
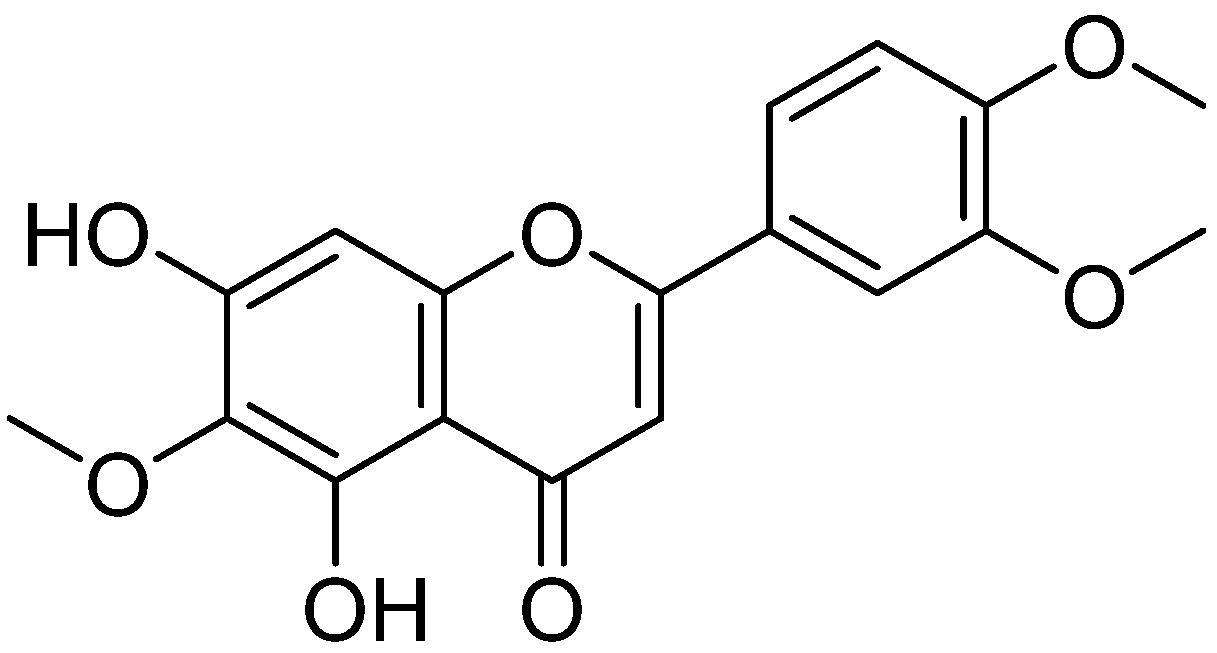
Eupatilin Suppresses OVA-Induced Asthma by Inhibiting NF-κB and MAPK and Activating Nrf2 Signaling Pathways in Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
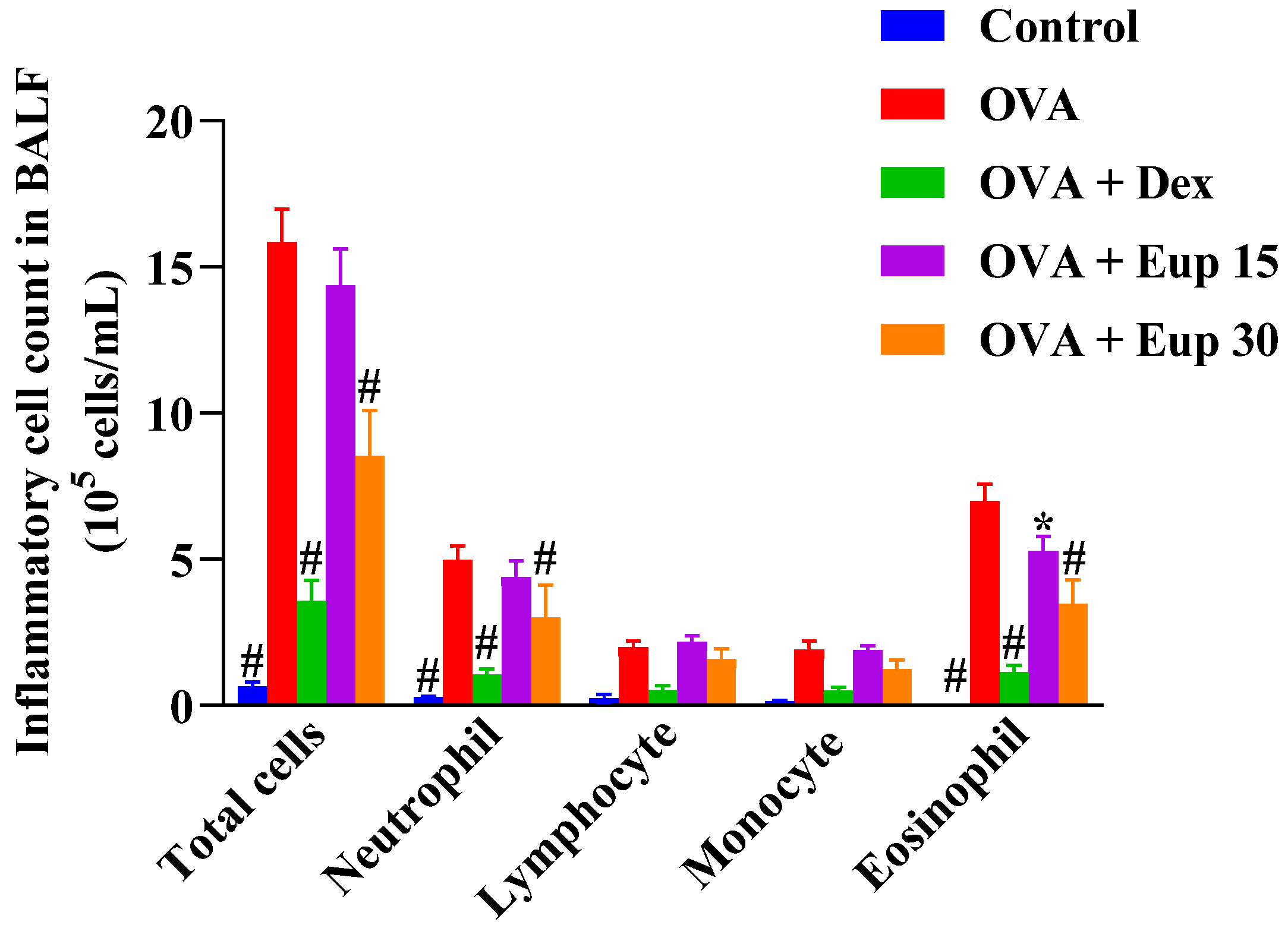
2.1. Effect of Eupatilin on Inflammatory Cells in the Bronchoalveolar Lavage Fluid (BALF)
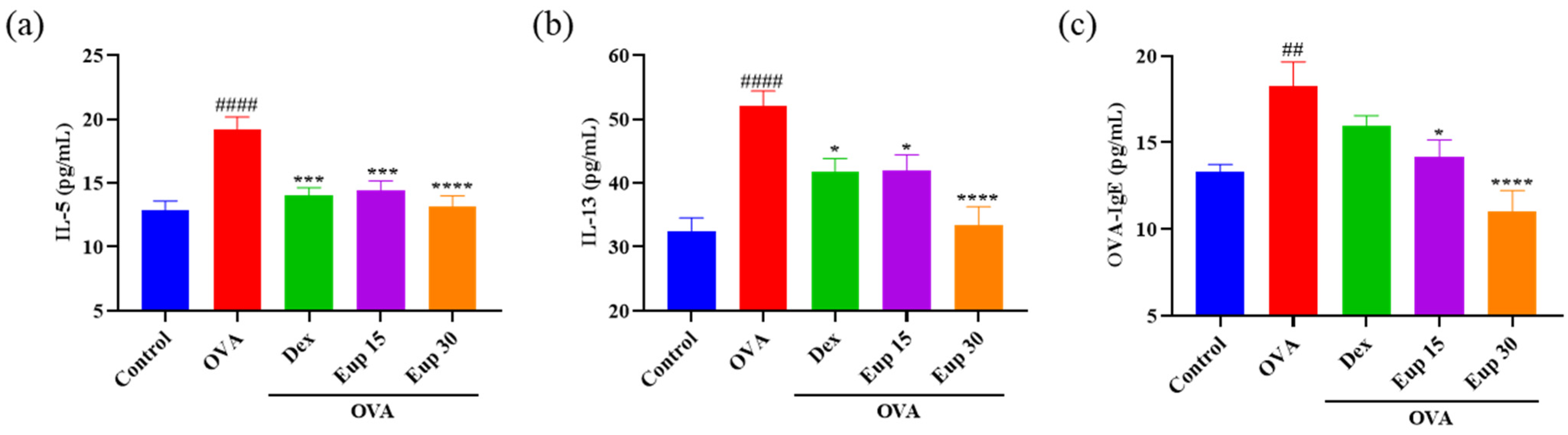
2.2. Eupatilin Reduces OVA-Induced Th2 Cytokine Levels in the BALF and OVA-IgE Levels in the Serum
2.3. Effect of Eupatilin on Lung Histological Changes in Asthmatic Mice
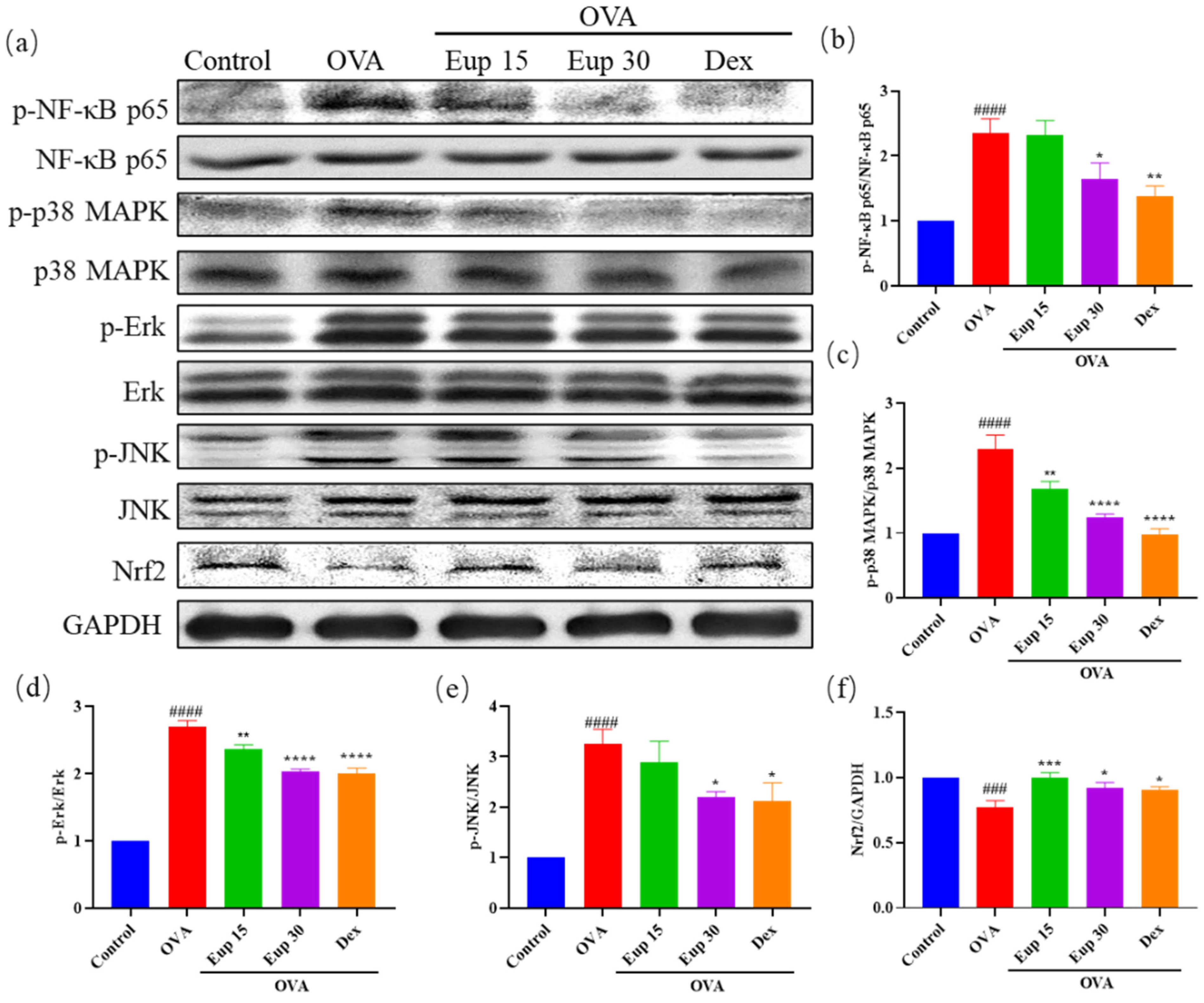
2.4. Effect of Eupatilin on NF-κB, MAPK and Nrf2 Signaling Pathways in Asthmatic Mice
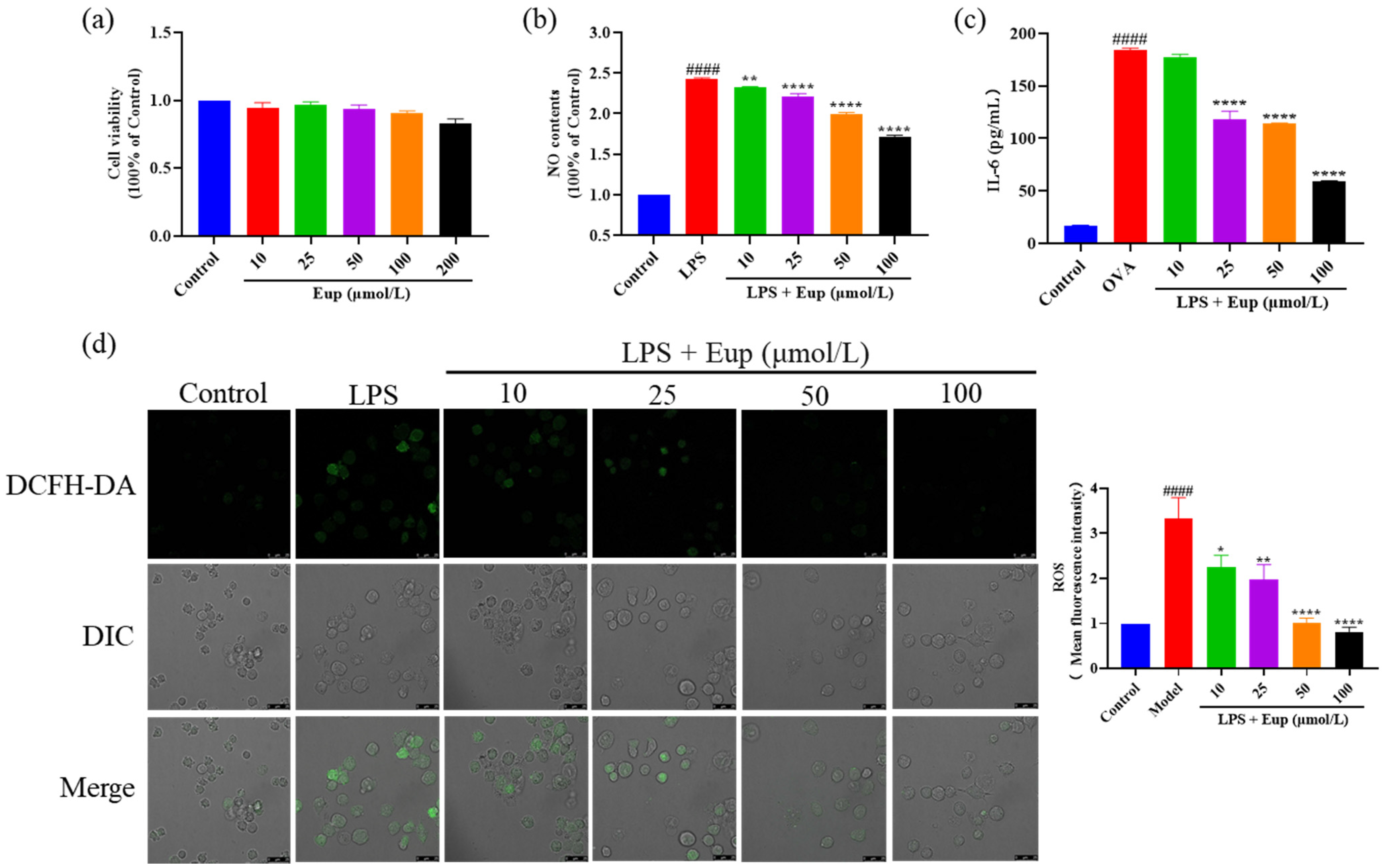
2.5. Effect of Eupatilin on Inflammatory Cytokines in RAW264.7 Cells
2.6. Effect of Eupatilin on NF-κB, MAPK, and Nrf2 Signaling Pathways in RAW264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Drugs and Reagents
4.2. Animals
4.3. Sensitization and Treatment Protocols
4.4. BALF Collection and Leukocyte Counts
4.5. Determination of IL-5, IL-13 and OVA-IgE Levels
4.6. H&E and PAS Staining
4.7. Cell Culture and Treatment
4.8. MTT Assay
4.9. Measurement of NO
4.10. Measurement of ROS
4.11. Measurement of IL-6
4.12. Western Blot Analysis
4.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
Abbreviations
| NF-κB | nuclear factor kappa B |
| MAPKs | mitogen-activated protein kinases |
| JNK | c-Jun N-terminal kinase |
| Nrf2 | nuclear factor-erythroid 2-related factor 2 |
| OVA | ovalbumin |
| Dex | dexamethasone |
| BALF | bronchoalveolar lavage fluid |
| LPS | lipopolysaccharide |
| FBS | fetal bovine serum |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| ROS | reactive oxygen species |
References
- Luo, X.Q.; Zhong, J.W.; Qiu, S.Y.; Zhi, M.; Yang, L.Q.; Zhou, Y.L.; Zhou, F.X.; Yang, P.C.; Liu, D.B.; Mo, L.H. A20-OVA nanoparticles inhibit allergic asthma in a murine model. Inflammation 2020, 43, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Yang, L.; Wang, S.; Zhu, Y.; Shi, W.; Zhang, C.; Jin, H.; Tian, Y.; Xu, H.; Sun, G.; et al. Double negative T cells mediate Lag3-dependent antigen-specific protection in allergic asthma. Nat. Commun. 