
Role of ANGPTL8 in NAFLD Improvement after Bariatric Surgery in Experimental and Human Obesity

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Prevalence of NAFLD and Metabolic Improvement after Bariatric Surgery in the Studied Population
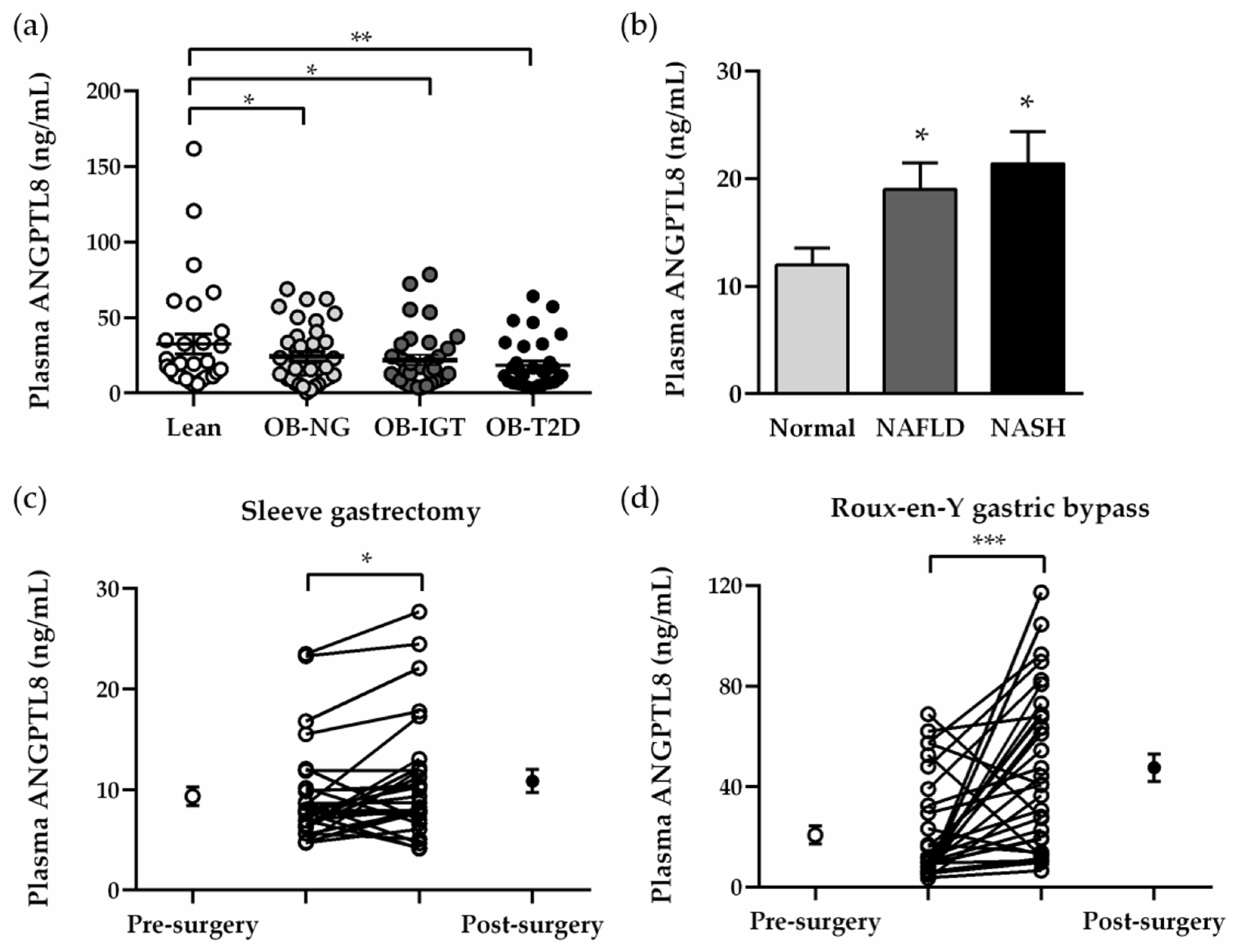
2.2. Plasma Concentrations of ANGPTL8 Are Increased in Obesity-Associated NAFLD
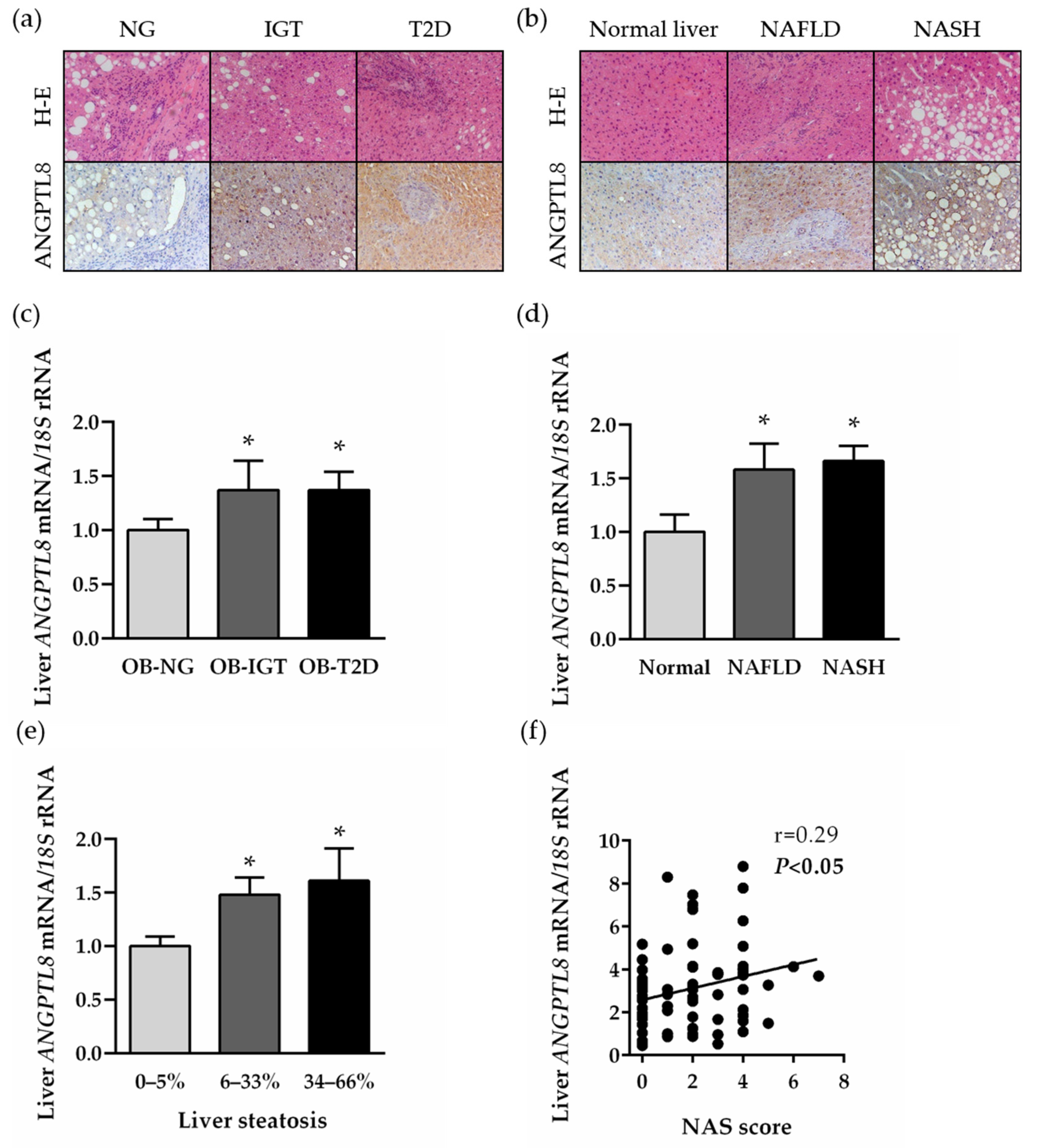
2.3. Hepatic Expression of ANGTL8 Is Upregulated in Patients with Morbid Obesity and NAFLD in Relation to Their Degree of Liver Steatosis
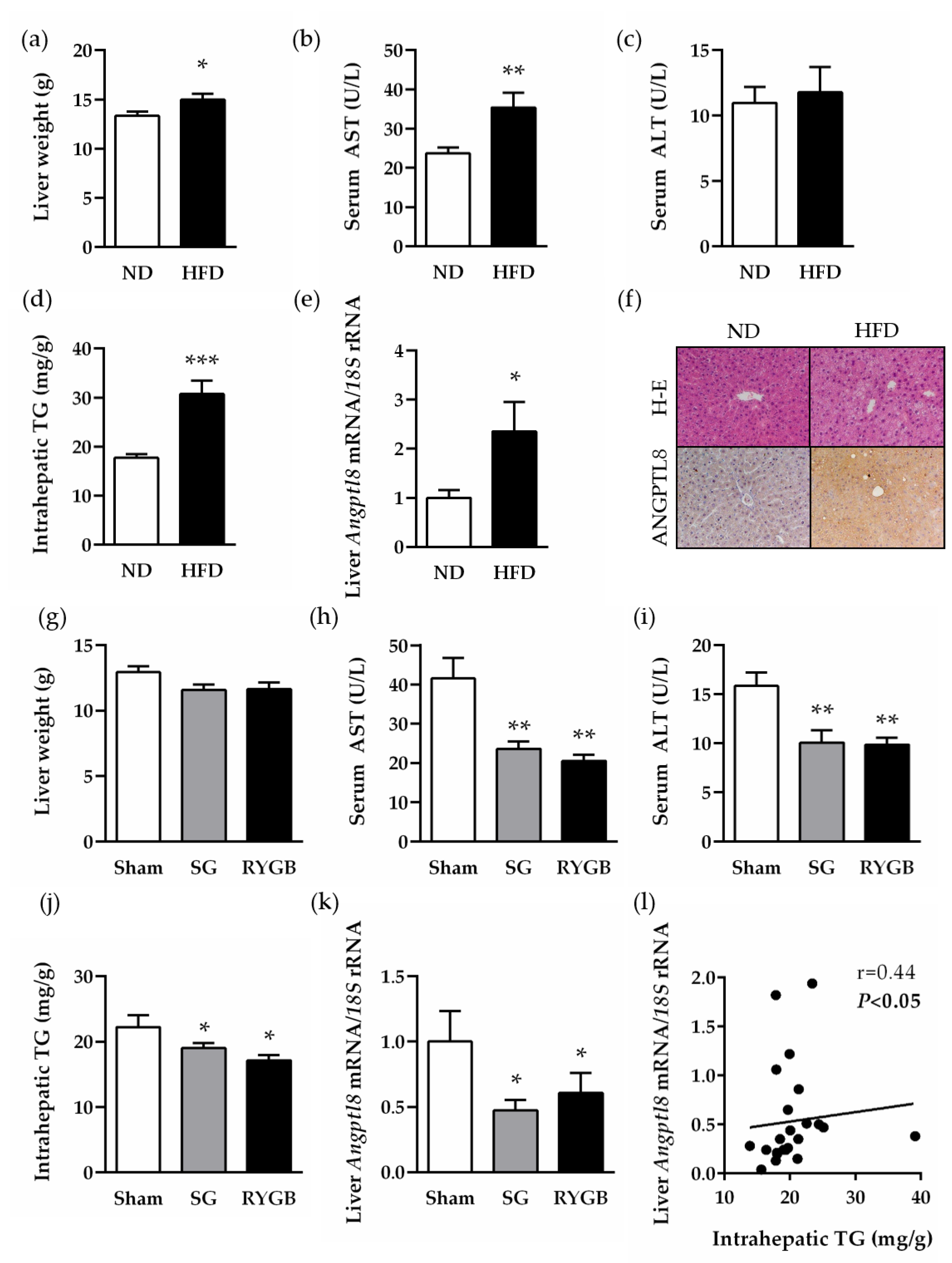
2.4. Increased Hepatic Expression of ANGPTL8 in a Preclinical Model of Diet-Induced Obesity Is Downregulated after Bariatric Surgery
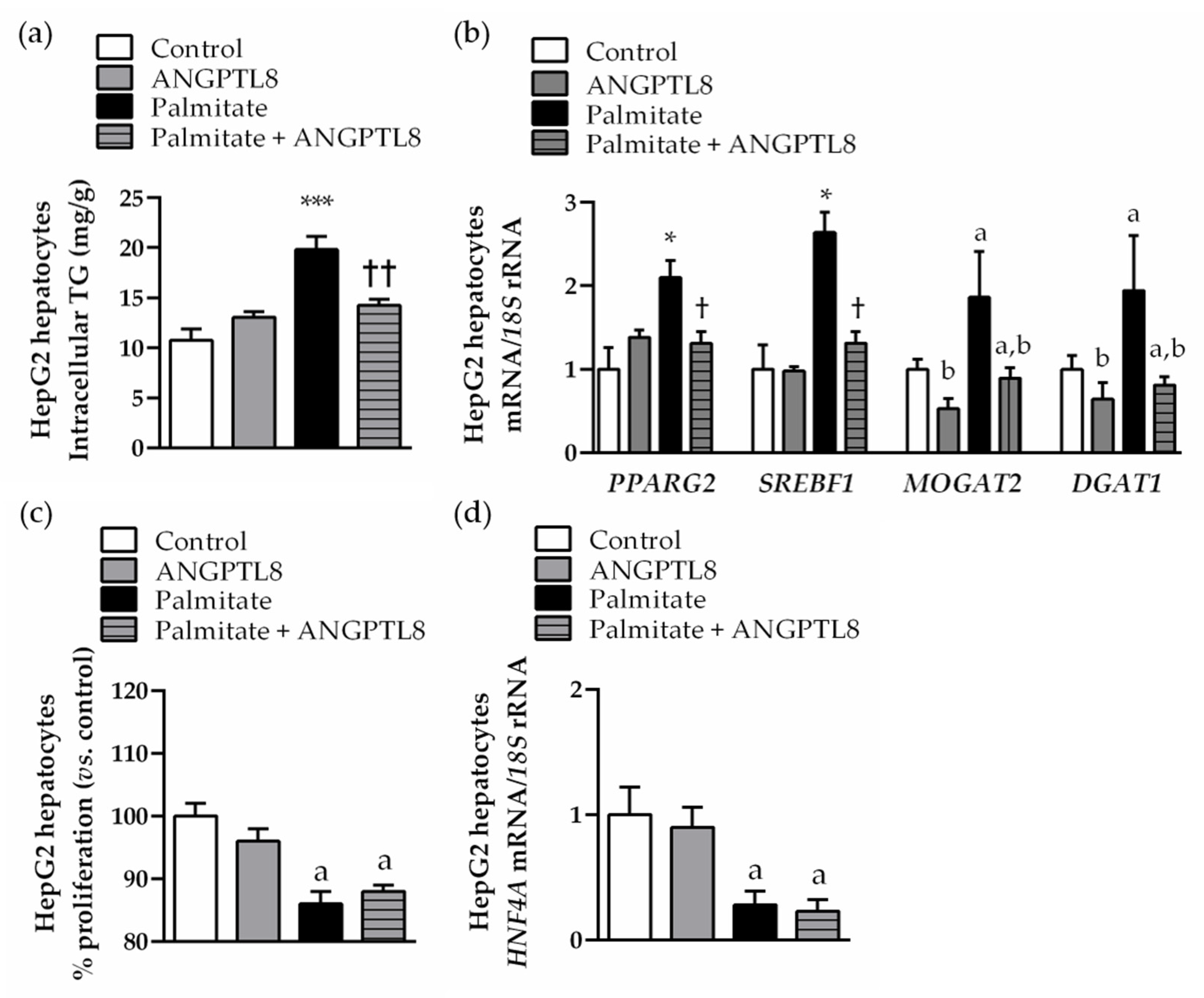
2.5. ANGPTL8 Inhibits Palmitate-Induced Lipogenesis, but Not Its Anti-Proliferative Action, in Human HepG2 Hepatocytes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patients
5.2. Experimental Animals
5.3. Blood Analysis
5.4. Real-Time PCR
5.5. Immunohistochemistry of ANGPTL8
5.6. Cell Culture and Treatment
5.7. Proliferation Assay
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powell, E.E.; Wong, V.W.; Rinella, M. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Marques-Vidal, P.; Cortez-Pinto, H. Hepatic histology in obese patients undergoing bariatric surgery. J. Hepatol. 2006, 45, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Golabi, P.; de Avila, L.; Paik, J.M.; Srishord, M.; Fukui, N.; Qiu, Y.; Burns, L.; Afendy, A.; Nader, F. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: A systematic review and meta-analysis. J. Hepatol. 2019, 71, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Cummings, D.E.; Rubino, F. Metabolic surgery for the treatment of type 2 diabetes in obese individuals. Diabetologia 2018, 61, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mingrone, G.; Panunzi, S.; De Gaetano, A.; Guidone, C.; Iaconelli, A.; Capristo, E.; Chamseddine, G.; Bornstein, S.R.; Rubino, F. Metabolic surgery versus conventional medical therapy in patients with type 2 diabetes: 10-year follow-up of an open-label, single-centre, randomised controlled trial. Lancet 2021, 397, 293–304. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar] [CrossRef]
