
Variability in Behavioral Phenotypes after Forced Swimming-Induced Stress in Rats Is Associated with Expression of the Glucocorticoid Receptor, Nurr1, and IL-1β in the Hippocampus

 , ,
, ,
Abstract
:1. Introduction
2. Results
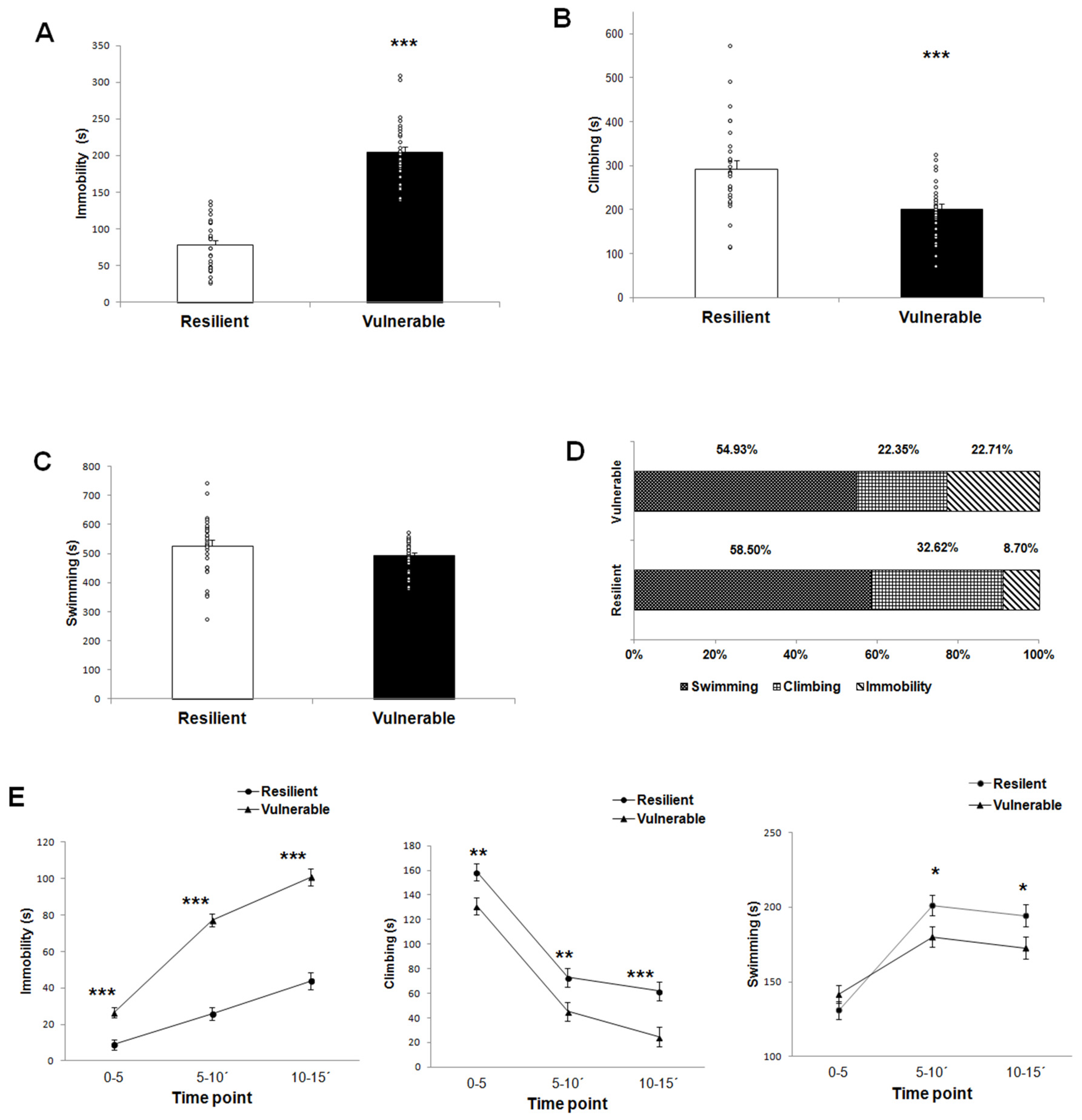
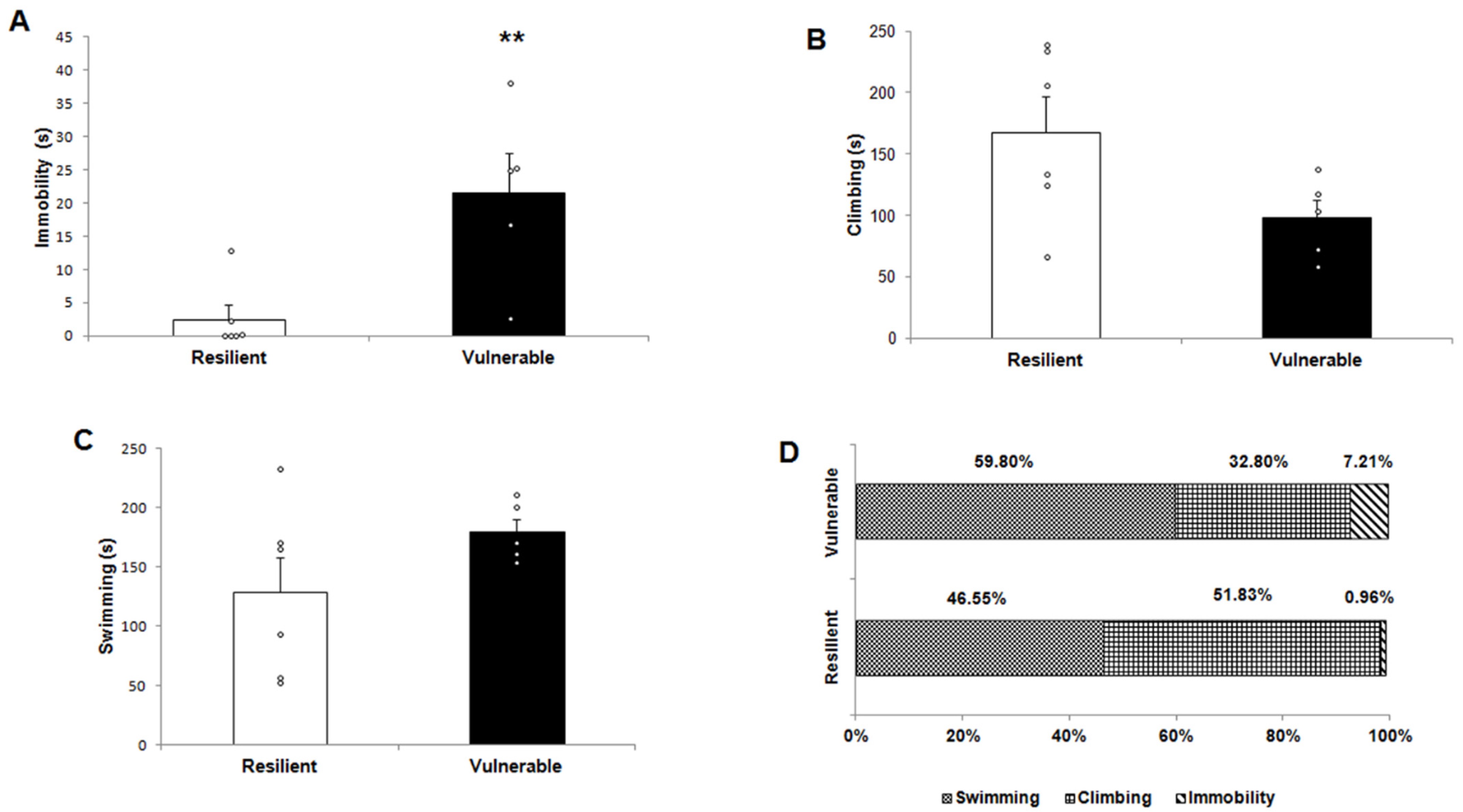
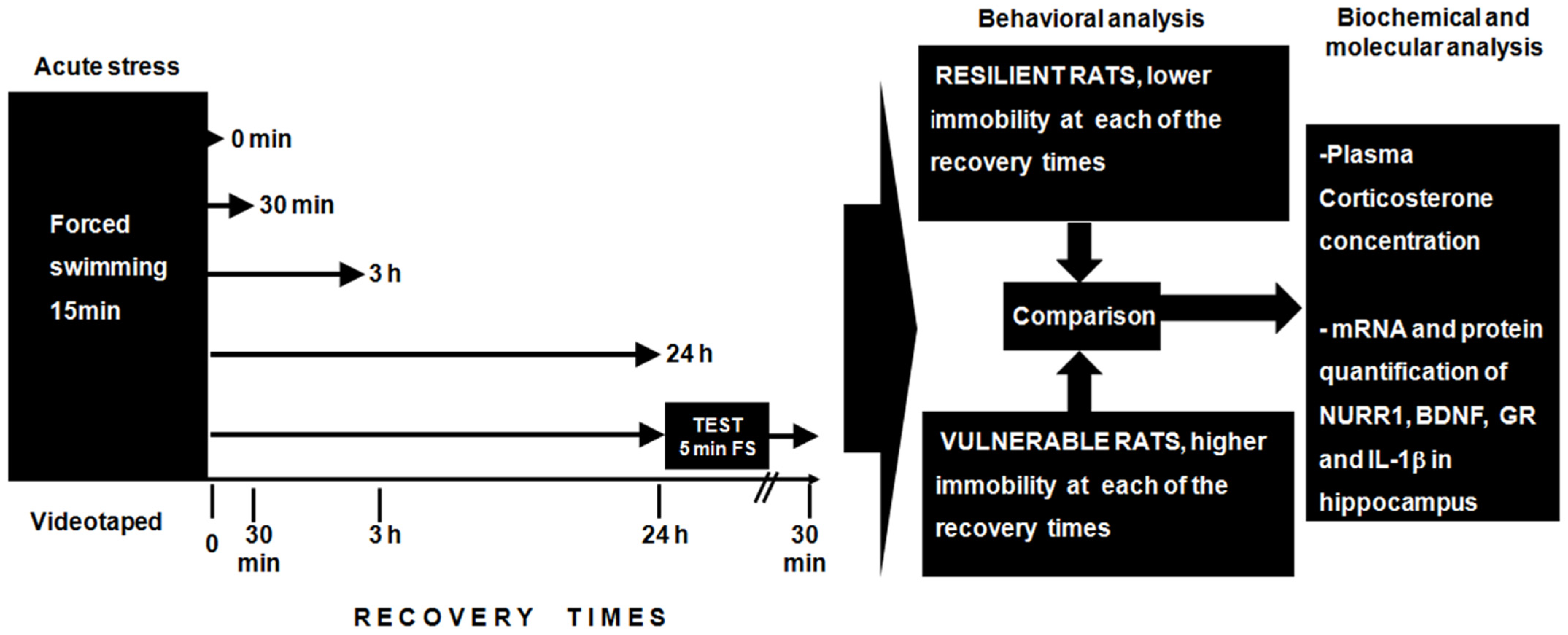
2.1. Behavioral Response
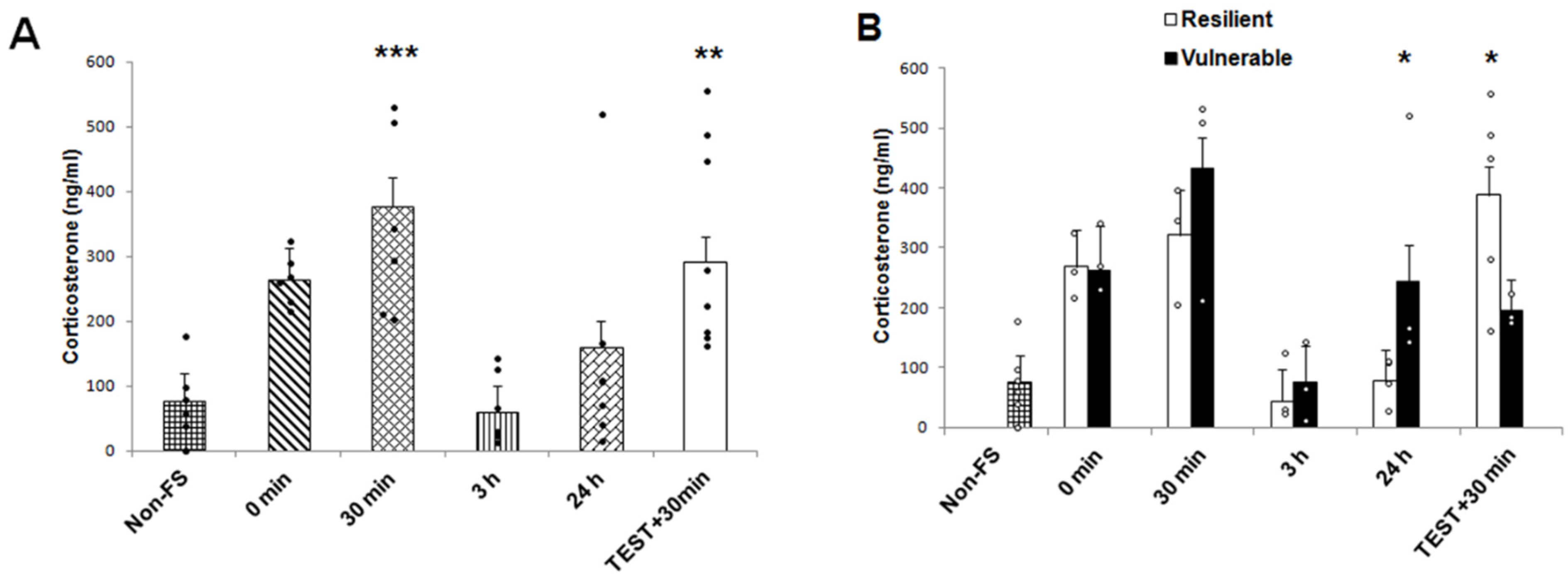
2.2. Plasma Corticosterone Concentration
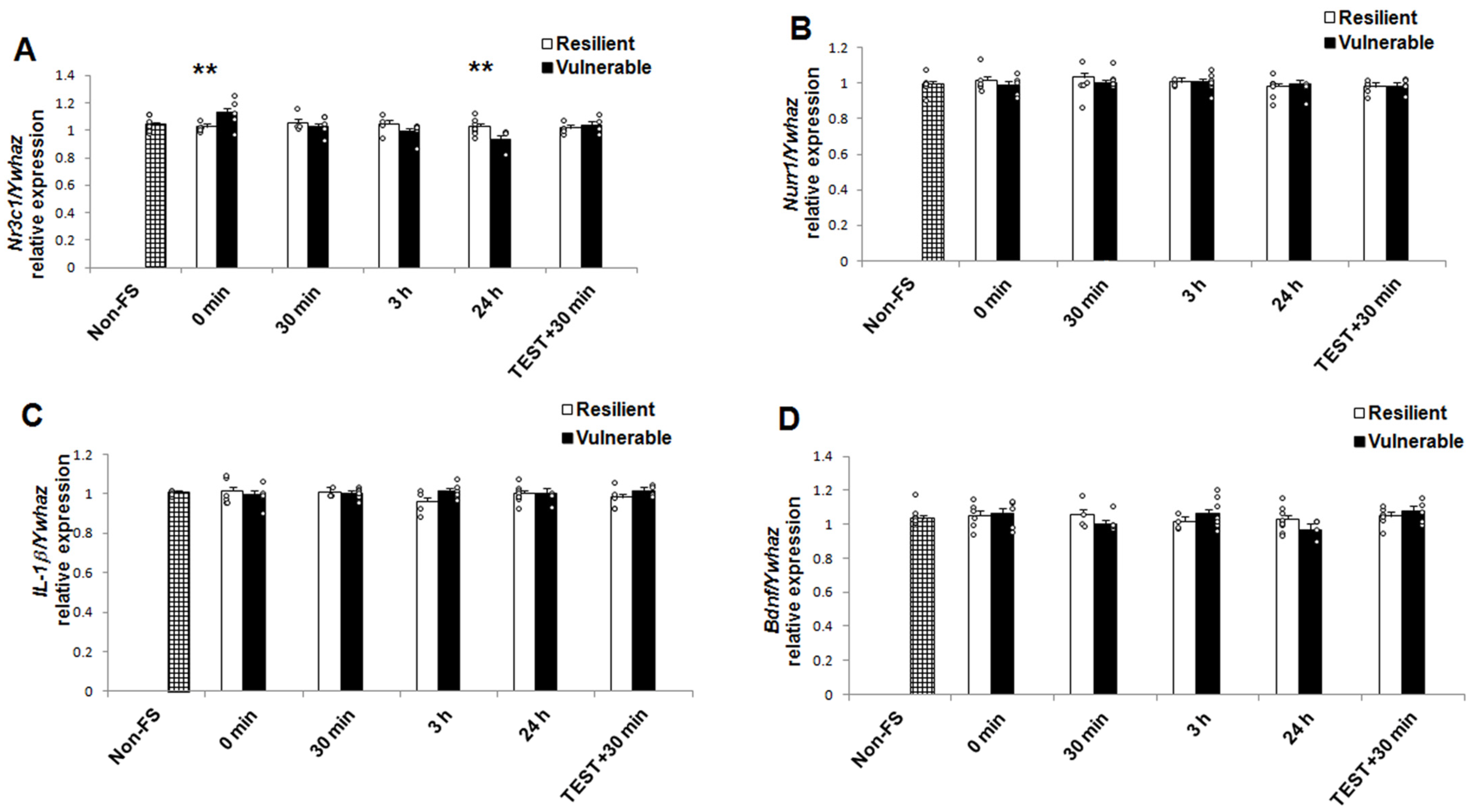
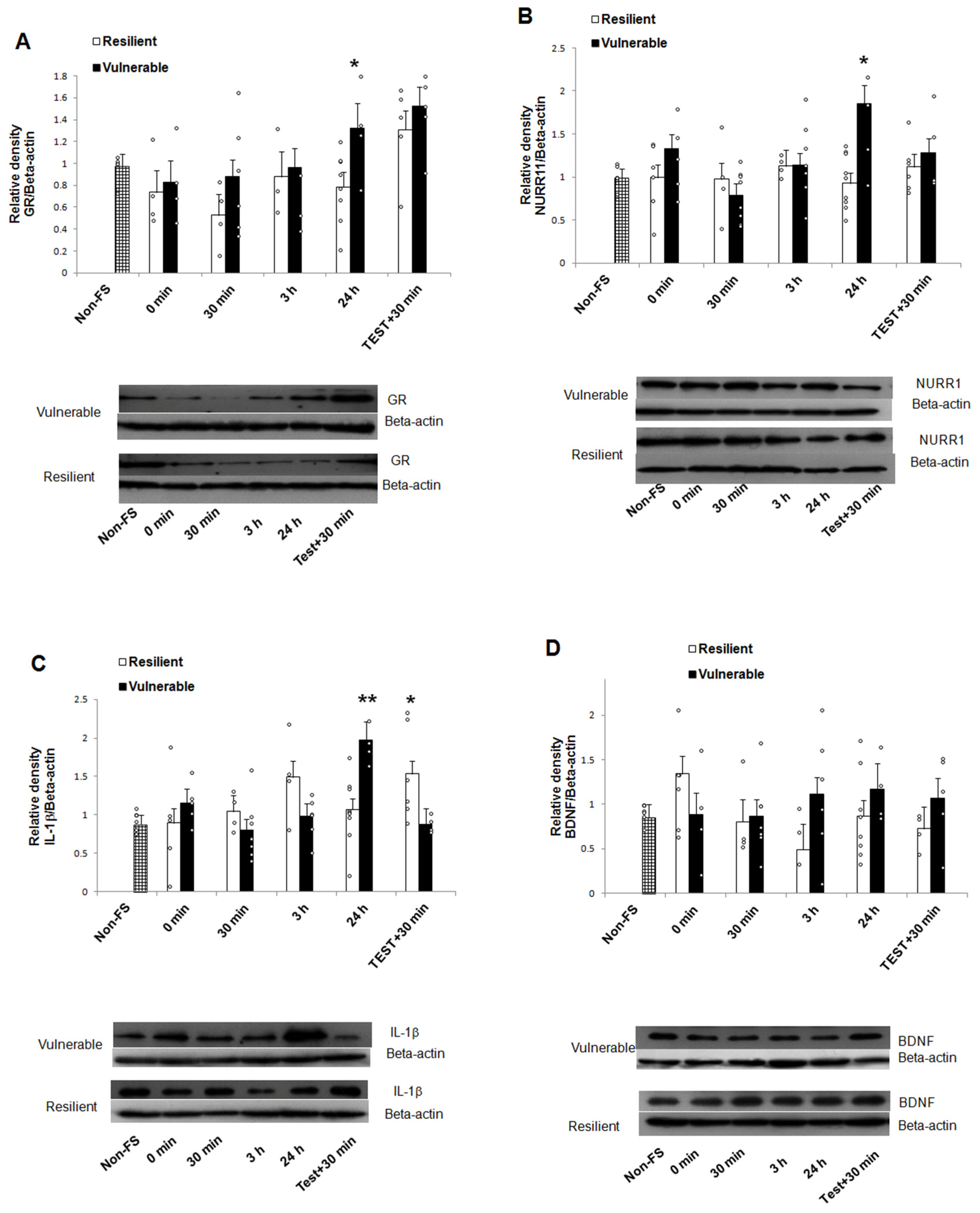
2.3. mRNA and Protein Expression
3. Discussion
4. Materials and Methods
4.1. Animals and Bioethical Guidelines
4.2. Acute Stress Induced by Forced Swimming
4.3. Quantification of Plasma Corticosterone Concentrations after Acute Stress
4.4. mRNA Quantification of Nr3c1, Nurr1, IL-1β, and BDNF
4.5. Protein Quantification of GR, Nurr1, IL-1β, and BDNF
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cathomas, F.; Murrough, J.W.; Nestler, E.J.; Han, M.-H.; Russo, S.J. Neurobiology of resilience: Interface between mind and body. Biol. Psychiatry 2019, 86, 410–420. [Google Scholar] [CrossRef]
- Gottschalk, M.G.; Domschke, K.; Schiele, M.A. Epigenetics underlying susceptibility and resilience relating to daily life stress, work stress, and socioeconomic status. Front. Psychiatry 2020, 11, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Kloet, E.R.; Molendijk, M.L. Coping with the forced swim stressor: Towards understanding an adaptive mechanism. Neural. Plast. 2016, 2016, 6503162. [Google Scholar] [CrossRef] [Green Version]
- Gradus, J.L. Prevalence and prognosis of stress disorders: A review of the epidemiologic literature. Clin. Epidemiol. 2017, 9, 251–260. [Google Scholar] [CrossRef]
- Armario, A.; Nadal, R. Individual differences and the characterization of animal models of psychopathology: A strong challenge and a good opportunity. Front. Pharmacol. 2013, 4, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S. Physiology and neurobiology of stress and adaptation: Central role of the brain. Physiol. Rev. 2002, 87, 873–904. [Google Scholar] [CrossRef] [Green Version]
