
The HLA-G Immune Checkpoint Plays a Pivotal Role in the Regulation of Immune Response in Autoimmune Diseases

 , , , and
, , , and
Abstract
:1. Introduction
2. Molecular Characterization of the HLA-G Antigen
Regulation of HLA-G Expression

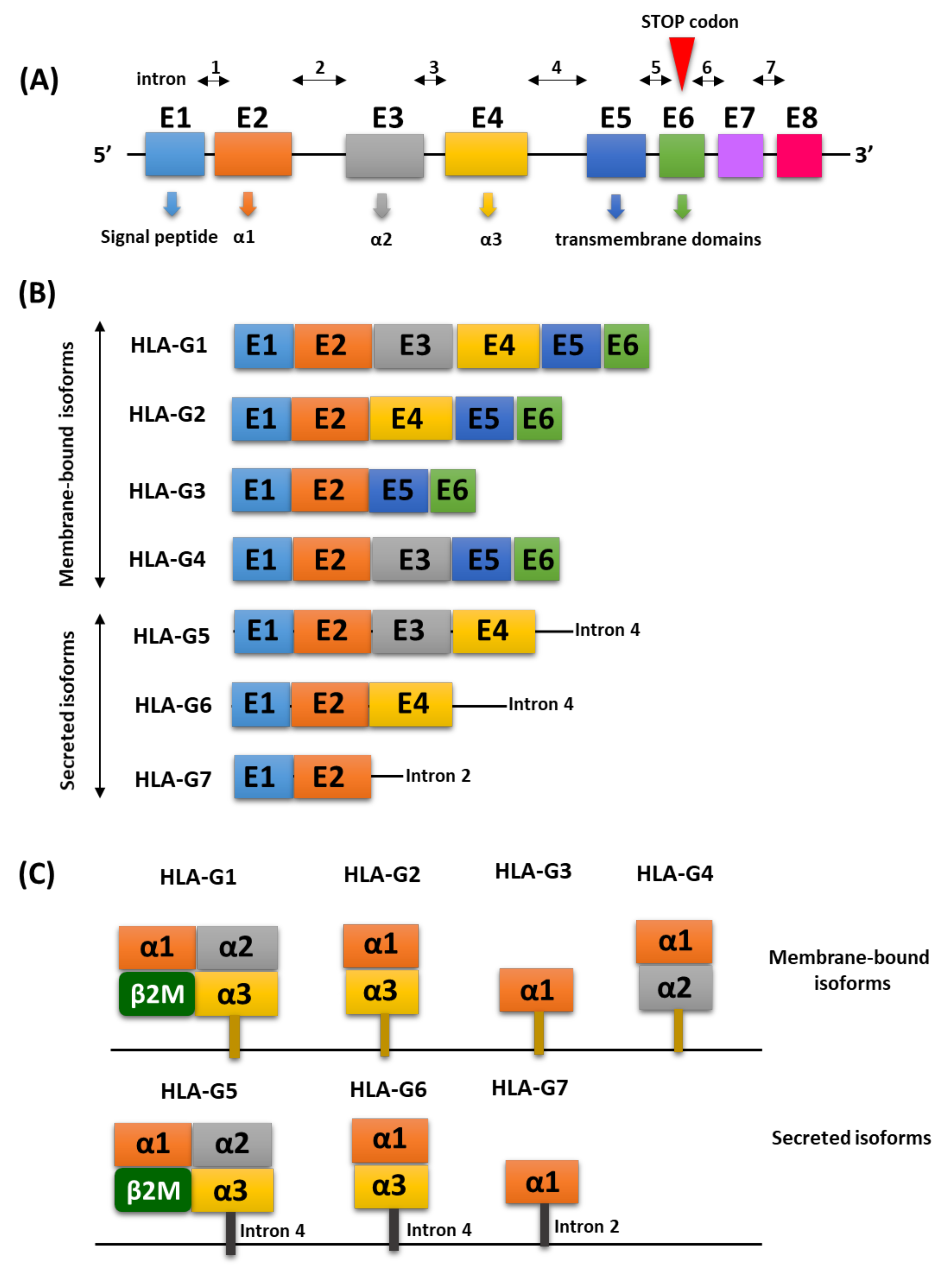{kind=link}
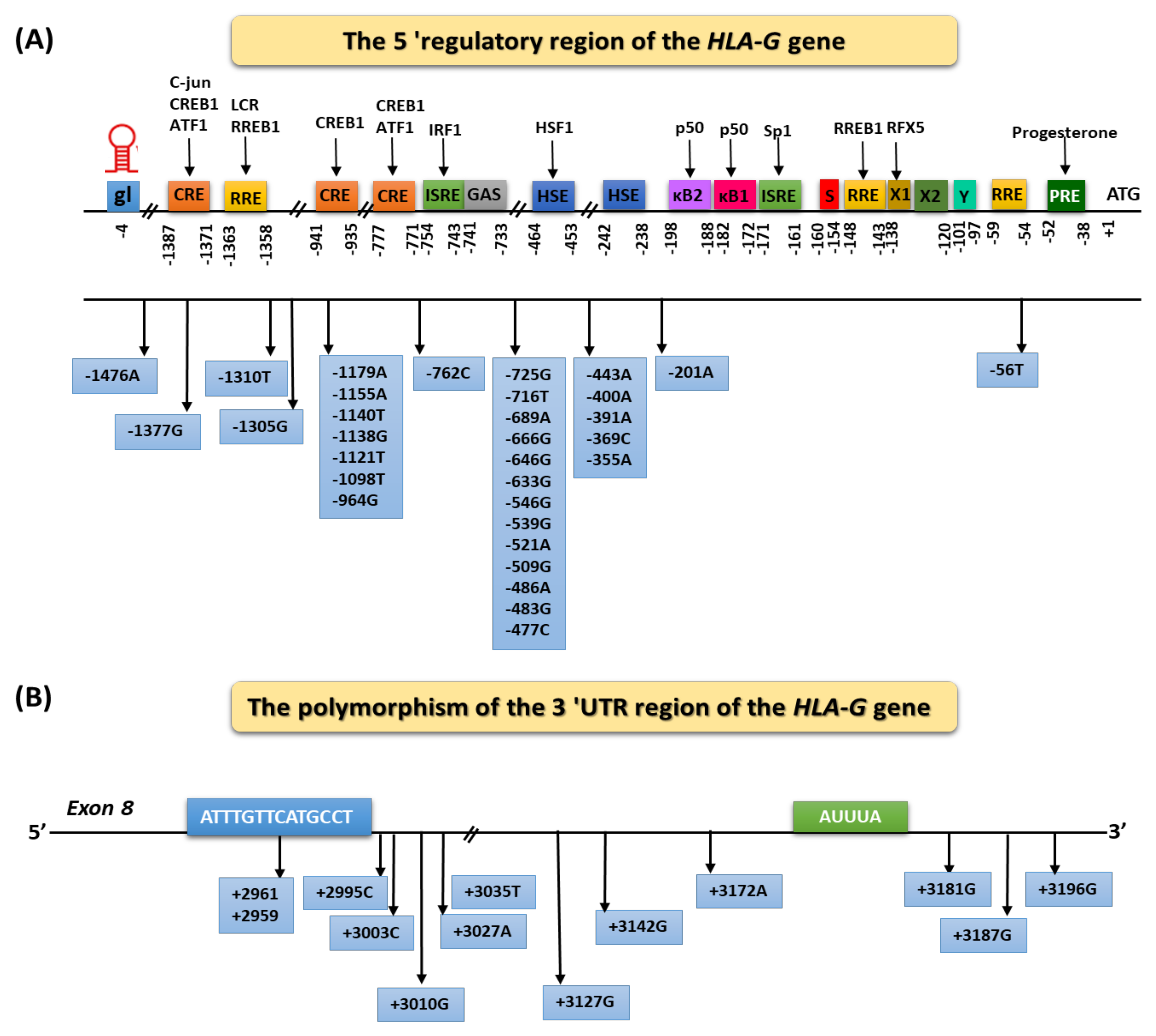{kind=link}
| Haplotype | Type of Mutation | Localization of Haplotype HLA-G in 3′UTR | ||||||
|---|---|---|---|---|---|---|---|---|
| +3003 | +3010 | +3027 | +3035 | +3142 | +3187 | +3196 | ||
| UTR-1 | Deletion | T | G | C | C | C | G | C |
| UTR-2 | Insertion | T | C | C | C | G | A | G |
| UTR-3 | Deletion | T | C | C | C | G | A | C |
| UTR-4 | Deletion | C | G | C | C | C | A | C |
| UTR-5 | Insertion | T | C | C | T | G | A | C |
| UTR-6 | Deletion | T | G | C | C | C | A | C |
| UTR-7 | Insertion | T | C | A | T | G | A | C |
| UTR-8 | Insertion | T | G | C | C | G | A | G |
3. Contribution of the HLA-G Antigens to the Immune Response
3.1. Characterization of Receptors for the HLA-G Molecule
3.2. The Role of HLA-G in Physiological Conditions
| Name of Immune Cell | Tolerogenic Function of HLA-G | Reference |
|---|---|---|
| T cells | Inhibition of proliferation, cytolisys, and chemotaxis, cytotoxicity, and INF-y secretion | [67,68,69] |
| Induction of Treg, cytokine produced by Th2 | [70] | |
| B cells | Inhibition of proliferation, Ig secretion, and chemotaxis | [71,72] |
| Neutrophils | Inhibition of phagocytosis and reactive oxygen species production | [73] |
| Dendritic cells | Inhibition of maturation and NK cell activation | [54,74] |
| Induction of anergic and suppressor T cells, and tolerogenic DC | [54,75,76] | |
| NK cells | Inhibition of chemotaxis, cytotoxicity, and INF-y secretion | [77,78] |
4. Selected Autoimmune Diseases Which Involve the HLA-G Molecule
4.1. The Role of the HLA-G Molecule in the Pathogenesis of Autoimmune Diseases of the Gastrointestinal Tract
4.1.1. Influence of the HLA-G Molecule on the Development of Celiac Disease
4.1.2. The Role of HLA-G in the Development of Crohn’s Disease
4.2. The Role of HLA-G in the Development of Multiple Sclerosis
4.3. The Role of HLA-G in the Development of Rheumatoid Diseases
4.3.1. The Role of HLA-G in the Development of Rheumatoid Arthritis
4.3.2. The Role of HLA-G in the Development of Systemic Scleroderma
4.3.3. The Role of HLA-G in the Development of Systemic Lupus Erythematosus
4.3.4. The Role of HLA-G in the Development of Behçet’s Disease
4.3.5. The Role of HLA-G in the Development of Kawasaki Disease
4.4. The Role of HLA-G in the Development of Selected Respiratory Diseases
4.4.1. The Role of HLA-G in the Development of Asthma
4.4.2. The Role of HLA-G in the Development of Allergic Rhinitis
4.5. The Role of HLA-G in the Development of Diabetes
4.6. The Role of HLA-G in the Development of Psoriasis
4.7. Role of HLA-G in Uveitis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carosella, E.D.; Moreau, P.; Lemaoult, J.; Rouas-Freiss, N. HLA-G: From Biology to Clinical Benefits. Trends Immunol. 2008, 29, 125–132. [Google Scholar] [CrossRef]
