
Aristolochic Acid Induces Renal Fibrosis and Senescence in Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
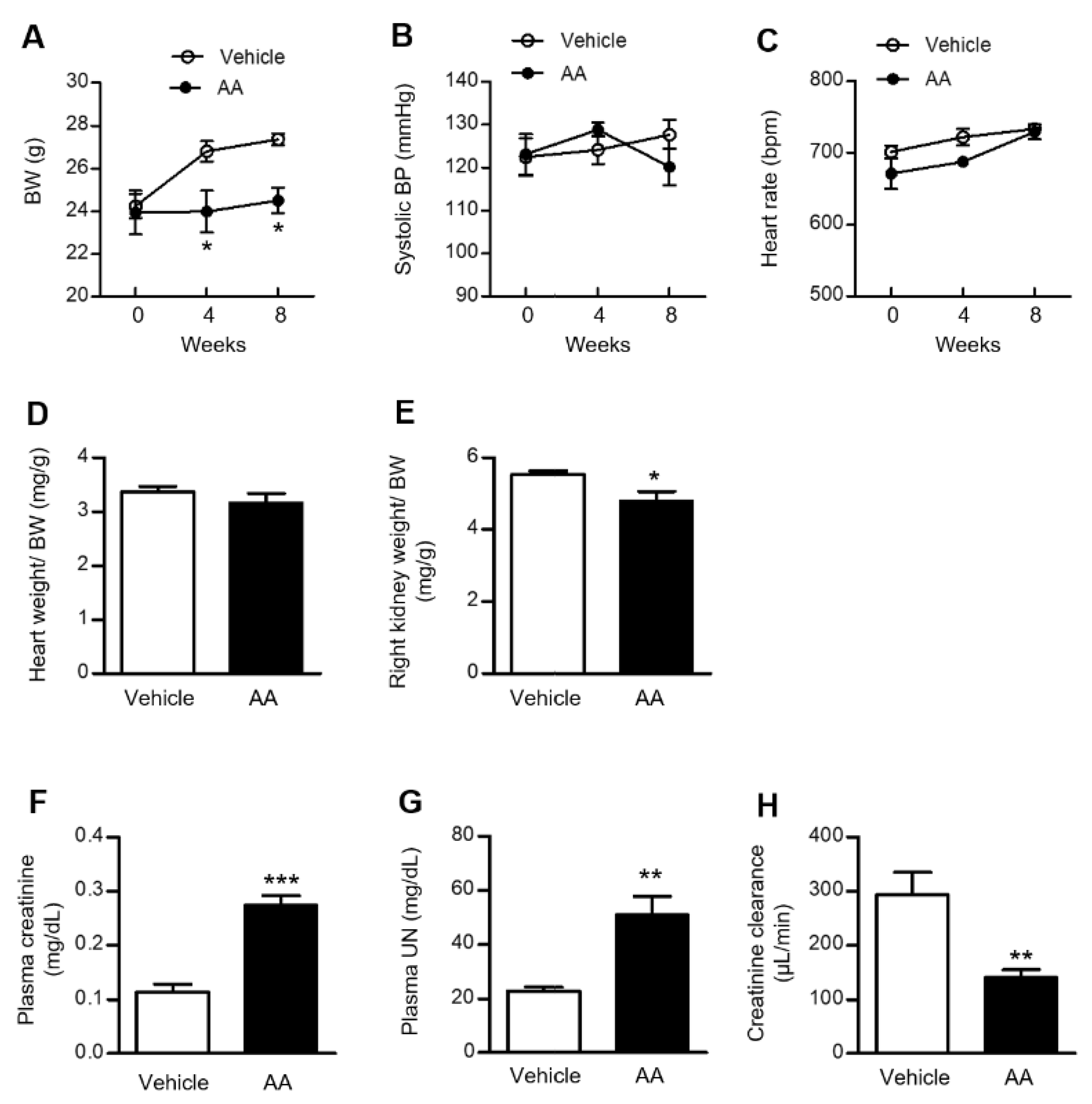
2.1. AA Administration Induced Significant Weight Loss, Renal Atrophy, and a Decline in Renal Function
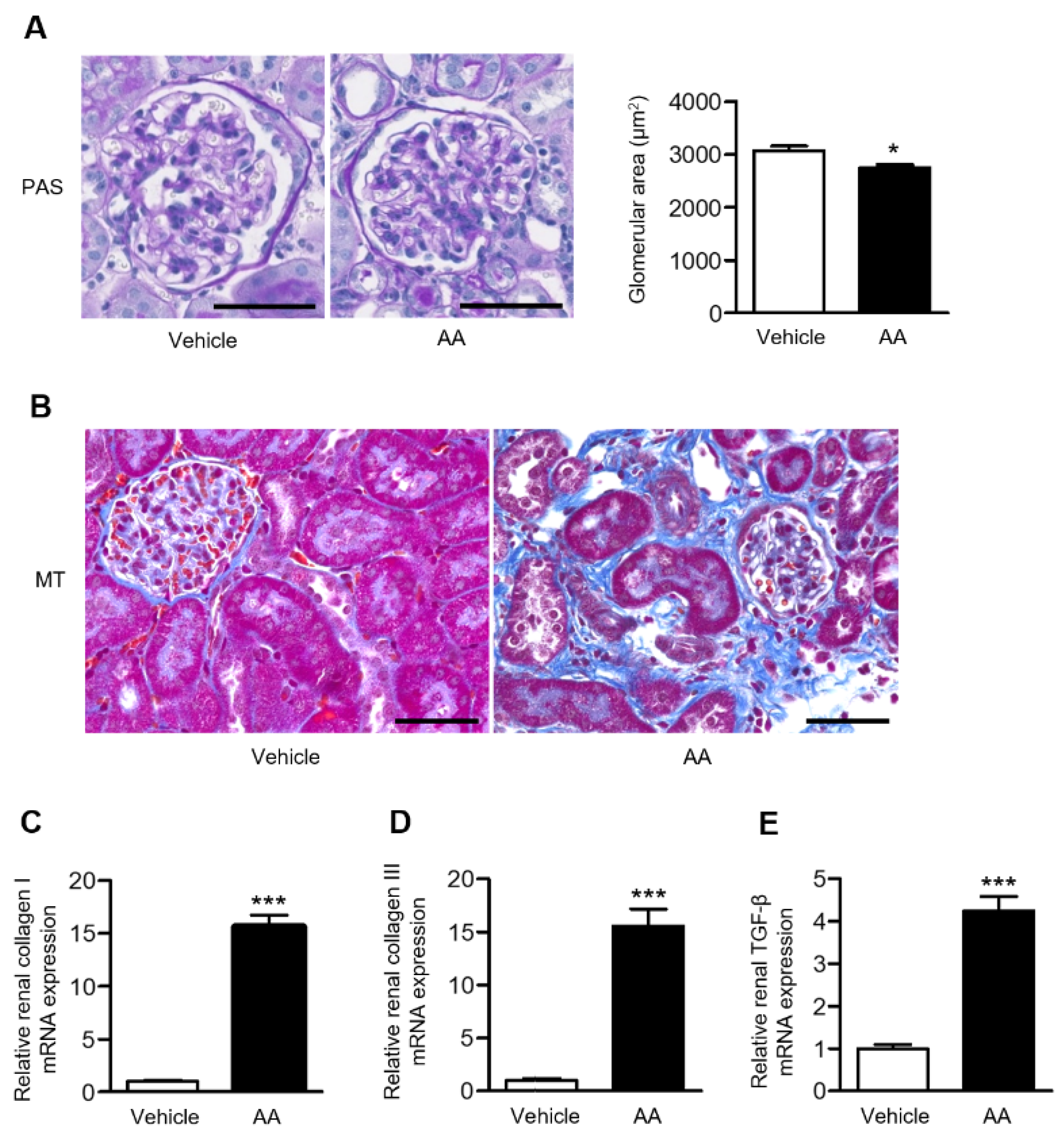
2.2. AA Administration Induced Overt Tubulointerstitial Fibrosis and Significant Upregulation of Fibrosis-Related Gene Expression in the Kidneys
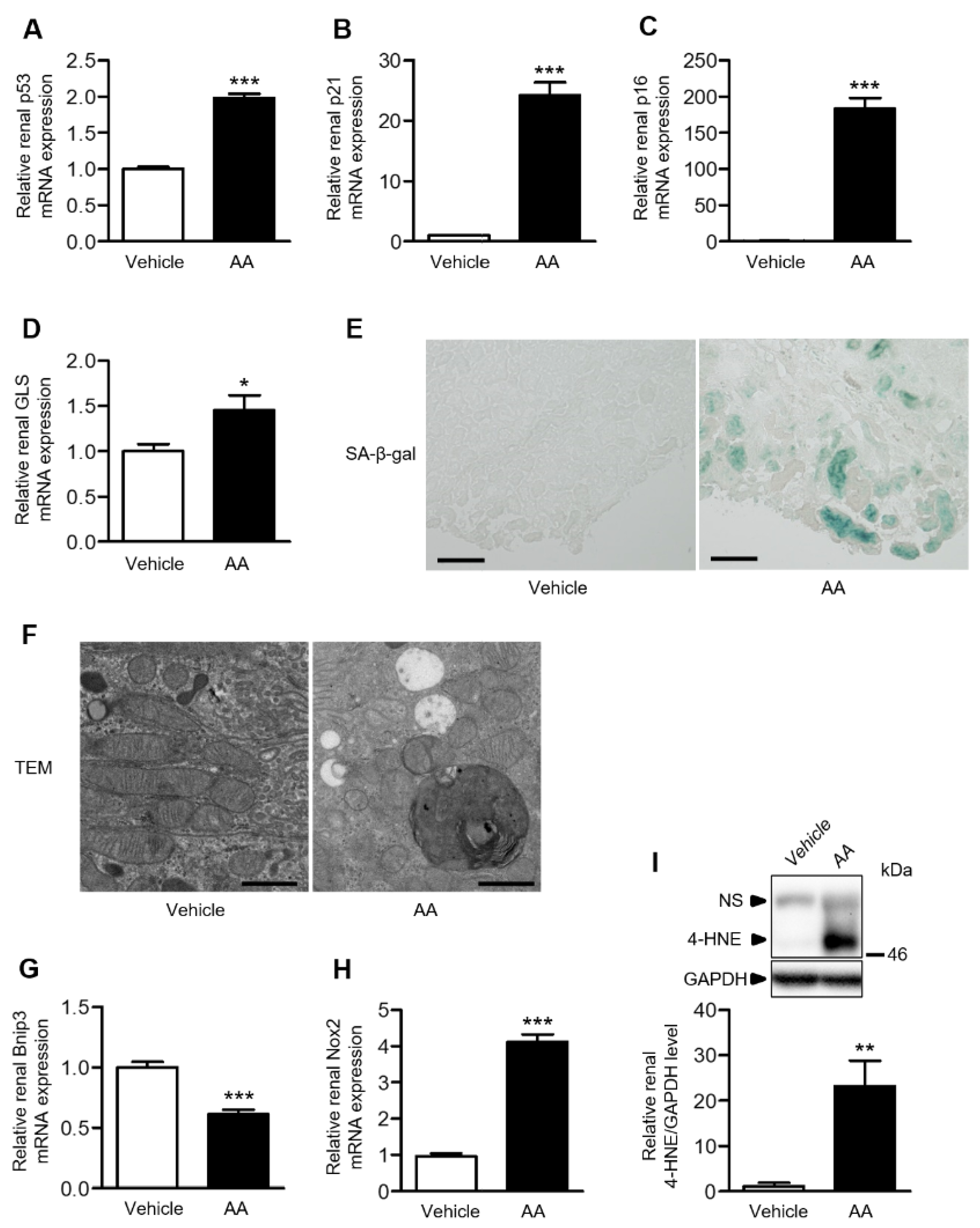
2.3. AA Administration Accelerated Cellular Senescence, Mitochondrial Dysfunction, and Accumulation of Reactive Oxygen Species (ROS) in the Kidneys
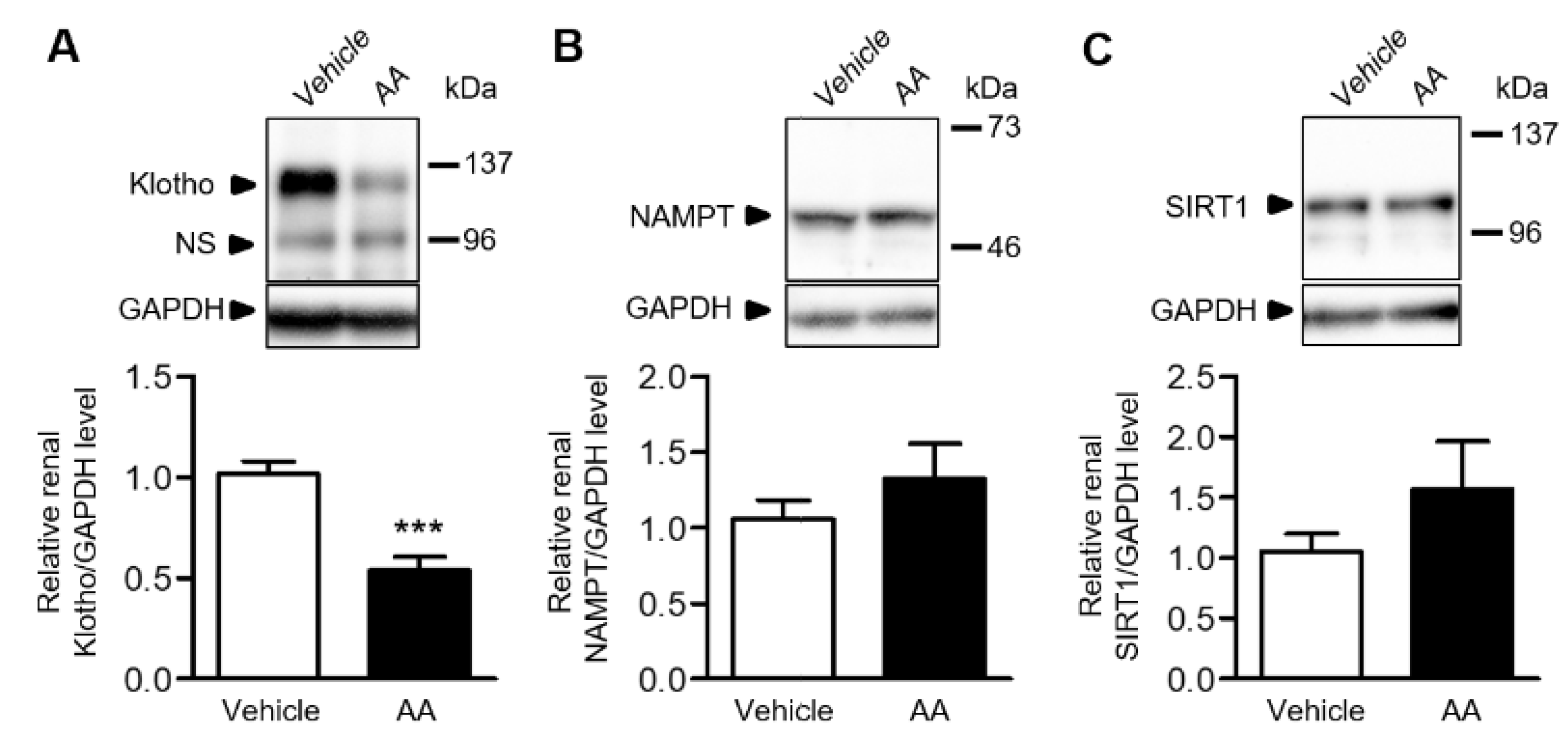
2.4. AA Reduced Renal Klotho Protein Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. BP Measurement
4.3. Real-Time Quantitative Reverse-Transcription PCR Analysis
4.4. Western Blot Analysis
4.5. Histological Analysis
4.6. SA-β-gal Staining
4.7. Electron Microscopy
4.8. Biochemical Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ebert, N.; Jakob, O.; Gaedeke, J.; van der Giet, M.; Kuhlmann, M.K.; Martus, P.; Mielke, N.; Schuchardt, M.; Tölle, M.; Wenning, V.; et al. Prevalence of reduced kidney function and albuminuria in older adults: The Berlin Initiative Study. Nephrol. Dial. Transplant. 2016, 32, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, E.D.; Hughes, J.; Ferenbach, D. Renal Aging: Causes and Consequences. J. Am. Soc. Nephrol. 