
Crosstalk during the Carbon–Nitrogen Cycle That Interlinks the Biosynthesis, Mobilization and Accumulation of Seed Storage Reserves

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Storage Reserves of Seeds
3. Seed Formation, Development and Filling
4. Source and Sink Relation
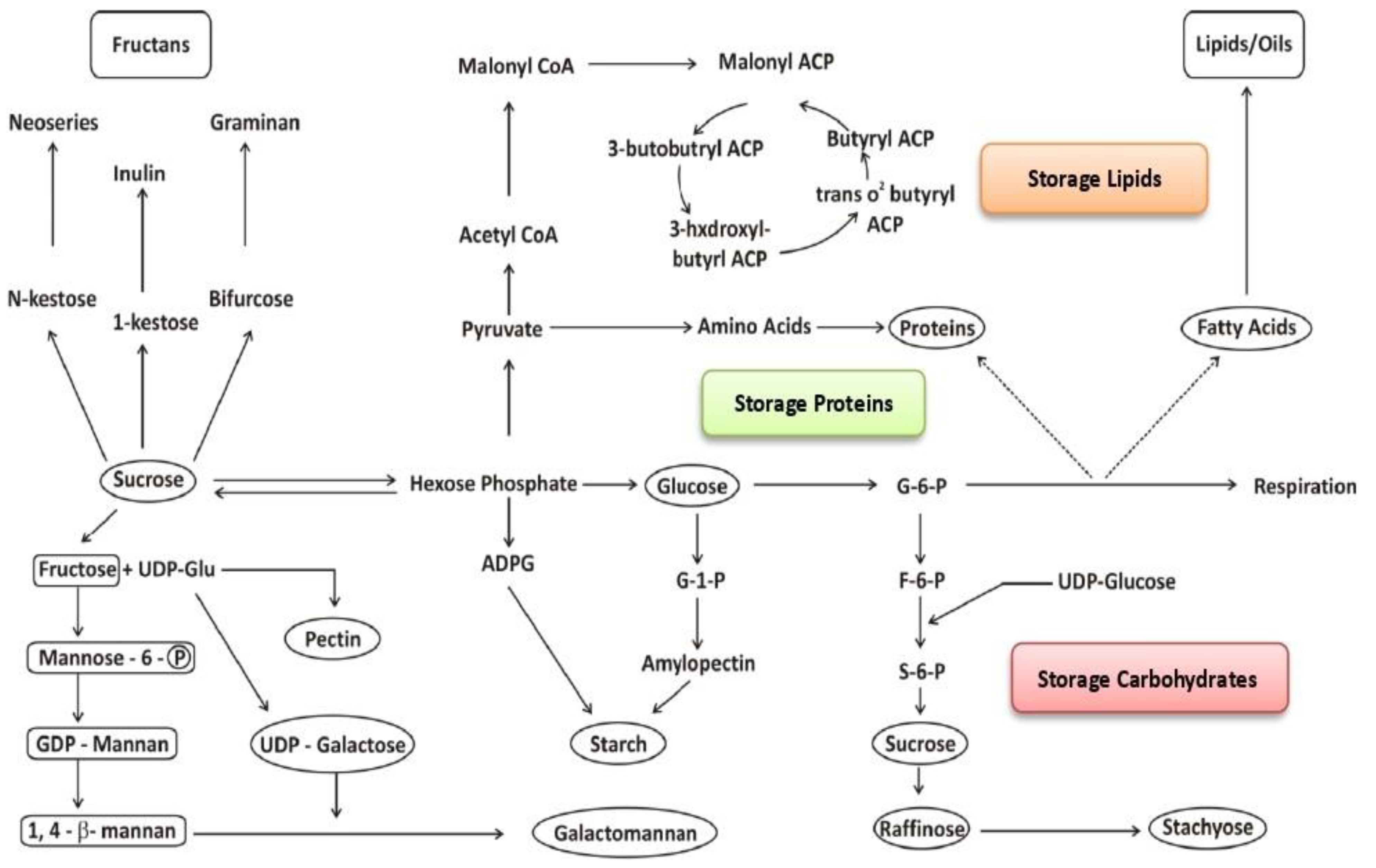
5. Sucrolytic Route Leading to Seed Storage Reserve Biosynthesis
5.1. Protein Biosynthesis
5.2. Lipid Biosynthesis
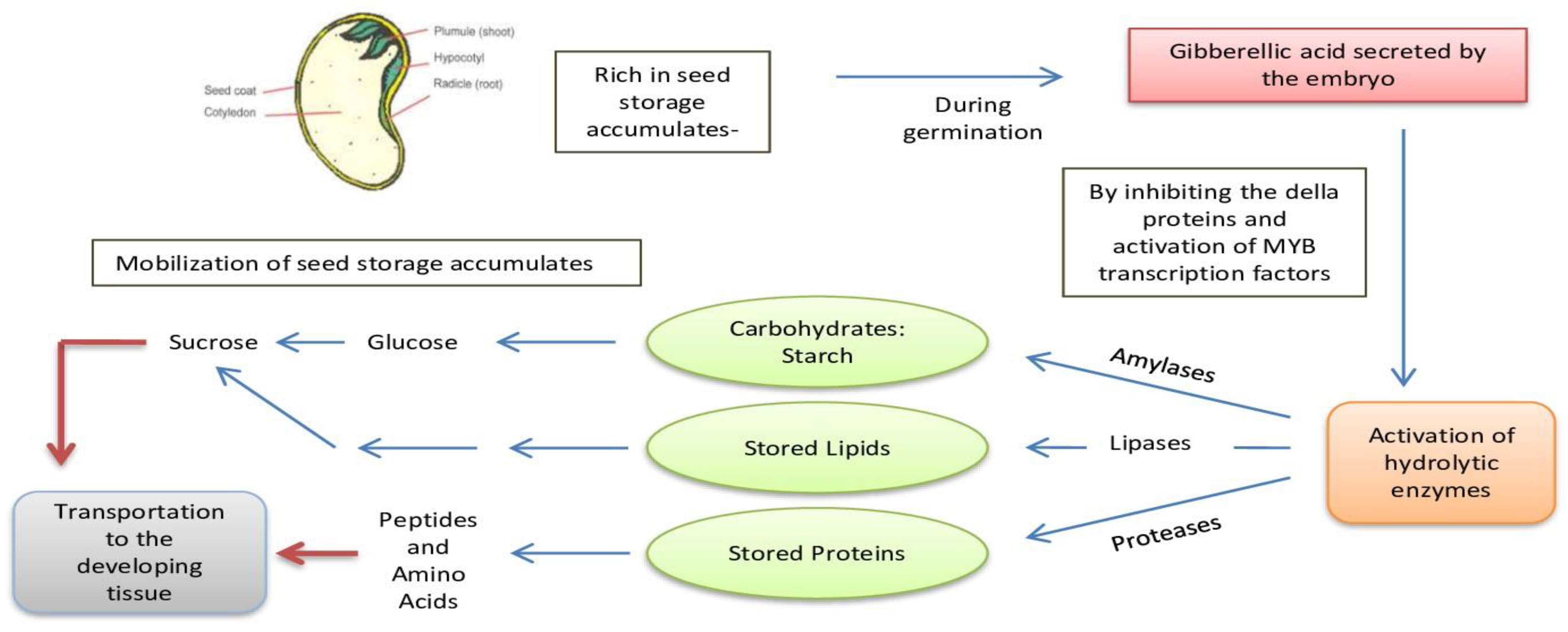
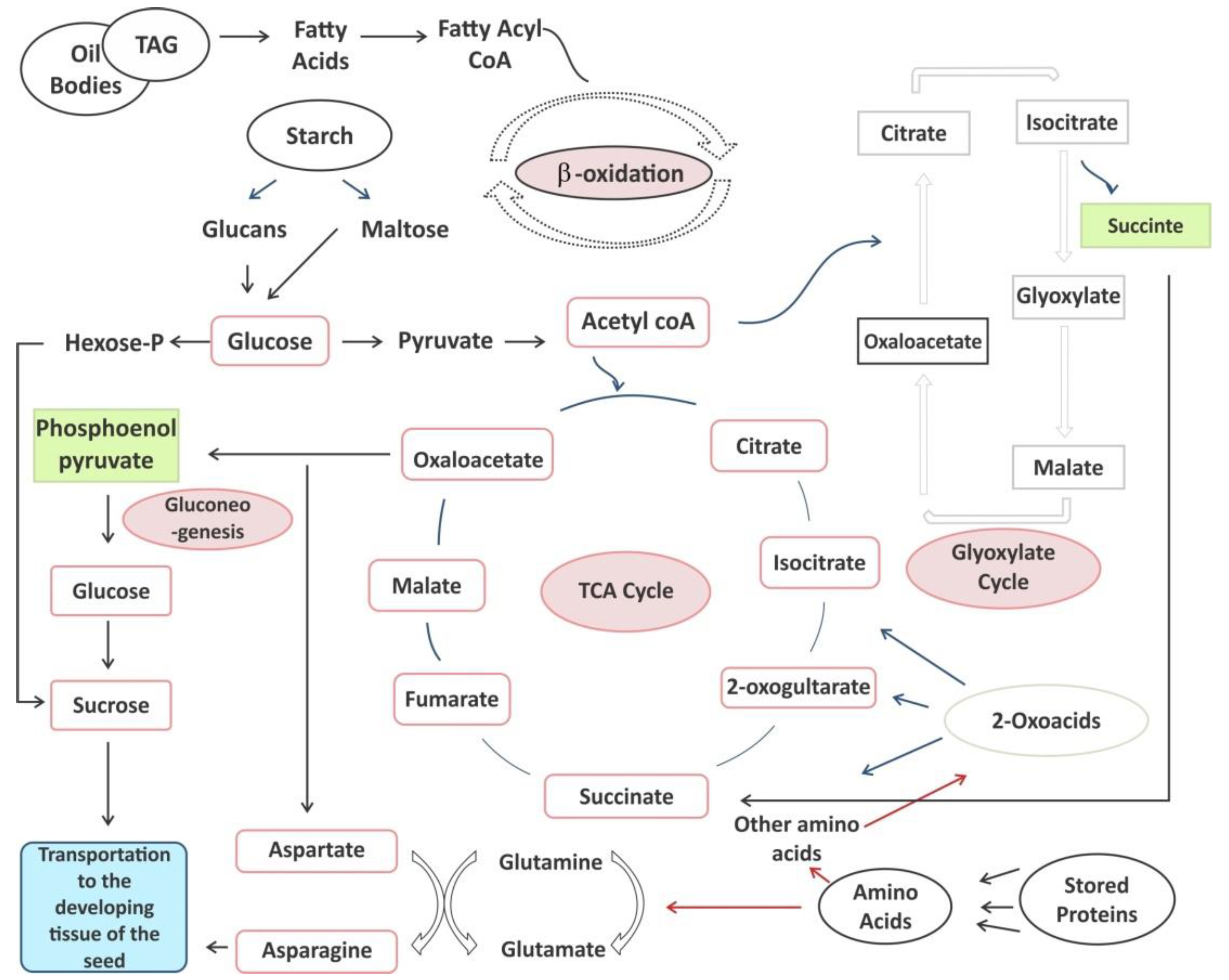
6. Mobilization of Storage Reserves Following Germination
Stored Lipids
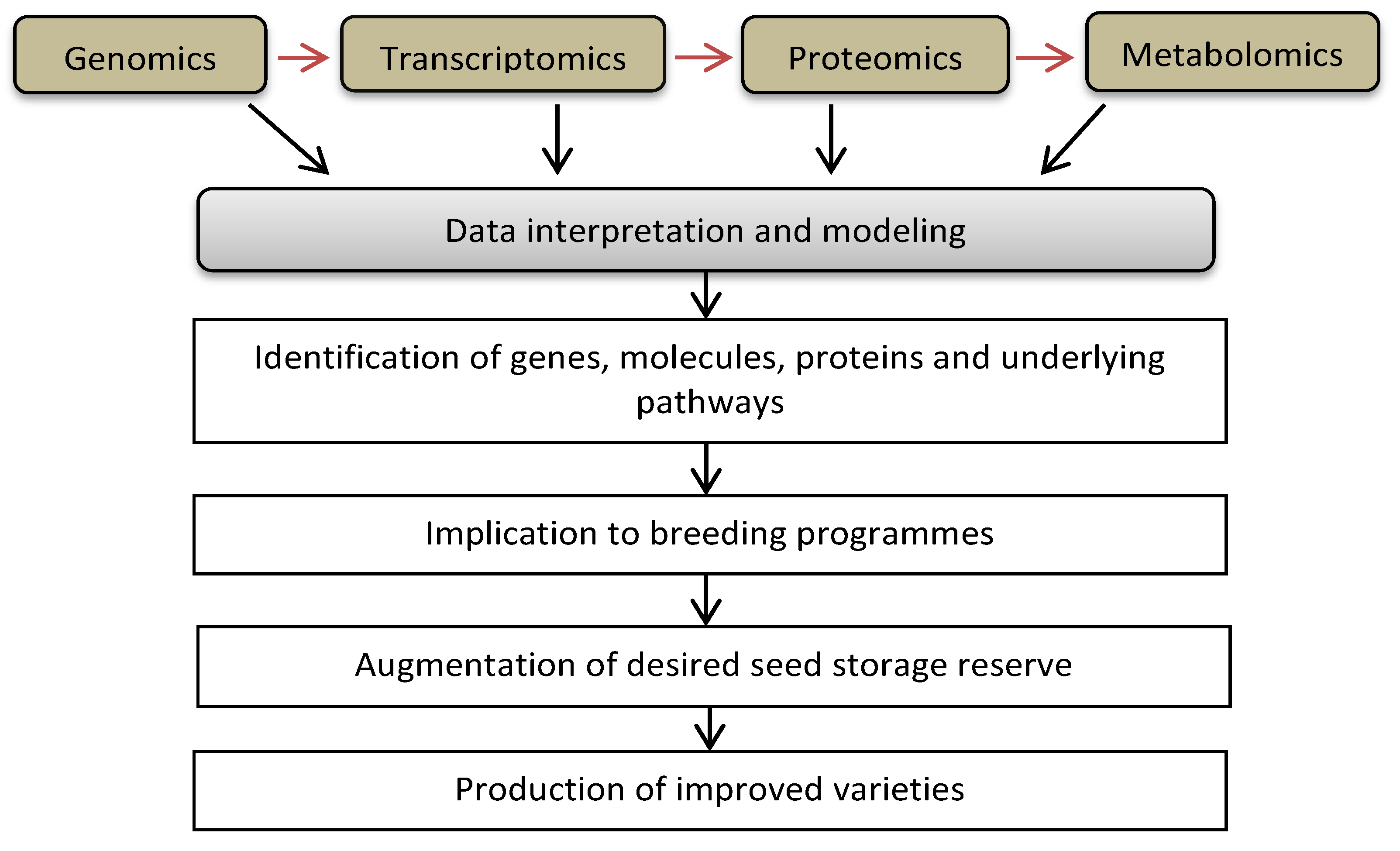
7. Omics Approaches to Augment the Targeted Seed Storage Reserve Accumulation
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mello, J.I.D.O.; Barbedo, C.J.; Salatino, A.; Figueiredo-Ribeiro, R.D.C.L. Reserve carbohydrates and lipids from the seeds of four tropical tree species with different sensitivity to desiccation. Braz. Arch. Biol. 2010, 53, 889–899. [Google Scholar] [CrossRef]
- Thomas, B. Sources Sinks. In Encyclopedia of Applied Plant Science; Elsevier: Oxford, UK, 2017; pp. 119–127. [Google Scholar]
- Chen, L.Q.; Qu, X.Q.; Hou, B.H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M.; Kiegle, E.; Leo, G.; Ezquer, I. Carbohydrate reserves and seed development: An overview. Plant Reprod. 2018, 31, 263–290. [Google Scholar] [CrossRef] [PubMed]
- Koehler, P.; Wieser, H. Chemistry of cereal grains. In Handbook on Sourdough Biotechnology; Springer: Boston, MA, USA, 2013; pp. 11–45. [Google Scholar] [CrossRef]
- Makri, E.; Papalamprou, E.; Doxastakis, G. Study of functional properties of seed storage proteins from indigenous European legume crops (lupin, pea, broad bean) in admixture with polysaccharides. Food Hydrocoll. 2005, 19, 583–594. [Google Scholar] [CrossRef]
- Alves, A.M.; Fernandes, D.C.; Borges, J.F.; Sousa, A.G.D.O.; Naves, M.M.V. Oilseeds native to the Cerrado have fatty acid profile beneficial for cardiovascular health. Rev. Nutr. 2016, 29, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Vera-Sirera, F.; Gomez, M.D.; Perez-Amador, M.A. DELLA proteins, a group of GRAS transcription regulators that mediate gibberellin signaling. In Plant Transcription Factors; Academic Press: Cambridge, MA, USA, 2016; pp. 313–328. [Google Scholar] [CrossRef]