- Kinlein, S.A.; Karatsoreos, I.N. The hypothalamic-pituitary-adrenal axis as a substrate for stress resilience: Interactions with the circadian clock. Front. Neuroendocrinol. 2020, 56, 100819. [Google Scholar] [CrossRef] [PubMed]
- Reul, J.M.; Collins, A.; Saliba, R.S.; Mifsud, K.R.; Carter, S.D.; Gutierrez-Mecinas, M.; Qian, X.; Linthorst, A.C. Glucocorticoids, epigenetic control and stress resilience. Neurobiol. Stress 2014, 1, 44–59. [Google Scholar] [CrossRef] [Green Version]
- Suri, D.; Vaidya, V.A. The adaptive and maladaptive continuum of stress responses- a hippocampal perspective. Rev. Neurosci. 2015, 26, 415–442. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.D.; Kogan, J.F.; Marrocco, J.; McEwen, B.S. Genomic and epigenomic mechanisms of glucocorticoids in the brain. Nat. Rev. Endocrinol. 2017, 13, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.P.; Conneely, O.M. Neuroendocrine regulation of the hypothalamic pituitary adrenal axis by the nurr1/nur77 subfamily of nuclear receptors. Mol. Endocrinol. 1997, 11, 39–47. [Google Scholar] [CrossRef]
- Campos-Melo, D.; Galleguillos, D.; Sánchez, N.; Gysling, K.; Andrés, M.E. Nur transcription factors in stress and addiction. Front. Mol. Neurosci. 2013, 6, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helbling, J.C.; Minni, A.M.; Pallet, V.; Moisan, M.P. Stress and glucocorticoid regulation of NR4A genes in mice. J. Neurosci. Res. 2014, 92, 825–834. [Google Scholar] [CrossRef]
- Carpentier, R.; Sacchetti, P.; Segard, P.; Staels, B.; Lefebvre, P. The glucocorticoid receptor is a co-regulator of the orphan nuclear receptor Nurr1. J. Neurochem. 2008, 104, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Rojas, P.; Joodmardi, E.; Hong, Y.; Perlmann, T.; Ogren, S.O. Adult mice with reduced Nurr1 expression: An animal model for schizophrenia. Mol. Psychiatry 2007, 12, 756–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eells, J.B.; Lipska, B.K.; Yeung, S.K.; Misler, J.A.; Nikodem, V.M. Nurr1-null heterozygous mice have reduced mesolimbic and mesocortical dopamine levels and increased stress-induced locomotor activity. Behav. Brain Res. 2002, 136, 267–275. [Google Scholar] [CrossRef]