2019, 10, 4246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, Y.; Gu, H.; Zhang, J.; Tang, H.; Rong, Q.; Gu, L.; Pan, J.; Zhao, D.; Liu, F. LncRNA-AK149641 associated with airway inflammation in an OVA-induced asthma mouse model. J. Bioenerg. Biomembr. 2020, 52, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.W.; Kwon, H.J.; Seo, C.S.; Choi, S.J.; Shin, N.R.; Kim, S.H.; Kim, Y.H.; Kim, J.C.; Kim, M.S.; Shin, I.S. 4-Hydroxycinnamic acid suppresses airway inflammation and mucus hypersecretion in allergic asthma induced by ovalbumin challenge. Phytother. Res. 2020, 34, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tang, J.; Qian, B.; Zeng, Z.; Gao, Y.; Song, J.L. Rubusoside alleviates the ovalbumin-induced mice allergic asthma by modulating the NF-κB activation. J. Food Biochem. 2020, 44, e13187. [Google Scholar] [CrossRef] [PubMed]
- Ji, N.F.; Xie, Y.C.; Zhang, M.S.; Zhao, X.; Cheng, H.; Wang, H.; Yin, K.S.; Huang, M. Ligustrazine corrects Th1/Th2 and Treg/Th17 imbalance in a mouse asthma model. Int. Immunopharmacol. 2014, 21, 76–81. [Google Scholar] [CrossRef]
- Franova, S.; Joskova, M.; Sadlonova, V.; Pavelcikova, D.; Mesarosova, L.; Novakova, E.; Sutovska, M. Experimental model of allergic asthma. Adv. Exp. Med. Biol. 2013, 756, 49–55. [Google Scholar]
- Bosnjak, B.; Stelzmueller, B.; Erb, K.J.; Epstein, M.M. Treatment of allergic asthma: Modulation of Th2 cells and their responses. Respir. Res. 2011, 12, 114. [Google Scholar] [CrossRef] [Green Version]
- Alrumaihi, F.; Almatroudi, A.; Allemailem, K.S.; Rahmani, A.H.; Khan, A.; Khan, M.A. Therapeutic effect of bilsaan, Sambucus nigra stem exudate, on the OVA-induced allergic asthma in mice. Oxid. Med. Cell. Longev. 2020, 2020, 3620192. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The cytokines of asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Walsh, G.M. Anti-IL-4/-13 based therapy in asthma. Expert Opin. Emerg. Drugs 2015, 20, 349–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, X.; Ci, X.; He, J.; Wei, M.; Yang, X.; Cao, Q.; Li, H.; Guan, S.; Deng, Y.; Pang, D.; et al. A novel anti-inflammatory role for ginkgolide B in asthma via inhibition of the ERK/MAPK signaling pathway. Molecules 2011, 16, 