
Ketoprofen Combined with UVA Irradiation Exerts Higher Selectivity in the Mode of Action against Melanotic Melanoma Cells than against Normal Human Melanocytes

 , , , and
, , , and 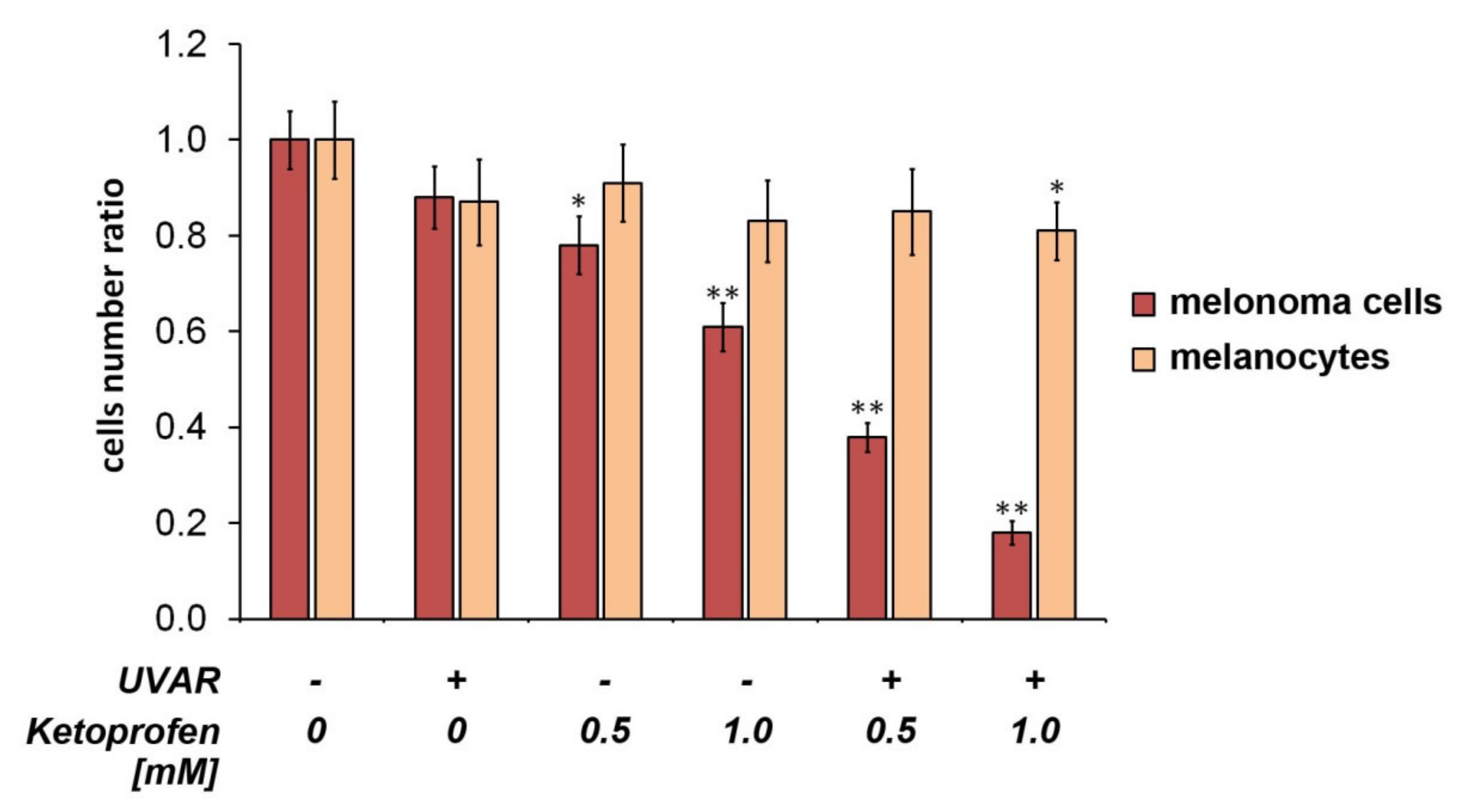{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect on Cell Proliferation
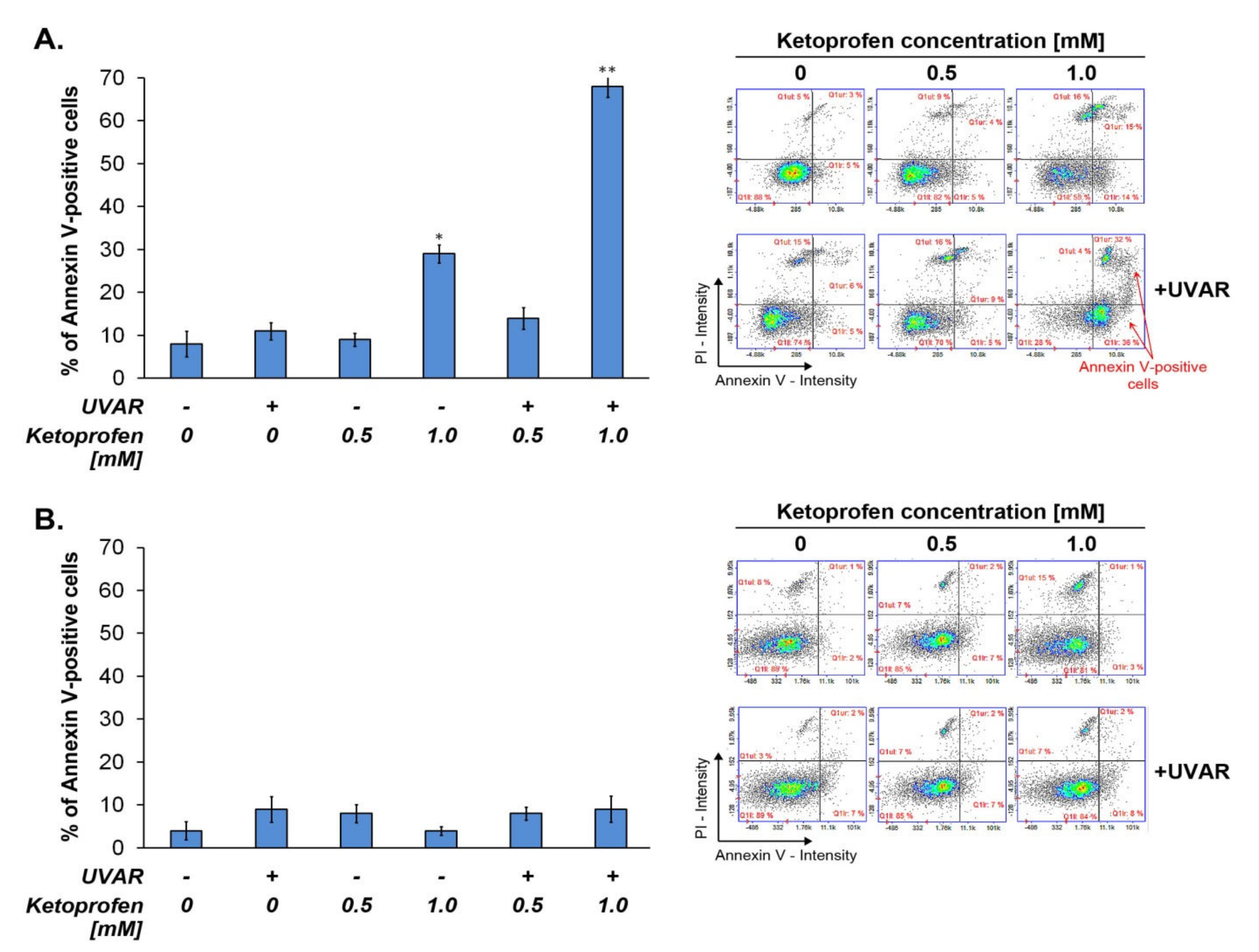
2.2. Co-Treatment with KTP and UVAR Induces Apoptosis in Melanoma Cells, but Not in Human Melanocytes
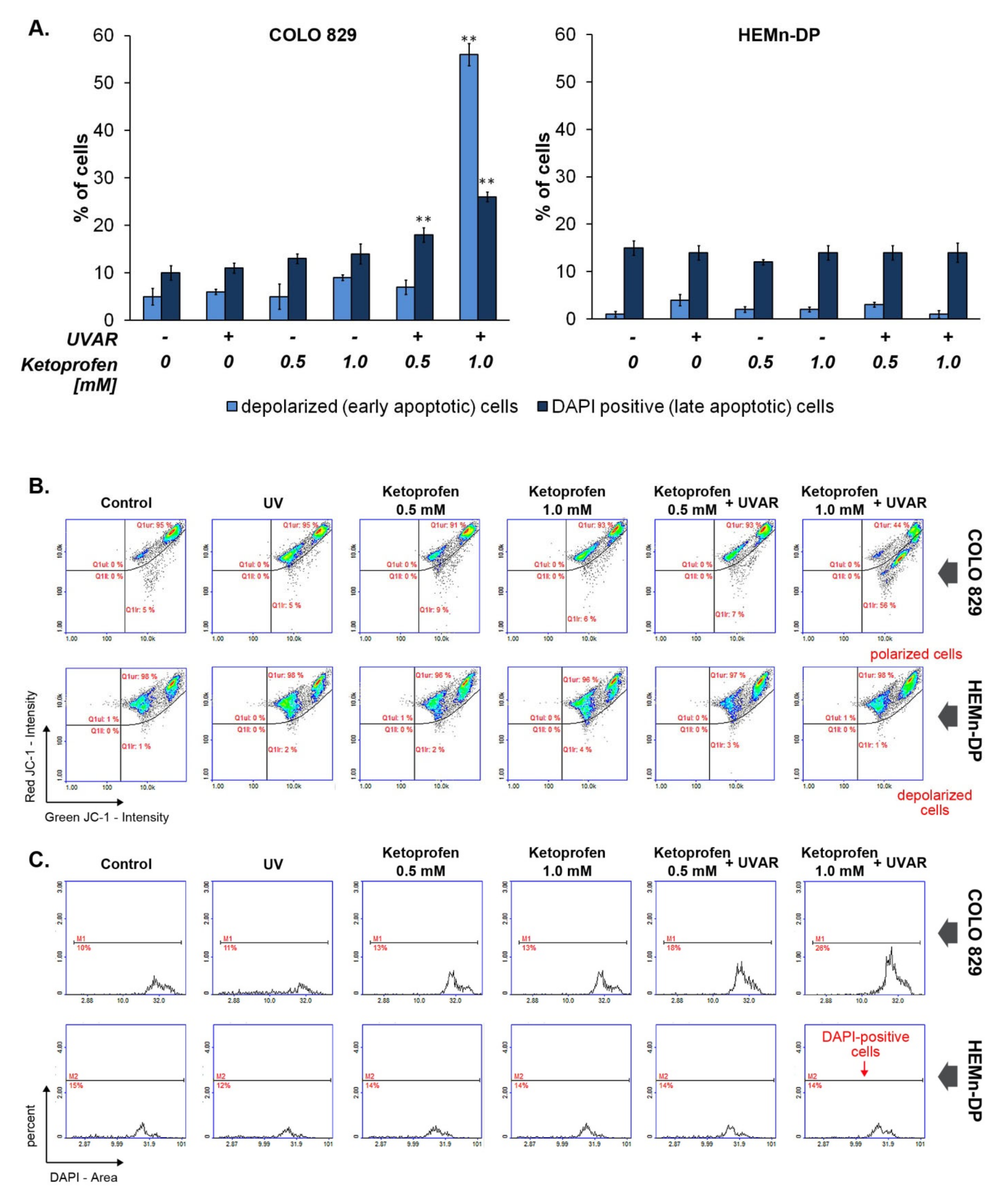