- Rojas, P.; Joodmardi, E.; Perlmann, T.; Ogren, S.O. Rapid increase of Nurr1 mRNA expression in limbic and cortical brain structures related to coping with depression-like behavior in mice. J. Neurosci. Res. 2010, 88, 2284–2293. [Google Scholar] [CrossRef] [PubMed]
- Honkaniemi, J.; Sharp, F.R. Prolonged expression of zinc finger immediate-early gene mRNAs and decreased protein synthesis following kainic acid induced seizures. Eur. J. Neurosci. 1999, 11, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Stroth, N.; Liu, Y.; Aguilera, G.; Eiden, L.E. Pituitary adenylate cyclase-activating polypeptide controls stimulus-transcription coupling in the hypothalamic-pituitary-adrenal axis to mediate sustained hormone secretion during stress. J. Neuroendocrinol. 2011, 23, 944–955. [Google Scholar] [CrossRef] [Green Version]
- Cernackova, A.; Durackova, Z.; Trebaticka, J.; Mravec, B. Neuroinflammation and depressive disorder: The role of the hypothalamus. J. Clin. Neurosci. 2020, 75, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Weber, M.D.; Watkins, L.R.; Maier, S.F. Stress-induced neuroinflammatory priming: A liability factor in the etiology of psychiatric disorders. Neurobiol. Stress 2015, 4, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Réus, G.Z.; Fries, G.R.; Stertz, L.; Badawy, M.; Passos, I.C.; Barichello, T.; Kapczinski, F.; Quevedo, J. The role of inflammation and microglial activation in the pathophysiology of psychiatric disorders. Neuroscience 2015, 300, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Shim, H.S.; An, K.; Starkweather, A.; Kim, K.S.; Shim, I. IL-1 inhibits IL-1β-induced depressive-like behavior and central neurotransmitter alterations. Mediat. Inflamm. 2015, 2015, 941413. [Google Scholar] [CrossRef] [Green Version]
- Koo, J.W.; Duman, R.S. Evidence for IL-1 receptor blockade as a therapeutic strategy for the treatment of depression. Curr. Opin. Investig. Drugs 2009, 10, 664–671. [Google Scholar]