7634–7648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, E.; Fu, Y.; Wei, Z.; Yu, Y.; Zhang, X.; Yang, Z. Thymol attenuates allergic airway inflammation in ovalbumin (OVA)-induced mouse asthma. Fitoterapia 2014, 96, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, L.; Chen, B.; Zhuo, Q.; Bao, C.; Lin, L. Propofol inhibits NF-κB activation to ameliorate airway inflammation in ovalbumin (OVA)-induced allergic asthma mice. Int. Immunopharmacol. 2017, 51, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Choi, Y.H.; Xian, Z.; Zheng, M.; Piao, H.; Yan, G. Aloperine suppresses allergic airway inflammation through NF-κB, MAPK, and Nrf2/HO-1 signaling pathways in mice. Int. Immunopharmacol. 2018, 65, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.W.; Chiou, W.F.; Chao, S.H.; Lee, M.H.; Chen, C.C.; Tsai, Y.C. Ligustilide prevents LPS-induced iNOS expression in RAW 264.7 macrophages by preventing ROS production and down-regulating the MAPK, NF-κB and AP-1 signaling pathways. Int. Immunopharmacol. 2011, 11, 1166–1172. [Google Scholar] [CrossRef]
- Gu, X.; Zhang, Q.; Du, Q.; Shen, H.; Zhu, Z. Pinocembrin attenuates allergic airway inflammation via inhibition of NF-κB pathway in mice. Int. Immunopharmacol. 2017, 53, 90–95. [Google Scholar] [CrossRef]
- Santana, F.P.R.; da Silva, R.C.; Ponci, V.; Pinheiro, A.; Olivo, C.R.; Caperuto, L.C.; Arantes-Costa, F.M.; Claudio, S.R.; Ribeiro, D.A.; Tibério, I.; et al. Dehydrodieugenol improved lung inflammation in an asthma model by inhibiting the STAT3/SOCS3 and MAPK pathways. Biochem. Pharmacol. 2020, 180, 114175. [Google Scholar] [CrossRef]
- Liu, L.; Shang, Y.; Li, M.; Han, X.; Wang, J.; Wang, J. Curcumin ameliorates asthmatic airway inflammation by activating nuclear factor-E2-related factor 2/haem oxygenase (HO)-1 signalling pathway. Clin. Exp. Pharmacol. Physiol. 2015, 42, 520–529. [Google Scholar] [CrossRef]
- Liu, L.; Wang, L.P.; He, S.; Ma, Y. Immune homeostasis: Effects of Chinese herbal formulae and herb-derived compounds on allergic asthma in different experimental models. Chin. J. Integr. Med. 2018, 24, 390–398. [Google Scholar] [CrossRef]