- Carosella, E.D.; Favier, B.; Rouas-Freiss, N.; Moreau, P.; LeMaoult, J. Beyond the Increasing Complexity of the Immunomodulatory HLA-G Molecule. Blood 2008, 111, 4862–4870. [Google Scholar] [CrossRef] [PubMed]
- LeMaoult, J.; Yan, W.-H. Editorial: The Biological and Clinical Aspects of HLA-G. Front. Immunol. 2021, 12, 441. [Google Scholar] [CrossRef]
- Xu, H.-H.; Lin, A.; Yan, W.-H. Chapter 14—HLA-G-Mediated Immunological Tolerance and Autoimmunity. In Translational Autoimmunity; Rezaei, N., Ed.; Translational Immunology; Academic Press: Cambridge, MA, USA, 2022; Volume 1, pp. 265–295. ISBN 978-0-12-822564-6. [Google Scholar]
- Ishitani, A.; Geraghty, D.E. Alternative Splicing of HLA-G Transcripts Yields Proteins with Primary Structures Resembling Both Class I and Class II Antigens. Proc. Natl. Acad. Sci. USA 1992, 89, 3947–3951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, P.; Cabestre, F.A.; Ibrahim, E.C.; Lefebvre, S.; Khalil-Daher, I.; Vazeux, G.; Quiles, R.M.; Bermond, F.; Dausset, J.; Carosella, E.D. Identification of HLA-G7 as a New Splice Variant of the HLA-G MRNA and Expression of Soluble HLA-G5, -G6, and -G7 Transcripts in Human Transfected Cells. Hum. Immunol. 2000, 61, 1138–1149. [Google Scholar] [CrossRef]
- Lila, N.; Rouas-Freiss, N.; Dausset, J.; Carpentier, A.; Carosella, E.D. Soluble HLA-G Protein Secreted by Allo-Specific CD4+ T Cells Suppresses the Allo-Proliferative Response: A CD4+ T Cell Regulatory Mechanism. Proc. Natl. Acad. Sci. USA 2001, 98, 12150–12155. [Google Scholar] [CrossRef] [Green Version]
- Le Friec, G.; Laupèze, B.; Fardel, O.; Sebti, Y.; Pangault, C.; Guilloux, V.; Beauplet, A.; Fauchet, R.; Amiot, L. Soluble HLA-G Inhibits Human Dendritic Cell-Triggered Allogeneic T-Cell Proliferation without Altering Dendritic Differentiation and Maturation Processes. Hum. Immunol. 2003, 64, 752–761. [Google Scholar] [CrossRef]
- Naji, A.; Menier, C.; Morandi, F.; Agaugué, S.; Maki, G.; Ferretti, E.; Bruel, S.; Pistoia, V.; Carosella, E.D.; Rouas-Freiss, N. Binding of HLA-G to ITIM-Bearing Ig-like Transcript 2 Receptor Suppresses B Cell Responses. J. Immunol. 2014, 192, 1536–1546. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, R.; Bortolotti, D.; Bolzani, S.; Fainardi, E. HLA-G Molecules in Autoimmune Diseases and Infections. Front. Immunol. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contini, P.; Murdaca, G.; Puppo, F.; Negrini, S. HLA-G Expressing Immune Cells in Immune Mediated Diseases. Front. Immunol. 2020, 11, 1613. [Google Scholar] [CrossRef]
- Morandi, F.; Rizzo, R.; Fainardi, E.; Rouas-Freiss, N.; Pistoia, V. Recent Advances in Our Understanding of HLA-G Biology: Lessons from a Wide Spectrum of Human Diseases. J. Immunol. Res. 2016, 2016, e4326495. [Google Scholar] [CrossRef] [Green Version]
- HLA-G-HLA-G Protein-Homo Sapiens (Human)-HLA-G Gene & Protein. Available online: https://www.uniprot.org/uniprot/O02924 (accessed on 24 October 2021).
- HLA-G Major Histocompatibility Complex, Class I, G [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3135 (accessed on 24 October 2021).