- Lassailly, G.; Caiazzo, R.; Buob, D.; Pigeyre, M.; Verkindt, H.; Labreuche, J.; Raverdy, V.; Leteurtre, E.; Dharancy, S.; Louvet, A.; et al. Bariatric surgery reduces features of nonalcoholic steatohepatitis in morbidly obese patients. Gastroenterology 2015, 149, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, D.; Chennakesavalu, M.; Gangemi, A. Systematic review and meta-analysis of Roux-en-Y gastric bypass against laparoscopic sleeve gastrectomy for amelioration of NAFLD using four criteria. Surg. Obes. Relat. Dis. 2019, 15, 2123–2130. [Google Scholar] [CrossRef]
- Zhang, R. Lipasin, a novel nutritionally-regulated liver-enriched factor that regulates serum triglyceride levels. Biochem. Biophys. Res. Commun. 2012, 424, 786–792. [Google Scholar] [CrossRef]
- Wang, Y.; Quagliarini, F.; Gusarova, V.; Gromada, J.; Valenzuela, D.M.; Cohen, J.C.; Hobbs, H.H. Mice lacking ANGPTL8 (Betatrophin) manifest disrupted triglyceride metabolism without impaired glucose homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, 16109–16114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Shannon, C.E.; Bakewell, T.M.; Abdul-Ghani, M.A.; Fourcaudot, M.; Norton, L. Regulation of ANGPTL8 in liver and adipose tissue by nutritional and hormonal signals and its effect on glucose homeostasis in mice. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E613–E624. [Google Scholar] [CrossRef] [PubMed]
- Sylvers-Davie, K.L.; Davies, B.S.J. Regulation of lipoprotein metabolism by ANGPTL3, ANGPTL4, and ANGPTL8. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E493–E508. [Google Scholar] [CrossRef]
- Quagliarini, F.; Wang, Y.; Kozlitina, J.; Grishin, N.V.; Hyde, R.; Boerwinkle, E.; Valenzuela, D.M.; Murphy, A.J.; Cohen, J.C.; Hobbs, H.H. Atypical angiopoietin-like protein that regulates ANGPTL3. Proc. Natl. Acad. Sci. USA 2012, 109, 19751–19756. [Google Scholar] [CrossRef] [Green Version]
- Alenad, A.; Alenezi, M.M.; Alokail, M.S.; Wani, K.; Mohammed, A.K.; Alnaami, A.M.; Sulimani, M.; Zargar, S.; Clerici, M.; Al-Daghri, N.M. Association of ANGPTL8 (betatrophin) gene variants with components of metabolic syndrome in Arab adults. Sci. Rep. 2020, 10, 6764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peloso, G.M.; Auer, P.L.; Bis, J.C.; Voorman, A.; Morrison, A.C.; Stitziel, N.O.; Brody, J.A.; Khetarpal, S.A.; Crosby, J.R.; Fornage, M.; et al. Association of low-frequency and rare coding-sequence variants with blood lipids and coronary heart disease in 56,000 whites and blacks. Am. J. Hum. Genet. 