- Jeong, J.H.; Moon, S.J.; Jhun, J.Y.; Yang, E.J.; Cho, M.L.; Min, J.K. Eupatilin exerts antinociceptive and chondroprotective properties in a rat model of osteoarthritis by downregulating oxidative damage and catabolic activity in chondrocytes. PLoS ONE 2015, 10, e0130882. [Google Scholar] [CrossRef] [PubMed]
- Nageen, B.; Sarfraz, I.; Rasul, A.; Hussain, G.; Rukhsar, F.; Irshad, S.; Riaz, A.; Selamoglu, Z.; Ali, M. Eupatilin: A natural pharmacologically active flavone compound with its wide range applications. J. Asian Nat. Prod. Res. 2020, 22, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Lee, S.; Chae, J.R.; Lee, H.S.; Jun, C.D.; Kim, S.H. Eupatilin inhibits lipopolysaccharide-induced expression of inflammatory mediators in macrophages. Life Sci. 2011, 88, 1121–1126. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.I.; Ko, S.H.; Kim, Y.J.; Choi, S.M.; Kang, K.K.; Kim, H.; Yoon, H.J.; Kim, J.M. The flavone eupatilin inhibits eotaxin expression in an NF-κB-dependent and STAT6-independent manner. Scand. J. Immunol. 2015, 81, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, E.H.; Chung, K.S.; Kang, Y.M.; Lee, J.H.; Lee, M.; An, H.J. Eupatilin suppresses the allergic inflammatory response in vitro and in vivo. Phytomedicine 2018, 42, 1–8. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kwon, E.Y.; Lee, Y.S.; Kim, W.B.; Ro, J.Y. Eupatilin blocks mediator release via tyrosine kinase inhibition in activated guinea pig lung mast cells. J. Toxicol. Environ. Health Part A 2005, 68, 2063–2080. [Google Scholar]
- Liu, H.; Hao, J.; Wu, C.; Liu, G.; Wang, X.; Yu, J.; Liu, Y.; Zhao, H. Eupatilin alleviates lipopolysaccharide-induced acute lung injury by inhibiting inflammation and oxidative stress. Med. Sci. Monit. 2019, 25, 8289–8296. [Google Scholar] [CrossRef]
- He, J.; Lv, L.; Wang, Z.; Huo, C.; Zheng, Z.; Yin, B.; Jiang, P.; Yang, Y.; Li, J.; Gao, Y.; et al. Pulvis Fellis Suis extract attenuates ovalbumin-induced airway inflammation in murine model of asthma. J. Ethnopharmacol. 2017, 207, 34–41. [Google Scholar] [CrossRef]
- Liu, J.; Xiong, H.; Cheng, Y.; Cui, C.; Zhang, X.; Xu, L.; Zhang, X. Effects of taraxasterol on ovalbumin-induced allergic asthma in mice. J. Ethnopharmacol. 2013, 148, 787–793. [Google Scholar] [CrossRef]