- Castelli, E.C.; Ramalho, J.; Porto, I.O.P.; Lima, T.H.A.; Felício, L.P.; Sabbagh, A.; Donadi, E.A.; Mendes-Junior, C.T. Insights into HLA-G Genetics Provided by Worldwide Haplotype Diversity. Front. Immunol. 2014, 5, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tronik-Le Roux, D.; Renard, J.; Vérine, J.; Renault, V.; Tubacher, E.; LeMaoult, J.; Rouas-Freiss, N.; Deleuze, J.-F.; Desgrandschamps, F.; Carosella, E.D. Novel Landscape of HLA-G Isoforms Expressed in Clear Cell Renal Cell Carcinoma Patients. Mol. Oncol. 2017, 11, 1561–1578. [Google Scholar] [CrossRef] [PubMed]
- Donadi, E.A.; Castelli, E.C.; Arnaiz-Villena, A.; Roger, M.; Rey, D.; Moreau, P. Implications of the Polymorphism of HLA-G on Its Function, Regulation, Evolution and Disease Association. Cell. Mol. Life Sci. 2011, 68, 369–395. [Google Scholar] [CrossRef] [Green Version]
- Arnaiz-Villena, A.; Juarez, I.; Suarez-Trujillo, F.; López-Nares, A.; Vaquero, C.; Palacio-Gruber, J.; Martin-Villa, J.M. HLA-G: Function, Polymorphisms and Pathology. Int. J. Immunogenet. 2021, 48, 172–192. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Nardi, F.; König, L.; Wagner, B.; Giebel, B.; Santos Manvailer, L.F.; Rebmann, V. Soluble Monomers, Dimers and HLA-G-Expressing Extracellular Vesicles: The Three Dimensions of Structural Complexity to Use HLA-G as a Clinical Biomarker. HLA 2016, 88, 77–86. [Google Scholar] [CrossRef] [PubMed]
- HoWangYin, K.-Y.; Loustau, M.; Wu, J.; Alegre, E.; Daouya, M.; Caumartin, J.; Sousa, S.; Horuzsko, A.; Carosella, E.D.; LeMaoult, J. Multimeric Structures of HLA-G Isoforms Function through Differential Binding to LILRB Receptors. Cell. Mol. Life Sci. 2012, 69, 4041–4049. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Yan, W.-H. Heterogeneity of HLA-G Expression in Cancers: Facing the Challenges. Front. Immunol. 2018, 9, 2164. [Google Scholar] [CrossRef] [PubMed]
- Moreau, P.; Flajollet, S.; Carosella, E.D. Non-Classical Transcriptional Regulation of HLA-G: An Update. J. Cell. Mol. Med. 2009, 13, 2973–2989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, E.C.; Veiga-Castelli, L.C.; Yaghi, L.; Moreau, P.; Donadi, E.A. Transcriptional and Posttranscriptional Regulations of the HLA-G Gene. J. Immunol. Res. 2014, 2014, 734068. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.H.; Hviid, T.V.F. Human Leukocyte Antigen-G Polymorphism in Relation to Expression, Function, and Disease. Hum. Immunol. 2009, 70, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.M.; Orr, H.T. HLA-G Transgenic Mice: A Model for Studying Expression and Function at the Maternal/Fetal Interface. Immunol. Rev. 1995, 147, 53–65. [Google Scholar] [CrossRef]
- Moreau, P.; Paul, P.; Gourand, L.; Prost, S.; Dausset, J.; Carosella, E.; Kirszenbaum, M. HLA-G Gene Transcriptional Regulation in Trophoblasts and Blood Cells: Differential Binding of Nuclear Factors to a Regulatory Element Located 1.1 Kb from Exon 1. Hum. Immunol. 1997, 52, 41–46. [Google Scholar] [CrossRef]
- Ober, C.; Aldrich, C.L.; Chervoneva, I.; Billstrand, C.; Rahimov, F.; Gray, H.L.; Hyslop, T. Variation in the HLA-G Promoter Region Influences Miscarriage Rates. Am. J. Hum. Genet. 2003, 72, 1425–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amodio, G.; Gregori, S. HLA-G Genotype/Expression/Disease Association Studies: Success, Hurdles, and Perspectives. Front. Immunol. 2020, 11, 1178. [Google Scholar] [CrossRef] [PubMed]
- Durmanova, V.; Bandzuchova, H.; Zilinska, Z.; Tirpakova, J.; Kuba, D.; Buc, M.; Polakova, K. Association of HLA-G Polymorphisms in the 3‘UTR Region and Soluble HLA-G with Kidney Graft Outcome. Immunol. Investig. 2019, 48, 644–658. [Google Scholar] [CrossRef]
- Garziera, M.; Bidoli, E.; Cecchin, E.; Mini, E.; Nobili, S.; Lonardi, S.; Buonadonna, A.; Errante, D.; Pella, N.; D’Andrea, M.; et al. HLA-G 3’UTR Polymorphisms Impact the Prognosis of Stage II-III CRC Patients in Fluoropyrimidine-Based Treatment. PLoS ONE 2015, 10, e0144000. [Google Scholar] [CrossRef]
- Hviid, T.V.F.; Hylenius, S.; Rørbye, C.; Nielsen, L.G. HLA-G Allelic Variants Are Associated with Differences in the HLA-G MRNA Isoform Profile and HLA-G MRNA Levels. Immunogenetics 2003, 55, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Hiby, S.E.; King, A.; Sharkey, A.; Loke, Y.W. Molecular Studies of Trophoblast HLA-G: Polymorphism, Isoforms, Imprinting and Expression in Preimplantation Embryo. Tissue Antigens 1999, 53, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Castelli, E.C.; de Almeida, B.S.; Muniz, Y.C.N.; Silva, N.S.B.; Passos, M.R.S.; Souza, A.S.; Page, A.E.; Dyble, M.; Smith, D.; Aguileta, G.; et al. HLA-G Genetic Diversity and Evolutive Aspects in Worldwide Populations. Sci. Rep. 2021, 11, 23070. [Google Scholar] [CrossRef] [PubMed]
- Harrison, G.A.; Humphrey, K.E.; Jakobsen, I.B.; Cooper, D.W. A 14 Bp Deletion Polymorphism in the HLA-G Gene. Hum. Mol. Genet. 1993, 2, 2200. [Google Scholar] [CrossRef]
- Castelli, E.C.; Mendes-Junior, C.T.; Deghaide, N.H.S.; de Albuquerque, R.S.; Muniz, Y.C.N.; Simões, R.T.; Carosella, E.D.; Moreau, P.; Donadi, E.A. The Genetic Structure of 3′untranslated Region of the HLA-G Gene: Polymorphisms and Haplotypes. Genes Immun. 2010, 11, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.H.; Lin, A.; Chen, X.J.; Dai, M.Z.; Gan, L.H.; Zhou, M.Y.; Zhu, M.; Shi, W.W.; Liu, J.M. Association of the Maternal 14-Bp Insertion Polymorphism in the HLA-G Gene in Women with Recurrent Spontaneous Abortions. Tissue Antigens 2006, 68, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, R.; Hviid, T.V.F.; Govoni, M.; Padovan, M.; Rubini, M.; Melchiorri, L.; Stignani, M.; Carturan, S.; Grappa, M.T.; Fotinidi, M.; et al. HLA-G Genotype and HLA-G Expression in Systemic Lupus Erythematosus: HLA-G as a Putative Susceptibility Gene in Systemic Lupus Erythematosus. Tissue Antigens 2008, 71, 520–529. [Google Scholar] [CrossRef]
- Poras, I.; Yaghi, L.; Martelli-Palomino, G.; Mendes-Junior, C.T.; Muniz, Y.C.N.; Cagnin, N.F.; de Almeida, B.S.; Castelli, E.C.; Carosella, E.D.; Donadi, E.A.; et al. Haplotypes of the HLA-G 3’ Untranslated Region Respond to Endogenous Factors of HLA-G+ and HLA-G- Cell Lines Differentially. PLoS ONE 2017, 12, e0169032. [Google Scholar] [CrossRef] [Green Version]
- Attia, J.V.D.; Dessens, C.E.; van de Water, R.; Houvast, R.D.; Kuppen, P.J.K.; Krijgsman, D. The Molecular and Functional Characteristics of HLA-G and the Interaction with Its Receptors: Where to Intervene for Cancer Immunotherapy? Int. J. Mol. Sci. 2020, 21, 8678. [Google Scholar] [CrossRef]
- Christmas, S.E. Expression of HLA-G and Its Receptors in Relation to Transplant Acceptance. Integr. Clin. Med. 2020, 4, 1–2. [Google Scholar] [CrossRef]
- Billadeau, D.D.; Leibson, P.J. ITAMs versus ITIMs: Striking a Balance during Cell Regulation. J. Clin. Investig. 2002, 109, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Yan, W.-H. The Emerging Roles of Human Leukocyte Antigen-F in Immune Modulation and Viral Infection. Front. Immunol. 2019, 10, 964. [Google Scholar] [CrossRef] [PubMed]
- Yusa, S.; Catina, T.L.; Campbell, K.S. SHP-1- and Phosphotyrosine-Independent Inhibitory Signaling by a Killer Cell Ig-Like Receptor Cytoplasmic Domain in Human NK Cells. J. Immunol. 2002, 168, 5047–5057. [Google Scholar] [CrossRef] [Green Version]
- Selvakumar, A.; Steffens, U.; Dupont, B. NK Cell Receptor Gene of the KIR Family with Two IG Domains but Highest Homology to KIR Receptors with Three IG Domains. Tissue Antigens 1996, 48, 285–295. [Google Scholar] [CrossRef]
- Loustau, M.; Anna, F.; Dréan, R.; Lecomte, M.; Langlade-Demoyen, P.; Caumartin, J. HLA-G Neo-Expression on Tumors. Front. Immunol. 2020, 11, 1685. [Google Scholar] [CrossRef] [PubMed]