2014, 94, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Barja-Fernández, S.; Folgueira, C.; Seoane, L.M.; Casanueva, F.F.; Dieguez, C.; Castelao, C.; Agüera, Z.; Banos, R.; Botella, C.; de la Torre, R.; et al. Circulating betatrophin levels are increased in anorexia and decreased in morbidly obese women. J. Clin. Endocrinol. Metab. 2015, 100, E1188–E1196. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ambrosi, J.; Pascual, E.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Silva, C.; Gil, M.J.; Salvador, J.; Frühbeck, G. Circulating betatrophin concentrations are decreased in human obesity and type 2 diabetes. J. Clin. Endocrinol. Metab. 2014, 99, E2004–E2009. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ambrosi, J.; Pascual-Corrales, E.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Romero, S.; Vila, N.; Ibañez, P.; Margall, M.A.; Silva, C.; et al. Altered concentrations in dyslipidemia evidence a role for ANGPTL8/betatrophin in lipid metabolism in humans. J. Clin. Endocrinol. Metab. 2016, 101, 3803–3811. [Google Scholar] [CrossRef]
- Pascual-Corrales, E.; Gómez-Ambrosi, J.; Moncada, R.; Valentí, V.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Silva, C.; Gil, M.J.; Salvador, J.; et al. Circulating ANGPTL8/betatrophin concentrations are increased after surgically induced weight loss, but not after diet-induced weight loss. Obes. Surg. 2016, 26, 1881–1889. [Google Scholar] [CrossRef]
- Guo, C.; Wang, C.; Deng, X.; He, J.; Yang, L.; Yuan, G. ANGPTL8 in metabolic homeostasis: More friend than foe? Open Biol. 2021, 11, 210106. [Google Scholar] [CrossRef] [PubMed]
- García-Monzón, C.; Petrov, P.D.; Rey, E.; Marañón, P.; Del Pozo-Maroto, E.; Guzmán, C.; Rodríguez de Cía, J.; Casado-Collado, A.J.; Vargas-Castrillón, J.; Saez, A.; et al. Angiopoietin-like protein 8 is a novel vitamin D receptor target gene involved in nonalcoholic fatty liver pathogenesis. Am. J. Pathol. 2018, 188, 2800–2810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.H.; Lee, S.G.; Lee, C.J.; Kim, S.H.; Song, Y.M.; Yoon, M.R.; Jeon, B.H.; Lee, J.H.; Lee, B.W.; Kang, E.S.; et al. Association between betatrophin/ANGPTL8 and non-alcoholic fatty liver disease: Animal and human studies. Sci. Rep. 2016, 6, 24013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, Y.; Liu, S.; Zhang, Z.; Hu, J. Circulating angiopoietin-like proteins in metabolic-associated fatty liver disease: A systematic review and meta-analysis. Lipids Health Dis. 2021, 20, 55. [Google Scholar] [CrossRef] [PubMed]