2017, 28, 407–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhooren, V.; Libert, C. The mouse as a model organism in aging research: Usefulness, pitfalls and possibilities. Ageing Res. Rev. 2013, 12, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Kuro-O, M.; Matsumura, Y.; Aizawa, H.; Kawaguchi, H.; Suga, T.; Utsugi, T.; Ohyama, Y.; Kurabayashi, M.; Kaname, T.; Kume, E.; et al. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature 1997, 390, 45–51. [Google Scholar] [CrossRef]
- Jadot, I.I.; Declèves, A.-E.; Nortier, J.; Caron, N. An Integrated View of Aristolochic Acid Nephropathy: Update of the Literature. Int. J. Mol. Sci. 2017, 18, 297. [Google Scholar] [CrossRef] [Green Version]
- Luciano, R.L.; Perazella, M.A. Aristolochic Acid Nephropathy: Epidemiology, Clinical Presentation, and Treatment. Drug Saf. 2014, 38, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Schmeiser, H.; Schoepe, K.-B.; Wiessler, M. DNA adduct formation of aristolochic acid I and II in vitro and in vivo. Carcinogenesis 1988, 9, 297–303. [Google Scholar] [CrossRef]
- Huang, L.; Scarpellini, A.; Funck, M.; Verderio, E.A.; Johnson, T.S. Development of a Chronic Kidney Disease Model in C57BL/6 Mice with Relevance to Human Pathology. Nephron Extra 2013, 3, 12–29. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Wakui, H.; Azushima, K.; Yamaji, T.; Urate, S.; Suzuki, T.; Abe, E.; Tanaka, S.; Taguchi, S.; Yamada, T. Tissue-specific expression of the SARS-CoV-2 receptor, angiotensin-converting enzyme 2, in mouse models of chronic kidney disease. Sci. Rep. 2021, 11, 16843. [Google Scholar] [CrossRef]
