
Comprehensive Simulation of Ca2+ Transients in the Continuum of Mouse Skeletal Muscle Fiber Types


Abstract
:1. Introduction
2. Results
3. Discussion
3.1. A Model which Includes Four Fiber Types
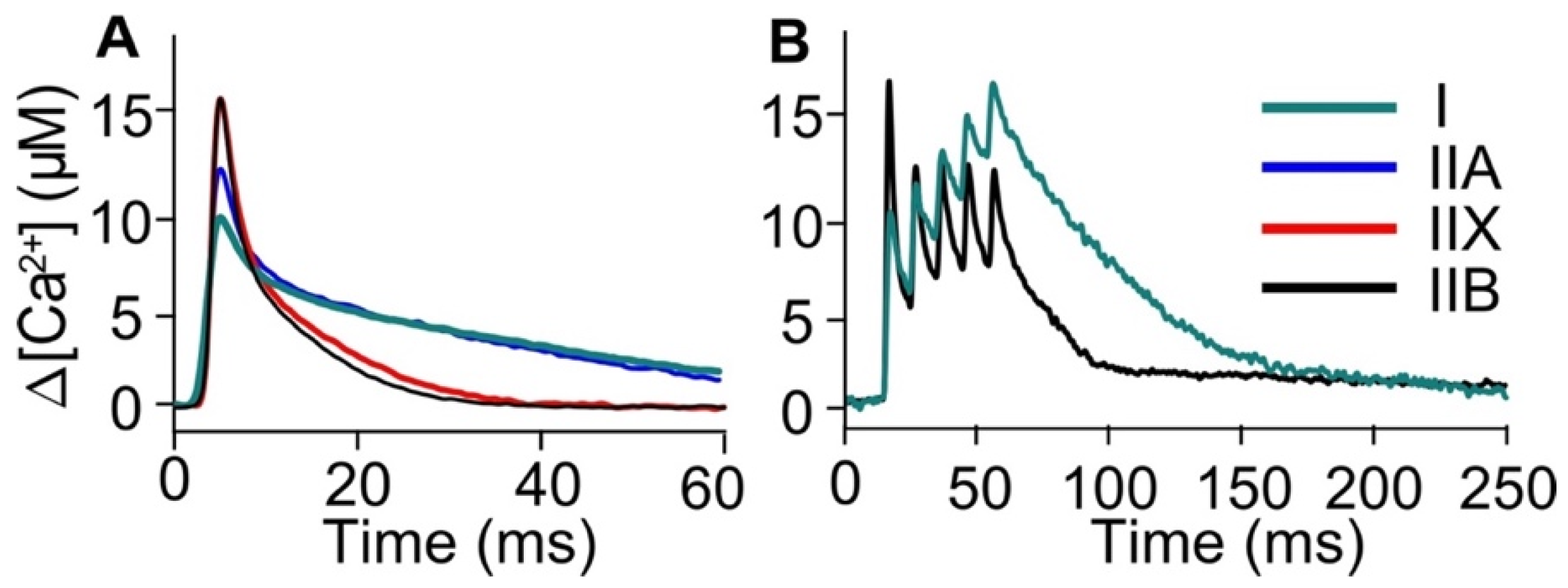
3.2. [Ca2+] Kinetics with a Fast Ca2+ Dye
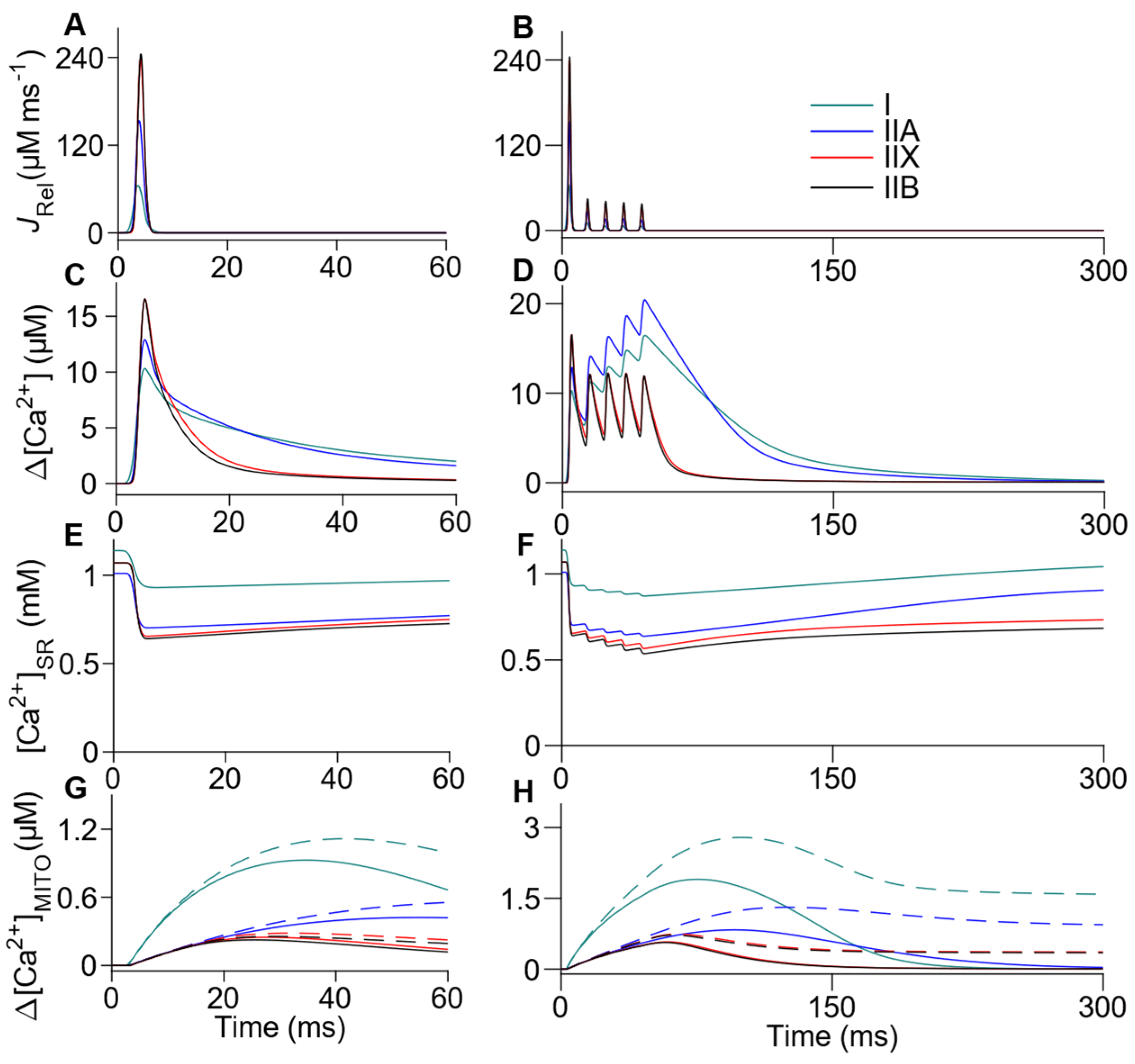
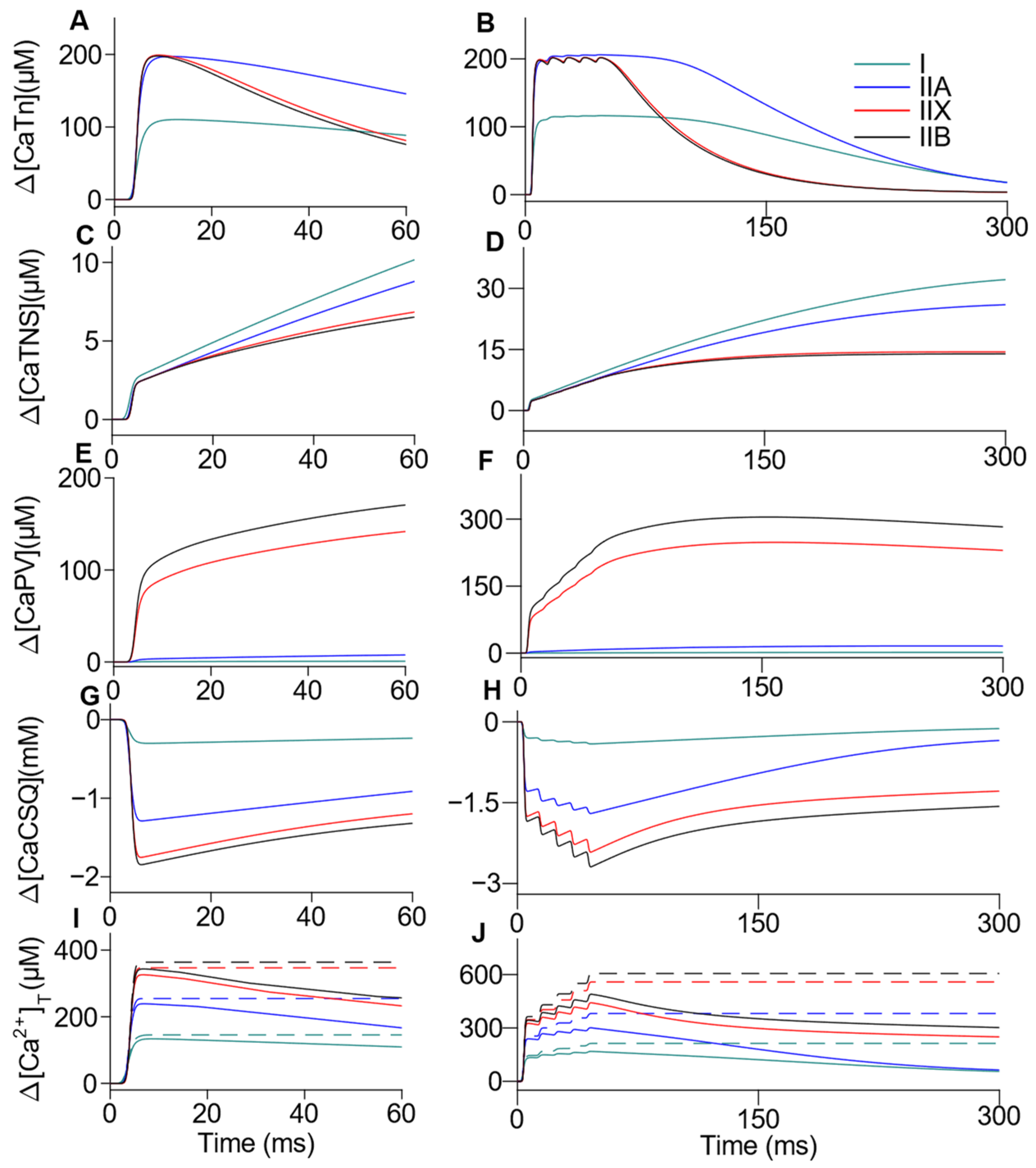
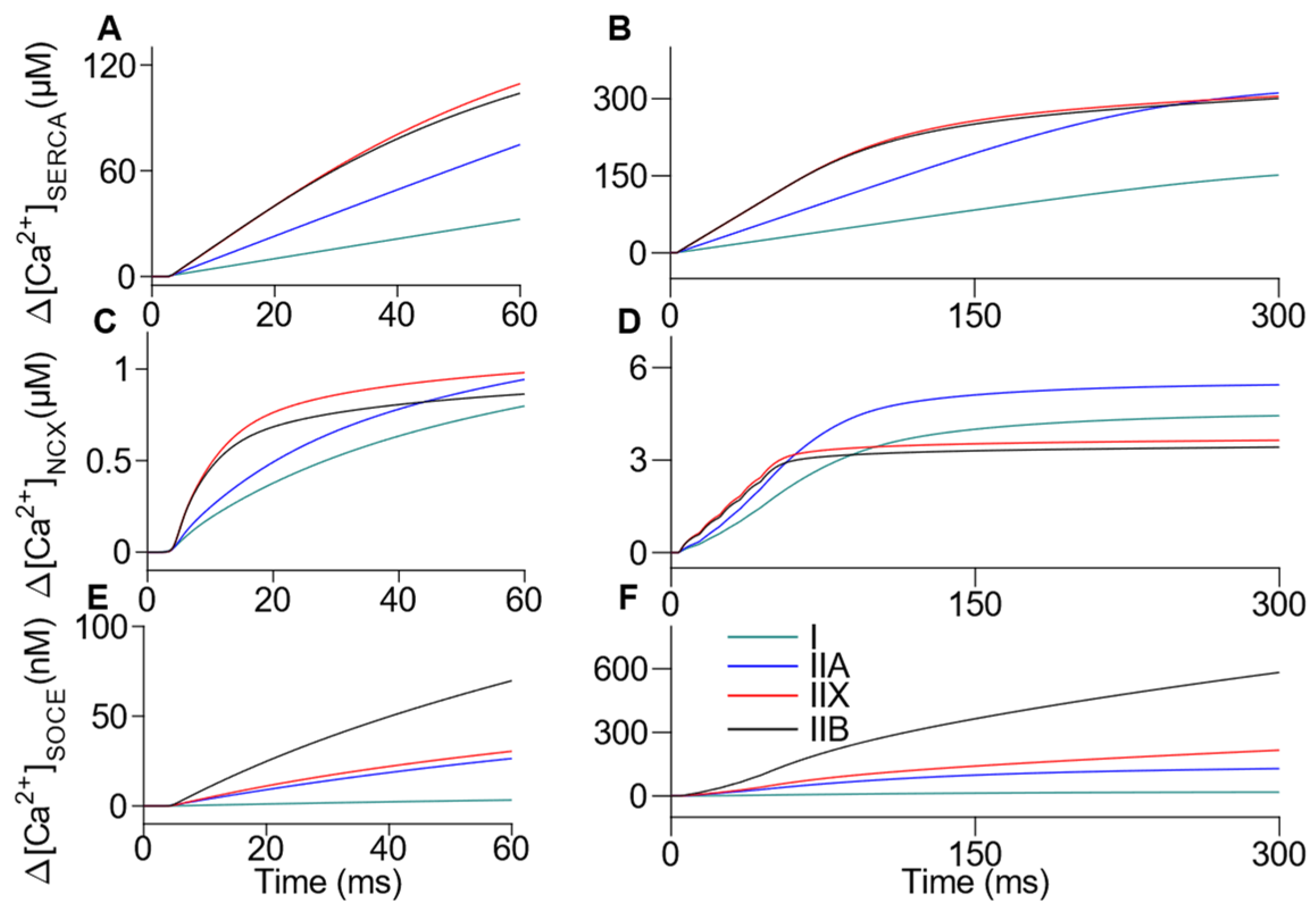
3.3. Comprehensive Integration of Mechanisms Involved in Ca2+ Handling: Sarcoplasm, SR, MITO, NCX and SOCE
3.4. Final Remarks
4. Materials and Methods
4.1. Experimental Single and Tetanic Ca2+ Transients
4.2. Model Description
4.2.1. Release Rate of Ca2+
4.2.2. Reaction of Ca2+ with Sarcomeric Buffers
4.2.3. Muscle Proteins Concentration
4.2.4. Reuptake Rate of Ca2+ by SERCA
4.2.5. The Mitochondrial Ca2+ Uniporter (MCU) Inflow and the Sodium Ca2+ Exchanger (NCE) Outflow
4.2.6. Ca2+ Flux through the NCX
4.2.7. The Effect of Store-operated Ca2+ Entry (SOCE)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calderón, J.C.; Bolaños, P.; Caputo, C. The excitation–contraction coupling mechanism in skeletal muscle. Biophys. Rev. 2014, 6, 133–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiber, J.; Hawkins, A.; Zhang, Z.S.; Wang, S.; Burch, J.; Graham, V.; Ward, C.C.; Seth, M.; Finch, E.; Malouf, N.; et al. STIM1 signalling controls store-operated calcium entry required for development and contractile function in skeletal muscle. Nat. Cell Biol. 2008, 10, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Wei-LaPierre, L.; Carrell, E.M.; Boncompagni, S.; Protasi, F.; Dirksen, R.T. Orai1-dependent calcium entry promotes skeletal muscle growth and limits fatigue. Nat. Commun. 2013, 4, 2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Protasi, F.; Pietrangelo, L.; Boncompagni, S. Calcium entry units (CEUs): Perspectives in skeletal muscle function and disease. J. Muscle Res. Cell Motil. 2021, 42, 233–249. [Google Scholar] [CrossRef] [PubMed]
- Koenig, X.; Choi, R.H.; Launikonis, B.S. Store-operated Ca2+ entry is activated by every action potential in skeletal muscle. Commun. Biol. 2018, 1, 31. [Google Scholar] [CrossRef] [PubMed]
- Calderón, J.C.; Bolaños, P.; Caputo, C. Myosin heavy chain isoform composition and Ca2+ transients in fibres from enzymatically dissociated murine soleus and extensor digitorum longus muscles. J. Physiol. 2010, 588, 267–279. [Google Scholar] [CrossRef]
- Leberer, E.; Pette, D. Immunochemical quantification of sarcoplasmic reticulum Ca-ATPase, of calsequestrin and of parvalbumin in rabbit skeletal muscles of defined fiber composition. Eur. J. Biochem. 1986, 156, 489–496. [Google Scholar] [CrossRef]