- von Loeffelholz, C.; Pfeiffer, A.F.H.; Lock, J.F.; Lieske, S.; Docke, S.; Murahovschi, V.; Kriebel, J.; de Las Heras Gala, T.; Grallert, H.; Rudovich, N.; et al. ANGPTL8 (Betatrophin) is Expressed in Visceral Adipose Tissue and Relates to Human Hepatic Steatosis in Two Independent Clinical Collectives. Horm. Metab. Res. 2017, 49, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, F.; Wu, R.; Wang, P.; Wu, Y.; Azam, M.S.; Xu, Q.; Chen, Y.; Liu, Y. Fasting and feeding signals control the oscillatory expression of Angptl8 to modulate lipid metabolism. Sci. Rep. 2016, 6, 36926. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Zhao, Z.; Deng, X.; Chen, Z.; Tu, Z.; Yuan, G. Regulation of angiopoietin-like protein 8 expression under different nutritional and metabolic status. Endocr. J. 2019, 66, 1039–1046. [Google Scholar] [CrossRef]
- Ren, G.; Kim, J.Y.; Smas, C.M. Identification of RIFL, a novel adipocyte-enriched insulin target gene with a role in lipid metabolism. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E334–E351. [Google Scholar] [CrossRef] [Green Version]
- Müller, F.A.; Sturla, S.J. Human in vitro models of nonalcoholic fatty liver disease. Curr. Opin. Toxicol. 2019, 16, 9–16. [Google Scholar] [CrossRef]
- Hu, W.; Shao, X.; Guo, D.; Hao, H.; Zhang, Y.; Xia, M.; Gong, Y.; Zhou, H.; Fan, Y.; Yu, W. Relationship of Serum Betatrophin with Nonalcoholic Fatty Liver in a Chinese Population. PLoS ONE 2017, 12, e0170758. [Google Scholar] [CrossRef]
- Hong, B.S.; Liu, J.; Zheng, J.; Ke, W.; Huang, Z.; Wan, X.; He, X.; Xiao, H.; Li, Y. Angiopoietin-like protein 8/betatrophin correlates with hepatocellular lipid content independent of insulin resistance in non-alcoholic fatty liver disease patients. J. Diabetes Investig. 2018, 9, 952–958. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, M.; Ozenirler, S.; Kocabiyik, M. Serum beta-trophin level as a new marker for noninvasive assessment of nonalcoholic fatty liver disease and liver fibrosis. Eur. J. Gastroenterol. Hepatol. 2016, 28, 57–63. [Google Scholar] [CrossRef]
- Fenzl, A.; Itariu, B.K.; Kosi, L.; Fritzer-Szekeres, M.; Kautzky-Willer, A.; Stulnig, T.M.; Kiefer, F.W. Circulating betatrophin correlates with atherogenic lipid profiles but not with glucose and insulin levels in insulin-resistant individuals. Diabetologia 2014, 57, 1204–1208. [Google Scholar] [CrossRef]