- Krishnamurthy, J.; Torrice, C.; Ramsey, M.R.; Kovalev, G.I.; Al-Regaiey, K.; Su, L.; Sharpless, N.E. Ink4a/Arf expression is a biomarker of aging. J. Clin. Investig. 2004, 114, 1299–1307. [Google Scholar] [CrossRef]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally occurring p16Ink4a-positive cells shorten healthy lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O.; et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [Green Version]
- Melk, A.; Kittikowit, W.; Sandhu, I.; Halloran, K.M.; Grimm, P.; Schmidt, B.M.; Halloran, P. Cell senescence in rat kidneys in vivo increases with growth and age despite lack of telomere shortening. Kidney Int. 2003, 63, 2134–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johmura, Y.; Yamanaka, T.; Omori, S.; Wang, T.-W.; Sugiura, Y.; Matsumoto, M.; Suzuki, N.; Kumamoto, S.; Yamaguchi, K.; Hatakeyama, S.; et al. Senolysis by glutaminolysis inhibition ameliorates various age-associated disorders. Science 2021, 371, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Youle, R.J.; Finkel, T. The Mitochondrial Basis of Aging. Mol. Cell 2016, 61, 654–666. [Google Scholar] [CrossRef] [Green Version]
- Perez-Campo, R.; López-Torres, M.; Cadenas, S.; Rojas, C.; Barja, G. The rate of free radical production as a determinant of the rate of aging: Evidence from the comparative approach. J. Comp. Physiol. B 1998, 168, 149–158. [Google Scholar] [CrossRef]
- Correia-Melo, C.; Marques, F.D.M.; Anderson, R.; Hewitt, G.; Hewitt, R.; Cole, J.; Carroll, B.M.; Miwa, S.; Birch, J.; Merz, A.; et al. Mitochondria are required for pro-ageing features of the senescent phenotype. EMBO J. 2016, 35, 724–742. [Google Scholar] [CrossRef] [PubMed]
