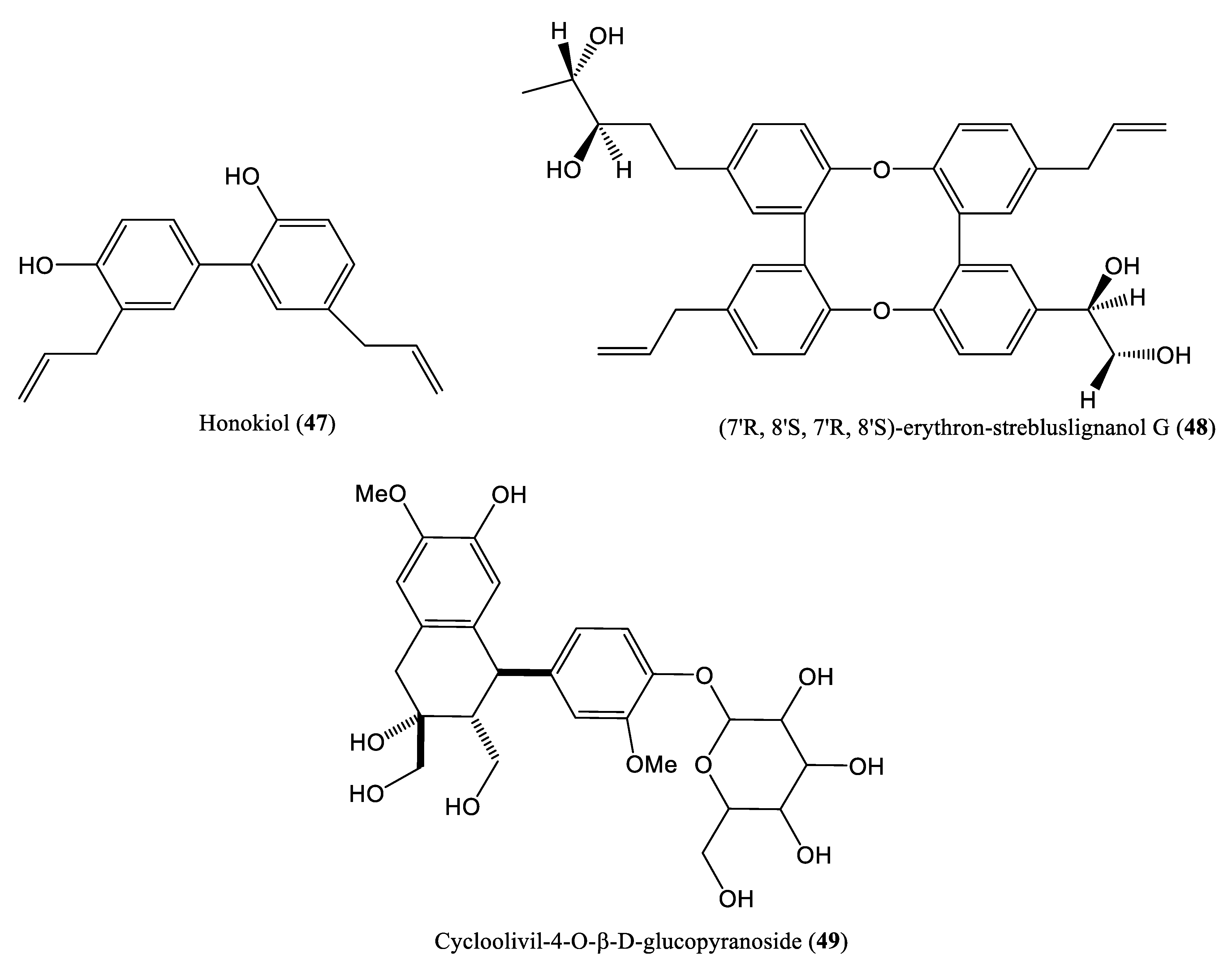
| honokiol and (7′R, 8′S, 7′R, 8′S)-erythron-strebluslignanol G | 8.67 μM | Inhibit DNA replication | HepG2.2.15 cells | Lignans | Streblus asper | [61,62] |
| Cycloolivil-4-O-β-D-glucopyranoside | 0.29 mM | Inhibit DNA replication | HepG2.2.15 cells | Lignans | Stereospermum cylindricum | [63] |
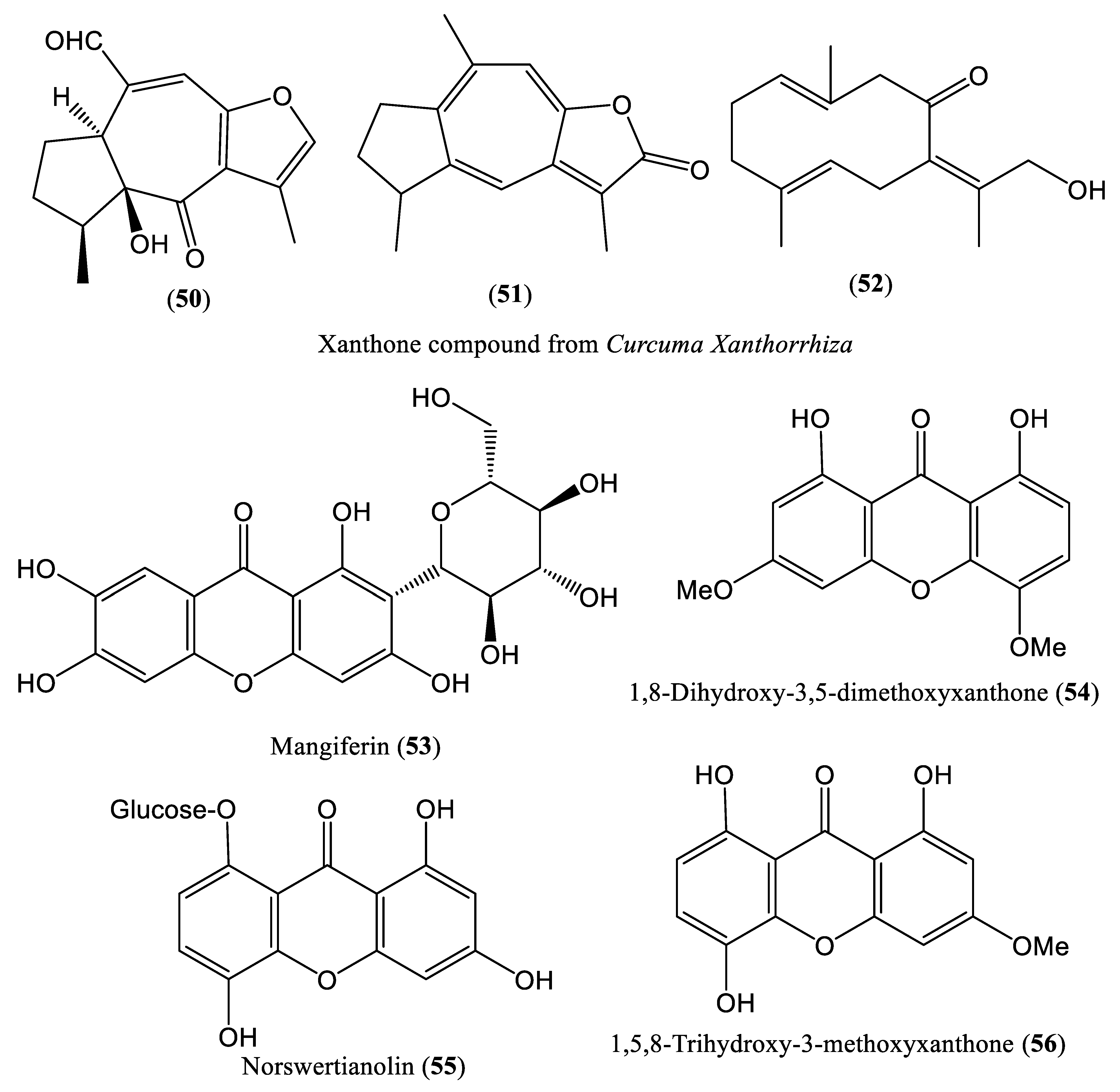
| Extract | 50 μg/mL | Reduce DNA levels | Hep38.7 Tet cells | Xanthones | Curcuma xanthorrhiza | [65] |
| Mangiferin | 0.13 mM | Inhibit DNA replication | HepG2.2.15 cells | Xanthones | Swertia mussotii | [66,67] |
| Dihydroxy-3,5-dimethoxyxanthone and norswertianolin | 0.21 mM | Inhibit DNA replication | HepG2.2.15 cells | Xanthones | Swertia yunnanensis | [38,66] |
| Trihydroxy-3-methoxyxanthone | 0.05 mM | Inhibit DNA replication | HepG2.2.15 cells | Xanthones | Swertia delavayi | [66,68] |
| β-Thujaplicinol | 10.1 mM | Inhibit RNAseH activity | Huh7 cells | Tropolone | Thuja plicata, Thuja occidentalis and Chamaecyparis obtusa | [70] |
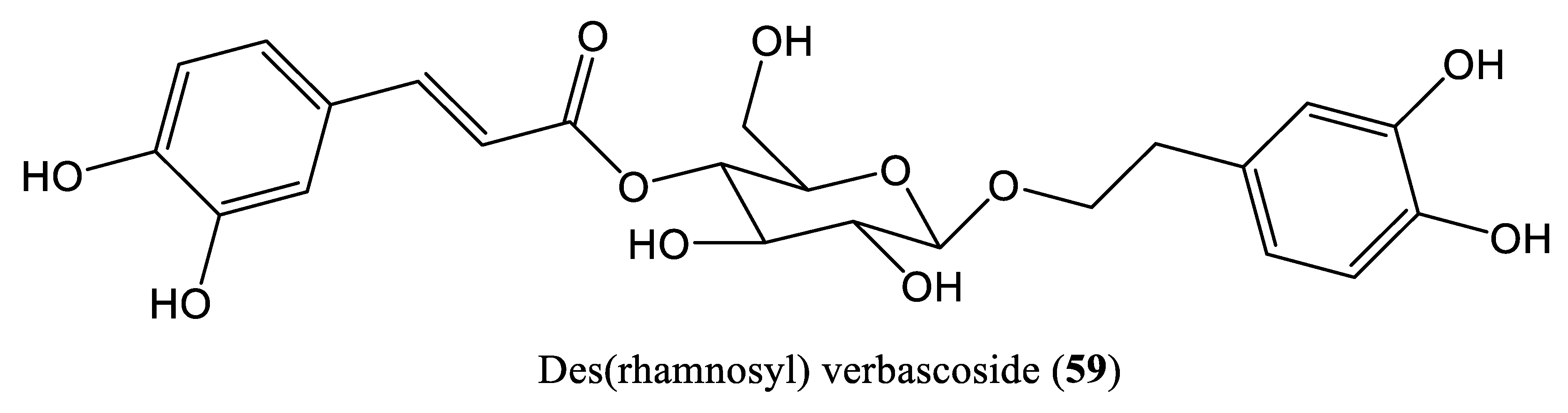
| Des(rhamnosyl) verbascoside | 12.5 mg/L | Downregulate HBx and inhibit DNA replication | HepG2.2.15 cells | Polysaccharide | Lindernia ruellioides | [71] |
| heteropolysaccharide (FP-1) | 250 μg/mL | Inhibit DNA replication | HepG2.2.15 cells | Polysaccharide | Flaxseed hull | [72] |
| Studies | Approaches (Compound and Methods) | Limitations | References |
|---|---|---|---|
| Zhao et al., 2008 | Saponin HepG2.2.15 cells and Peking ducklings | No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [24] |
| Yao et al., 2020 | Saponin HepG2.2.15 cells and C57BL/6 mice | No investigation on specific step of HBV DNA replication Only HBV genome containing recombinant adenovirus is used in mammalian infection system | [25] |
| Li and Wang, 2006 | Ganoderic acid HepG2.2.15 cells | No investigation on specific step of HBV DNA replication | [26] |
