
Transcriptome Analysis Reveals Sexual Disparities between Olfactory and Immune Gene Expression in the Olfactory Epithelium of Megalobrama amblycephala


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
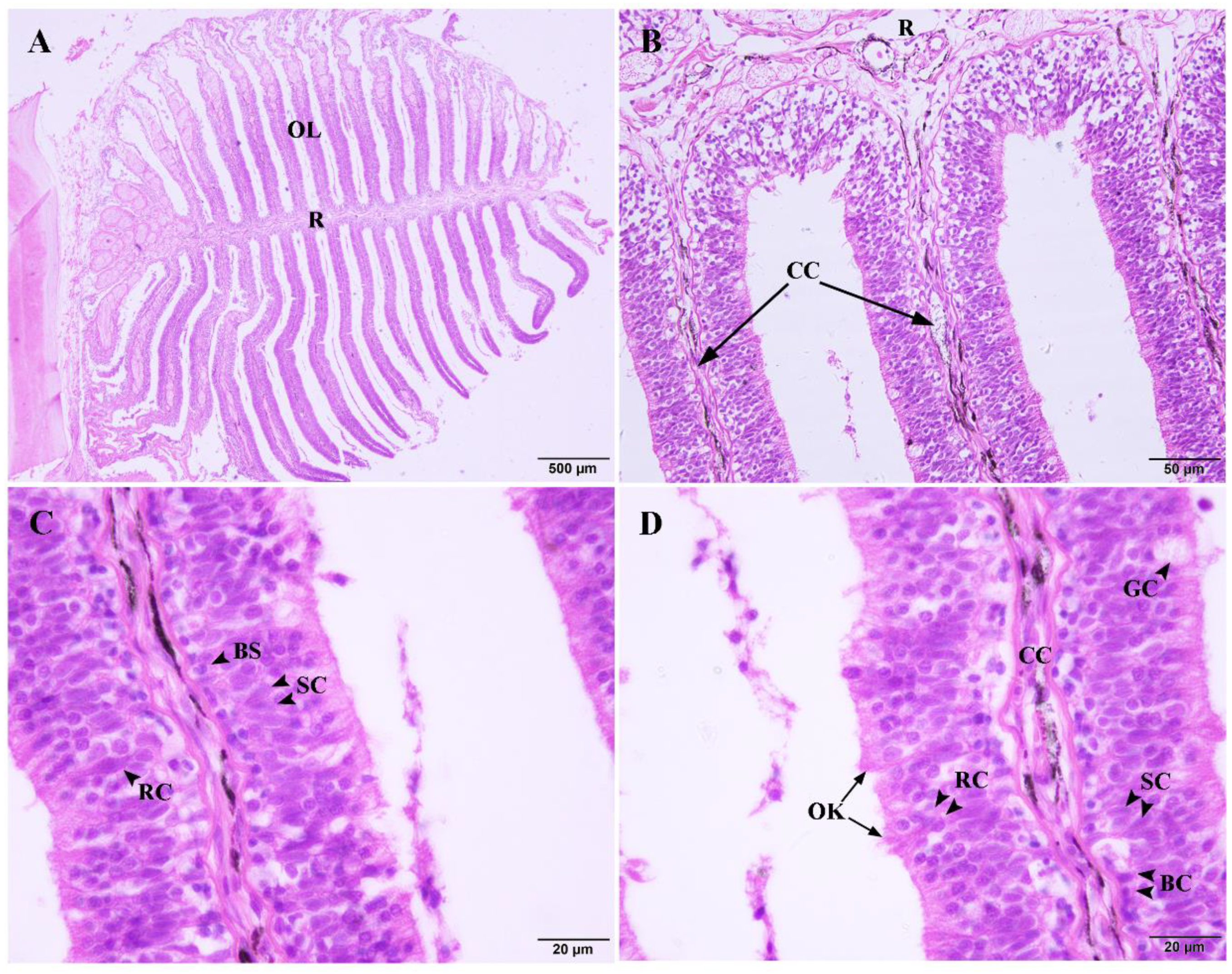
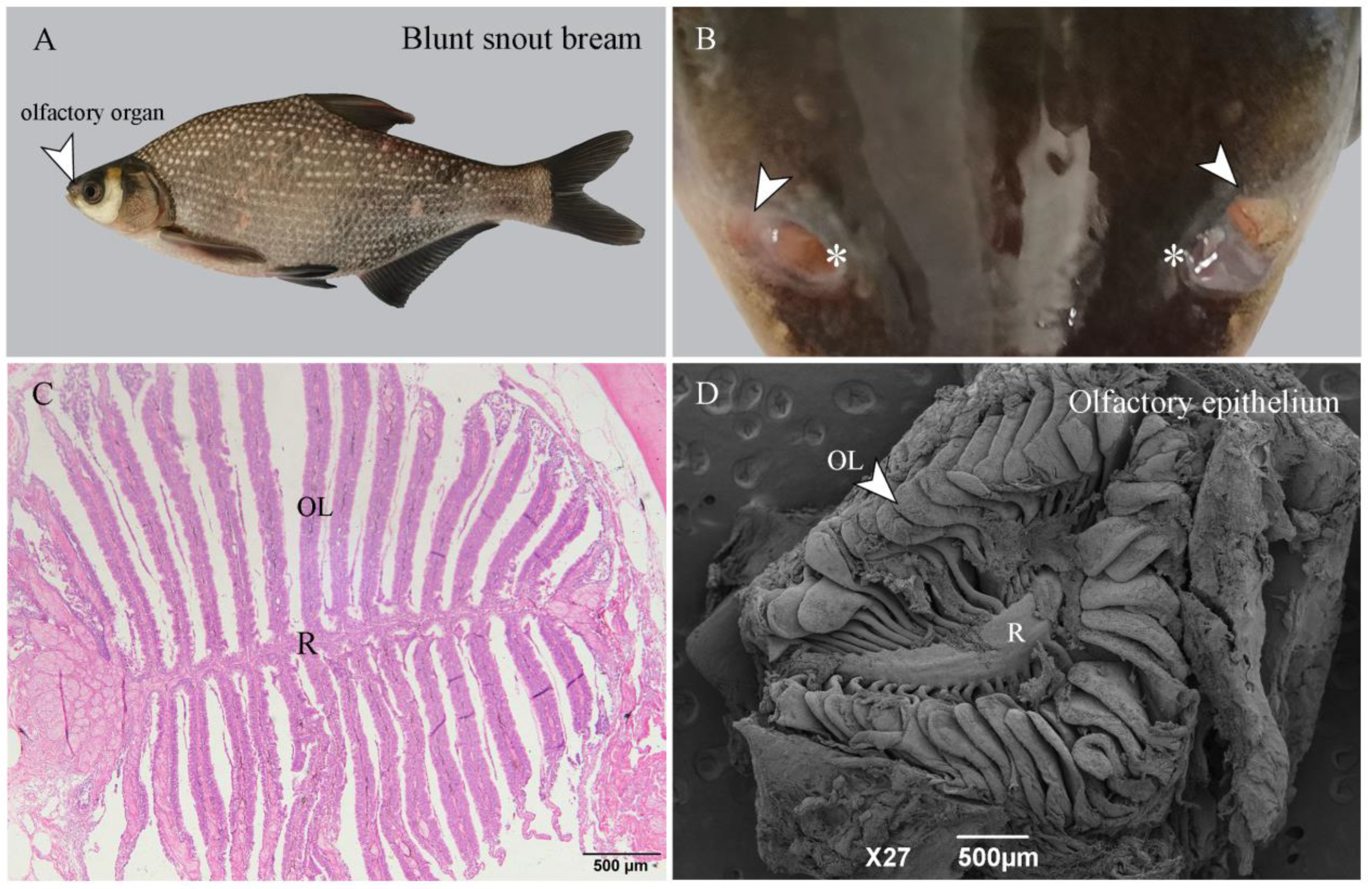
2.1. The Structure of the Olfactory Epithelium
2.2. The SEM of the Olfactory Epithelium
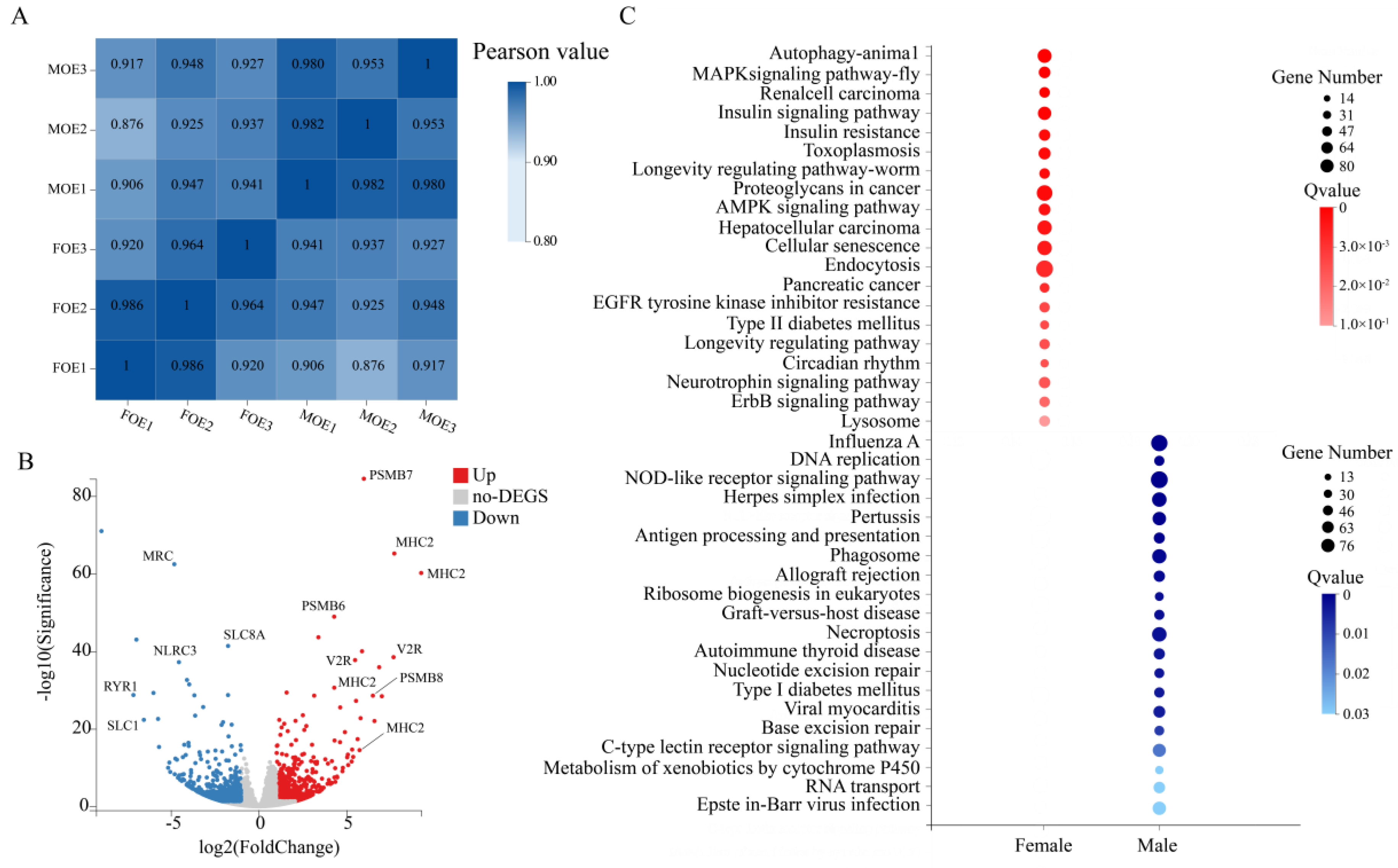
2.3. Transcriptome Analysis
2.4. Differentially Expressed Genes
2.5. The Olfactory Receptor Repertoires
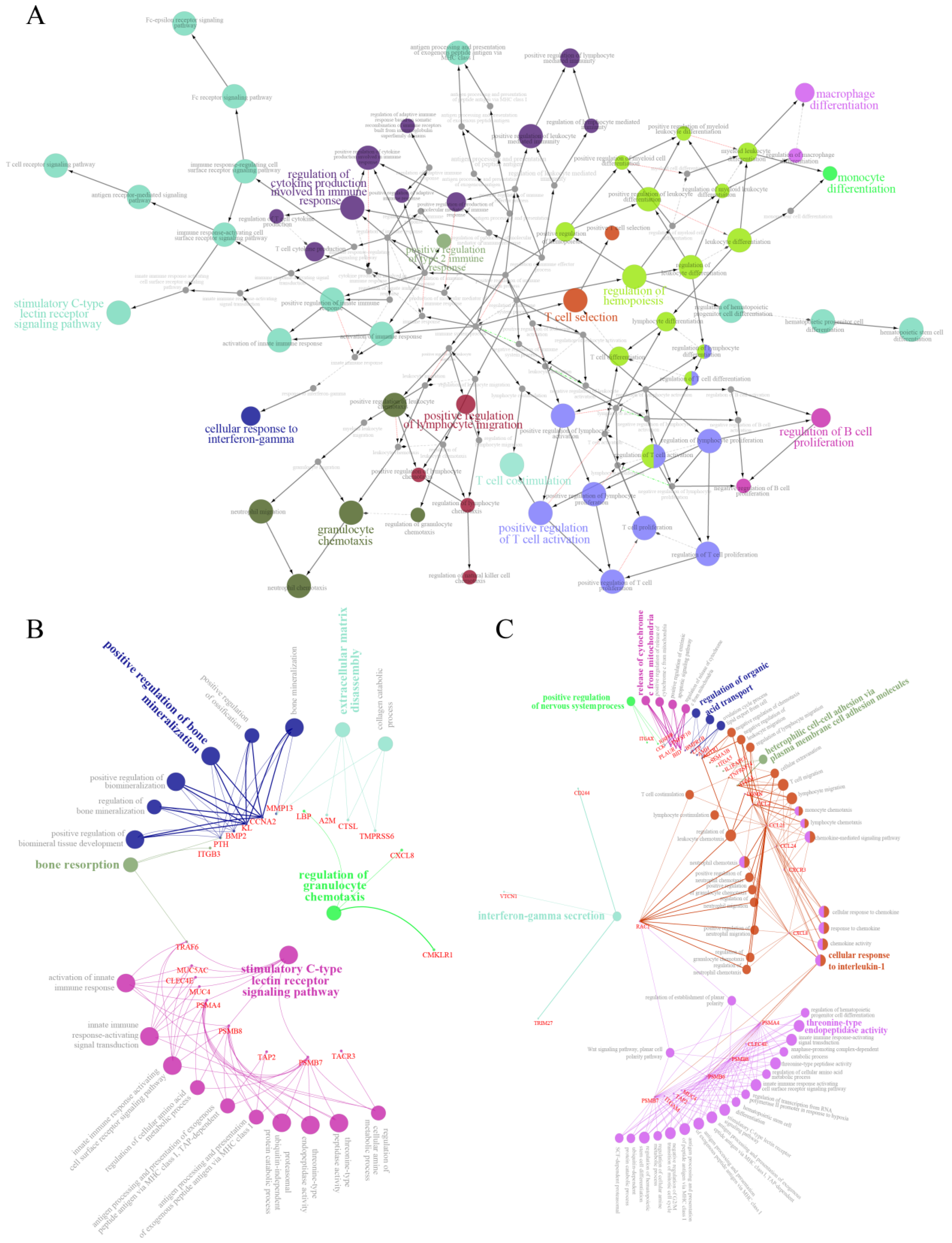
2.6. Regulatory Network of Immune Genes
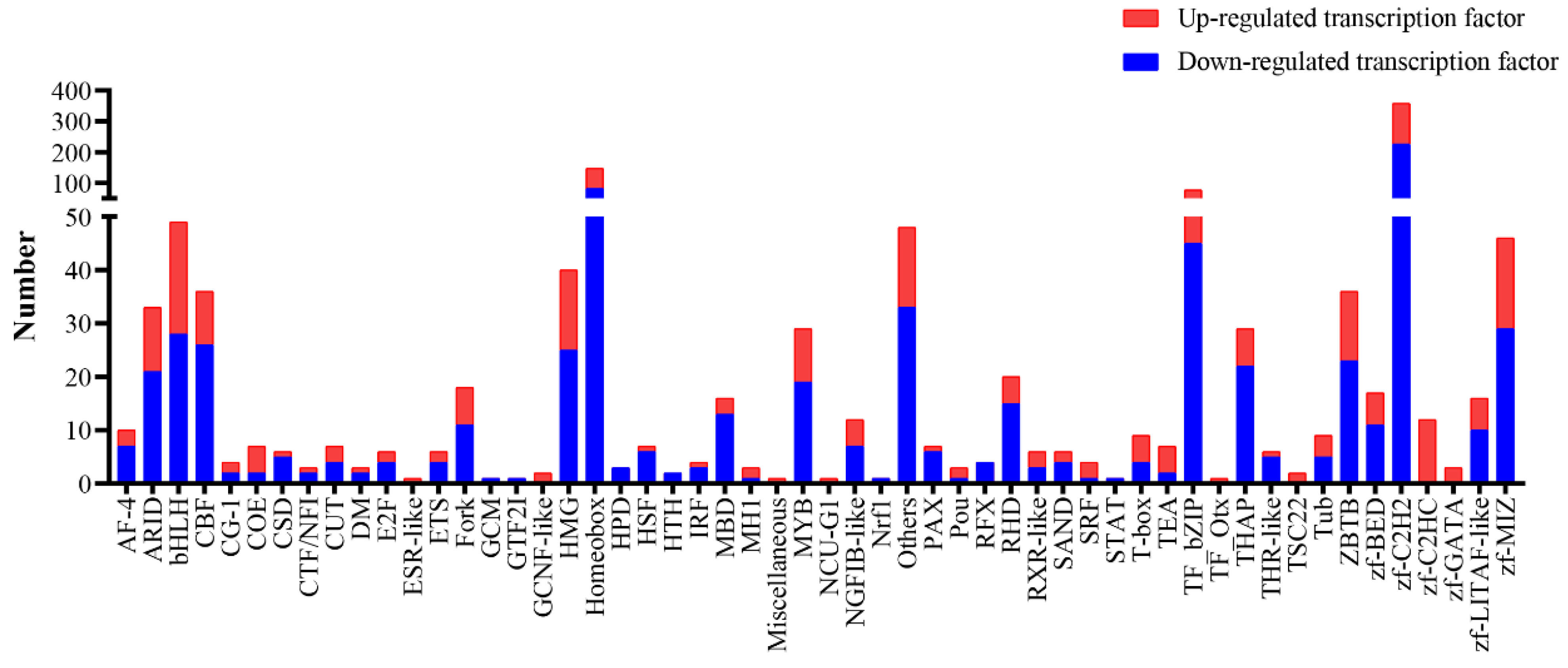
2.7. Analysis of Differentially Expressed Transcription Factors
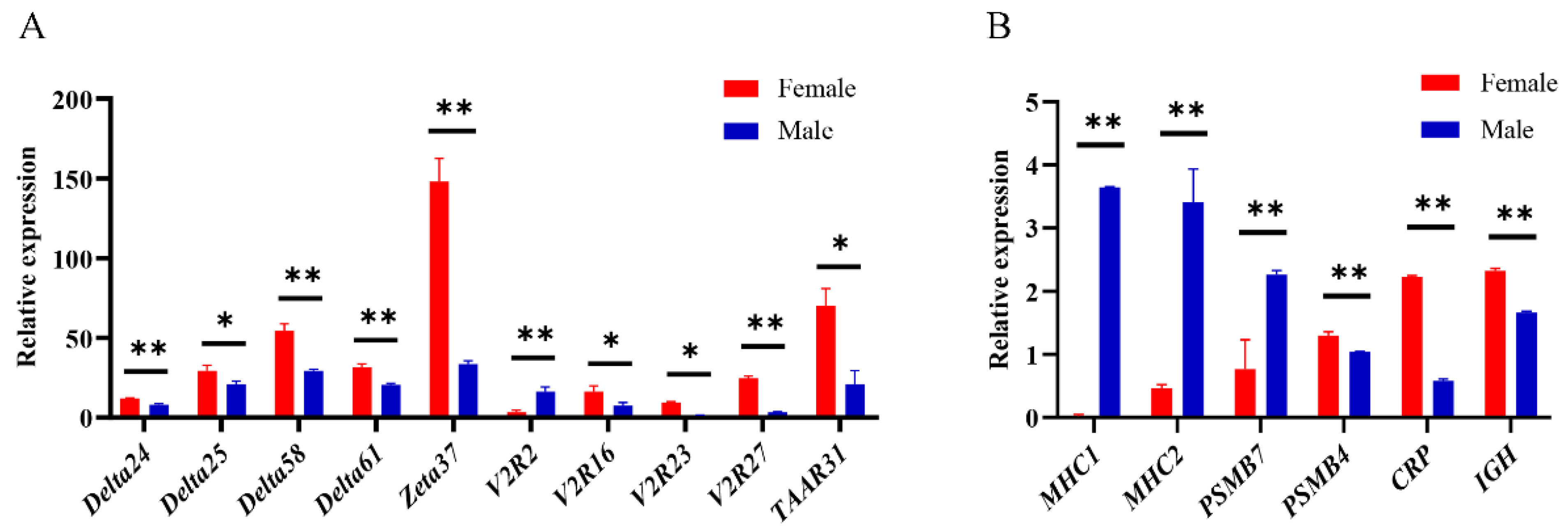
2.8. Quantitative Real-Time PCR (qPCR)
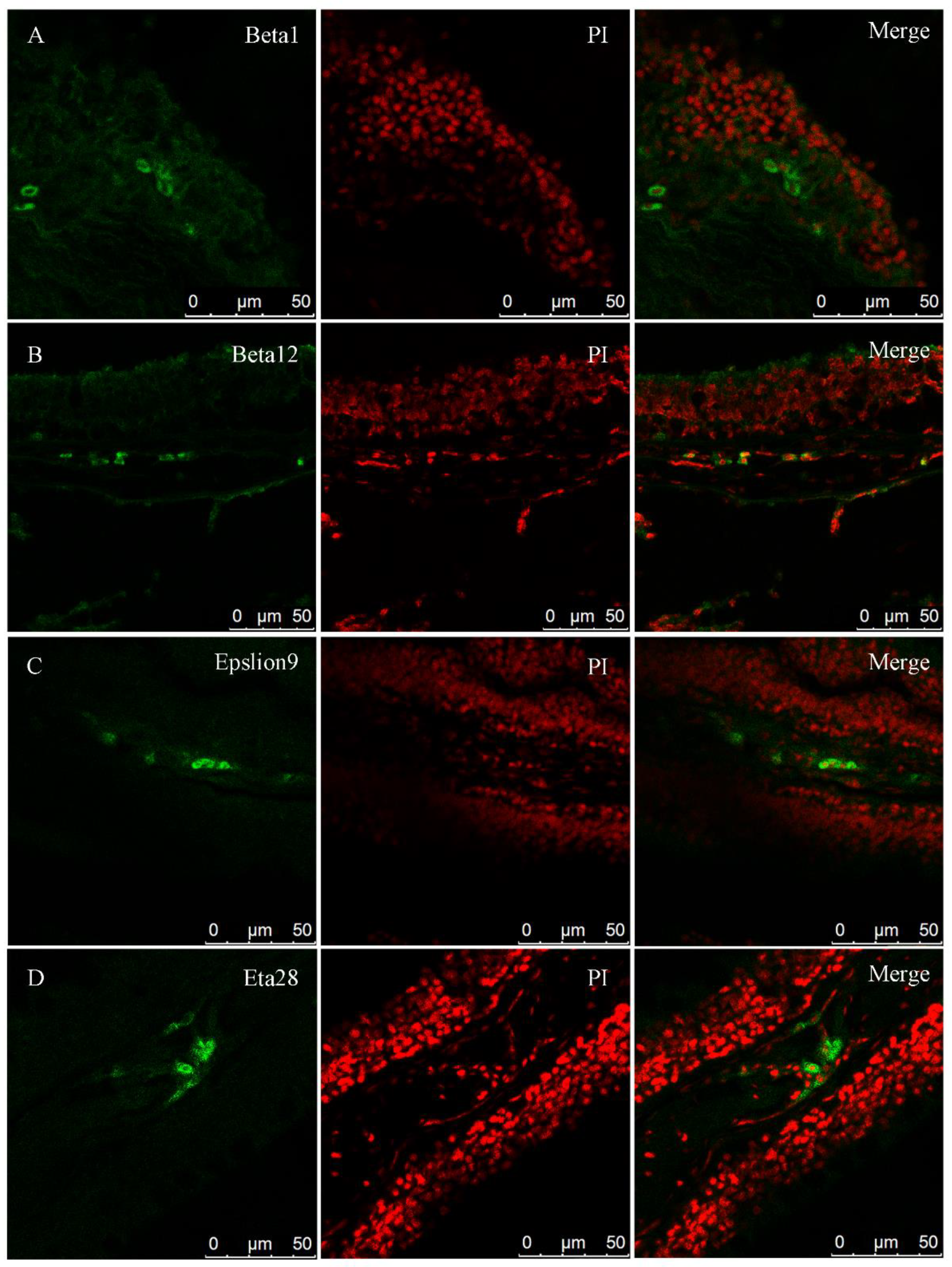
2.9. Fluorescence In Situ Hybridization
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. RNA Extraction and BGISEQ Library Construction
4.3. Analysis of RNA-Seq Data
4.4. Data Mining for the Olfactory Receptor Genes
4.5. Gene Network Analyses
4.6. Quantitative Real-Time PCR (qPCR)
4.7. Fluorescence In Situ Hybridization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Dulac, C.; Axel, R. A novel family of genes encoding putative pheromone receptors in mammals. Cell 1995, 83, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Ryba, N.J.P.; Tirindelli, R. A new multigene family of putative pheromone receptors. Neuron 1997, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Liberles, S.D.; Buck, L.B. A second class of chemosensory receptors in the olfactory epithelium. Nature 2006, 442, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Rivière, S.; Challet, L.; Fluegge, D.; Spehr, M.; Rodriguez, I. Formyl peptide receptor-like proteins are a novel family of vomeronasal chemosensors. Nature 2009, 459, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Niimura, Y.; Matsui, A.; Touhara, K. Extreme expansion of the olfactory receptor gene repertoire in African elephants and evolutionary dynamics of orthologous gene groups in 13 placental mammals. Genome Res. 2014, 24, 1485–1496. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Du, K.; Gan, X.; Yang, L.; He, S. Massive loss of olfactory receptors but not trace amine-associated receptors in the world’s deepest-living fish (Pseudoliparis swirei). Genes 2019, 10, 910. [Google Scholar] [CrossRef] [Green Version]
- Boschat, C.; Pélofi, C.; Randin, O.; Roppolo, D.; Lüscher, C.; Broillet, M.C.; Rodriguez, I. Pheromone detection mediated by a V1r vomeronasal receptor. Nat. Neurosci. 2002, 5, 1261–1262. [Google Scholar] [CrossRef]
- Sorensen, P.W.; Fine, J.M.; Dvornikovs, V.; Jeffrey, C.S.; Shao, F.; Wang, J.; Vrieze, L.A.; Anderson, K.R.; Hoye, T.R. Mixture of new sulfated steroids functions as a migratory pheromone in the sea lamprey. Nat. Chem. Biol. 2005, 1, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.A.; Stacey, N.E.; Corkum, L.D. Putative steroidal pheromones in the round goby, Neogobius melanostomus: Olfactory and behavioral responses. J. Chem. Ecol. 2001, 24, 443–470. [Google Scholar] [CrossRef]
- Behrens, M.; Frank, O.; Rawel, H.; Ahuja, G.; Potting, C.; Hofmann, T.; Meyerhof, W.; Korsching, S. ORA1, a zebrafish olfactory receptor ancestral to all mammalian V1R genes, recognizes 4-hydroxyphenylacetic acid, a putative reproductive pheromone. J. Biol. Chem. 2014, 289, 19778–19788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, A.; Rolen, S.H.; Anderson, K.; Morita, Y.; Caprio, J.; Finger, T.E. Correlation between olfactory receptor cell type and function in the channel catfish. J. Neurosci. 2003, 23, 9328–9339. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, H.; Haga, S.; Sato, K.; Touhara, K. Sex-specific peptides from exocrine glands stimulate mouse vomeronasal sensory neurons. Nature 2005, 437, 898–901. [Google Scholar] [CrossRef] [PubMed]