- Shiroishi, M.; Kuroki, K.; Ose, T.; Rasubala, L.; Shiratori, I.; Arase, H.; Tsumoto, K.; Kumagai, I.; Kohda, D.; Maenaka, K. Efficient Leukocyte Ig-like Receptor Signaling and Crystal Structure of Disulfide-Linked HLA-G Dimer. J. Biol. Chem. 2006, 281, 10439–10447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonen-Gross, T.; Achdout, H.; Arnon, T.I.; Gazit, R.; Stern, N.; Hořejší, V.; Goldman-Wohl, D.; Yagel, S.; Mandelboim, O. The CD85J/Leukocyte Inhibitory Receptor-1 Distinguishes between Conformed and Β2-Microglobulin-Free HLA-G Molecules. J. Immunol. 2005, 175, 4866–4874. [Google Scholar] [CrossRef] [Green Version]
- Gros, F.; Cabillic, F.; Toutirais, O.; Maux, A.L.; Sebti, Y.; Amiot, L. Soluble HLA-G Molecules Impair Natural Killer/Dendritic Cell Crosstalk via Inhibition of Dendritic Cells. Eur. J. Immunol. 2008, 38, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Riteau, B.; Menier, C.; Khalil-Daher, I.; Martinozzi, S.; Pla, M.; Dausset, J.; Carosella, E.D.; Rouas-Freiss, N. HLA-G1 Co-Expression Boosts the HLA Class I-Mediated NK Lysis Inhibition. Int. Immunol. 2001, 13, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.-G.; Xu, D.-P.; Lin, A.; Yan, W.-H. NK Cytolysis Is Dependent on the Proportion of HLA-G Expression. Hum. Immunol. 2013, 74, 286–289. [Google Scholar] [CrossRef]
- Manaster, I.; Mandelboim, O. REVIEW ARTICLE: The Unique Properties of Uterine NK Cells. Am. J. Reprod. Immunol. 2010, 63, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, S.; Fu, J.; Long, E.O. Cutting Edge: Induction of IFN-γ Production but Not Cytotoxicity by the Killer Cell Ig-Like Receptor KIR2DL4 (CD158d) in Resting NK Cells. J. Immunol. 2001, 167, 1877–1881. [Google Scholar] [CrossRef] [Green Version]
- Rajagopalan, S.; Long, E.O. A Human Histocompatibility Leukocyte Antigen (HLA)-G–Specific Receptor Expressed on All Natural Killer Cells. J. Exp. Med. 1999, 189, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Page, M.E.L.L.; Goodridge, J.P.; John, E.; Christiansen, F.T.; Witt, C.S. Killer Ig-like Receptor 2DL4 Does Not Mediate NK Cell IFN-γ Responses to Soluble HLA-G Preparations. J. Immunol. 2014, 192, 732–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, I.L.; Montero-Montero, L.; Ferreira, E.; Quintanilla, M. New Insights Into the Role of Qa-2 and HLA-G Non-Classical MHC-I Complexes in Malignancy. Front. Immunol. 2018, 9, 2894. [Google Scholar] [CrossRef] [PubMed]
- Morandi, F.; Ferretti, E.; Bocca, P.; Prigione, I.; Raffaghello, L.; Pistoia, V. A Novel Mechanism of Soluble HLA-G Mediated Immune Modulation: Downregulation of T Cell Chemokine Receptor Expression and Impairment of Chemotaxis. PLoS ONE 2010, 5, e11763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fons, P.; Chabot, S.; Cartwright, J.E.; Lenfant, F.; L’Faqihi, F.; Giustiniani, J.; Herault, J.-P.; Gueguen, G.; Bono, F.; Savi, P.; et al. Soluble HLA-G1 Inhibits Angiogenesis through an Apoptotic Pathway and by Direct Binding to CD160 Receptor Expressed by Endothelial Cells. Blood 2006, 108, 2608–2615. [Google Scholar] [CrossRef] [Green Version]
- Amiot, L.; Vu, N.; Samson, M. Immunomodulatory Properties of HLA-G in Infectious Diseases. J. Immunol. Res. 2014, 2014, e298569. [Google Scholar] [CrossRef] [PubMed]
- Carosella, E.D.; Moreau, P.; Le Maoult, J.; Le Discorde, M.; Dausset, J.; Rouas-freiss, N. HLA-G Molecules: From Maternal–Fetal Tolerance to Tissue Acceptance. In Advances in Immunology; Academic Press: Cambridge, MA, USA, 2003; Volume 81, pp. 199–252. [Google Scholar]
- Feger, U.; Tolosa, E.; Huang, Y.-H.; Waschbisch, A.; Biedermann, T.; Melms, A.; Wiendl, H. HLA-G Expression Defines a Novel Regulatory T-Cell Subset Present in Human Peripheral Blood and Sites of Inflammation. Blood 2007, 110, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Pankratz, S.; Ruck, T.; Meuth, S.G.; Wiendl, H. CD4(+)HLA-G(+) Regulatory T Cells: Molecular Signature and Pathophysiological Relevance. Hum. Immunol. 2016, 77, 727–733. [Google Scholar] [CrossRef]
- LeMaoult, J.; Caumartin, J.; Daouya, M.; Favier, B.; Rond, S.L.; Gonzalez, A.; Carosella, E.D. Immune Regulation by Pretenders: Cell-to-Cell Transfers of HLA-G Make Effector T Cells Act as Regulatory Cells. Blood 2006, 109, 2040–2048. [Google Scholar] [CrossRef]
- Caumartin, J.; Favier, B.; Daouya, M.; Guillard, C.; Moreau, P.; Carosella, E.D.; LeMaoult, J. Trogocytosis-based generation of suppressive NK cells. EMBO J. 2007, 26, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Mitsdoerffer, M.; Schreiner, B.; Kieseier, B.C.; Neuhaus, O.; Dichgans, J.; Hartung, H.-P.; Weller, M.; Wiendl, H. Monocyte-Derived HLA-G Acts as a Strong Inhibitor of Autologous CD4 T Cell Activation and Is Upregulated by Interferon-β in Vitro and in Vivo: Rationale for the Therapy of Multiple Sclerosis. J. Neuroimmunol. 2005, 159, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Bahri, R.; Hirsch, F.; Josse, A.; Rouas-Freiss, N.; Bidere, N.; Vasquez, A.; Carosella, E.D.; Charpentier, B.; Durrbach, A. Soluble HLA-G Inhibits Cell Cycle Progression in Human Alloreactive T Lymphocytes. J. Immunol. 2006, 176, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, F.-S.; Gershwin, M.E. Human Autoimmune Diseases: A Comprehensive Update. J. Intern. Med. 2015, 278, 369–395. [Google Scholar] [CrossRef] [PubMed]
- Sallese, M.; Lopetuso, L.R.; Efthymakis, K.; Neri, M. Beyond the HLA Genes in Gluten-Related Disorders. Front. Nutr. 2020, 7, 239. [Google Scholar] [CrossRef] [PubMed]
- Catamo, E.; Zupin, L.; Segat, L.; Celsi, F.; Crovella, S. HLA-G and Susceptibility to Develop Celiac Disease. Hum. Immunol. 2015, 76, 36–41. [Google Scholar] [CrossRef]
- Loftus, E.V., Jr.; Schoenfeld, P.; Sandborn, W.J. The Epidemiology and Natural History of Crohn’s Disease in Population-Based Patient Cohorts from North America: A Systematic Review. Aliment. Pharmacol. Ther. 2002, 16, 51–60. [Google Scholar] [CrossRef]
- Sood, A.; Midha, V.; Sood, N.; Bhatia, A.S.; Avasthi, G. Incidence and Prevalence of Ulcerative Colitis in Punjab, North India. Gut 2003, 52, 1587–1590. [Google Scholar] [CrossRef] [PubMed]
- Möller, E. Mechanisms for Induction of Autoimmunity in Humans. Acta Paediatr. 1998, 87, 16–20. [Google Scholar] [CrossRef]
- Suzuki, K.; Mori, A.; Ishii, K.J.; Saito, J.; Singer, D.S.; Klinman, D.M.; Krause, P.R.; Kohn, L.D. Activation of Target-Tissue Immune-Recognition Molecules by Double-Stranded Polynucleotides. Proc. Natl. Acad. Sci. USA 1999, 96, 2285–2290. [Google Scholar] [CrossRef] [Green Version]
- Radaev, S.; Sun, P.D. Structure and Function of Natural Killer Cell Surface Receptors. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 93–114. [Google Scholar] [CrossRef]
- Torres, M.I.; Le Discorde, M.; Lorite, P.; Ríos, A.; Gassull, M.A.; Gil, A.; Maldonado, J.; Dausset, J.; Carosella, E.D. Expression of HLA-G in Inflammatory Bowel Disease Provides a Potential Way to Distinguish between Ulcerative Colitis and Crohn’s Disease. Int. Immunol. 2004, 16, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, R.; Melchiorri, L.; Simone, L.; Stignani, M.; Marzola, A.; Gullini, S.; Baricordi, O.R. Different Production of Soluble HLA-G Antigens by Peripheral Blood Mononuclear Cells in Ulcerative Colitis and Crohn’s Disease: A Noninvasive Diagnostic Tool? Inflamm. Bowel Dis. 2008, 14, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Canto, E.; Oksenberg, J.R. Multiple Sclerosis Genetics. Mult. Scler. Houndmills Basingstoke Engl. 2018, 24, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Hollenbach, J.A.; Oksenberg, J.R. The Immunogenetics of Multiple Sclerosis: A Comprehensive Review. J. Autoimmun. 2015, 64, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramagopalan, S.V.; Ebers, G.C. Multiple Sclerosis: Major Histocompatibility Complexity and Antigen Presentation. Genome Med. 2009, 1, 105. [Google Scholar] [CrossRef] [PubMed]
- Europe Autoimmune Disease Diagnostics Market |2021–26| Industry Share, Size, Growth-Mordor Intelligence. Available online: https://www.mordorintelligence.com/industry-reports/european-autoimmune-disease-diagnostics-market-industry (accessed on 24 October 2021).