- Rodríguez-Calvo, R.; Tajes, M.; Vázquez-Carrera, M. The NR4A subfamily of nuclear receptors: Potential new therapeutic targets for the treatment of inflammatory diseases. Expert Opin. Ther. Targets 2017, 21, 291–304. [Google Scholar] [CrossRef]
- Murphy, E.P.; Crean, D. Molecular interactions between NR4A orphan nuclear receptors and NF-kB are required for appropriate inflammatory responses and immune cell homeostasis. Biomolecules 2015, 5, 1302–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kageyama, K.; Kagaya, S.; Takayasu, S.; Hanada, K.; Iwasaki, Y.; Suda, T. Cytokines induce NF-κB, Nurr1 and corticotropin-releasing factor gene transcription in hypothalamic 4B cells. Neuroimmunomodulation 2010, 17, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Takayasu, S.; Iwasaki, Y.; Nigawara, T.; Asai, M.; Yoshida, M.; Kageyama, K.; Suda, T. Involvement of nuclear factor-kB and Nurr-1 in cytokine-induced transcription of proopiomelanocortin gene in AtT20 corticotroph cells. Neuroimmunomodulation 2010, 17, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Bensinger, S.; Tontonoz, P. A Nurr1 pathway for neuroprotection. Cell 2009, 137, 26–28. [Google Scholar] [CrossRef] [Green Version]
- Notaras, M.; van den Buuse, M. Neurobiology of BDNF in fear memory, sensitivity to stress, and stress-related disorders. Mol. Psychiatry 2020, 25, 2251–2274. [Google Scholar] [CrossRef] [PubMed]
- Molendijk, M.L.; de Kloet, E.R. Coping with the forced swim stressor: Current state-of-the-art. Behav. Brain Res. 2019, 364, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Volpicelli, F.; Caiazzo, M.; Grego, D.; Consales, C.; Leone, L.; Perrone-Capano, P.; D’Amato, L.C.; di Porzio, U. Bdnf gene is a downstream target of Nurr1 transcription factor in rat midbrain neurons in vitro. J. Neurochem. 2007, 102, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Veldhuis, H.D.; de Krte, C.C.M.M.; de Kloet, E.R. Glucocorticoids facilitate the retention of acquired immobility during forced swimming. Eur. J. Pharmacol. 1985, 115, 211–217. [Google Scholar] [CrossRef]