- Guzmán-Ortiz, F.A.; Castro-Rosas, J.; Gómez-Aldapa, C.A.; Mora-Escobedo, R.; Rojas-León, A.; Rodríguez-Marín, M.L.; Falfán-Cortés, R.N.; Román-Gutiérrez, A.D. Enzyme activity during germination of different cereals: A review. Food Rev. Int. 2019, 35, 177–200. [Google Scholar] [CrossRef]
- Cimini, S.; Locato, V.; Vergauwen, R.; Paradiso, A.; Cecchini, C.; Vandenpoel, L.; Verspreet, J.; Courtin, C.M.; D’Egidio, M.G.; Van den Ende, W.; et al. Fructan biosynthesis and degradation as part of plant metabolism controlling sugar fluxes during durum wheat kernel maturation. Front. Plant Sci. 2015, 6, 89. [Google Scholar] [CrossRef] [Green Version]
- Raigond, P.; Ezekiel, R.; Raigond, B. Resistant starch in food: A review. J. Sci. Food Agric. 2015, 95, 1968–1978. [Google Scholar] [CrossRef]
- De Melo, R.B.; Franco, A.C.; Silva, C.O.; Piedade, M.T.F.; Ferreira, C.S. Seed germination and seedling development in response to submergence in tree species of the Central Amazonian floodplains. AoB Plants 2015, 7, plv041. [Google Scholar] [CrossRef] [Green Version]
- Guillon, F.; Larre, C.; Petipas, F.; Berger, A.; Moussawi, J.; Rogniaux, H.; Santoni, A.; Saulnier, L.; Jamme, F.; Miquel, M.; et al. A comprehensive overview of grain development in Brachypodium distachyon variety Bd21. J. Exp. Bot. 2012, 63, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Nishinari, K.; Takemasa, M.; Zhang, H.; Takahashi, R. Storage Plant Polysaccharides: Xyloglucans, Galactomannans, Glucomannans. In Comprehensive Glycoscience; Kamerling, H., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 613–652. [Google Scholar] [CrossRef]
- Kennedy, I.R.; Mwandemele, O.D.; McWhirter, K.S. Estimation of sucrose, raffinose and stachyose in soybean seeds. Food Chem. 1985, 17, 85–93. [Google Scholar] [CrossRef]
- Kermode, A.R.; Finch-Savage, B.E. Desiccation sensitivity in orthodox and recalcitrant seeds in relation to development. In Desiccation and Survival in Plants: Drying without Dying; CABI: Wallingford, UK, 2002; pp. 149–184. [Google Scholar] [CrossRef]
- Dea, I.C.; Morrison, A. Chemistry and interactions of seed galactomannans. Adv. Carbohydr. Chem. Biochem. 1975, 31, 241–312. [Google Scholar] [CrossRef]