| Zhou et al., 2013 | Astataricusones B and epishionol HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [27] |
| Guo et al., 2013 | Hemslecin A HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [28] |
| Wang et al., 2012a | Caudatin HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [29] |
| Wang et al., 2012b | Glycyrrhizin and glycyrrhetinic acid HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [30] |
| Tseng et al., 2008 | Helioxanthin HepA2 cells | Only cell culture system (in vitro) is used No mammalian infection (in vivo) system is used | [31] |
| Huang et al., 2013b | Asiaticoside HepG2.2.15 cells and ducklings | No mammalian infection (in vivo) system is used | [32] |
| Huang et al., 2013a | Methyl helicterate HepG2.2.15 cells and ducklings | No mammalian infection (in vivo) system is used | [33] |
| Song, 2018; Ye et al., 2009 | Epigallocatechin gallate (EGCG) HepG2.117 cells | Only cell culture system (in vitro) is used No mammalian infection (in vivo) system is used | [36,37] |
| Cao et al., 2013a | Isovitexin HepG2.117 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [38] |
| Huang et al., 2014 | LPRP-Et-97543 HepG2.2.15 cells and Huh7 cells | Only cell culture system (in vitro) is used No mammalian infection (in vivo) system is used | [39] |
| Cao et al., 2015b | Isooriention HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [40] |
| Zembower et al., 1998 | Robustaflavone HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [41] |
| Huang et al., 2000 | Wogonin HBV-producing cell line (MS-G2) | Only cell culture system (in vitro) is used No mammalian infection (in vivo) system is used | [42] |
| Li et al., 2008 | Polyphenolic extract HepG2.2.15 cells and ducklings | No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [44] |
| Han et al., 2008 | Phenolics extract HepG2.2.15 cells and ducks | No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [45] |