- de Brito, E.S.M.B.; Tustumi, F.; de Miranda Neto, A.A.; Dantas, A.C.B.; Santo, M.A.; Cecconello, I. Gastric bypass compared with sleeve gastrectomy for nonalcoholic fatty liver disease: A systematic review and meta-analysis. Obes. Surg. 2021, 31, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Cherla, D.V.; Rodriguez, N.A.; Vangoitsenhoven, R.; Singh, T.; Mehta, N.; McCullough, A.J.; Brethauer, S.A.; Schauer, P.R.; Aminian, A. Impact of sleeve gastrectomy and Roux-en-Y gastric bypass on biopsy-proven non-alcoholic fatty liver disease. Surg. Endosc. 2020, 34, 2266–2272. [Google Scholar] [CrossRef] [PubMed]
- Faramia, J.; Ostinelli, G.; Drolet-Labelle, V.; Picard, F.; Tchernof, A. Metabolic adaptations after bariatric surgery: Adipokines, myokines and hepatokines. Curr. Opin. Pharmacol. 2020, 52, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ambrosi, J.; Gallego-Escuredo, J.M.; Catalán, V.; Rodríguez, A.; Domingo, P.; Moncada, R.; Valentí, V.; Salvador, J.; Giralt, M.; Villarroya, F.; et al. FGF19 and FGF21 serum concentrations in human obesity and type 2 diabetes behave differently after diet- or surgically-induced weight loss. Clin. Nutr. 2017, 36, 861–868. [Google Scholar] [CrossRef]
- Poloczek, J.; Kazura, W.; Kwasnicka, E.; Gumprecht, J.; Jochem, J.; Stygar, D. Effects of bariatric surgeries on fetuin-A, selenoprotein P, angiopoietin-like protein 6, and fibroblast growth factor 21 concentration. J. Diabetes Res. 2021, 2021, 5527107. [Google Scholar] [CrossRef] [PubMed]
- Al-Shawaf, E.; Al-Ozairi, E.; Al-Asfar, F.; Al-Beloushi, S.; Kumari, S.; Tuomilehto, J.; Arefanian, H. Biphasic changes in angiopoietin-like 8 level after laparoscopic sleeve gastrectomy and type 2 diabetes remission during a 1-year follow-up. Surg. Obes. Relat. Dis. 2018, 14, 1284–1294. [Google Scholar] [CrossRef]
- Guo, K.; Yu, H.; Lu, J.; Bao, Y.; Chen, H.; Jia, W. Decreased serum betatrophin levels correlate with improved fasting plasma glucose and insulin secretion capacity after Roux-en-Y gastric bypass in obese Chinese patients with type 2 diabetes: A 1-year follow-up. Surg. Obes. Relat. Dis. 2016, 12, 1343–1348. [Google Scholar] [CrossRef]
- Song, Z.; Guo, K.; Huang, W.; Xu, H.; Liu, Y.; Guan, J.; Yin, S.; Yu, H.; Yi, H.; Zou, J. Decreased serum betatrophin may correlate with the improvement of obstructive sleep apnea after Roux-en-Y Gastric Bypass surgery. Sci. Rep. 2021, 11, 1808. [Google Scholar] [CrossRef] [PubMed]