- Lee, I.T.; Yang, C.M. Inflammatory signalings involved in airway and pulmonary diseases. Mediat. Inflamm. 2013, 2013, 791231. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Kumar, R.K.; Hansbro, P.M.; Foster, P.S. Emerging roles of pulmonary macrophages in driving the development of severe asthma. J. Leukoc. Biol. 2012, 91, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Wu, J.; Wang, J.; Guo, Y.; Gao, L.; Liang, H.; Huang, J.; Wang, J. Ma Huang Tang ameliorates bronchial asthma symptoms through the TLR9 pathway. Pharm. Biol. 2018, 56, 580–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.Q.; Wei, H.L.; Zhang, X. TLR2 regulates allergic airway inflammation through NF-κB and MAPK signaling pathways in asthmatic mice. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3138–3146. [Google Scholar] [PubMed]
- Abdelaziz, R.R.; Elmahdy, M.K.; Suddek, G.M. Flavocoxid attenuates airway inflammation in ovalbumin-induced mouse asthma model. Chem. Biol. Interact. 2018, 292, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ge, A.; Zhu, W.; Liu, Y.N.; Ji, N.F.; Zha, W.J.; Zhang, J.X.; Zeng, X.N.; Huang, M. Morin attenuates ovalbumin-induced airway inflammation by modulating oxidative stress-responsive MAPK signaling. Oxid. Med. Cell. Longev. 2016, 2016, 5843672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ci, X.; Chu, X.; Wei, M.; Yang, X.; Cai, Q.; Deng, X. Different effects of farrerol on an OVA-induced allergic asthma and LPS-induced acute lung injury. PLoS ONE 2012, 7, e34634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, C.; Gao, F.; Li, X.; Li, Z. Icariside II attenuates eosinophils-induced airway inflammation and remodeling via inactivation of NF-κB and STAT3 in an asthma mouse model. Exp. Mol. Pathol. 2020, 113, 104373. [Google Scholar] [CrossRef]
- Cho, J.Y. Recent advances in mechanisms and treatments of airway remodeling in asthma: A message from the bench side to the clinic. Korean J. Intern. Med. 2011, 26, 367–383. [Google Scholar] [CrossRef]
- Gauvreau, G.M.; El-Gammal, A.I.; O’Byrne, P.M. Allergen-induced airway responses. Eur. Respir. J. 2015, 46, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Zhi, Y.; Huang, H.; Liang, L. MFG-E8/integrin β3 signaling contributes to airway inflammation response and airway remodeling in an ovalbumin-induced murine model of asthma. J. Cell. Biochem. 2018, 119, 8887–8896. [Google Scholar] [CrossRef]