- Milinski, M.; Griffiths, S.; Wegner, K.M.; Reusch, T.B.H.; Haas-Assenbaum, A.; Boehm, T. Mate choice decisions of stickleback females predictably modified by MHC peptide ligands. Proc. Natl. Acad. Sci. USA 2005, 102, 4414–4418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belal, A.A.; El-Gohery, Y.; Talaat, M. Nasal and paranasal pathology in experimental bilharziasis. J. Laryngol. Otol. 1977, 91, 391–400. [Google Scholar] [CrossRef]
- Kelemen, G. The junction of the nasal cavity and the pharyngeal tube in the rat. Arch. Otolaryngol. 1947, 45, 159–168. [Google Scholar] [CrossRef]
- Harkema, J.R.; Plopper, C.G.; Hyde, D.M.; St. George, J.A. Regional differences in quantities of histochemically detectable mucosubstances in nasal, paranasal, and nasopharyngeal epithelium of the bonnet monkey. J. Histochem. Cytochem. 1987, 35, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Mair, T.S.; Batten, E.H.; Stokes, C.R.; Bourne, F.J. The histological features of the immune system of the equine respiratory tract. J. Comp. Pathol. 1987, 97, 575–586. [Google Scholar] [CrossRef]
- Yu, Y.Y.; Kong, W.; Yin, Y.X.; Dong, F.; Huang, Z.Y.; Yin, G.M.; Dong, S.; Salinas, I.; Zhang, Y.A.; Xu, Z. Mucosal immunoglobulins protect the olfactory organ of teleost fish against parasitic infection. PLoS Pathog. 2018, 14, e1007251. [Google Scholar] [CrossRef] [Green Version]
- Das, P.K.; Salinas, I. Fish nasal immunity: From mucosal vaccines to neuroimmunology. Fish Shellfish Immunol. 2020, 104, 165–171. [Google Scholar] [CrossRef]
- Sorokowski, P.; Karwowski, M.; Misiak, M.; Marczak, M.K.; Dziekan, M.; Hummel, T.; Sorokowska, A. Sex differences in human olfaction: A meta-analysis. Front. Psychol. 2019, 10, 242. [Google Scholar] [CrossRef] [Green Version]
- Haga, S.; Hattori, T.; Sato, T.; Sato, K.; Matsuda, S.; Kobayakawa, R.; Sakano, H.; Yoshihara, Y.; Kikusui, T.; Touhara, K. The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature 2010, 466, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Simpson, D.M.; Armstrong, S.D.; Davidson, A.J.; Robertson, D.H.; McLean, L.; Beynon, R.J.; Hurst, J.L. Darcin: A male pheromone that stimulates female memory and sexual attraction to an individual male’s odour. BMC Biol. 2010, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Doty, R.L.; Cameron, E.L. Sex differences and reproductive hormone influences on human odor perception. Physiol. Behav. 2009, 97, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, C.; Gao, Z.; Min, J.; Gu, Y.; Jian, J.; Jiang, X.; Cai, H.; Ebersberger, I.; Xu, M.; et al. The draft genome of blunt snout bream (Megalobrama amblycephala) reveals the development of intermuscular bone and adaptation to herbivorous diet. Gigascience 2017, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, C.; Lv, M.; Liu, N.; Hu, Y.; Zhang, H.; Enbody, E.D.; Gao, Z.; Andersson, L.; Wang, W. A chromosome-level assembly of blunt snout bream (Megalobrama amblycephala) reveals an expansion of olfactory receptor genes in freshwater fish. Mol. Biol. Evol. 2021, 38, 4238–4251. [Google Scholar] [CrossRef] [PubMed]
- Alioto, T.S.; Ngai, J. The odorant receptor repertoire of teleost fish. BMC Genom. 2005, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yan, X.; Xu, S.; Zhu, P.; He, X.; Liu, J. Family structure and phylogenetic analysis of odorant receptor genes in the large yellow croaker (Larimichthys crocea). BMC Evol. Biol. 2011, 11, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.Y.; Liang, X.F.; He, S. Genome-wide identification and characterization of olfactory receptor genes in chinese perch, Siniperca chuatsi. Genes (Basel) 2019, 10, 178. [Google Scholar] [CrossRef] [Green Version]
- Lastein, S.; Hamdani, E.H.; Døving, K.B. Gender distinction in neural discrimination of sex pheromones in the olfactory bulb of Crucian carp, Carassius carassius. Chem. Senses 2006, 31, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jiang, H.; Yang, L. Transcriptome analysis of zebrafish olfactory epithelium reveal sexual differences in odorant detection. Genes (Basel) 2020, 15, 592. [Google Scholar] [CrossRef]