- Passos, J.F.; Nelson, G.; Wang, C.; Richter, T.; Simillion, C.; Proctor, C.J.; Miwa, S.; Olijslagers, S.; Hallinan, J.; Wipat, A.; et al. Feedback between p21 and reactive oxygen production is necessary for cell senescence. Mol. Syst. Biol. 2010, 6, 347. [Google Scholar] [CrossRef]
- Kume, S.; Uzu, T.; Horiike, K.; Chin-Kanasaki, M.; Isshiki, K.; Araki, S.-I.; Sugimoto, T.; Haneda, M.; Kashiwagi, A.; Koya, D. Calorie restriction enhances cell adaptation to hypoxia through Sirt1-dependent mitochondrial autophagy in mouse aged kidney. J. Clin. Investig. 2010, 120, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Declèves, A.-E.; Jadot, I.; Colombaro, V.; Martin, B.; Voisin, V.; Nortier, J.; Caron, N.; Habsch, I.; De Prez, E. Protective effect of nitric oxide in aristolochic acid-induced toxic acute kidney injury: An old friend with new assets. Exp. Physiol. 2015, 101, 193–206. [Google Scholar] [CrossRef]
- Zuo, Z.; Lei, H.; Wang, X.; Wang, Y.; Sonntag, W.; Sun, Z. Aging-related kidney damage is associated with a decrease in klotho expression and an increase in superoxide production. AGE 2010, 33, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Kuro-O, M. Klotho as a regulator of oxidative stress and senescence. Biol. Chem. 2008, 389, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Kawamata, Y.; Takahashi, A.; Imai, Y.; Hanyu, A.; Okuma, A.; Takasugi, M.; Yamakoshi, K.; Sorimachi, H.; Kanda, H.; et al. Ablation of the p16INK4a tumour suppressor reverses ageing phenotypes of klotho mice. Nat. Commun. 2015, 6, 7035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigenaga, A.