- Feng, L.; Yin, J.; Nie, S.; Wan, Y.; Xie, M. Structure and conformation characterization of galactomannan from seeds of Cassia obtusifolia. Food Hydrocoll. 2018, 76, 67–77. [Google Scholar] [CrossRef]
- Busch, V.M.; Delgado, J.F.; Santagapita, P.R.; Wagner, J.R.; Buera, M.D.P. Rheological characterization of vinal gum, a galactomannan extracted from Prosopis ruscifolia seeds. Food Hydrocoll. 2018, 74, 333–341. [Google Scholar] [CrossRef]
- Rashid, F.; Hussain, S.; Ahmed, Z. Extraction purification and characterization of galactomannan from fenugreek for industrial utilization. Carbohydr. Polym. 2018, 180, 88–95. [Google Scholar] [CrossRef]
- Zandi, P.; Basu, S.K.; Khatibani, L.B.; Balogun, M.O.; Aremu, M.O.; Sharma, M.; Kumar, A.; Sengupta, R.; Li, X.; Li, Y.; et al. Fenugreek (Trigonella foenum-graecum L.) seed: A review of physiological and biochemical properties and their genetic improvement. Acta Physiol. Planta 2015, 37, 1714. [Google Scholar] [CrossRef]
- Wieling, W.C. Galactomannans. In Handbook of Hydrocolloids; Woodhead: London, UK, 2009; pp. 228–251. [Google Scholar]
- Behera, S.S.; Ray, R.C. Konjac glucomannan, a promising polysaccharide of Amorphophallus konjac K. Koch in health care. Int. J. Biol. Macromol. 2016, 92, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.S.; Ray, R.C. Nutritional and potential health benefits of konjac glucomannan, a promising polysaccharide of elephant foot yam, Amorphophallus konjac K. Koch: A review. Food Rev. Int. 2017, 33, 22–43. [Google Scholar] [CrossRef]
- Singh, S.; Singh, G.; Arya, S.K. Mannans: An overview of properties and application in food products. Int. J. Biol. Macromol. 2018, 119, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.D.; Yin, J.Y.; Cui, S.W.; Wang, Q.; Wang, S.Y.; Nie, S.P. Plant-derived glucomannans: Sources, preparation methods, structural features, and biological properties. Trends Food Sci. Tech. 2020, 99, 101–116. [Google Scholar] [CrossRef]
- Schulze, E.; Steiger, E. Zur Kenntnis des Paragalaktans. Landwirtsch Vers Stn. 1892, 41, 223–229. [Google Scholar]
- Hirst, E.L.; Jones, J.K.N.; Walder, W.O. Pectic substances. Part 7: The constitution of the galactan from Lupinus albus. J. Chem. Soc. 1947, 25, 1225–1229. [Google Scholar] [CrossRef] [PubMed]