- Armario, A. The forced swim test: Historical, conceptual and methodological considerations and its relationship with individ ual behavioral traits. Neurosci. Biobehav. Rev. 2021, 128, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Shishkina, G.T.; Kalinina, T.S.; Berezova, I.V.; Bulygina, V.V.; Dygalo, N.N. Resistance to the development of stress-induced behavioral despair in the forced swim test associated with elevated hippocampal Bcl-xl expression. Behav. Brain Res. 2010, 213, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Sequeira-Cordero, A.; Mora-Gallegos, A.; Cuenca-Berger, P.; Fornaguera-Trías, J. Individual differences in the immobility behavior in juvenile and adult rats are associated with monoaminergic neurotransmission and with the expression of corticotropin-releasing factor receptor 1 in the nucleus accumbens. Behav. Brain Res. 2013, 252, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Sequeira-Cordero, A.; Mora-Gallegos, A.; Cuenca-Berger, P.; Fornaguera-Trías, J. Individual differences in the forced swimming test and neurochemical kinetics in the rat brain. Physiol. Behav. 2014, 128, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Sequeira-Cordero, A.; Mora-Gallegos, A.; Cuenca-Berger, P.; Fornaguera-Trías, J. Individual differences in the forced swimming test and the effect of environmental enrichment: Searching for an interaction. Neuroscience 2014, 265, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The rodent forced swim test measures-coping strategy, not depression-like behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef] [Green Version]
- Knight, R.T.; Nakada, T. Cortico-limbic circuits and novelty: A review of EEG and blood flow data. Rev. Neurosci. 1998, 9, 57–70. [Google Scholar] [CrossRef]
- Wingenfeld, K.; Wolf, O.T. Stress, memory, and the hippocampus. Front. Neurol. Neurosci. 2014, 34, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Mifsud, K.R.; Reul, J.M.H.M. Mineralocorticoid and glucocorticoid receptor-mediated control of genomic responses to stress in the brain. Stress 2018, 21, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, A.A.; Lapp, H.E.; Hunter, R.G. Epigenetic mechanisms of the glucocorticoid receptor. Trends Endocrinol. Metab. 2019, 30, 807–818. [Google Scholar] [CrossRef]