- Olender, T.; Keydar, I.; Pinto, J.M.; Tatarskyy, P.; Alkelai, A.; Chien, M.S.; Fishilevich, S.; Restrepo, D.; Matsunami, H.; Gilad, Y.; et al. The human olfactory transcriptome. BMC Genom. 2016, 17, 619. [Google Scholar] [CrossRef] [Green Version]
- Shiao, M.S.; Chang, A.Y.F.; Liao, B.Y.; Ching, Y.H.; Lu, M.Y.J.; Chen, S.M.; Li, W.H. Transcriptomes of mouse olfactory epithelium reveal sexual differences in odorant. Genome Biol. Evol. 2012, 4, 703–712. [Google Scholar] [CrossRef] [Green Version]
- Magklara, A.; Yen, A.; Colquitt, B.M.; Clowney, E.J.; Allen, W.; Markenscoff-papadimitriou, E.; Evans, Z.A.; Kheradpour, P.; Mountoufaris, G.; Carey, C.; et al. An epigenetic signature for monoallelic olfactory receptor expression. Cell 2012, 145, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Chess, A.; Simon, I.; Cedar, H.; Axel, R. Allelic inactivation regulates olfactory receptor gene expression. Cell 1994, 78, 823–834. [Google Scholar] [CrossRef]
- Croy, I.; Bendas, J.; Wittrodt, N.; Lenk, M.; Joraschky, P.; Weidner, K. Gender-specific relation between olfactory sensitivity and disgust perception. Chem. Senses 2017, 42, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Ueda, K. Comparative morphology of fish olfactory epithelium—I*1salmoniformes. Nippon Suisan Gakkaishi. 1977, 43, 1163–1174. [Google Scholar] [CrossRef]
- Mokhtar, D.; Abd-Elhafeez, H. Light- and electron-microscopic studies of olfactory organ of Red-tail shark, Epalzeorhynchos bicolor (Teleostei: Cyprinidae). J. Microsc. Ultrastruct. 2014, 2, 182. [Google Scholar] [CrossRef] [Green Version]
- Doty, R.L.; Shaman, P.; Dann, M. Development of the university of pennsylvania smell identification test: A standardized microencapsulated test of olfactory function. Physiol. Behav. 1984, 32, 489–502. [Google Scholar] [CrossRef]