- Warsame, A.O.; Sullivan, D.M.O.; Tosi, P. Seed storage proteins of faba bean (Vicia faba L): Current status and prospects for genetic improvement. J. Agric. Food Chem. 2018, 66, 12617–12626. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Radhika, V.; Rao, V.S.H. Computational approaches for the classification of seed storage proteins. J. Food Sci. Tech. 2015, 52, 4246–4255. [Google Scholar] [CrossRef] [Green Version]
- Frakolaki, G.; Giannou, V.; Topakas, E.; Tzia, C. Chemical characterization and breadmaking potential of spelt versus wheat flour. J. Cereal Sci. 2018, 79, 50–56. [Google Scholar] [CrossRef]
- Jacobsen, S.E. The worldwide potential for quinoa (Chenopodium quinoa Willd.). Food Rev. Int. 2003, 19, 167–177. [Google Scholar] [CrossRef]
- Machado Alencar, N.M.; Steel, C.J.; Alvim, I.D.; de Morais, E.C.; Andre Bolini, H.M. Addition of quinoa and amaranth flour in gluten-free breads: Temporal profile and instrumental analysis. LWT Food Sci. Technol. 2015, 62, 1011–1018. [Google Scholar] [CrossRef]
- Pagamunici, L.M.; Gohara, A.K.; Souza, A.H.P.; Bittencourt, P.R.S.; Torquato, A.S.; Batiston, W.P.; Gomes, S.T.M.; Souza, N.R.; Visentainer, J.V.; Matsushita, M. Using chemometric techniques to characterize gluten-free cookies containing the whole flour of a new quinoa cultivar. J. Braz. Chem. Soc. 2014, 25, 219–228. [Google Scholar] [CrossRef]
- Ahamed, N.T.; Singhal, R.S.; Kulkami, P.R.; Palb, M. Physicochemical and Functional Properties of Chenopodium quinoa Starch. Carbohydr. Polym. 1996, 31, 99–103. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Stølen, O. Quinoa—morphology, phenology and prospects for its production as a new crop in Europe. Eur. J. Agron. 1993, 2, 19–29. [Google Scholar] [CrossRef]
- Guerrieri, N.; Cavaletto, M. Cereals proteins. In Proteins in Food Processing; Yada, R.Y., Ed.; Woodhead: Oxford, UK, 2018; pp. 223–244. [Google Scholar]
- Larkins, B.A.; Wu, Y.; Song, R.; Messing, J. Maize seed storage proteins. In Maize Kernel Development; Larkins, B.A., Ed.; CABI: Wallingford, UK, 2017; pp. 175–189. [Google Scholar] [CrossRef]
- Filho, A.M.M.; Pirozi, M.R.; Borges, J.T.D.S.; Sant’Ana, H.M.P.; Chaves, J.B.P.; Coimbra, J.S.D.R. Quinoa: Nutritional, functional, and antinutritional aspects. Crit. Rev. Food Sci. Nutr. 2017, 57, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Nowak, V.; Du., J.; Charrondière, U.R. Assessment of the nutritional composition of quinoa (Chenopodium quinoa Willd.). Food Chem. 2016, 193, 47–54. [Google Scholar] [CrossRef]