2.3. KTP in Combination with UVAR Decreases Mitochondrial Membrane Potential of Melanoma Cells
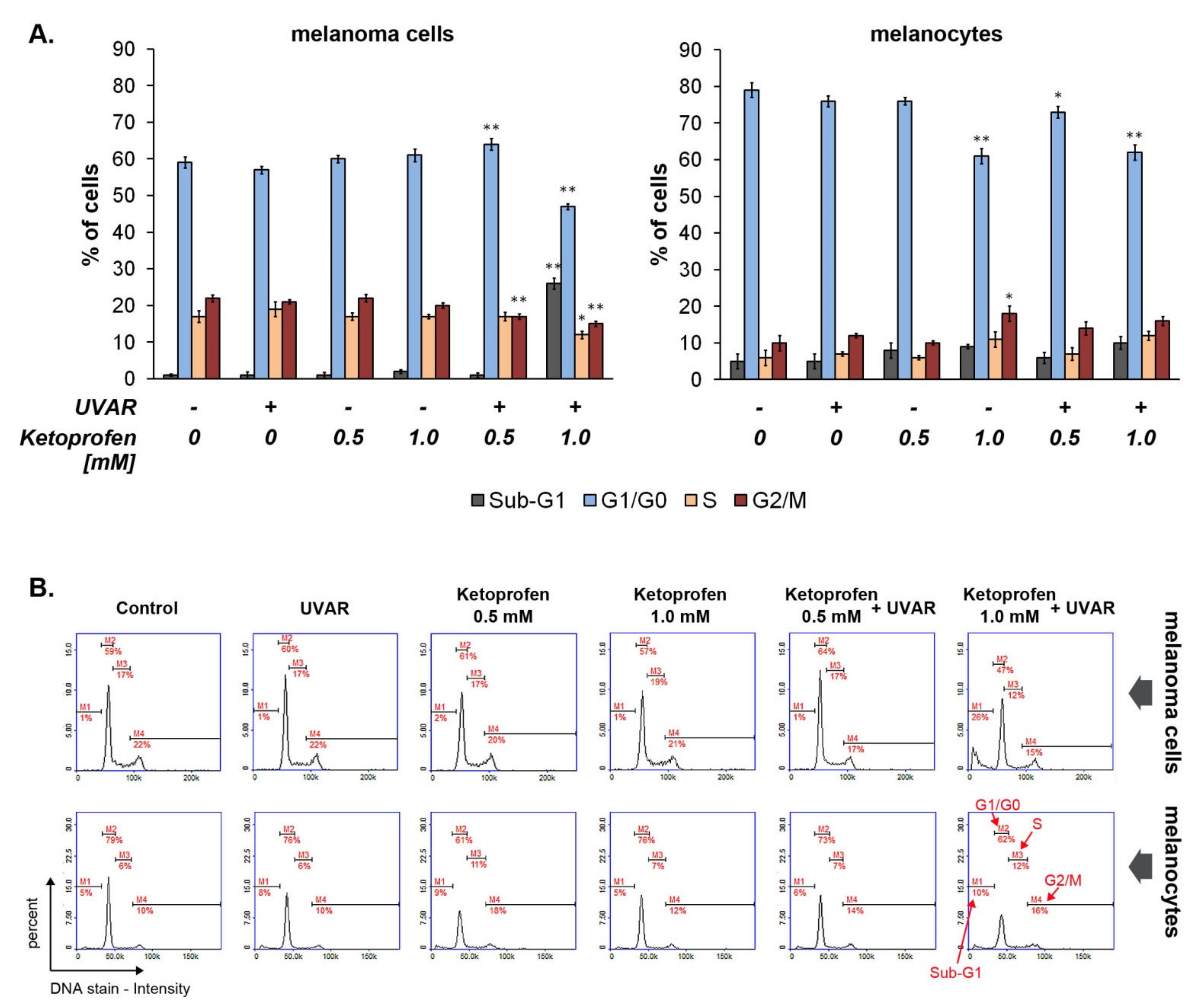
2.4. The Effect of KTP and UVAR on Melanoma and Melanocytes Cycle
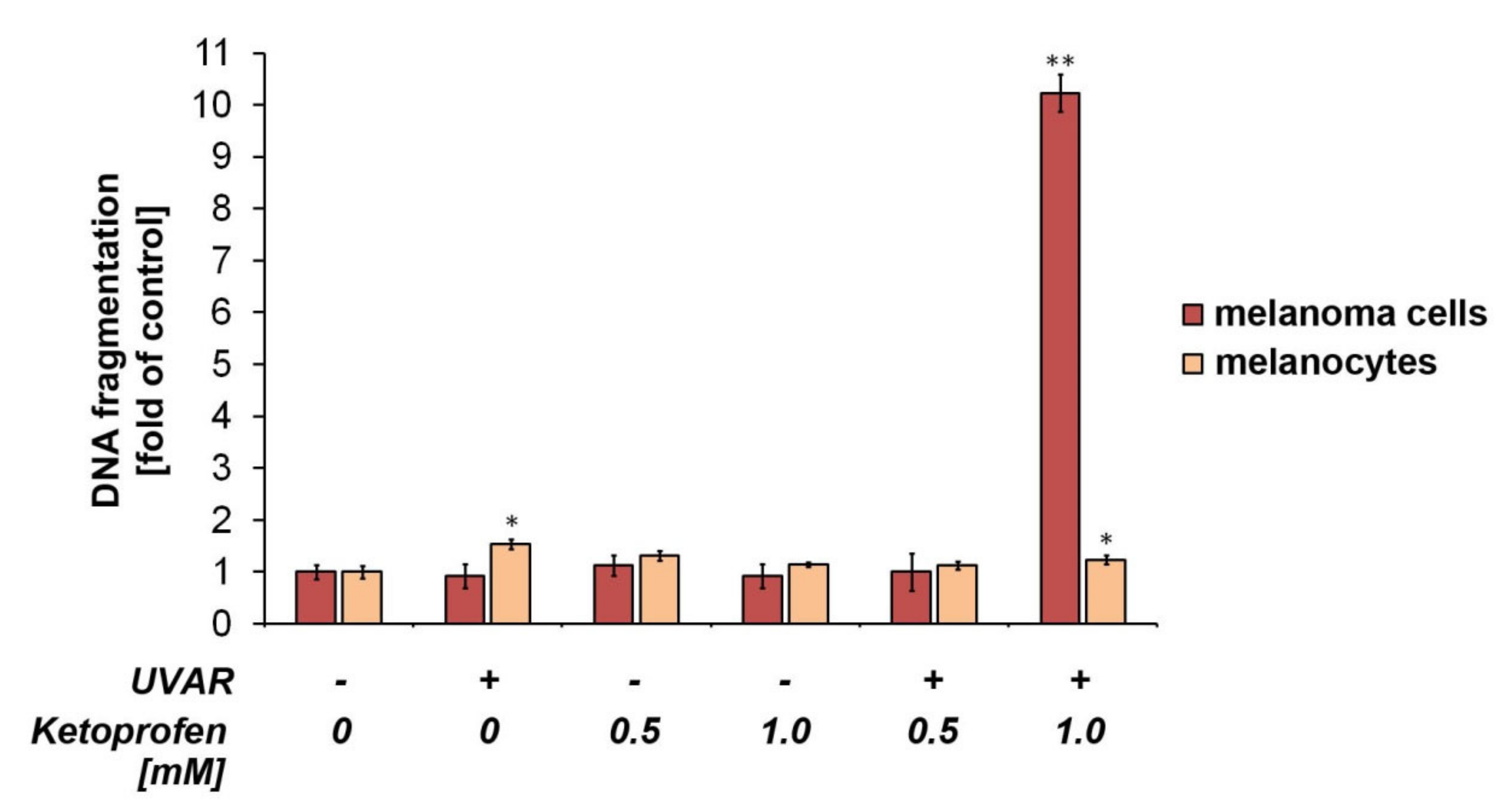
2.5. The Influence of KTP and UVA Irradiation on DNA Fragmentation in Melanoma Cells and Melanocytes
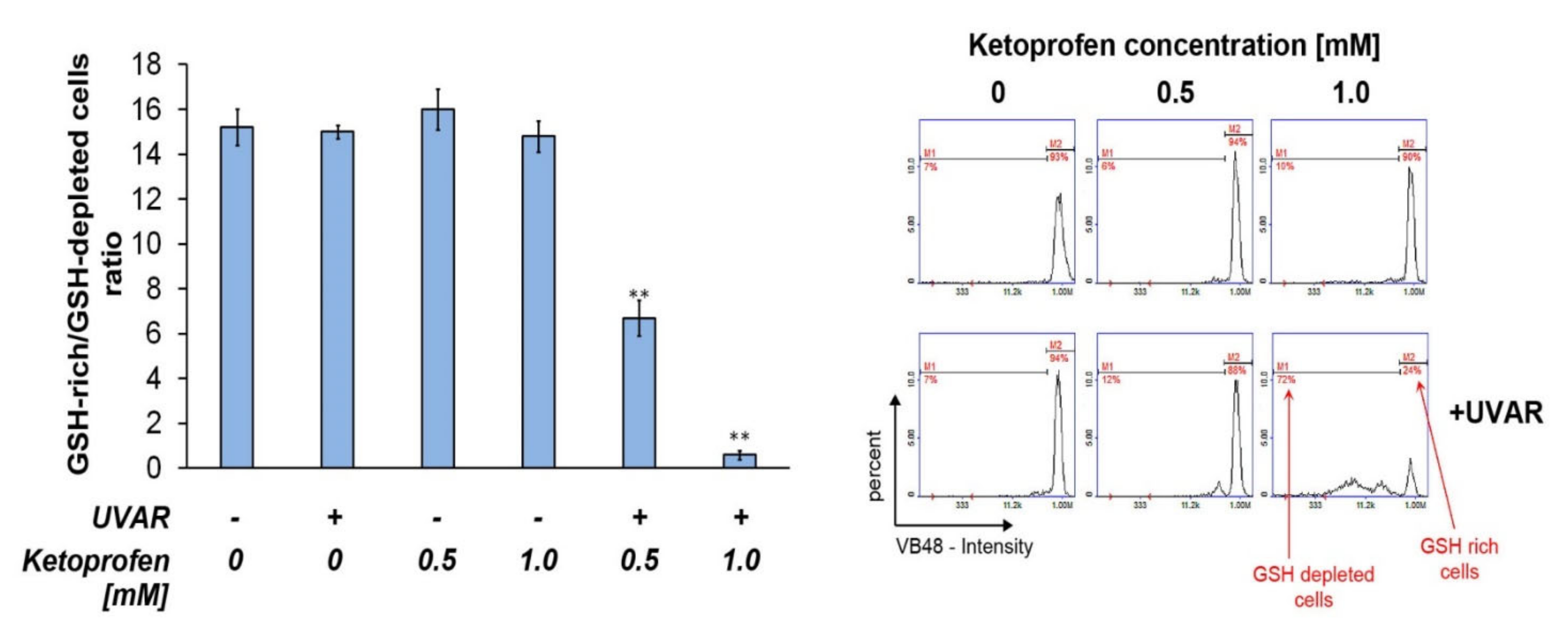
2.6. Co-Therapy with KTP and UVAR Decreases the Intracellular Level of Reduced Thiols
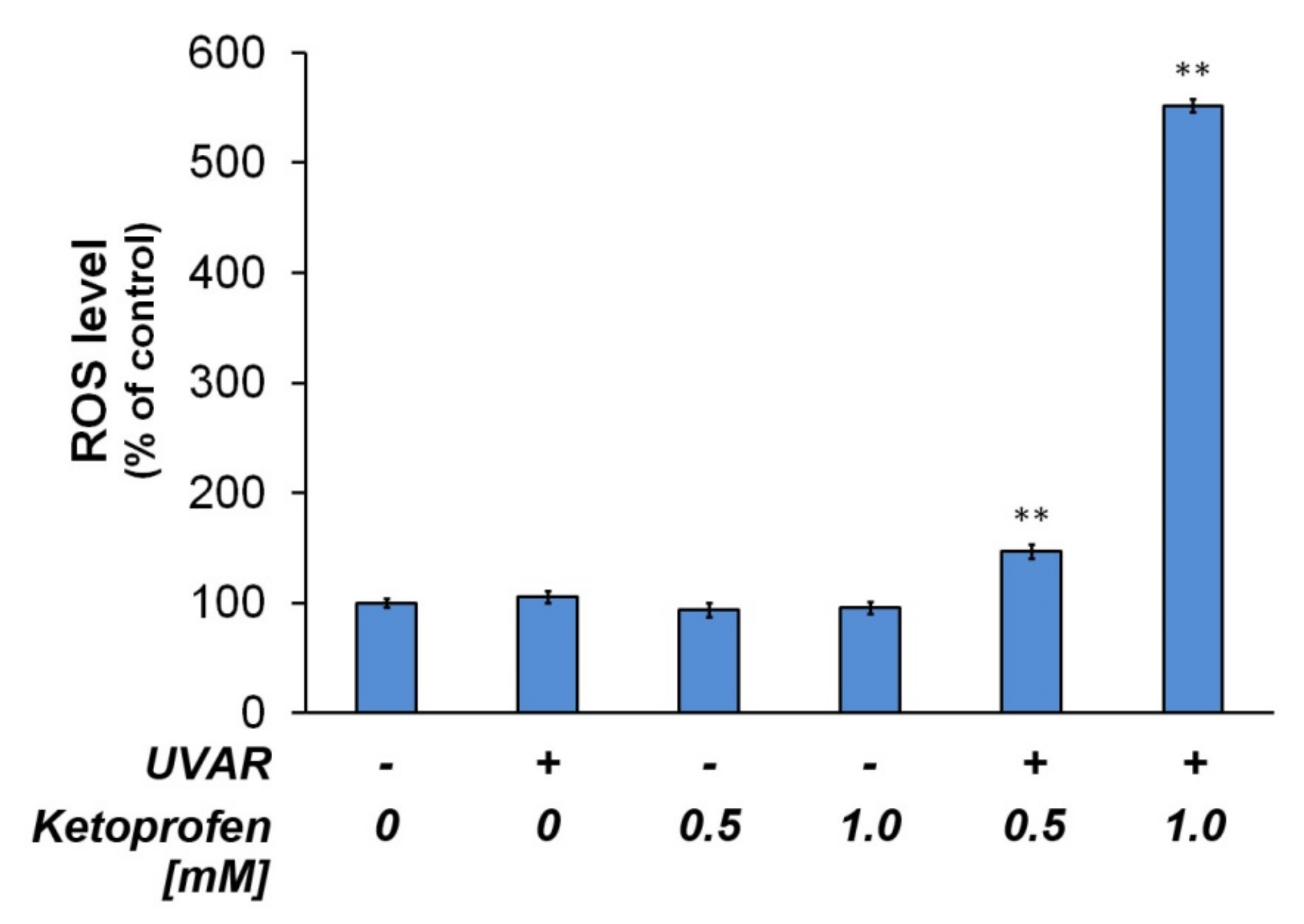
2.7. Redox Homeostasis of Melanotic Melanoma Exposed to KTP and UVAR