- Ferguson, D.G.; Franzini-Armstrong, C. The Ca2+ ATPase content of slow and fast twitch fibers of guinea pig. Muscle Nerve 1988, 11, 561–570. [Google Scholar] [CrossRef]
- Heizmann, C.W. Parvalbumin, and intracellular calcium-binding protein; distribution, properties and possible roles in mammalian cells. Experientia 1984, 40, 910–921. [Google Scholar] [CrossRef]
- Cannell, M.B.; Allen, D.G. Model of calcium movements during activation in the sarcomere of frog skeletal muscle. Biophys. J. 1984, 45, 913–925. [Google Scholar] [CrossRef] [Green Version]
- Baylor, S.M.; Hollingworth, S. Model of Sarcomeric Ca2+ Movements, Including ATP Ca2+ Binding and Diffusion, during Activation of Frog Skeletal Muscle. J. Gen. Physiol. 1998, 112, 297–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderón, J.C.; Bolaños, P.; Caputo, C. Tetanic Ca2+ transient differences between slow- and fast-twitch mouse skeletal muscle fibres: A comprehensive experimental approach. J. Muscle Res. Cell Motil. 2014, 35, 279–293. [Google Scholar] [CrossRef]
- Hollingworth, S.; Kim, M.M.; Baylor, S.M. Measurement and simulation of myoplasmic calcium transients in mouse slow-twitch muscle fibres. J. Physiol. 2012, 590, 575–594. [Google Scholar] [CrossRef] [Green Version]
- Marcucci, L.; Canato, M.; Protasi, F.; Stienen, G.J.M.; Reggiani, C. A 3D diffusional-compartmental model of the calcium dynamics in cytosol, sarcoplasmic reticulum and mitochondria of murine skeletal muscle fibers. PLoS ONE 2018, 13, e0201050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylor, S.M.; Hollingworth, S. Calcium indicators and calcium signalling in skeletal muscle fibres during excitation–contraction coupling. Prog. Biophys. Mol. Biol. 2011, 105, 162–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylor, S.M.; Hollingworth, S. Simulation of Ca2+ movements within the sarcomere of fast-twitch mouse fibers stimulated by action potentials. J. Gen. Physiol. 2007, 130, 283–302. [Google Scholar] [CrossRef] [Green Version]
- Milán, A.F.; Rincón, O.A.; Arango, L.B.; Reutovich, A.A.; Smith, G.L.; Giraldo, M.A.; Bou-Abdallah, F.; Calderón, J.C. Calibration of mammalian skeletal muscle Ca2+ transients recorded with the fast Ca2+ dye Mag-Fluo-4. Biochim. Biophys. Acta-Gen. Subj. 2021, 1865, 129939. [Google Scholar] [CrossRef]
- Luff, A.R.; Atwood, H.L. Changes in the sarcoplasmic reticulum and transverse tubular system of fast and slow skeletal muscles of the mouse during postnatal development. J. Cell Biol. 1971, 51, 369–383. [Google Scholar] [CrossRef]