- Ruales, J.; Nair, B.M. Nutritional quality of the protein in quinoa (Chenopodium quinoa, Willd.) seeds. Plant Foods Hum. Nutr. 1992, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Templeman, T.S.; Demaggio, A.E.; Stetler, D.A. Biochemistry of fern spore germination: Globulin storage proteins in Matteuccia struthiopteris L. Plant Physiol. 1987, 85, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Landriscina, L.; Agnello, P.D.; Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M.; Lamacchia, C. Impact of gluten-friendly™ technology on wheat kernel endosperm and gluten protein structure in seeds by light and electron microscopy. Food Chem. 2017, 221, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Rasane, P.; Jha, A.; Sabikhi, L.; Kumar, A.; Unnikrishnan, V.S. Nutritional advantages of oats and opportunities for its processing as value added foods—A review. J. Food Sci. Tech. 2015, 52, 662–675. [Google Scholar] [CrossRef] [Green Version]
- Nuttall, J.G.; O’Leary, G.J.; Panozzo, J.F.; Walker, C.K.; Barlow, K.M.; Fitzgerald, G.J. Models of grain quality in wheat—A review. Field Crops Res. 2017, 202, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.; Cheryan, M. Zein: The industrial protein from corn. Indust. Crops Prod. 2001, 13, 171–192. [Google Scholar] [CrossRef]
- Giteru, S.G.; Oey, I.; Ali, M.A.; Johnson, S.K.; Fang, Z. Effect of kafirin-based films incorporating citral and quercetin on storage of fresh chicken fillets. Food Control. 2017, 80, 37–44. [Google Scholar] [CrossRef]
- Houde, M.; Khodaei, N.; Benkerroum, N.; Karboune, S. Barley protein concentrates: Extraction, structural and functional properties. Food Chem. 2018, 254, 367–376. [Google Scholar] [CrossRef]
- Dias, C.C.; Madruga, M.S.; Pintado, M.M.E.; Almeida, G.H.O.; Alves, A.P.V.; Dantas, F.A.; Bezerra, J.K.G.; de Melo, M.F.F.T.; Vanessa Bordin Viera, V.B.; Soares, J.K.B. Cashew nuts (Anacardium occidentale L.) decrease visceral fat, yet augment glucose in dyslipidemic rats. PLoS ONE 2019, 14, e0225736. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Ge, Z.; Feng, Y.; Wang, X. Lipid characterization of Chinese wild hazelnuts (Corylus mandshurica Maxim.). J. Oleo Sci. 2018, ess18132. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, G.D.; Gómez-Coca, R.B.; Pérez Camino, M.D.C.; Moreda, W.; Barrera-Arellano, D. Chemical characterization of commercial and single-variety avocado oils. Grasas Aceites 2018, 69, e256. [Google Scholar] [CrossRef] [Green Version]