-I.; Tamura, K.; Wakui, H.; Masuda, S.-I.; Azuma, K.; Tsurumi-Ikeya, Y.; Ozawa, M.; Mogi, M.; Matsuda, M.; Uchino, K.; et al. Effect of Olmesartan on Tissue Expression Balance Between Angiotensin II Receptor and Its Inhibitory Binding Molecule. Hypertension 2008, 52, 672–678. [Google Scholar] [CrossRef] [Green Version]
- Azushima, K.; Uneda, K.; Wakui, H.; Ohki, K.; Haruhara, K.; Kobayashi, R.; Haku, S.; Kinguchi, S.; Yamaji, T.; Minegishi, S.; et al. Effects of rikkunshito on renal fibrosis and inflammation in angiotensin II-infused mice. Sci. Rep. 2019, 9, 6201. [Google Scholar] [CrossRef]
- Wakui, H.; Tamura, K.; Masuda, S.-I.; Tsurumi-Ikeya, Y.; Fujita, M.; Maeda, A.; Ohsawa, M.; Azushima, K.; Uneda, K.; Matsuda, M.; et al. Enhanced Angiotensin Receptor-Associated Protein in Renal Tubule Suppresses Angiotensin-Dependent Hypertension. Hypertension 2013, 61, 1203–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohki, K.; Wakui, H.; Kishio, N.; Azushima, K.; Uneda, K.; Haku, S.; Kobayashi, R.; Haruhara, K.; Kinguchi, S.; Yamaji, T.; et al. Angiotensin II Type 1 Receptor-associated Protein Inhibits Angiotensin II-induced Insulin Resistance with Suppression of Oxidative Stress in Skeletal Muscle Tissue. Sci. Rep. 2018, 8, 2846. [Google Scholar] [CrossRef]
- Azushima, K.; Ohki, K.; Wakui, H.; Uneda, K.; Haku, S.; Kobayashi, R.; Haruhara, K.; Kinguchi, S.; Matsuda, M.; Maeda, A.; et al. Adipocyte-Specific Enhancement of Angiotensin II Type 1 Receptor-Associated Protein Ameliorates Diet-Induced Visceral Obesity and Insulin Resistance. J. Am. Heart Assoc. 