- Mifsud, K.R.; Saunderson, E.A.; Spiers, H.; Carter, S.D.; Trollope, A.F.; Mill, J.; Reul, J.M. Rapid down-regulation of glucocorticoid receptor gene expression in the dentate gyrus after acute stress in vivo: Role of DNA methylation and MicroRNA activity. Neuroendocrinology 2017, 104, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Ridder, S.; Chourbaji, S.; Hellweg, R.; Urani, A.; Zacher, C.; Schmid, W.; Zink, M.; Hörtnagl, H.; Flor, H.; Henn, F.A.; et al. Mice with genetically altered glucocorticoid receptor expression show altered sensitivity for stress-induced depressive reactions. J. Neurosci. 2005, 25, 6243–6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetterström, R.H.; Williams, R.; Perlmann, T.; Olson, L. Cellular expression of the immediate early transcription factors Nurr1 and NGF1-B suggests a gene regulatory role in several brain regions including the nigrostriatal dopamine system. Brain Res. Mol. Brain Res. 1996, 41, 111–120. [Google Scholar] [CrossRef]
- Saucedo-Cárdenas, O.; Conneely, O.M. Comparative distribution of NURR1 and NUR77 nuclear receptors in the mouse central nervous system. J. Mol. Neurosci. 1996, 7, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Ota, K.T.; Duman, R.S. The inflammasome: Pathways linking psychological stress, depression, and systemic illnesses. Brain Behav. Immun. 2013, 31, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.S.; Adibfar, A.; Herrmann, N.; Gallagher, D.; Lanctôt, K.L. Evidence for Inflammation-Associated Depression. Curr. Top. Behav. Neurosci. 2017, 31, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Baratta, M.V.; Sprunger, D.B.; Watkins, L.R.; Maier, S.F. Microglia serve as a neuroimmune substrate for stress-induced potentiation of CNS pro-inflammatory cytokine responses. Brain Behav. Immun. 2007, 21, 47–59. [Google Scholar] [CrossRef]
- Ramirez, K.; Fornaguera-Trías, J.; Sheridan, J.F. Stress-Induced microglia activation and monocyte trafficking to the brain underlie the development of anxiety and depression. Curr. Top. Behav. Neurosci. 2017, 31, 155–172. [Google Scholar] [CrossRef]