- Aung, W.P.; Bjertness, E.; Htet, A.S.; Stigum, H.; Chongsuvivatwong, V.; Soe, P.P.; Kjøllesdal, M.K.R. Fatty acid profiles of various vegetable oils and the association between the use of palm oil vs. peanut oil and risk factors for non-communicable diseases in Yangon region, Myanmar. Nutrients 2018, 10, 1193. [Google Scholar] [CrossRef] [Green Version]
- Awogbemi, O.; Onuh, E.I.; Inambao, F.L. Comparative study of properties and fatty acid composition of some neat vegetable oils and waste cooking oils. Int. J. Low-Carbon Tech. 2019, 14, 417–425. [Google Scholar] [CrossRef]
- Endo, Y.; Hayashi, C.; Yamanaka, T.; Takayose, K.; Yamaoka, M.; Tsuno, T.; Nakajima, S. Linolenic acid as the main source of acrolein formed during heating of vegetable oils. J. Am. Oil Chem. Soc. 2013, 90, 959–964. [Google Scholar] [CrossRef]
- Gunstone, F.D.; Harwood, J.L.; Dijkstra, A.J. The Lipid Handbook, 3rd ed.; Taylor and Francis: Abingdon, UK, 2007; pp. 703–782. [Google Scholar]
- Gunstone, F.D. Supplies of vegetable oils for non-food purposes. Eur. J. Lipid Sci. Tech. 2011, 113, 3–7. [Google Scholar] [CrossRef]
- Boateng, L.; Ansong, R.; Owusu, W.; Steiner-Asiedu, M. Coconut oil and palm oil’s role in nutrition, health and national development: A review. Ghana Med. J. 2016, 50, 189–196. [Google Scholar] [CrossRef]
- Taylor, D.C. New very long chain fatty acid seed oils produced through introduction of strategic genes into Brassica carinata. Inform. AOCS 2010, 21, 602–607. [Google Scholar]
- Kim, H.U.; Park, M.E.; Lee, K.R.; Suh, M.C.; Chen, G.Q. Variant castor lysophosphatidic acid acyltransferases acylate ricinoleic acid in seed oil. Ind. Crops Prod. 2020, 150, 112245. [Google Scholar] [CrossRef]
- Vijayakumar, B.; Nagendrappa, G.; Prakash, B.S. Synthesis of ester components of spermaceti and a jojoba oil analogue catalyzed by acid activated Indian bentonite under microwave irradiation. Ind. J. Chem. Technol. 2009, 16, 377–381. Available online: http://hdl.handle.net/123456789/6160 (accessed on 5 November 2021).