2017, 6, 004488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsawa, M.; Tamura, K.; Wakui, H.; Maeda, A.; Dejima, T.; Kanaoka, T.; Azushima, K.; Uneda, K.; Tsurumi-Ikeya, Y.; Kobayashi, R.; et al. Deletion of the angiotensin II type 1 receptor–associated protein enhances renal sodium reabsorption and exacerbates angiotensin II–mediated hypertension. Kidney Int. 2014, 86, 570–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, A.; Atobe, Y.; Kadota, T.; Goris, R.; Funakoshi, K. Axonal regeneration through the fibrous scar in lesioned goldfish spinal cord. Neuroscience 2015, 284, 134–152. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urate, S.; Wakui, H.; Azushima, K.; Yamaji, T.; Suzuki, T.; Abe, E.; Tanaka, S.; Taguchi, S.; Tsukamoto, S.; Kinguchi, S.; et al. Aristolochic Acid Induces Renal Fibrosis and Senescence in Mice. Int. J. Mol. Sci. 2021, 22, 12432. https://doi.org/10.3390/ijms222212432
Urate S, Wakui H, Azushima K, Yamaji T, Suzuki T, Abe E, Tanaka S, Taguchi S, Tsukamoto S, Kinguchi S, et al. Aristolochic Acid Induces Renal Fibrosis and Senescence in Mice. International Journal of Molecular Sciences. 2021; 22(22):12432. https://doi.org/10.3390/ijms222212432
Chicago/Turabian StyleUrate, Shingo, Hiromichi Wakui, Kengo Azushima, Takahiro Yamaji, Toru Suzuki, Eriko Abe, Shohei Tanaka, Shinya Taguchi, Shunichiro Tsukamoto, Sho Kinguchi, and et al. 2021. "Aristolochic Acid Induces Renal Fibrosis and Senescence in Mice" International Journal of Molecular Sciences 22, no. 22: 12432. https://doi.org/10.3390/ijms222212432