- Yabuki, Y.; Koide, T.; Miyasaka, N.; Wakisaka, N.; Masuda, M.; Ohkura, M.; Nakai, J.; Tsuge, K.; Tsuchiya, S.; Sugimoto, Y.; et al. Olfactory receptor for prostaglandin F2α mediates male fish courtship behavior. Nat. Neurosci. 2016, 19, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Villamayor, P.R.; Robledo, D.; Fernández, C.; Gullón, J.; Quintela, L.; Sánchez-Quinteiro, P.; Martínez, P. Analysis of the vomeronasal organ transcriptome reveals variable gene expression depending on age and function in rabbits. Genomics 2021, 113, 2240–2252. [Google Scholar] [CrossRef] [PubMed]
- Lewcock, J.W.; Reed, R.R. A feedback mechanism regulates monoallelic odorant receptor expression. Proc. Natl. Acad. Sci. USA 2004, 117, 801–815. [Google Scholar] [CrossRef] [Green Version]
- Shykind, B.M.; Rohani, S.C.; O’Donnell, S.; Nemes, A.; Mendelsohn, M.; Sun, Y.; Axel, R.; Barnea, G. Gene switching and the stability of odorant receptor gene choice. Cell 2004, 117, 801–815. [Google Scholar] [CrossRef] [Green Version]
- Serizawa, S.; Miyamichi, K.; Nakatani, H.; Suzuki, M.; Saito, M.; Yoshihara, Y.; Sakano, H. Negative feedback regulation ensures the one receptor-one olfactory neuron rule in mouse. Science 2003, 302, 2088–2094. [Google Scholar] [CrossRef] [Green Version]
- Torcia, M.G.; Nencioni, L.; Clemente, A.M.; Civitelli, L.; Celestino, I.; Limongi, D.; Fadigati, G.; Perissi, E.; Cozzolino, F.; Garaci, E.; et al. Sex differences in the response to viral infections: TLR8 and TLR9 ligand stimulation induce higher IL10 production in males. PLoS ONE 2012, 7, e39853. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L.; Jedlicka, A.; Pekosz, A. The Xs and Y of immune responses to viral vaccines. Lancet Infect. 2010, 10, 338–349. [Google Scholar] [CrossRef]
- Hill-Burns, E.M.; Clark, A.G. X-linked variation in immune response in Drosophila melanogaster. Genetics 2009, 183, 1477–1491. [Google Scholar] [CrossRef] [Green Version]
- Pap, P.L.; Czirják, G.Á.; Vágási, C.I.; Barta, Z.; Hasselquist, D. Sexual dimorphism in immune function changes during the annual cycle in house sparrows. Naturwissenschaften 2010, 97, 891–901. [Google Scholar] [CrossRef]