| Kim et al., 2007 | Quinic acid derivatives HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [46] |
| Wang et al., 2009 | Chlorogenic acid HepG2.2.15 cells and ducklings | No mammalian infection (in vivo) system is used | [47] |
| Zhou et al., 2007 | Protocatechuic aldehyde HepG2.2.15 cells and ducks | No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [48] |
| Geng et al., 2012 | Mulberrofuran G HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [49] |
| Zhao et al., 2015 | p-Hydroxyacetophenone HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [50] |
| Geng et al., 2018 | Atractylodin HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [51] |
| Chen et al., 2014 | Dehydroandrographolide and andrographolide HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [53] |
| Romero et al., 2005 | Artemisinin HepG2.2.15 cells | Only cell culture system (in vitro) is used No mammalian infection (in vivo) system is used | [54] |
| Geng et al., 2011 | Swerilactones H-K HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [55] |
| Wang et al., 2013 | Xanthrone and secoiridoid lactone HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [56] |
| Zhong et al., 2020 | Herpetolide A HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [58] |
| Xu et al., 2020 | Swertisin HepG2.2.15 cells and HBV transgenic mice | No investigation on specific step of HBV DNA replication HBV producing transgenic mice is used No de novo infection in mice is performed | [59] |
| Li et al., 2012; Li et al., 2013 | Honokiol and (7′R, 8′S, 7′R, 8′S)-erythron-strebluslignanol G HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [61,62] |
| Zhou et al., 2015 | Cycloolivil-4-O-β-D-glucopyranoside HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [63] |
| Tutik et al., 2020 | Extract Hep38.7 Tet cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [65] |
| Cao et al., 2013b; Liu et al., 2020 | Mangiferin HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [66,67] |