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Lines and Cell Culture
4.3. Cells Treatment and Counting Assay
4.4. Detection of Phosphatidylserine Externalization in Apoptotic Cells
4.5. Detection of Mitochondrial Depolarization
4.6. Cell Cycle Analysis and DNA Fragmentation Assay
4.7. Assessment of Intracellular GSH Levels
4.8. ROS Detection Assay—H2DCFDA
4.9. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 5-MOP | 5-methoxypsoralen |
| 8-MOP | 8-methoxypsoralen |
| AO | acridine orange |
| COLO829 | metastatic melanotic melanoma cell line |
| DMSO | dimethyl sulfoxide |
| GSH | reduced glutathione |
| HEMn-DP | human epidermal melanocytes, neonatal, darkly pigmented |
| HMGS-2 | human melanocyte growth supplement-2 |
| KTP | ketoprofen |
| PBS | phosphate buffered saline |
| PI | propidium iodide |
| PS | phosphatidylserine |
| PUVA | Psoralen Ultra-Violet A |
| ROS | reactive oxygen species |
| UVAR | ultraviolet A radiation |
References
- Devi, S.; Sinha, R.; Singh, R.K. Malignant melanoma maxilla. Natl. J. Maxillofac. Surg. 2015, 6, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Longo, C.; Pellacani, G. Melanomas. Dermatol. Clin. 2016, 34, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Moran, B.; Silva, R.; Perry, A.; Gallagher, W.M. Epigenetics of malignant melanoma. Semin. Cancer Biol. 2018, 51, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Respondek, M.; Beberok, A.; Rzepka, Z.; Rok, J.; Wrześniok, D. Mcl-1 Inhibitor Induces Cells Death in BRAF-Mutant Amelanotic