- Moustakas, A.K.; van de Wal, Y.; Routsias, J.; Kooy, Y.M.C.; van Veelen, P.; Drijfhout, J.W.; Koning, F.; Papadopoulos, G.K. Structure of Celiac Disease-Associated HLA-DQ8 and Non-Associated HLA-DQ9 Alleles in Complex with Two Disease-Specific Epitopes. Int. Immunol. 2000, 12, 1157–1166. [Google Scholar] [CrossRef] [Green Version]
- De Lourdes Moreno, M.; Cebolla, Á.; Muñoz-Suano, A.; Carrillo-Carrion, C.; Comino, I.; Pizarro, Á.; León, F.; Rodríguez-Herrera, A.; Sousa, C. Detection of Gluten Immunogenic Peptides in the Urine of Patients with Coeliac Disease Reveals Transgressions in the Gluten-Free Diet and Incomplete Mucosal Healing. Gut 2017, 66, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Comino, I.; Real, A.; Vivas, S.; Síglez, M.Á.; Caminero, A.; Nistal, E.; Casqueiro, J.; Rodríguez-Herrera, A.; Cebolla, Á.; Sousa, C. Monitoring of Gluten-Free Diet Compliance in Celiac Patients by Assessment of Gliadin 33-Mer Equivalent Epitopes in Feces. Am. J. Clin. Nutr. 2012, 95, 670–677. [Google Scholar] [CrossRef] [Green Version]
- Sciurti, M.; Fornaroli, F.; Gaiani, F.; Bonaguri, C.; Leandro, G.; Mario, F.D.; de’Angelis, G.L. Genetic Susceptibilty and Celiac Disease: What Role Do HLA Haplotypes Play? Acta Biomed. Atenei Parm. 2018, 89, 17–21. [Google Scholar] [CrossRef]
- Bergamaschi, L.; Ban, M.; Barizzone, N.; Leone, M.; Ferrante, D.; Fasano, M.E.; Guerini, F.R.; Corrado, L.; Naldi, P.; Dametto, E.; et al. Association of HLA Class I Markers with Multiple Sclerosis in the Italian and UK Population: Evidence of Two Independent Protective Effects. J. Med. Genet. 2011, 48, 485–492. [Google Scholar] [CrossRef]
- Link, J.; Kockum, I.; Lorentzen, Å.R.; Lie, B.A.; Celius, E.G.; Westerlind, H.; Schaffer, M.; Alfredsson, L.; Olsson, T.; Brynedal, B.; et al. Importance of Human Leukocyte Antigen (HLA) Class I and II Alleles on the Risk of Multiple Sclerosis. PLoS ONE 2012, 7, e36779. [Google Scholar] [CrossRef] [Green Version]
- Maghbooli, Z.; Sahraian, M.A.; Naser Moghadasi, A. Multiple Sclerosis and Human Leukocyte Antigen Genotypes: Focus on the Middle East and North Africa Region. Mult. Scler. J.-Exp. Transl. Clin. 2020, 6, 2055217319881775. [Google Scholar] [CrossRef] [Green Version]
- Fainardi, E.; Rizzo, R.; Melchiorri, L.; Vaghi, L.; Castellazzi, M.; Marzola, A.; Govoni, V.; Paolino, E.; Tola, M.R.; Granieri, E.; et al. Presence of Detectable Levels of Soluble HLA-G Molecules in CSF of Relapsing–Remitting Multiple Sclerosis: Relationship with CSF Soluble HLA-I and IL-10 Concentrations and MRI Findings. J. Neuroimmunol. 2003, 142, 149–158. [Google Scholar] [CrossRef]
- Airas, L.; Nikula, T.; Huang, Y.-H.; Lahesmaa, R.; Wiendl, H. Postpartum-Activation of Multiple Sclerosis Is Associated with down-Regulation of Tolerogenic HLA-G. J. Neuroimmunol. 2007, 187, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Wiendl, H.; Feger, U.; Mittelbronn, M.; Jack, C.; Schreiner, B.; Stadelmann, C.; Antel, J.; Brueck, W.; Meyermann, R.; Bar-Or, A.; et al. Expression of the Immune-Tolerogenic Major Histocompatibility Molecule HLA-G in Multiple Sclerosis: Implications for CNS Immunity. Brain 2005, 128, 2689–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-H.; Zozulya, A.L.; Weidenfeller, C.; Metz, I.; Buck, D.; Toyka, K.V.; Brück, W.; Wiendl, H. Specific Central Nervous System Recruitment of HLA-G+ Regulatory T Cells in Multiple Sclerosis. Ann. Neurol. 2009, 66, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Waschbisch, A.; Sandbrink, R.; Hartung, H.-P.; Kappos, L.; Schwab, S.; Pohl, C.; Wiendl, H. Evaluation of Soluble HLA-G as a Biomarker for Multiple Sclerosis. Neurology 2011, 77, 596–598. [Google Scholar] [CrossRef]
- Ben Fredj, N.; Sakly, K.; Bortolotti, D.; Aissi, M.; Frih-Ayed, M.; Rotola, A.; Caselli, E.; Cura, F.; Sakly, N.; Aouni, M.; et al. The Association between Functional HLA-G 14bp Insertion/Deletion and +3142 C>G Polymorphisms and Susceptibility to Multiple Sclerosis. Immunol. Lett. 2016, 180, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, A.; Bilińska, M.; Klimczak, A.; Wagner, M.; Majorczyk, E.; Nowak, I.; Pokryszko-Dragan, A.; Kuśnierczyk, P. Association of the HLA-G Gene Polymorphism with Multiple Sclerosis in a Polish Population. Int. J. Immunogenet. 2010, 37, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Brenol, C.V.; Veit, T.D.; Chies, J.A.B.; Xavier, R.M. The Role of the HLA-G Gene and Molecule on the Clinical Expression of Rheumatologic Diseases. Rev. Bras. Reumatol. 2012, 52, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Ongaro, A.; Stignani, M.; Pellati, A.; Melchiorri, L.; Massari, L.; Caruso, G.; De Mattei, M.; Caruso, A.; Baricordi, O.R.; Rizzo, R. Human Leukocyte Antigen-G Molecules Are Constitutively Expressed by Synovial Fibroblasts and Upmodulated in Osteoarthritis. Hum. Immunol. 2010, 71, 342–350. [Google Scholar] [CrossRef] [PubMed]
- LeMaoult, J.; Le Discorde, M.; Rouas-Freiss, N.; Moreau, P.; Menier, C.; McCluskey, J.; Carosella, E.D. Biology and Functions of Human Leukocyte Antigen-G in Health and Sickness*. Tissue Antigens 2003, 62, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Prigione, I.; Penco, F.; Martini, A.; Gattorno, M.; Pistoia, V.; Morandi, F. HLA-G and HLA-E in Patients with Juvenile Idiopathic Arthritis. Rheumatology 2011, 50, 966–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbruggen, L.A.; Rebmann, V.; Demanet, C.; De Cock, S.; Grosse-Wilde, H. Soluble HLA-G in Rheumatoid Arthritis. Hum. Immunol. 2006, 67, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Catamo, E.; Addobbati, C.; Segat, L.; Sotero Fragoso, T.; Domingues Barbosa, A.; Tavares Dantas, A.; de Ataíde Mariz, H.; da Rocha, L.F.; Branco Pinto Duarte, A.L.; Monasta, L.; et al. HLA-G Gene Polymorphisms Associated with Susceptibility to Rheumatoid Arthritis Disease and Its Severity in Brazilian Patients. Tissue Antigens 2014, 84, 308–315. [Google Scholar] [CrossRef]
- Hashemi, M.; Sandoughi, M.; Fazeli, S.A.; Bahari, G.; Rezaei, M.; Zakeri, Z. Evaluation of HLA-G 14 Bp Ins/Del and +3142G>C Polymorphism with Susceptibility and Early Disease Activity in Rheumatoid Arthritis. Adv. Med. 2016, 2016, e4985745. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.; Kumar, U.; Kumar, M.; Kanga, U.; Dada, R. Association of HLA-G 3’UTR Polymorphisms with Soluble HLA-G Levels and Disease Activity in Patients with Rheumatoid Arthritis: A Case-Control Study. Immunol. Investig. 2020, 49, 88–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Bae, S.-C.; Song, G.G. Meta-Analysis of Associations between Functional HLA-G Polymorphisms and Susceptibility to Systemic Lupus Erythematosus and Rheumatoid Arthritis. Rheumatol. Int. 2015, 35, 953–961. [Google Scholar] [CrossRef]