- Gold, P.W. The organization of the stress system and its dysregulation in depressive illness. Mol. Psychiatry 2015, 20, 32–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.K.; Won, E. The influence of stress on neuroinflammation and alterations in brain structure and function in major depressive disorder. Behav. Brain Res. 2017, 329, 6–11. [Google Scholar] [CrossRef]
- Barrientos, R.M.; Sprunger, D.B.; Campeau, S.; Higgins, E.A.; Watkins, L.R.; Rudy, J.W.; Maier, S.F. Brain-derived neurotrophic factor mRNA downregulation produced by social isolation is blocked by intrahippocampal interleukin-1 receptor antagonist. Neuroscience 2003, 121, 847–853. [Google Scholar] [CrossRef]
- Gutiérrez-Encinas, M.; Trollope, A.F.; Collins, A.; Morfett, H.; Hesketh, S.A.; Kersanté, F.; Reul, J.M.H.M. Long-lasting behavioral responses to stress involve a direct interaction of glucocorticoid receptors with ERK1/2-MSK1-Elk-1 signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 13806–13811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freudenberg, F. Quantitative analysis of Gria1, Gria 2, Dlg1, Dlg4 expression levels in hippocampus following forced swim stress in mice. Sci. Rep. 2019, 9, 14060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Beyer, A.; Aebersold, R. On the dependency of cellular protein levels on mRNA abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Guasti, A.; Olivares-Nazario, M.; Reyes, R.; Martínez-Mota, L. Sex differences in the antidepressant-like effect of fluoxetine in the forced swim test. Pharmacol. Biochem. Behav. 2017, 152, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Montes, P.; Ruiz-Sánchez, E.; Calvillo, M.; Rojas, P. Active coping of prenatally stressed rats in the forced swimming test: Involvement of the Nurr1 gene. Stress 2016, 19, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Lino-de-Oliveira, C.; De Lima, T.C.; de Pádua Carobrez, A. Structure of the rat behaviour in the forced swimming test. Behav. Brain Res. 2005, 158, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Kinoscope. Scoring Behavioral Experiments. Available online: https://sourceforge.net/projects/kinoscope (accessed on 6 August 2020).