- Weber, H.; Borisjuk, L.; Wobus, U. Molecular physiology of legume seed development. Annu. Rev. Plant Biol. 2005, 56, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Berger, F.; Hamamura, Y.; Ingouff, M.; Higashiyama, T. Double fertilization–caught in the act. Trends Plant Sci. 2008, 13, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Luo, Y.; Wang, X.; Yu, F. Fruit spray of 24-Epibrassinolide and fruit shade alter pericarp photosynthesis activity and seed lipid accumulation in Styrax tonkinensis. J. Plant Growth Regul. 2018, 37, 1066–1084. [Google Scholar] [CrossRef]
- Shakya, R.; Lal, M.A. Photoassimilate translocation. In Plant Physiology, Development and Metabolism; Springer: Berlin/Heidelberg, Germany, 2018; pp. 227–251. [Google Scholar] [CrossRef]
- Khan, M.A.; Castro-Guerrero, N.A.; Mendoza-Cozatl, D. Moving toward a precise nutrition: Preferential loading of seeds with essential nutrients over non-essential toxic elements. Front. Plant Sci. 2014, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Lin, I.W.; Qu, X.Q.; Sosso, D.; McFarlane, H.E.; Londoño, A.; Samuels, A.L.; Frommer, W.B. A cascade of sequentially expressed sucrose transporters in the seed coat and endosperm provides nutrition for the Arabidopsis embryo. Plant Cell 2015, 27, 607–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matilla, A.J. Seed coat formation: Its evolution and regulation. Seed Sci. Res. 2019, 29, 215–226. [Google Scholar] [CrossRef]
- Fettke, J.; Fernie, A.R. Intracellular and cell-to-apoplast compartmentation of carbohydrate metabolism. Trends Plant Sci. 2015, 20, 490–497. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, H.; Wu, J.; Xie, F. Effect of drought stress during Soybean R2–R6 growth stages on sucrose metabolism in leaf and seed. Int. J. Mol. Sci. 2020, 21, 618. [Google Scholar] [CrossRef] [Green Version]
- Punia, S.; Dhull, S.B.; Sandhu, K.S.; Kaur, M.; Purewal, S.S. Kidney bean (Phaseolus vulgaris) starch: A review. Legume Sci. 2020, 2, e52. [Google Scholar] [CrossRef]
- Liepman, A.H.; Wilkerson, C.G.; Keegstra, K. Expression of cellulose synthase-like (Csl) genes in insect cells reveals that CslA family members encode mannan synthases. Proc. Nat. Acad. Sci. USA 2005, 102, 2221–2226. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.B.; Wang, C.; Muench, D.G.; Ozawa, K.; Franceschi, V.R.; Wu, Y.; Okita, T.W. Messenger RNA targeting of rice seed storage proteins to specific ER subdomains. Nature 2000, 407, 765–767. [Google Scholar] [CrossRef] [PubMed]
- Savojardo, C.; Martelli, P.L.; Fariselli, P.; Casadio, R. DeepSig: Deep learning improves signal peptide detection in proteins. Bioinformatics 2018, 34, 1690–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubis, S.E.; Pike, M.J.; Everett, C.J.; Hill, L.M.; Rawsthorne, S. The import of phosphoenolpyruvate by plastids from developing embryos of oilseed rape, Brassica napus (L.), and its potential as a substrate for fatty acid synthesis. J. Exp. Bot. 2004, 55, 1455–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, Y.; Nagano, Y. Plant acetyl-CoA carboxylase: Structure, biosynthesis, regulation, and gene manipulation for plant breeding. Biosci. Biotech. Biochem. 2004, 68, 1175–1184. [Google Scholar] [CrossRef]
- Fatiha, A.I.D. Plant lipid metabolism. In Advances in Lipid Metabolism; InTech Open: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Tonini, P.P.; Purgatto, E.; Buckeridge, M.S. Effects of abscisic acid, ethylene and sugars on the mobilization of storage proteins and carbohydrates in seeds of the tropical tree Sesbania virgata (Leguminosae). Ann. Bot. 2010, 106, 607–616. [Google Scholar] [CrossRef]
- Graham, I.A. Seed storage oil mobilization. Annu. Rev. Plant Biol. 2008, 59, 115–142. [Google Scholar] [CrossRef] [PubMed]
- Theodoulou, F.L.; Eastmond, P.J. Seed storage oil catabolism: A story of give and take. Curr. Opin. Plant Biol. 2012, 15, 322–328. [Google Scholar] [CrossRef]
- Penfield, S.; Pinfield-Wells, H.; Graham, I.A. Lipid metabolism in seed dormancy. Annu. Plant. Rev. 2018, 27, 133–152. [Google Scholar] [CrossRef]
- Quettier, A.L.; Eastmond, P.J. Storage oil hydrolysis during early seedling growth. Plant Physiol. Biochem. 2009, 47, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Benning, C. Functions of triacylglycerols during plant development and stress. Curr. Opin. Biotechnol. 2018, 49, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Masterson, C.; Wood, C. Mitochondrial β-oxidation of fatty acids in higher plants. Physiol. Planta 2000, 109, 217–224. [Google Scholar] [CrossRef]
- Glevarec, G.; Bouton, S.; Jaspard, E.; Riou, M.T.; Cliquet, J.B.; Suzuki, A.; Limami, A.M. Respective roles of the glutamine synthetase/glutamate synthase cycle and glutamate dehydrogenase in ammonium and amino acid metabolism during germination and post-germinative growth in the model legume Medicago truncatula. Planta 2004, 219, 286–297. [Google Scholar] [CrossRef]
- Lea, P.J.; Sodek, L.; Parry, M.A.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Ann. App. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]
- Lehmann, T.; Ratajczak, L. The pivotal role of glutamate dehydrogenase (GDH) in the mobilization of N and C from storage material to asparagine in germinating seeds of yellow lupine. J. Plant Physiol. 2008, 165, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Tan-Wilson, A.L.; Wilson, K.A. Mobilization of seed protein reserves. Physiol. Planta. 2012, 145, 140–153. [Google Scholar] [CrossRef]