- Mariaselvam, C.M.; Chaaben, A.B.; Salah, S.; Charron, D.; Krishnamoorthy, R.; Tamouza, R.; Negi, V.S. Human Leukocyte Antigen-G Polymorphism Influences the Age of Onset and Autoantibody Status in Rheumatoid Arthritis. Tissue Antigens 2015, 85, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Rudwaleit, M.; Yin, Z.; Siegert, S.; Grolms, M.; Radbruch, A.; Braun, J.; Sieper, J. Response to Methotrexate in Early Rheumatoid Arthritis Is Associated with a Decrease of T Cell Derived Tumour Necrosis Factor Alpha, Increase of Interleukin 10, and Predicted by the Initial Concentration of Interleukin 4. Ann. Rheum. Dis. 2000, 59, 311–314. [Google Scholar] [CrossRef]
- Rizzo, R.; Hviid, T.V.F.; Stignani, M.; Balboni, A.; Grappa, M.T.; Melchiorri, L.; Baricordi, O.R. The HLA-G Genotype Is Associated with IL-10 Levels in Activated PBMCs. Immunogenetics 2005, 57, 172–181. [Google Scholar] [CrossRef]
- Rizzo, R.; Rubini, M.; Govoni, M.; Padovan, M.; Melchiorri, L.; Stignani, M.; Carturan, S.; Ferretti, S.; Trotta, F.; Baricordi, O.R. HLA-G 14-Bp Polymorphism Regulates the Methotrexate Response in Rheumatoid Arthritis. Pharmacogenet. Genom. 2006, 16, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Stamp, L.K.; O’Donnell, J.L.; Chapman, P.T.; Barclay, M.L.; Kennedy, M.A.; Frampton, C.M.A.; Roberts, R.L. Lack of Association between HLA-G 14 Bp Insertion/Deletion Polymorphism and Response to Long-Term Therapy with Methotrexate Response in Rheumatoid Arthritis. Ann. Rheum. Dis. 2009, 68, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Kooloos, W.M.; Wessels, J.A.; van der Straaten, T.; Allaart, C.F.; Huizinga, T.W.; Guchelaar, H.-J. Functional Polymorphisms and Methotrexate Treatment Outcome in Recent-Onset Rheumatoid Arthritis. Pharmacogenomics 2010, 11, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Wastowski, I.J.; Sampaio-Barros, P.D.; Amstalden, E.M.I.; Palomino, G.M.; Marques-Neto, J.F.; Crispim, J.C.O.; Biral, A.C.; Rassi, D.M.; Carosella, E.D.; Moreau, P.; et al. HLA-G Expression in the Skin of Patients with Systemic Sclerosis. J. Rheumatol. 2009, 36, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Negrini, S.; Contini, P.; Pupo, F.; Greco, M.; Murdaca, G.; Puppo, F. Expression of Membrane-Bound Human Leucocyte Antigen-G in Systemic Sclerosis and Systemic Lupus Erythematosus. Hum. Immunol. 2020, 81, 162–167. [Google Scholar] [CrossRef]
- Mok, C.C.; Lau, C.S. Pathogenesis of Systemic Lupus Erythematosus. J. Clin. Pathol. 2003, 56, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Shen, B.; Jiang, Y.; Jun, L.; Zhu, M.; Chen, B.; Liu, C. Analysis of Immunoglobulin-like Transcripts (ILTs) in Lymphocytes with SHLA-G and IL10 from SLE Patients. Clin. Exp. Med. 2013, 13, 135–142. [Google Scholar] [CrossRef]
- Rosado, S.; Perez-Chacon, G.; Mellor-Pita, S.; Sanchez-Vegazo, I.; Bellas-Menendez, C.; Citores, M.J.; Losada-Fernandez, I.; Martin-Donaire, T.; Rebolleda, N.; Perez-Aciego, P. Expression of Human Leukocyte Antigen-G in Systemic Lupus Erythematosus. Hum. Immunol. 2008, 69, 9–15. [Google Scholar] [CrossRef]
- Rudstein-Svetlicky, N.; Loewenthal, R.; Horejsi, V.; Gazit, E. HLA-G Levels in Serum and Plasma. Tissue Antigens 2006, 67, 111–116. [Google Scholar] [CrossRef]
- Monsiváis-Urenda, A.E.; Baranda, L.; Alvarez-Quiroga, C.; Abud-Mendoza, C.; González-Amaro, R. Expression and Functional Role of HLA-G in Immune Cells from Patients with Systemic Lupus Erythematosus. J. Clin. Immunol. 2011, 31, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Fernando, M.M.A.; Freudenberg, J.; Lee, A.; Morris, D.L.; Boteva, L.; Rhodes, B.; Gonzalez-Escribano, M.F.; Lopez-Nevot, M.A.; Navarra, S.V.; Gregersen, P.K.; et al. Transancestral Mapping of the MHC Region in Systemic Lupus Erythematosus Identifies New Independent and Interacting Loci at MSH5, HLA-DPB1 and HLA-G. Ann. Rheum. Dis. 2012, 71, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Veit, T.; Cordero, E.; Mucenic, T.; Monticielo, O.; Brenol, J.; Xavier, R.; Delgado-Cañedo, A.; Chies, J. Association of the HLA-G 14 Bp Polymorphism with Systemic Lupus Erythematosus. Lupus 2009, 18, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Pedroza, L.; Sauma, M.; Vasconcelos, J.; Takeshita, L.; Ribeiro-Rodrigues, E.; Sastre, D.; Barbosa, C.; Chies, J.; Veit, T.; Lima, C.; et al. Systemic Lupus Erythematosus: Association with KIR and SLC11A1 Polymorphisms, Ethnic Predisposition and Influence in Clinical Manifestations at Onset Revealed by Ancestry Genetic Markers in an Urban Brazilian Population. Lupus 2011, 20, 265–273. [Google Scholar] [CrossRef]
- Lucena-Silva, N.; de Souza, V.S.B.; Gomes, R.G.; Fantinatti, A.; Muniz, Y.C.N.; de Albuquerque, R.S.; Monteiro, A.L.R.; Diniz, G.T.N.; Coelho, M.R.C.D.; Mendes-Junior, C.T.; et al. HLA-G 3′ Untranslated Region Polymorphisms Are Associated with Systemic Lupus Erythematosus in 2 Brazilian Populations. J. Rheumatol. 2013, 40, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Consiglio, C.R.; Veit, T.D.; Monticielo, O.A.; Mucenic, T.; Xavier, R.M.; Brenol, J.C.T.; Chies, J.A.B. Association of the HLA-G Gene +3142C>G Polymorphism with Systemic Lupus Erythematosus. Tissue Antigens 2011, 77, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Hamzaoui, K.; Hamzaoui, A. Immunological Responses in Patients with Behçet’s Disease: Advances in Understanding. Expert Rev. Ophthalmol. 2012, 7, 261–270. [Google Scholar] [CrossRef]
- Kaya, T.İ. Genetics of Behçet’s Disease. Pathol. Res. Int. 2011, 2012, e912589. [Google Scholar] [CrossRef]
- Kaya, T.I.; Tursen, U.; Gurler, A.; Dur, H. Association of Class I HLA Antigens with the Clinical Manifestations of Turkish Patients with Behçet’s Disease. Clin. Exp. Dermatol. 2002, 27, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Fietta, P. Behçet’s Disease: Familial Clustering and Immunogenetics. Clin. Exp. Rheumatol. 2005, 23, S96–105. [Google Scholar] [PubMed]
- Mizuki, N.; Ohno, S.; Ando, H.; Chen, L.; Palimeris, G.D.; Stavropoulos-Ghiokas, E.; Ishihara, M.; Goto, K.; Nakamura, S.; Shindo, Y.; et al. A Strong Association between HLA-B*5101 and Behçet’s Disease in Greek Patients. Tissue Antigens 1997, 50, 57–60. [Google Scholar] [CrossRef]
- Sakly, K.; Maatouk, M.; Hammami, S.; Harzallah, O.; Sakly, W.; Feki, S.; Mirshahi, M.; Ghedira, I.; Sakly, N. HLA-G 14bp Insertion/Deletion Polymorphism and Its Association with SHLA-G Levels in Behçet’s Disease Tunisian Patients. Hum. Immunol. 2016, 77, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Nam, J.H.; Lee, E.S.; Choi, J.S.; Bang, D.; Lee, S. Increased Risk of Human Leukocyte Antigen-G Gene Variants in Behçet’s Disease. Clin. Exp. Rheumatol. 2006, 24, S126–S127. [Google Scholar] [PubMed]