- Chen, G.; Carroll, S.; Racay, P.; Dick, J.; Pette, D.; Traub, I.; Vrbova, G.; Eggli, P.; Celio, M.; Schwaller, B. Deficiency in parvalbumin increases fatigue resistance in fast-twitch muscle and upregulates mitochondria. Am. J. Physiol. Physiol. 2001, 281, C114–C122. [Google Scholar] [CrossRef]
- Eisenberg, B.R. Quantitative Ultrastructure of Mammalian Skeletal Muscle. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 1983; pp. 73–112. [Google Scholar]
- Williams, D.A.; Head, S.I.; Bakker, A.J.; Stephenson, D.G. Resting calcium concentrations in isolated skeletal muscle fibres of dystrophic mice. J. Physiol. 1990, 428, 243–256. [Google Scholar] [CrossRef]
- Fryer, M.W.; Stephenson, D.G. Total and sarcoplasmic reticulum calcium contents of skinned fibres from rat skeletal muscle. J. Physiol. 1996, 493, 357–370. [Google Scholar] [CrossRef]
- Westerblad, H.; Allen, D.G. Myoplasmic free Mg2+ concentration during repetitive stimulation of single fibres from mouse skeletal muscle. J. Physiol. 1992, 453, 413–434. [Google Scholar] [CrossRef]
- Westerblad, H.; Allen, D.G. The role of sarcoplasmic reticulum in relaxation of mouse muscle; effects of 2,5-di(tert-butyl)-1,4-benzohydroquinone. J. Physiol. 1994, 474, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Sperelakis, N. Cell Physiology Source Book; Elsevier Inc.: Amsterdam, The Netherlands, 2012; ISBN 9780123877383. [Google Scholar]
- Boyman, L.; Chikando, A.C.; Williams, G.S.B.; Khairallah, R.J.; Kettlewell, S.; Ward, C.W.; Smith, G.L.; Kao, J.P.Y.; Lederer, W.J. Calcium movement in cardiac mitochondria. Biophys. J. 2014, 107, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Barclay, C.J.; Launikonis, B.S. Components of activation heat in skeletal muscle. J. Muscle Res. Cell Motil. 2021, 42. [Google Scholar] [CrossRef] [PubMed]
- Kushmerick, M.J.; Moerland, T.S.; Wiseman, R.W. Mammalian skeletal muscle fibers distinguished by contents of phosphocreatine, ATP, and Pi. Proc. Natl. Acad. Sci. USA 1992, 89, 7521–7525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lytton, J.; Westlin, M.; Burk, S.E.; Shull, G.E.; MacLennan, D.H. Functional comparisons between isoforms of the sarcoplasmic or endoplasmic reticulum family of calcium pumps. J. Biol. Chem. 1992, 267, 14483–14489. [Google Scholar] [CrossRef]
- Sembrowich, W.L.; Quintinskie, J.J.; Li, G. Calcium uptake in mitochondria from different skeletal muscle types. J. Appl. Physiol. 1985, 59, 137–141. [Google Scholar] [CrossRef]