- Fargallo, J.A.; Martínez-Padilla, J.; Toledano-Díaz, A.; Santiago-Moreno, J.; Dávila, J.A. Sex and testosterone effects on growth, immunity and melanin coloration of nestling Eurasian kestrels. J. Anim. Ecol. 2007, 76, 201–209. [Google Scholar] [CrossRef]
- Taylor, K.; Kimbrell, D.A. Host immune response and differential survival of the sexes in Drosophila. Fly (Austin) 2007, 1, 197–204. [Google Scholar] [CrossRef]
- Mondal, S.; Rai, U. Sexual dimorphism in phagocytic activity of wall lizard’s splenic macrophages and its control by sex steroids. Gen. Comp. Endocrinol. 1999, 116, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Lotter, H.; Altfeld, M. Sex differences in immunity. Semin. Immunopathol. 2019, 41, 133–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radkiewicz, C.; Johansson, A.L.V.; Dickman, P.W.; Lambe, M.; Edgren, G. Sex differences in cancer risk and survival: A swedish cohort study. Eur. J. Cancer 2017, 84, 130–140. [Google Scholar] [CrossRef]
- Scelo, G.; Li, P.; Chanudet, E.; Muller, D.C. Variability of sex disparities in cancer incidence over 30 years: The striking case of kidney cancer. Eur. Urol. Focus 2018, 4, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Grossman, C. Possible underlying mechanisms of sexual dimorphism in the immune response, fact and hypothesis. J. Steroid Biochem. 1989, 34, 241–251. [Google Scholar] [CrossRef]
- Butterworth, M.; McClellan, B.; Aklansmith, M. Influence of sex on immunoglobulin levels. Nature 1967, 214, 1224–1225. [Google Scholar] [CrossRef]
- Terres, G.; Morrison, S.L.; Habicht, G.S. A quantitative difference in the immune response between male and female mice. Proc. Soc. Exp. Biol. Med. 1968, 127, 664–667. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Hannah, M.F.; Bajic, V.B.; Klein, S.L. Sex differences in the recognition of and innate antiviral responses to Seoul virus in Norway rats. Brain Behav. Immun. 