- Park, K.S.; Park, J.S.; Nam, J.H.; Bang, D.; Sohn, S.; Lee, E.S. HLA-E*0101 and HLA-G*010101 Reduce the Risk of Behcet’s Disease. Tissue Antigens 2007, 69, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Newburger, J.W.; Takahashi, M.; Burns, J.C. Kawasaki Disease. J. Am. Coll. Cardiol. 2016, 67, 1738–1749. [Google Scholar] [CrossRef] [PubMed]
- Onouchi, Y. Molecular Genetics of Kawasaki Disease. Pediatr. Res. 2009, 65, 46–54. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Diseases and Injuries Collaborators. Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990-2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet Lond. Engl. 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Nicolae, D.; Cox, N.J.; Lester, L.A.; Schneider, D.; Tan, Z.; Billstrand, C.; Kuldanek, S.; Donfack, J.; Kogut, P.; Patel, N.M.; et al. Fine Mapping and Positional Candidate Studies Identify HLA-G as an Asthma Susceptibility Gene on Chromosome 6p21. Am. J. Hum. Genet. 2005, 76, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Tahan, F.; Patıroglu, T. Plasma Soluble Human Leukocyte Antigen G Levels in Asthmatic Children. Int. Arch. Allergy Immunol. 2006, 141, 213–216. [Google Scholar] [CrossRef]
- Zheng, X.-Q.; Li, C.-C.; Xu, D.-P.; Lin, A.; Bao, W.-G.; Yang, G.-S.; Yan, W.-H. Analysis of the Plasma Soluble Human Leukocyte Antigen-G and Interleukin-10 Levels in Childhood Atopic Asthma. Hum. Immunol. 2010, 71, 982–987. [Google Scholar] [CrossRef]
- Ribeyre, C.; Carlini, F.; René, C.; Jordier, F.; Picard, C.; Chiaroni, J.; Abi-Rached, L.; Gouret, P.; Marin, G.; Molinari, N.; et al. HLA-G Haplotypes Are Differentially Associated with Asthmatic Features. Front. Immunol. 2018, 9, 278. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.; Randall, G.; Fan, J.; Camoretti-Mercado, B.; Brockman-Schneider, R.; Pan, L.; Solway, J.; Gern, J.E.; Lemanske, R.F.; Nicolae, D.; et al. Allele-Specific Targeting of MicroRNAs to HLA-G and Risk of Asthma. Am. J. Hum. Genet. 2007, 81, 829–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, R.; Mapp, C.E.; Melchiorri, L.; Maestrelli, P.; Visentin, A.; Ferretti, S.; Bononi, I.; Miotto, D.; Baricordi, O.R. Defective Production of Soluble HLA-G Molecules by Peripheral Blood Monocytes in Patients with Asthma. J. Allergy Clin. Immunol. 2005, 115, 508–513. [Google Scholar] [CrossRef]
- Mapp, C.E.; Ferrazzoni, S.; Rizzo, R.; Miotto, D.; Stignani, M.; Boschetto, P.; Maestrelli, P.; Baricordi, O.R. Soluble Human Leucocyte Antigen-G and Interleukin-10 Levels in Isocyanate-Induced Asthma. Clin. Exp. Allergy 2009, 39, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Umetsu, D.T.; DeKruyff, R.H. The Regulation of Allergy and Asthma. Immunol. Rev. 2006, 212, 238–255. [Google Scholar] [CrossRef]
- Bousquet, J.; Anto, J.M.; Bachert, C.; Baiardini, I.; Bosnic-Anticevich, S.; Walter Canonica, G.; Melén, E.; Palomares, O.; Scadding, G.K.; Togias, A.; et al. Allergic Rhinitis. Nat. Rev. Dis. Primers 2020, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. Biology of Human TH1 and TH2 Cells. J. Clin. Immunol. 1995, 15, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Contini, P.; Negrini, S.; Ciprandi, G.; Puppo, F. Immunoregulatory Role of HLA-G in Allergic Diseases. J. Immunol. Res. 2016, 2016, 6865758. [Google Scholar] [CrossRef] [Green Version]
- Ciprandi, G.; Colombo, B.M.; Contini, P.; Cagnati, P.; Pistorio, A.; Puppo, F.; Murdaca, G. Soluble HLA-G and HLA-A,-B,-C Serum Levels in Patients with Allergic Rhinitis. Allergy 2008, 63, 1335–1338. [Google Scholar] [CrossRef] [PubMed]
- Ciprandi, G.; Contini, P.; Murdaca, G.; DeAmici, M.; Gallina, A.M.; Puppo, F. Soluble Serum HLA-G and HLA-A, -B, -C Molecules in Patients with Seasonal Allergic Rhinitis Exposed to Pollens. Int. Immunopharmacol. 2009, 9, 1058–1062. [Google Scholar] [CrossRef]
- Ciprandi, G.; Contini, P.; Murdaca, G.; Gallina, A.M.; Puppo, F. Soluble HLA-G Molecule in Patients with Perennial Allergic Rhinitis. Int. Arch. Allergy Immunol. 2009, 150, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Ciprandi, G.; Contini, P.; Pistorio, A.; Murdaca, G.; Puppo, F. Sublingual Immunotherapy Reduces Soluble HLA-G and HLA-A,-B,-C Serum Levels in Patients with Allergic Rhinitis. Int. Immunopharmacol. 2009, 9, 253–257. [Google Scholar] [CrossRef]
- Ciprandi, G.; De Amici, M.; Caimmi, S.; Marseglia, A.; Marchi, A.; Castellazzi, A.M.; Marseglia, G. Soluble Serum HLA-G in Children with Allergic Rhinitis and Asthma. J. Biol. Regul. Homeost. Agents 2010, 24, 221–224. [Google Scholar]
- Ciprandi, G.; Corsico, A.; Pisati, P. Serum-Soluble HLA-G Is Associated with Specific IgE in Patients with Allergic Rhinitis and Asthma. Inflammation 2014, 37, 1630–1634. [Google Scholar] [CrossRef]
- Wyatt, R.C.; Lanzoni, G.; Russell, M.A.; Gerling, I.; Richardson, S.J. What the HLA-I!—Classical and Non-Classical HLA Class I and Their Potential Roles in Type 1 Diabetes. Curr. Diab. Rep. 2019, 19, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirulli, V.; Zalatan, J.; McMaster, M.; Prinsen, R.; Salomon, D.R.; Ricordi, C.; Torbett, B.E.; Meda, P.; Crisa, L. The Class I HLA Repertoire of Pancreatic Islets Comprises the Nonclassical Class Ib Antigen HLA-G. Diabetes 2006, 55, 1214–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommese, L.; Benincasa, G.; Schiano, C.; Marfella, R.; Grimaldi, V.; Sorriento, A.; Lucchese, R.; Fiorito, C.; Sardu, C.; Nicoletti, G.F.; et al. Genetic and Epigenetic-Sensitive Regulatory Network in Immune Response: A Putative Link between HLA-G and Diabetes. Expert Rev. Endocrinol. Metab. 2019, 14, 233–241. [Google Scholar] [CrossRef]
- Amodio, G.; Mandelli, A.; Curto, R.; Rancoita, P.M.V.; Stabilini, A.; Bonfanti, R.; de Pellegrin, M.; Bosi, E.; Di Serio, C.; Battaglia, M.; et al. Altered Frequency and Phenotype of HLA-G-Expressing DC-10 in Type 1 Diabetes Patients at Onset and in Subjects at Risk to Develop the Disease. Front. Immunol. 2021, 12, 3874. [Google Scholar] [CrossRef]
- Silva, H.P.V.; Ururahy, M.A.G.; Souza, K.S.C.; Loureiro, M.B.; Oliveira, Y.M.C.; Oliveira, G.H.M.; Luchessi, A.D.; Carvalho, K.T.C.; Freitas, J.C.O.C.; Donadi, E.A.; et al. The Association between the HLA-G 14-Bp Insertion/Deletion Polymorphism and Type 1 Diabetes. Genes Immun. 2016, 17, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Gerasimou, P.; Skordis, N.; Picolos, M.; Spyridonidis, A.; Costeas, P. HLA-G 14-Bp Polymorphism Affects the Age of Onset in Type I Diabetes Mellitus. Int. J. Immunogenet. 2016, 43, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Solini, A.; Muscelli, E.; Stignani, M.; Melchiorri, L.; Santini, E.; Rossi, C.; Astiarraga, B.D.; Rizzo, R.; Baricordi, O.R. Soluble Human Leukocyte Antigen-G Expression and Glucose Tolerance in Subjects with Different Degrees of Adiposity. J. Clin. Endocrinol. Metab. 2010, 95, 3342–3346. [Google Scholar] [CrossRef] [Green Version]