- Dash, R.K.; Beard, D.A. Analysis of cardiac mitochondrial Na2+-Ca2+ exchanger kinetics with a biophysical model of mitochondrial Ca2+ handing suggests a 3:1 stoichiometry. J. Physiol. 2008, 586, 3267–3285. [Google Scholar] [CrossRef] [PubMed]
- Donoso, P.; Hidalgo, C. Sodium-calcium exchange in transverse tubules isolated from frog skeletal muscle. Biochim. Biophys. Acta-Biomembr. 1989, 978, 8–16. [Google Scholar] [CrossRef]
- Koenig, X.; Choi, R.H.; Schicker, K.; Singh, D.P.; Hilber, K.; Launikonis, B.S. Mechanistic insights into store-operated Ca 2+ entry during excitation-contraction coupling in skeletal muscle. Biochim. Biophys. Acta-Mol. Cell Res. 2019, 1866, 1239–1248. [Google Scholar] [CrossRef]
- Gillis, J.M.; Thomason, D.; Lefèvre, J.; Kretsinger, R.H. Parvalbumins and muscle relaxation: A computer simulation study. J. Muscle Res. Cell Motil. 1982, 3, 377–398. [Google Scholar] [CrossRef]
- Baylor, S.M.; Hollingworth, S. Sarcoplasmic reticulum calcium release compared in slow-twitch and fast-twitch fibres of mouse muscle. J. Physiol. 2003, 551, 125–138. [Google Scholar] [CrossRef]
- Bakker, A.J.; Cully, T.R.; Wingate, C.D.; Barclay, C.J.; Launikonis, B.S. Doublet stimulation increases Ca2+ binding to troponin C to ensure rapid force development in skeletal muscle. J. Gen. Physiol. 2017, 149, 323–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senneff, S.; Lowery, M.M. Effects of extracellular potassium on calcium handling and force generation in a model of excitation-contraction coupling in skeletal muscle. J. Theor. Biol. 2021, 519, 110656. [Google Scholar] [CrossRef]
- Wang, M.; Sun, J.; Yang, Q. Modeling and simulation of excitation-contraction coupling of fast-twitch skeletal muscle fibers. Technol. Health Care 2020, 28, 13–24. [Google Scholar] [CrossRef]
- Holash, R.J.; MacIntosh, B.R. A stochastic simulation of skeletal muscle calcium transients in a structurally realistic sarcomere model using MCell. PLoS Comput. Biol. 2019, 15, e1006712. [Google Scholar] [CrossRef]
- Bottinelli, R.; Reggiani, C. Human skeletal muscle fibres: Molecular and functional diversity. Prog. Biophys. Mol. Biol. 2000, 73, 195–262. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C. Fiber Types in Mammalian Skeletal Muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, S.; Gorza, L.; Sartore, S.; Saggin, L.; Ausoni, S.; Vianello, M.; Gundersen, K.; LØmo, T. Three myosin heavy chain isoforms in type 2 skeletal muscle fibres. J. Muscle Res. Cell Motil. 1989, 10, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Olsson, K.; Cheng, A.J.; Al-Ameri, M.; Wyckelsma, V.L.; Rullman, E.; Westerblad, H.; Lanner, J.T.; Gustafsson, T.; Bruton, J.D. Impaired sarcoplasmic reticulum Ca2+ release is the major cause of fatigue-induced force loss in intact single fibres from human intercostal muscle. J. Physiol. 2020, 598, 773–787. [Google Scholar] [CrossRef]