- Wu, D.; Li, S.; Liu, X.; Xu, J.; Jiang, A.; Zhang, Y.; Liu, Z.; Wang, J.; Zhou, E.; Wei, Z.; et al. Alpinetin prevents inflammatory responses in OVA-induced allergic asthma through modulating PI3K/AKT/NF-κB and HO-1 signaling pathways in mice. Int. Immunopharmacol. 2020, 89, 107073. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, M. Th2 cells as targets for therapeutic intervention in allergic bronchial asthma. Expert Rev. Mol. Diagn. 2009, 9, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Shin, I.S.; Jeon, W.Y.; Shin, H.K.; Lee, M.Y. Effects of montelukast on subepithelial/peribronchial fibrosis in a murine model of ovalbumin induced chronic asthma. Int. Immunopharmacol. 2013, 17, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Umetsu, D.T.; DeKruyff, R.H. TH1 and TH2 CD4+ cells in human allergic diseases. J. Allergy Clin. Immunol. 1997, 100, 1–6. [Google Scholar] [CrossRef]
- Mishra, V.; Banga, J.; Silveyra, P. Oxidative stress and cellular pathways of asthma and inflammation: Therapeutic strategies and pharmacological targets. Pharmacol. Ther. 2018, 181, 169–182. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Shabnam, B.; Girisa, S.; Harsha, C.; Banik, K.; Devi, T.B.; Choudhury, R.; Sahu, H.; Parama, D.; Sailo, B.L.; et al. Inflammation, NF-κB, and chronic diseases: How are they linked? Crit. Rev. Immunol. 2020, 40, 1–39. [Google Scholar] [CrossRef]
- Roy, S.; Manna, K.; Jha, T.; Saha, K.D. Chrysin-loaded PLGA attenuates OVA-induced allergic asthma by modulating TLR/NF-κB/NLRP3 axis. Nanomed. Nanotechnol. Biol. Med. 2020, 30, 102292. [Google Scholar] [CrossRef]
- Dai, R.; Niu, M.; Wang, N.; Wang, Y. Syringin alleviates ovalbumin-induced lung inflammation in BALB/c mice asthma model via NF-κB signaling pathway. Environ. Toxicol. 2021, 36, 433–444. [Google Scholar] [CrossRef]
- Yoon, W.J.; Moon, J.Y.; Song, G.; Lee, Y.K.; Han, M.S.; Lee, J.S.; Ihm, B.S.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Artemisia fukudo essential oil attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 1222–1229. [Google Scholar] [CrossRef]