| Cao et al., 2013a; Liu et al., 2020 | Dihydroxy-3,5-dimethoxyxanthone and norswertianolin HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [38,66] |
| Cao et al., 2015a; Liu et al., 2020 | Trihydroxy-3-methoxyxanthone HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [66,68] |
| Hu et al., 2013 | β-Thujaplicinol Huh7 cells | Only cell culture system (in vitro) is used No mammalian infection (in vivo) system is used | [70] |
| Mou et al., 2021 | Des(rhamnosyl) verbascoside HepG2.2.15 cells | Only cell culture system (in vitro) is used No mammalian infection (in vivo) system is used | [71] |
| Liang et al., 2019 | Heteropolysaccharide (FP-1) HepG2.2.15 cells | Only cell culture system (in vitro) is used No investigation on specific step of HBV DNA replication No mammalian infection (in vivo) system is used | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadiea, R.Z.; Sultana, S.; Chaki, B.M.; Islam, T.; Dash, S.; Akter, S.; Islam, M.S.; Kazi, T.; Nagata, A.; Spagnuolo, R.; et al. Phytomedicines to Target Hepatitis B Virus DNA Replication: Current Limitations and Future Approaches. Int. J. Mol. Sci. 2022, 23, 1617. https://doi.org/10.3390/ijms23031617
Sadiea RZ, Sultana S, Chaki BM, Islam T, Dash S, Akter S, Islam MS, Kazi T, Nagata A, Spagnuolo R, et al. Phytomedicines to Target Hepatitis B Virus DNA Replication: Current Limitations and Future Approaches. International Journal of Molecular Sciences. 2022; 23(3):1617. https://doi.org/10.3390/ijms23031617
Chicago/Turabian StyleSadiea, Rahila Zannat, Shahnaj Sultana, Bijan Mohon Chaki, Tasnim Islam, Sharmy Dash, Sharmin Akter, Md Sayeedul Islam, Taheruzzaman Kazi, Abir Nagata, Rocco Spagnuolo, and et al. 2022. "Phytomedicines to Target Hepatitis B Virus DNA Replication: Current Limitations and Future Approaches" International Journal of Molecular Sciences 23, no. 3: 1617. https://doi.org/10.3390/ijms23031617