- Weiss, N.; Andrianjafiniony, T.; Dupré-Aucouturier, S.; Pouvreau, S.; Desplanches, D.; Jacquemond, V. Altered myoplasmic Ca2+ handling in rat fast-twitch skeletal muscle fibres during disuse atrophy. Pflugers Arch. Eur. J. Physiol. 2010, 459, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Launikonis, B.S.; Stephenson, D.G.; Friedrich, O. Rapid Ca2+ flux through the transverse tubular membrane, activated by individual action potentials in mammalian skeletal muscle. J. Physiol. 2009, 587, 2299–2312. [Google Scholar] [CrossRef]
- Launikonis, B.S.; Murphy, R.M.; Edwards, J.N. Toward the roles of store-operated Ca2+ entry in skeletal muscle. Pflügers Arch.-Eur. J. Physiol. 2010, 460, 813–823. [Google Scholar] [CrossRef]
- Yi, J.; Ma, C.; Li, Y.; Weisleder, N.; Ríos, E.; Ma, J.; Zhou, J. Mitochondrial Calcium Uptake Regulates Rapid Calcium Transients in Skeletal Muscle during Excitation-Contraction (E-C) Coupling. J. Biol. Chem. 2011, 286, 32436–32443. [Google Scholar] [CrossRef] [Green Version]
- Canato, M.; Scorzeto, M.; Giacomello, M.; Protasi, F.; Reggiani, C.; Stienen, G.J.M. Massive alterations of sarcoplasmic reticulum free calcium in skeletal muscle fibers lacking calsequestrin revealed by a genetically encoded probe. Proc. Natl. Acad. Sci. USA 2010, 107, 22326–22331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appelt, D.; Buenviaje, B.; Champ, C.; Franzini-Armstrong, C. Quantitation of ‘junctional feet’ content in two types of muscle fiber from hind limb muscles of the rat. Tissue Cell 1989, 21, 783–794. [Google Scholar] [CrossRef]
- Franzini-Armstrong, C.; Ferguson, D.G.; Champ, C. Discrimination between fast- and slow-twitch fibres of guinea pig skeletal muscle using the relative surface density of junctional transverse tubule membrane. J. Muscle Res. Cell Motil. 1988, 9, 403–414. [Google Scholar] [CrossRef]
- Damiani, E.; Margreth, A. Characterization study of the ryanodine receptor and of calsequestrin isoforms of mammalian skeletal muscles in relation to fibre types. J. Muscle Res. Cell Motil. 1994, 15, 86–101. [Google Scholar] [CrossRef]
- Lee, Y.S.; Ondrias, K.; Duhl, A.J.; Ehrlich, B.E.; Kim, D.H. Comparison of calcium release from sarcoplasmic reticulum of slow and fast twitch muscles. J. Membr. Biol. 1991, 122, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Ecob-Prince, M.S.; Leberer, E. Parvalbumin in mouse muscle in vivo and in vitro. Differentiation 1989, 40, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, K.; Leberer, E.; Lømo, T.; Pette, D.; Staron, R.S. Fibre types, calcium-sequestering proteins and metabolic enzymes in denervated and chronically stimulated muscles of the rat. J. Physiol. 1988, 398, 177–189. [Google Scholar] [CrossRef]