- Das, J.; Chen, C.H.; Yang, L.; Cohn, L.; Ray, P.; Ray, A. A critical role for NF-kappa B in GATA3 expression and TH2 differentiation in allergic airway inflammation. Nat. Immunol. 2001, 2, 45–50. [Google Scholar] [CrossRef]
- Abohassan, M.; Al Shahrani, M.; Alshahrani, M.Y.; Begum, N.; Radhakrishnan, S.; Rajagopalan, P. FNF-12, a novel benzylidene-chromanone derivative, attenuates inflammatory response in in vitro and in vivo asthma models mediated by M2-related Th2 cytokines via MAPK and NF-kB signaling. Pharmacol. Rep. 2022, 74, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Son, E.S.; Park, J.W.; Kim, S.H.; Park, H.R.; Han, W.; Kwon, O.C.; Nam, J.Y.; Jeong, S.H.; Lee, C.S. Anti-inflammatory activity of 3,5,6,7,3′,4′-hexamethoxyflavone via repression of the NF-κB and MAPK signaling pathways in LPS-stimulated RAW264.7 cells. Mol. Med. Rep. 2020, 22, 1985–1993. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.I.; Hong, S.H.; Ku, J.M.; Kang, S.; Kim, T.Y.; Shin, Y.C.; Ko, S.G. Tonggyu-tang, a traditional Korean medicine, suppresses pro-inflammatory cytokine production through inhibition of MAPK and NF-κB activation in human mast cells and keratinocytes. BMC Complement. Altern. Med. 2017, 17, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryter, S.W.; Choi, A.M. Targeting heme oxygenase-1 and carbon monoxide for therapeutic modulation of inflammation. Transl. Res. J. Lab. Clin. Med. 2016, 167, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, M.C.; McGrath, K.W.; Hawkins, G.A.; Hastie, A.T.; Levy, B.D.; Israel, E.; Phillips, B.R.; Mauger, D.T.; Comhair, S.A.; Erzurum, S.C.; et al. Plasma interleukin-6 concentrations, metabolic dysfunction, and asthma severity: A cross-sectional analysis of two cohorts. Lancet Respir. Med. 2016, 4, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Luu Quoc, Q.; Cao Thi Bich, T.; Kim, S.H.; Park, H.S.; Shin, Y.S. Administration of vitamin E attenuates airway inflammation through restoration of Nrf2 in a mouse model of asthma. J. Cell. Mol. Med. 2021, 25, 6721–6732. [Google Scholar] [CrossRef]
- Fricker, M.; Gibson, P.G. Macrophage dysfunction in the pathogenesis and treatment of asthma. Eur. Respir. J. 2017, 50, 1700196. [Google Scholar] [CrossRef]