- Ejarque, M.; Borlaug, M.; Vilarrasa, N.; Martinez-Perez, B.; Llauradó, G.; Megía, A.; Helland, T.; Gutierrez, C.; Serena, C.; Folkestad, O.; et al. Angiopoietin-like protein 8/betatrophin as a new determinant of type 2 diabetes remission after bariatric surgery. Transl. Res. 2017, 184, 35–44.e4. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2021. Diabetes Care 2021, 44, S15–S33. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Ezquerro, S.; Becerril, S.; Tuero, C.; Méndez-Giménez, L.; Mocha, F.; Moncada, R.; Valentí, V.; Cienfuegos, J.A.; Catalán, V.; Gómez-Ambrosi, J.; et al. Role of ghrelin isoforms in the mitigation of hepatic inflammation, mitochondrial dysfunction, and endoplasmic reticulum stress after bariatric surgery in rats. Int. J. Obes. 2020, 44, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Ezquerro, S.; Mocha, F.; Frühbeck, G.; Guzmán-Ruiz, R.; Valentí, V.; Mugueta, C.; Becerril, S.; Catalán, V.; Gómez-Ambrosi, J.; Silva, C.; et al. Ghrelin reduces TNF-alpha-induced human hepatocyte apoptosis, autophagy, and pyroptosis: Role in obesity-associated NAFLD. J. Clin. Endocr. Metab. 2019, 104, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Unamuno, X.; Gómez-Ambrosi, J.; Ramírez, B.; Rodríguez, A.; Becerril, S.; Valentí, V.; Moncada, R.; Silva, C.; Salvador, J.; Frühbeck, G.; et al. NLRP3 inflammasome blockade reduces adipose tissue inflammation and extracellular matrix remodeling. Cell. Mol. Immunol. 2021, 18, 1045–1057. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lean | Obese NG | Obese IGT | Obese T2D | p | |
|---|---|---|---|---|---|
| n | 30 | 48 | 47 | 45 | - |
| Sex (male/female) | 16/14 | 17/31 | 17/30 | 20/25 | 0.199 |
| Age (years) | 43 ± 3 | 40 ± 2 | 44 ± 2 | 48 ± 2 b | 0.031 |
| BMI (kg/m2) | 22.6 ± 0.5 | 44.5 ± 1.3 a | 43.2 ± 0.9 a | 45.4 ± 1.4 a | <0.0001 |
| Body fat (%) | 21.2 ± 1.4 | 50.7 ± 0.9 a | 51.0 ± 1.3 a | 50.1 ± 1.2 a | <0.0001 |
| Waist circumference (cm) | 77 ± 3 | 125 ± 2 a | 125 ± 2 a | 132 ± 3 a | <0.0001 |
| Glucose (mg/dL) | 87 ± 2 | 92 ± 1 | 105 ± 1 a,b | 142 ± 9 a,b | <0.0001 |
| Glucose 2-h OGTT (mg/dL) | - | 121 ± 4 | 154 ± 5 b | 250 ± 16 b | <0.0001 |
| Insulin (µU/mL) | 5.7 ± 0.6 | 20.9 ± 2.7 a | 19.6 ± 1.7 a | 26.3 ± 3.1 a | <0.0001 |
| Insulin 2-h OGTT (µU/mL) | - | 97.0 ± 9.5 | 141.1 ± 12.4 b | 133.3 ± 19.2 b | 0.034 |
| HOMA | 1.2 ± 0.1 | 4.3 ± 0.4 | 5.2 ± 0.5 a | 9.2 ± 1.7 a,b | <0.0001 |