- Grudkowska, M.; Zagdańska, B. Multifunctional role of plant cysteine proteinases. Acta. Biochim. Pol. 2004, 51, 609–624. [Google Scholar] [CrossRef] [Green Version]
- Boyd, P.M.; Barnaby, N.; Tan-Wilson, A.; Wilson, K.A. Cleavage specificity of the subtilisin-like protease C1 from soybean. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2002, 1596, 269–282. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.; Nonogaki, H. Mobilization of stored reserves. In Seeds; Springer: Berlin/Heidelberg, Germany, 2013; pp. 183–246. [Google Scholar] [CrossRef]
- Robinson, D.G.; Neuhaus, J.M.M. Receptor-mediated sorting of soluble vacuolar proteins: Myths, facts, and a new model. J. Exp. Bot. 2016, 67, 4435–4449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrazzini, E.; Mainieri, D.; Marrano, C.A.; Vitale, A. Where do protein bodies of cereal seeds come from? Front. Plant Sci. 2016, 7, 1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Hao, P.; Chen, G.; Han, C.; Li, X.; Zeller, F.J.; Hsam, S.L.; Hu, Y.; Yan, Y. Molecular cloning, phylogenetic analysis, and expression profiling of endoplasmic reticulum molecular chaperone BiP genes from bread wheat (Triticum aestivum L.). BMC Plant Biol. 2014, 14, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangelsen, E.; Kilian, J.; Harter, K.; Jansson, C.; Wanke, D.; Sundberg, E. Transcriptome analysis of high-temperature stress in developing barley caryopses: Early stress responses and effects on storage compound biosynthesis. Mol. Plant. 2011, 4, 97–115. [Google Scholar] [CrossRef]
- Peng, T.; Sun, H.; Du, Y.; Zhang, J.; Li, J.; Liu, Y.; Zhao, Y.; Zhao, Q. Characterization and expression patterns of microRNAs involved in rice grain filling. PLoS ONE. 2013, 8, e54148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, R.; Zhu, Z.; Hu, J.; Qian, Q.; Dai, J.; Ding, Y. Identification and expression analysis of microRNAs at the grain filling stage in rice (Oryza sativa L.) via deep sequencing. PLoS ONE 2013, 8, e0057863. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Fu, Z.; Lv, P.; Peng, Q.; Ding, D.; Li, W.; Tang, J. Identification and characterization of microRNAs during maize grain filling. PLoS ONE 2015, 10, e0125800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurai, T.; Wakayama, M.; Abiko, T.; Yanagisawa, S.; Aoki, N.; Ohsugi, R. Introduction of the ZmDof1 gene into rice enhances carbon and nitrogen assimilation under low-nitrogen conditions. Plant Biotechnol. J. 2011, 9, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Weselake, R.J.; Taylor, D.C.; Rahman, M.H.; Shah, S.; Laroche, A.; McVetty, P.B.; Harwood, J.L. Increasing the flow of carbon into seed oil. Biotech. Adv. 2009, 27, 866–878. [Google Scholar] [CrossRef]
- Li, J.; Baroja-Fernández, E.; Bahaji, A.; Muñoz, F.J.; Ovecka, M.; Montero, M.; Sesma, M.T.; Alonso-Casajús, N.; Almagro, G.; Sánchez-López, A.M.; et al. Enhancing sucrose synthase activity results in increased levels of starch and ADP-glucose in maize (Zea mays L.) seed endosperms. Plant Cell Physiol. 2013, 54, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.J.; Oszvald, M.; Jesus, C.; Rajulu, C.; Griffiths, C.A. Increasing crop yield and resilience with trehalose 6-phosphate: Targeting a feast–famine mechanism in cereals for better source–sink optimization. J. Exp. Bot. 2017, 68, 4455–4462. [Google Scholar] [CrossRef] [Green Version]
- Terskikh, V.V.; Feurtado, J.A.; Borchardt, S.; Giblin, M.; Abrams, S.R.; Kermode, A.R. In vivo 13C NMR metabolite profiling: Potential for understanding and assessing conifer seed quality. J. Exp. Bot. 2005, 56, 2253–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terskikh, V.V.; Zeng, Y.; Feurtado, J.A.; Giblin, M.; Abrams, S.R.; Kermode, A.R. Deterioration of western red cedar (Thuja plicata Donn ex D. Don) seeds: Protein oxidation and in vivo NMR monitoring of storage oils. J. Exp. Bot. 2008, 59, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Terskikh, V.; Kermode, A.R. In vivo nuclear magnetic resonance metabolite profiling in plant seeds. In Seed Dormancy; Humana Press: Totowa, NJ, USA, 2011; pp. 307–318. [Google Scholar] [CrossRef]
- Sun, Y.; Jiao, G.; Liu, Z.; Zhang, X.; Li, J.; Guo, X.; Du, W.; Du, J.; Francis, F.; Zhao, Y.; et al. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes. Front. Plant Sci. 2017, 8, 298. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, M.; Tak, Y.; Bhatia, S.; Asthir, B.; Lorenzo, J.M.; Amarowicz, R. Crosstalk during the Carbon–Nitrogen Cycle That Interlinks the Biosynthesis, Mobilization and Accumulation of Seed Storage Reserves. Int. J. Mol. Sci. 2021, 22, 12032. https://doi.org/10.3390/ijms222112032
Kaur M, Tak Y, Bhatia S, Asthir B, Lorenzo JM, Amarowicz R. Crosstalk during the Carbon–Nitrogen Cycle That Interlinks the Biosynthesis, Mobilization and Accumulation of Seed Storage Reserves. International Journal of Molecular Sciences. 2021; 22(21):12032. https://doi.org/10.3390/ijms222112032
Chicago/Turabian StyleKaur, Manpreet, Yamini Tak, Surekha Bhatia, Bavita Asthir, José M. Lorenzo, and Ryszard Amarowicz. 2021. "Crosstalk during the Carbon–Nitrogen Cycle That Interlinks the Biosynthesis, Mobilization and Accumulation of Seed Storage Reserves" International Journal of Molecular Sciences 22, no. 21: 12032. https://doi.org/10.3390/ijms222112032