- Shobeiri, S.; Abediankenari, S.; Lashtoo-Aghaee, B.; Rahmani, Z.; Esmaeili-gorji, B. Evaluation of Soluble Human Leukocyte Antigen-G in Peripheral Blood of Pregnant Women with Gestational Diabetes Mellitus. Casp. J. Intern. Med. 2016, 7, 178–182. [Google Scholar]
- Abdel Hameed, M.R.; Ibrahiem, O.A.; Ahmed, E.H.; Sedky, P.R.; Mousa, N.M.M.A. Soluble Human Leukocyte Antigen-G Evaluation in Pregnant Women with Gestational Diabetes Mellitus. Egypt. J. Intern. Med. 2020, 32, 7. [Google Scholar] [CrossRef]
- Oztekin, O.; Cabus, U.; Enli, Y. Decreased Serum Human Leukocyte Antigen-G Levels Are Associated with Gestational Diabetes Mellitus. J. Obstet. Gynaecol. Res. 2021, 47, 2329–2337. [Google Scholar] [CrossRef] [PubMed]
- Kowalska-Kępczyńska, A.; Mleczko, M.; Domerecka, W.; Mazurek, M.; Krasowska, D.; Małecka-Massalska, T.; Donica, H. Extended Inflammation Parameters (EIP) as Markers of Immune System Cell Activation in Psoriasis. Int. J. Inflamm. 2021, 2021, e9216528. [Google Scholar] [CrossRef] [PubMed]
- Yendo, T.M.; Sato, M.N.; Branco, A.C.C.C.; Pietrobon, A.J.; Teixeira, F.M.E.; Ramos, Y.Á.L.; Alberca, R.W.; Valêncio, C.G.; Arruda, V.N.; Romiti, R.; et al. Impact of Inflammatory Immune Dysfunction in Psoriasis Patients at Risk for COVID-19. Vaccines 2021, 9, 478. [Google Scholar] [CrossRef] [PubMed]
- Prinz, J.C. Human Leukocyte Antigen-Class I Alleles and the Autoreactive T Cell Response in Psoriasis Pathogenesis. Front. Immunol. 2018, 9, 954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, R.P.; Stuart, P.E.; Nistor, I.; Hiremagalore, R.; Chia, N.V.C.; Jenisch, S.; Weichenthal, M.; Abecasis, G.R.; Lim, H.W.; Christophers, E.; et al. Sequence and Haplotype Analysis Supports HLA-C as the Psoriasis Susceptibility 1 Gene. Am. J. Hum. Genet. 2006, 78, 827–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Cao, H.; Zuo, X.; Zhang, T.; Zhang, X.; Liu, X.; Xu, R.; Chen, G.; Zhang, Y.; Zheng, X.; et al. Deep Sequencing of the MHC Region in the Chinese Population Contributes to Studies of Complex Disease. Nat. Genet. 2016, 48, 740–746. [Google Scholar] [CrossRef] [PubMed]
- De Freire Cassia, F.; Cardoso, J.F.; Porto, L.C.; Ramos-E-Silva, M.; Carneiro, S. Association of HLA Alleles and HLA Haplotypes with Psoriasis, Psoriatic Arthritis and Disease Severity in a Miscegenated Population. Psoriasis 2021, 11, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Borghi, A.; Fogli, E.; Stignani, M.; Melchiorri, L.; Altieri, E.; Baricordi, O.; Rizzo, R.; Virgili, A. Soluble Human Leukocyte Antigen-G and Interleukin-10 Levels in Plasma of Psoriatic Patients: Preliminary Study on a Possible Correlation between Generalized Immune Status, Treatments and Disease. Arch. Dermatol. Res. 2008, 300, 551–559. [Google Scholar] [CrossRef]
- Forrester, J.V.; Kuffova, L.; Dick, A.D. Autoimmunity, Autoinflammation, and Infection in Uveitis. Am. J. Ophthalmol. 2018, 189, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Amin, S.; Lung, K.I.; Seabury, S.; Rao, N.; Toy, B.C. Incidence, Prevalence, and Risk Factors of Infectious Uveitis and Scleritis in the United States: A Claims-Based Analysis. PLoS ONE 2020, 15, e0237995. [Google Scholar] [CrossRef] [PubMed]
- Neti, N.; Pimsri, A.; Boonsopon, S.; Tesavibul, N.; Choopong, P. Triggering Factors Associated with a New Episode of Recurrent Acute Anterior Uveitis. Sci. Rep. 2021, 11, 12156. [Google Scholar] [CrossRef]
- Takeuchi, M.; Mizuki, N.; Ohno, S. Pathogenesis of Non-Infectious Uveitis Elucidated by Recent Genetic Findings. Front. Immunol. 2021, 12, 1150. [Google Scholar] [CrossRef] [PubMed]
- Wildner, G.; Diedrichs-Möhring, M. Molecular Mimicry and Uveitis. Front. Immunol. 2020, 11, 2847. [Google Scholar] [CrossRef]
- Huang, X.-F.; Li, Z.; De Guzman, E.; Robinson, P.; Gensler, L.; Ward, M.M.; Rahbar, M.H.; Lee, M.; Weisman, M.H.; Macfarlane, G.J.; et al. Genomewide Association Study of Acute Anterior Uveitis Identifies New Susceptibility Loci. Investig. Ophthalmol. Vis. Sci. 2020, 61, 3. [Google Scholar] [CrossRef]
- Sanjanwala, B.; Draghi, M.; Norman, P.J.; Guethlein, L.A.; Parham, P. Polymorphic Sites Away from the Bw4 Epitope That Affect Interaction of Bw4+ HLA-B with KIR3DL1. J. Immunol. 2008, 181, 6293–6300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crabtree, E.; Song, L.; Llanga, T.; Bower, J.J.; Cullen, M.; Salmon, J.H.; Hirsch, M.L.; Gilger, B.C. AAV-Mediated Expression of HLA-G1/5 Reduces Severity of Experimental Autoimmune Uveitis. Sci. Rep. 2019, 9, 19864. [Google Scholar] [CrossRef]


| Receptor | Gene | Protein Mass (kDa) | The Number of Amino Acids | Expression | Interactions | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| NK | CD4+ T Cell | CD8+ T Cell | B Cell | Monocytes | Macrophage | DC | |||||
| ILT2 | LILRB1 | 65,039 | 598 | some | some | some | + | + | + | + | α3+β2m (dimer) |
| ILT4 | LILRB2 | 44,601 | 400 | - | - | - | - | + | + | + | α3 (dimer) |
| KIR2DL4 | KIR2DL4 | 41,487 | 377 | + | some | - | - | - | - | - | α1 |
| CD8 | CD8B | 21,524 | 192 | some | + | - | - | - | - | - | α3 |
| CD160 | CD160 | 19,810 | 181 | + | + | + | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaborek-Łyczba, M.; Łyczba, J.; Mertowska, P.; Mertowski, S.; Hymos, A.; Podgajna, M.; Niedźwiedzka-Rystwej, P.; Grywalska, E. The HLA-G Immune Checkpoint Plays a Pivotal Role in the Regulation of Immune Response in Autoimmune Diseases. Int. J. Mol. Sci. 2021, 22, 13348. https://doi.org/10.3390/ijms222413348
Zaborek-Łyczba M, Łyczba J, Mertowska P, Mertowski S, Hymos A, Podgajna M, Niedźwiedzka-Rystwej P, Grywalska E. The HLA-G Immune Checkpoint Plays a Pivotal Role in the Regulation of Immune Response in Autoimmune Diseases. International Journal of Molecular Sciences. 2021; 22(24):13348. https://doi.org/10.3390/ijms222413348
Chicago/Turabian StyleZaborek-Łyczba, Monika, Jakub Łyczba, Paulina Mertowska, Sebastian Mertowski, Anna Hymos, Martyna Podgajna, Paulina Niedźwiedzka-Rystwej, and Ewelina Grywalska. 2021. "The HLA-G Immune Checkpoint Plays a Pivotal Role in the Regulation of Immune Response in Autoimmune Diseases" International Journal of Molecular Sciences 22, no. 24: 13348. https://doi.org/10.3390/ijms222413348