| QUICKI | 0.38 ± 0.01 | 0.32 ± 0.01 a | 0.31 ± 0.01 a | 0.30 ± 0.01 a,b | <0.0001 |
| FFA (mmol/L) | 15.7 ± 0.1 | 16.7 ± 1.1 | 22.5 ± 2.2 | 28.9 ± 4.0 a,b | 0.001 |
| Glycerol (mg/dL) | 18.2 ± 3.5 | 19.8 ± 2.1 | 22.1 ± 2.5 | 21.9 ± 2.3 | 0.591 |
| Adipo-IR index | 20.2 ± 3.4 | 71.3 ± 7.7 | 98.9 ± 13.7 | 188.4 ± 36.3 a,b | <0.0001 |
| Triacylglycerol (mg/dL) | 70 ± 5 | 111 ± 7 a | 132 ± 11 a | 147 ± 11 a,b | <0.0001 |
| Total cholesterol (mg/dL) | 186 ± 7 | 191 ± 4 | 203 ± 6 | 198 ± 6 | 0.234 |
| LDL cholesterol (mg/dL) | 110 ± 6 | 118 ± 3 | 127 ± 5 | 116 ± 5 | 0.145 |
| HDL cholesterol (mg/dL) | 61 ± 3 | 50 ± 4 | 49 ± 2 a | 44 ± 2 a | 0.001 |
| CRP (mg/L) | 1.8 ± 0.3 | 8.3 ± 1.1 | 9.5 ± 1.5 a | 8.8 ± 1.8 a | 0.035 |
| Uric acid (mg/dL) | 4.3 ± 0.2 | 5.7 ± 0.2 a | 5.9 ± 0.2 a | 6.0 ± 0.2 a | 0.001 |
| Leptin (ng/mL) | 6.1 ± 0.7 | 45.4 ± 3.2 a | 48.9 ± 3.6 a | 46.0 ± 5.8 a | <0.0001 |
| AST (IU/L) | 13 ± 1 | 16 ± 1 | 17 ± 1 | 17 ± 1 | 0.244 |
| ALT (IU/L) | 10 ± 2 | 21 ± 2 | 28 ± 2 a | 27 ± 2 a | <0.0001 |
| Alkaline phosphatase (IU/L) | 86 ± 5 | 68 ± 5 | 77 ± 6 | 73 ± 5 | 0.334 |
| γ-GT (IU/L) | 11 ± 1 | 21 ± 2 | 29 ± 4 a | 30 ± 4 a | 0.015 |
| Daily alcohol intake (g) | 0.0 ± 0.0 | 1.1 ± 0.9 | 3.6 ± 2.7 | 4.6 ± 2.7 | 0.576 |
| Antihypertensive therapy, n (%) | 0 (0%) | 11 (23%) | 17 (32%) | 19 (49%) | 0.003 |
| Antidiabetic therapy, n (%) | 0 (0%) | 2 (4%) | 1 (2%) | 20 (51%) | <0.0001 |
| Lipid-lowering therapy, n (%) | 0 (0%) | 6 (13%) | 7 (15%) | 11 (28%) | 0.061 |
| Sleeve Gastrectomy (n = 34) | Roux-en-Y Gastric Bypass (n = 41) | |||||
|---|---|---|---|---|---|---|
| Pre-Surgery | Post-Surgery | p | Pre-Surgery | Post-Surgery | p | |
| Sex (male/female) | 10/24 | 10/24 | - | 15/26 | 15/26 | - |
| BMI (kg/m2) | 41.0 ± 1.7 | 31.1 ± 1.6 | <0.0001 | 44.7 ± 0.8 | 31.9 ± 0.6 | <0.0001 |
| Body fat (%) | 50.9 ± 1.2 | 39.5 ± 1.8 | <0.0001 | 50.7 ± 0.8 | 38.1 ± 1.0 | <0.0001 |
| Waist circumference (cm) | 120 ± 3 | 98 ± 3 | <0.0001 | 127 ± 2 | 100 ± 2 | <0.0001 |
| Glucose (mg/dL) | 106 ± 9 | 92 ± 4 | 0.022 | 115 ± 5 | 95 ± 4 | <0.0001 |
| Insulin (µU/mL) | 30.9 ± 6.6 | 10.0 ± 1.6 | 0.005 | 23.0 ± 3.3 | 8.6 ± 1.1 | <0.0001 |
| HOMA | 8.3 ± 2.1 | 2.1 ± 0.4 | 0.011 | 6.2 ± 1.3 | 2.3 ± 0.6 | <0.0001 |
| QUICKI | 0.30 ± 0.01 | 0.36 ± 0.01 | <0.0001 | 0.31 ± 0.01 | 0.36 ± 0.01 | <0.0001 |
| FFA (mmol/L) | 18.8 ± 1.2 | 14.3 ± 1.5 | 0.052 | 21.5 ± 2.2 | 13.2 ± 1.0 | 0.001 |
| Glycerol (mg/dL) | 19.4 ± 2.2 | 14.0 ± 0.8 | 0.043 | 22.4 ± 2.1 | 12.9 ± 1.3 | 0.001 |