- Han, Y.; Yuan, C.; Zhou, X.; Han, Y.; He, Y.; Ouyang, J.; Zhou, W.; Wang, Z.; Wang, H.; Li, G. Anti-inflammatory activity of three triterpene from Hippophae rhamnoides L. in lipopolysaccharide-stimulated RAW264.7 cells. Int. J. Mol. Sci. 2021, 22, 12009. [Google Scholar] [CrossRef]
- Ou, G.; Liu, Q.; Yu, C.; Chen, X.; Zhang, W.; Chen, Y.; Wang, T.; Luo, Y.; Jiang, G.; Zhu, M.; et al. The protective effects of maresin 1 in the OVA-induced asthma mouse model. Mediat. Inflamm. 2021, 2021, 4131420. [Google Scholar] [CrossRef]
- Li, Q.; Bai, D.; Qin, L.; Shao, M.; Liu, X.; Zhang, S.; Yan, C.; Yu, G.; Hao, J. Protective effect of L-hexaguluroic acid hexasodium salt on UVA-induced photo-aging in HaCaT cells. Int. J. Mol. Sci. 2020, 21, 1201. [Google Scholar] [CrossRef] [Green Version]
- Xin, W.; Huang, C.; Zhang, X.; Zhang, G.; Ma, X.; Sun, L.; Wang, C.; Zhang, D.; Zhang, T.; Du, G. Evaluation of the new anti-inflammatory compound ethyl salicylate 2-O-β-d-glucoside and its possible mechanism of action. Int. Immunopharmacol. 2013, 15, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Zhang, Y.; Sun, J.; Wang, X.; Tu, H.; Geng, S.; Liu, R.; Chen, Y.; Bi, Z. Deoxyelephantopin induces reactive oxygen species-mediated apoptosis and autophagy in human osteosarcoma cells. Cell. Physiol. Biochem. 2017, 42, 1812–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, D.; Sun, T.; Lu, F.; Shen, Y.; Zhang, Y.; Zhang, B.; Yu, G.; Li, H.; Hao, J. Eupatilin Suppresses OVA-Induced Asthma by Inhibiting NF-κB and MAPK and Activating Nrf2 Signaling Pathways in Mice. Int. J. Mol. Sci. 2022, 23, 1582. https://doi.org/10.3390/ijms23031582
Bai D, Sun T, Lu F, Shen Y, Zhang Y, Zhang B, Yu G, Li H, Hao J. Eupatilin Suppresses OVA-Induced Asthma by Inhibiting NF-κB and MAPK and Activating Nrf2 Signaling Pathways in Mice. International Journal of Molecular Sciences. 2022; 23(3):1582. https://doi.org/10.3390/ijms23031582
Chicago/Turabian StyleBai, Donghui, Tianxiao Sun, Fang Lu, Yancheng Shen, Yan Zhang, Bo Zhang, Guangli Yu, Haihua Li, and Jiejie Hao. 2022. "Eupatilin Suppresses OVA-Induced Asthma by Inhibiting NF-κB and MAPK and Activating Nrf2 Signaling Pathways in Mice" International Journal of Molecular Sciences 23, no. 3: 1582. https://doi.org/10.3390/ijms23031582