- Rudolf, R.; Mongillo, M.; Magalhaães, P.J.; Pozzan, T. In vivo monitoring of Ca2+ uptake into mitochondria of mouse skeletal muscle during contraction. J. Cell Biol. 2004, 166, 527–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wüst, R.C.I.; Helmes, M.; Martin, J.L.; van der Wardt, T.J.T.; Musters, R.J.P.; van der Velden, J.; Stienen, G.J.M. Rapid frequency-dependent changes in free mitochondrial calcium concentration in rat cardiac myocytes. J. Physiol. 2017, 595, 2001–2019. [Google Scholar] [CrossRef] [Green Version]
- Reddish, F.N.; Miller, C.L.; Deng, X.; Dong, B.; Patel, A.A.; Ghane, M.A.; Mosca, B.; McBean, C.; Wu, S.; Solntsev, K.M.; et al. Rapid subcellular calcium responses and dynamics by calcium sensor G-CatchER+. iScience 2021, 24, 102129. [Google Scholar] [CrossRef] [PubMed]
- Heizmann, C.W.; Berchtold, M.W.; Rowlerson, A.M. Correlation of parvalbumin concentration with relaxation speed in mammalian muscles. Proc. Natl. Acad. Sci. USA 1982, 79, 7243–7247. [Google Scholar] [CrossRef] [Green Version]
- Periasamy, M.; Kalyanasundaram, A. SERCA pump isoforms: Their role in calcium transport and disease. Muscle Nerve 2007, 35, 430–442. [Google Scholar] [CrossRef]
- Linck, B.; Qiu, Z.; He, Z.; Tong, Q.; Hilgemann, D.W.; Philipson, K.D. Functional comparison of the three isoforms of the Na+/Ca2+ exchanger (NCX1, NCX2, NCX3). Am. J. Physiol. Physiol. 1998, 274, C415–C423. [Google Scholar] [CrossRef] [PubMed]
- Mickelson, J.R.; Beaudry, T.M.; Louis, C.F. Regulation of skeletal muscle sarcolemmal ATP-dependent calcium transport by calmodulin and cAMP-dependent protein kinase. Arch. Biochem. Biophys. 1985, 242, 127–136. [Google Scholar] [CrossRef]
- Sacchetto, R.; Margreth, A.; Pelosi, M.; Carafoli, E. Colocalization of the Dihydropyridine Receptor, the Plasma-Membrane Calcium ATPase Isoform 1 and the Sodium/Calcium Exchanger to the Junctional-Membrane Domain of Transverse Tubules of Rabbit Skeletal Muscle. Eur. J. Biochem. 1996, 237, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Activation of Store-Operated Calcium Entry in Airway Smooth Muscle Cells: Insight from a Mathematical Model. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0069598 (accessed on 19 March 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Fiber Type | References | |
|---|---|---|---|---|