2008, 22, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg1, S.L. HISAT: A fast spliced aligner with low memory requirements Daehwan HHS Public Access. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Parrish, N.; Hormozdiari, F.; Eskin, E. Assembly of non-unique insertion content using next-generation sequencing. Bioinform. Impact Accurate Quantif. Proteom. Genet. Anal. Res. 2014, 12, 21–40. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.M.; Liu, T.; Liu, C.J.; Song, S.; Zhang, X.; Liu, W.; Jia, H.; Xue, Y.; Guo, A.Y. AnimalTFDB 2.0: A resource for expression, prediction and functional study of animal transcription factors. Nucleic Acids Res. 2015, 43, D76–D81. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- IBM Corp. IBM SPSS Statistics for Windows, Version 22.0; IBM Corp.: Armonk, NY, USA, 2013. [Google Scholar]
- Alamri, A.; Nam, J.Y.; Blancato, J.K. Fluorescence in situ hybridization of cells, chromosomes, and formalin-fixed paraffin-embedded tissues. Methods Mol. Biol. 2017, 1606, 265–279. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, M.; Chen, X.; Huang, X.; Liu, N.; Wang, W.; Liu, H. Transcriptome Analysis Reveals Sexual Disparities between Olfactory and Immune Gene Expression in the Olfactory Epithelium of Megalobrama amblycephala. Int. J. Mol. Sci. 2021, 22, 13017. https://doi.org/10.3390/ijms222313017
Lv M, Chen X, Huang X, Liu N, Wang W, Liu H. Transcriptome Analysis Reveals Sexual Disparities between Olfactory and Immune Gene Expression in the Olfactory Epithelium of Megalobrama amblycephala. International Journal of Molecular Sciences. 2021; 22(23):13017. https://doi.org/10.3390/ijms222313017
Chicago/Turabian StyleLv, Maolin, Xiuli Chen, Xin Huang, Ning Liu, Weimin Wang, and Han Liu. 2021. "Transcriptome Analysis Reveals Sexual Disparities between Olfactory and Immune Gene Expression in the Olfactory Epithelium of Megalobrama amblycephala" International Journal of Molecular Sciences 22, no. 23: 13017. https://doi.org/10.3390/ijms222313017