- Bonefeld, B.E.; Elfving, B.; Wegener, G. Reference genes for normalization: A study of rat brain tissue. Synapse 2008, 62, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Gubern, C.; Hurtado, O.; Rodríguez, R.; Morales, J.R.; Romera, V.G.; Moro, M.A.; Lizasoain, I.; Serena, J.; Mallolas, J. Validation of housekeeping genes for quantitative real-time PCR in in-vivo and in-vitro models of cerebral ischaemia. BMC Mol. Biol. 2009, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Expression Measure | Individual Behavioral Phenotype (Vulnerable vs. Resilient) | Recovery Times (0, 30 min, 3 h, 24 h, Test + 30 min) | Individual Behavioral Phenotype × Recovery Times | ||||||
|---|---|---|---|---|---|---|---|---|---|
| F | df | p | F | df | p | F | df | p | |
| Nurr1 mRNA | 0.491 | 1.58 | 0.486 | 0.904 | 4.58 | 0.468 | 0.370 | 4.58 | 0.829 |
| Nr3c1 mRNA | 0.279 | 1.58 | 0.600 | 3.575 | 4.58 | 0.011 | 4.397 | 4.58 | 0.004 |
| BDNF mRNA | 0.082 | 1.58 | 0.776 | 1.481 | 4.58 | 0.220 | 1.515 | 4.58 | 0.210 |
| IL-1β mRNA | 0.982 | 1.58 | 0.326 | 0.347 | 4.58 | 0.845 | 1.243 | 4.58 | 0.303 |
| Nurr1 protein | 5.818 | 1.58 | 0.019 | 2.343 | 4.58 | 0.065 | 3.042 | 4.58 | 0.024 |
| GR protein | 4.625 | 1.48 | 0.037 | 4.889 | 4.48 | 0.002 | 0.496 | 4.48 | 0.739 |
| BDNF protein | 1.404 | 1.51 | 0.241 | 0.630 | 4.51 | 0.643 | 1.539 | 4.51 | 0.205 |
| IL-1β protein | 0.165 | 1.55 | 0.686 | 2.676 | 4.55 | 0.027 | 5.482 | 4.55 | 0.001 |
| Genes | Accession Number | Sequence (5′-3′) | Annealing Temp (°C) |
|---|---|---|---|
| Nurr1 | NM_019328.3 | F: AGT CTG ATC AGT GCC CTC GT R: TCA GCA AAG CCA GGA ATC TT | 60 |
| BDNF | NM_001270631 | F: TCC ACC AGG TGA GAA GAG TG R: CGT GGA CGT TTG CTT CTT TC | 61 |
| Nr3c1 | NM_012576 | F: CCT CCC ATT CTA ACC ATC CT R: CTC CCT CTG CTA ACC TGT G | 60 |
| IL-1β | NM_000576.2 | F: CAC CTC TCA AGC AGA GCA CAG R: GGG TTC CAT GGT GAA GTC AAC | 64 |
| Ywhaz | NM_013011.3 | F: TTG AGC AGA AGA CGG AAG GT R: GAA GCA TTG GGG ATC AAG AA | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Sánchez, E.; López-Ramírez, A.M.; Ruiz-Chow, Á.; Calvillo, M.; Reséndiz-Albor, A.A.; Anguiano, B.; Rojas, P. Variability in Behavioral Phenotypes after Forced Swimming-Induced Stress in Rats Is Associated with Expression of the Glucocorticoid Receptor, Nurr1, and IL-1β in the Hippocampus. Int. J. Mol. Sci. 2021, 22, 12700. https://doi.org/10.3390/ijms222312700
Ruiz-Sánchez E, López-Ramírez AM, Ruiz-Chow Á, Calvillo M, Reséndiz-Albor AA, Anguiano B, Rojas P. Variability in Behavioral Phenotypes after Forced Swimming-Induced Stress in Rats Is Associated with Expression of the Glucocorticoid Receptor, Nurr1, and IL-1β in the Hippocampus. International Journal of Molecular Sciences. 2021; 22(23):12700. https://doi.org/10.3390/ijms222312700
Chicago/Turabian StyleRuiz-Sánchez, Elizabeth, Arely M. López-Ramírez, Ángel Ruiz-Chow, Minerva Calvillo, Aldo A. Reséndiz-Albor, Brenda Anguiano, and Patricia Rojas. 2021. "Variability in Behavioral Phenotypes after Forced Swimming-Induced Stress in Rats Is Associated with Expression of the Glucocorticoid Receptor, Nurr1, and IL-1β in the Hippocampus" International Journal of Molecular Sciences 22, no. 23: 12700. https://doi.org/10.3390/ijms222312700