| IIA, IIX, and IIB | I | |||
| Sarcomeric volumes | ||||
| VSR | Fiber volume occupied by the SR | 5.5% | 2.9% | [18] |
| VT-tubule | Fiber volume occupied by the T-tubule | 0.4% | 0.2% | [18] |
| Vmito | Fiber volume occupied by MITO | 8.5% | 15.4% | [19] |
| VMS | Fiber volume occupied by the MS | 75% | 81% | [20] |
| Vsarc* | Fiber volume occupied by the sarcoplasm | 85.6% | 81.5% | (Present study) |
| Concentrations | ||||
| [Ca2+]rest,sarc | Resting free Ca2+ in sarcoplasm | 106 nM | 106 nM | [21] |
| [Ca2+]rest,SR | Resting free Ca2+ in SR | 1.01 mM | 1.14 mM | [22] |
| [Mg2+]rest,sarc | Resting free Mg2+ in sarcoplasm | 0.78 mM | 0.78 mM | [23] |
| [Mg2+]T | Total [Mg2+] | 3300 µM | 3300 µM | [24] |
| [Na+]sarc | Sarcoplasmic [Na+] | 10 mM | 10 mM | [25] |
| [Na+]mito | Mitochondrial [Na+] | 5 mM | 5 mM | [26] |
| [Na+]extra | Extracellular [Na+] | 140 mM | 140 mM | [25] |
| [Ca2+]extra | Extracellular [Ca2+] | 1 mM | 1 mM | [25] |
| [Tn]T | Total [Tn] | 240 µM | 120 µM | [13] |
| [TNS]T | Total [TNS] | 240 µM | 240 µM | [13] |
| [PV]T | Total [PV] | 1900–60 µM | 6 µM | [7,9,27] |
| [ATP]T | Total [ATP] | 8 mM | 5 mM | [28] |
| [CSQ]T | Total [CSQ] | 46 mM | 23 mM | [27] |
| [B]T | Total [B] | 20 µM | 20 µM | [14] |
| SERCA | ||||
| VSERCA | Maximum flux rate for SERCA | 2.5–1.4 µM ms−1 | 0.6 µM ms−1 | (Present study) |
| KSERCA | SERCA half-maximum pump [Ca2+] | 0.44 µM | 0.38 µM | [29] |
| hSERCA | SERCA Hill coefficient | 2.1 | 2.2 | [29] |
| MCU | ||||
| VMCU | Maximum flux rate MCU | 18.2 µM s−1 | 74.3 µM s−1 | [13,16] and (Present study) |
| KMCU | MCU half-maximum pump [Ca2+] | 1.2 µM | 1.97 µM | [30] |
| hMCU | MCU Hill coefficient | 2 | 3.5 | [30] |
| NCE | ||||
| VNCE | Maximum flux rate NCE | 2.25 µM s−1 | 9.19 µM s−1 | (Present study) |
| KNCE,Ca2+ | Ca2+ binding constant of NCE | 1.1 mM | 1.1 mM | [14] |
| KNCE,Na+ | Na+ binding constant of NCE | 8.2 mM | 8.2 mM | [31] |
| ΔΨm,mito | Mitochondrial membrane potential | 190 mV | 190 mV | [31] |
| NCX | ||||
| VNCX | Maximum flux rate NCX | 214.5–120.1 µM s−1 | 93.2 µM s−1 | (Present study) |
| KNCX,Ca2+ | Ca2+ binding constant of NCX | 140 µM | 130 µM | [32] |
| KNCX,Na+ | Na+ binding constant of NCX | 14 mM | 11 mM | [32] |
| ΔΨm | Sarcolemmal membrane potential | 80 mV | 80 mV | [25] |
| SOCE | ||||
| VSOCE | Maximum flux rate for SOCE | 35–19 µM s−1 | 15 µM s−1 | [33] and (Present study) |
| KSOCE | SOCE half-maximum pump [Ca2+] | 0.35 mM | 0.35 mM | [33] |
| hSOCE | SOCE Hill coefficient | 4.7 | 4.7 | [33] |
| *Vsarc was calculated as 100%-VSR-VT-tubule-Vmito. | ||||