| Adipo-IR index | 138.7 ± 40.0 | 32.6 ± 6.3 | 0.020 | 91.9 ± 12.7 | 28.7 ± 5.6 | <0.0001 |
| Triacylglycerol (mg/dL) | 110 ± 15 | 98 ± 10 | 0.380 | 136 ± 9 | 90 ± 4 | <0.0001 |
| Total cholesterol (mg/dL) | 185 ± 8 | 186 ± 9 | 0.919 | 199 ± 5 | 158 ± 4 | <0.0001 |
| LDL cholesterol (mg/dL) | 115 ± 7 | 119 ± 7 | 0.576 | 122 ± 5 | 92 ± 3 | <0.0001 |
| HDL cholesterol (mg/dL) | 47 ± 3 | 52 ± 3 | 0.111 | 49 ± 3 | 48 ± 1 | 0.666 |
| CRP (mg/L) | 4.6 ± 1.5 | 3.0 ± 1.2 | 0.252 | 7.9 ± 1.1 | 2.4 ± 0.3 | <0.0001 |
| Uric acid (mg/dL) | 5.0 ± 0.3 | 4.7 ± 0.3 | 0.203 | 6.1 ± 0.2 | 4.9 ± 0.1 | <0.0001 |
| Leptin (ng/mL) | 77.0 ± 12.7 | 25.3 ± 5.9 | 0.004 | 51.0 ± 4.6 | 16.2 ± 2.5 | <0.0001 |
| AST (IU/L) | 17 ± 1 | 17 ± 1 | 0.983 | 18 ± 1 | 18 ± 1 | 0.849 |
| ALT (IU/L) | 22 ± 3 | 19 ± 2 | 0.318 | 27 ± 2 | 26 ± 2 | 0.606 |
| Alkaline phosphatase (IU/L) | 64 ± 6 | 65 ± 5 | 0.916 | 82 ± 5 | 84 ± 5 | 0.529 |
| γ-GT (IU/L) | 29 ± 6 | 22 ± 4 | 0.027 | 24 ± 3 | 14 ± 2 | <0.0001 |
| Antihypertensive therapy, n (%) | 15 (44%) | 12 (34%) | 0.001 | 17 (41%) | 9 (22%) | 0.011 |
| Antidiabetic therapy, n (%) | 8 (24%) | 5 (15%) | 0.085 | 12 (29%) | 4 (10%) | 0.001 |
| Lipid-lowering therapy, n (%) | 8 (24%) | 3 (9%) | 0.113 | 11 (27%) | 3 (7%) | 0.081 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perdomo, C.M.; Gómez-Ambrosi, J.; Becerril, S.; Valentí, V.; Moncada, R.; Fernández-Sáez, E.M.; Méndez-Giménez, L.; Ezquerro, S.; Catalán, V.; Silva, C.; et al. Role of ANGPTL8 in NAFLD Improvement after Bariatric Surgery in Experimental and Human Obesity. Int. J. Mol. Sci. 2021, 22, 12945. https://doi.org/10.3390/ijms222312945
Perdomo CM, Gómez-Ambrosi J, Becerril S, Valentí V, Moncada R, Fernández-Sáez EM, Méndez-Giménez L, Ezquerro S, Catalán V, Silva C, et al. Role of ANGPTL8 in NAFLD Improvement after Bariatric Surgery in Experimental and Human Obesity. International Journal of Molecular Sciences. 2021; 22(23):12945. https://doi.org/10.3390/ijms222312945
Chicago/Turabian StylePerdomo, Carolina M., Javier Gómez-Ambrosi, Sara Becerril, Víctor Valentí, Rafael Moncada, Eva M. Fernández-Sáez, Leire Méndez-Giménez, Silvia Ezquerro, Victoria Catalán, Camilo Silva, and et al. 2021. "Role of ANGPTL8 in NAFLD Improvement after Bariatric Surgery in Experimental and Human Obesity" International Journal of Molecular Sciences 22, no. 23: 12945. https://doi.org/10.3390/ijms222312945