| Fiber Type | I | IIA | IIX | IIB |
|---|---|---|---|---|
| Peak amplitude (µM ms−1) | 64.8 | 153.6 | 238.8 | 244.5 |
| 10–90% Rise time (ms) | 1.7 | 1.2 | 1.2 | 1.2 |
| Half-width (ms) | 1.9 | 1.4 | 1.3 | 1.3 |
| 90–10% Decay time (ms) | 2.4 | 1.8 | 1.5 | 1.5 |
| fRel (2nd AP) | 0.17 | 0.18 | 0.18 | 0.17 |
| fRel (3rd AP) | 0.10 | 0.11 | 0.17 | 0.15 |
| fRel (4th AP) | 0.10 | 0.11 | 0.16 | 0.15 |
| fRel (5th AP) | 0.10 | 0.10 | 0.15 | 0.14 |
| Fiber Type | I | IIA | IIX | IIB | I | IIA | IIX | IIB |
|---|---|---|---|---|---|---|---|---|
| Single | Tetanic | |||||||
| ∆[Ca2+] (µM) | 10.31 | 12.89 | 16.55 | 16.56 | 16.47 | 20.44 | 16.55 | 16.56 |
| [Ca2+]SR (mM) | 0.93 | 0.7 | 0.65 | 0.64 | 0.87 | 0.64 | 0.57 | 0.54 |
| ∆[Ca2+]MITO (µM) | 0.93 | 0.42 | 0.25 | 0.23 | 1.9 | 0.83 | 0.58 | 0.56 |
| ∆[CaB] (µM) | 0.33 | 0.14 | 0.08 | 0.07 | 1.61 | 0.9 | 0.36 | 0.34 |
| ∆[CaTn] (µM) | 110.23 | 197.28 | 199.09 | 197.75 | 116.39 | 206.05 | 202.16 | 201.85 |
| ∆[CaTNS] (µM) | 10.17 | 8.79 | 6.84 | 6.52 | 32.15 | 25.99 | 14.42 | 13.92 |
| ∆[CaPv] (µM) | 0.78 | 7.63 | 141.78 | 170.66 | 1.76 | 16.27 | 247.96 | 304.5 |
| ∆[CaATP] (µM) | 23.34 | 46.61 | 59.75 | 59.79 | 37.16 | 73.65 | 59.75 | 59.79 |
| ∆[CaDye] (µM) | 3.23 | 4.02 | 5.14 | 5.14 | 5.11 | 6.31 | 5.14 | 5.14 |
| ∆[Ca2+]T (µM) | 133.69 | 238.93 | 325.8 | 343.28 | 168.29 | 300.92 | 441.78 | 489.53 |
| ∆[Ca2+]Rel (µM) | 145.48 | 254.58 | 346.68 | 363.71 | 213.43 | 381.58 | 558.4 | 606.1 |
| ∆[CaCSQ] (mM) | −0.3 | −1.29 | −1.75 | −1.85 | −0.41 | −1.7 | −2.42 | −2.69 |
| ∆[Ca2+]SERCA (µM) | 32.48 | 74.84 | 109.39 | 103.96 | 151.42 | 311.23 | 304.76 | 300.58 |
| ∆[Ca2+]NCX (µM) | 0.8 | 0.94 | 0.98 | 0.86 | 4.44 | 5.44 | 3.64 | 3.42 |
| ∆[Ca2+]SOCE (nM) | 3.29 | 26.39 | 30.44 | 69.73 | 18 | 129 | 215.9 | 582.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rincón, O.A.; Milán, A.F.; Calderón, J.C.; Giraldo, M.A. Comprehensive Simulation of Ca2+ Transients in the Continuum of Mouse Skeletal Muscle Fiber Types. Int. J. Mol. Sci. 2021, 22, 12378. https://doi.org/10.3390/ijms222212378
Rincón OA, Milán AF, Calderón JC, Giraldo MA. Comprehensive Simulation of Ca2+ Transients in the Continuum of Mouse Skeletal Muscle Fiber Types. International Journal of Molecular Sciences. 2021; 22(22):12378. https://doi.org/10.3390/ijms222212378
Chicago/Turabian StyleRincón, Oscar A., Andrés F. Milán, Juan C. Calderón, and Marco A. Giraldo. 2021. "Comprehensive Simulation of Ca2+ Transients in the Continuum of Mouse Skeletal Muscle Fiber Types" International Journal of Molecular Sciences 22, no. 22: 12378. https://doi.org/10.3390/ijms222212378