Melanoma Trough GSH Depletion, DNA Damage and Cell Cycle Changes. Pathol. Oncol. Res. 2019, 26, 1465–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Respondek, M.; Beberok, A.; Rzepka, Z.; Rok, J.; Wrześniok, D. MIM1 induces COLO829 melanoma cell death through mitochondrial membrane breakdown, GSH depletion, and DNA damage. Fundam. Clin. Pharmacol. 2020, 34, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Lo, J.A.; Fisher, D.E. The melanoma revolution: From UV carcinogenesis to a new era in therapeutics. Science 2014, 346, 945–949. [Google Scholar] [CrossRef] [Green Version]
- Ho, A.; Tsao, H. Targeted Therapies in Melanoma: Translational Research at Its Finest. J. Investig. Dermatol. 2015, 135, 1929–1933. [Google Scholar] [CrossRef] [Green Version]
- Andreazza, N.L.; Vevert-Bizet, C.; Bourg-Heckly, G.; Sureau, F.; Salvador, M.; Bonneau, S. Berberine as a photosensitizing agent for antitumoral photodynamic therapy: Insights into its association to low density lipoproteins. Int. J. Pharm. 2016, 510, 240–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rok, J.; Karkoszka, M.; Rzepka, Z.; Respondek, M.; Banach, K.; Beberok, A.; Wrześniok, D. Cytotoxic and proapoptotic effect of doxycycline—An in vitro study on the human skin melanoma cells. Toxicol. Vitr. 2020, 65, 104790. [Google Scholar] [CrossRef]
- Obrador, E.; Liu-Smith, F.; Dellinger, R.W.; Salvador, R.; Meyskens, F.L.; Estrela, J.M. Oxidative stress and antioxidants in the pathophysiology of malignant melanoma. Biol. Chem. 2019, 400, 589–612. [Google Scholar] [CrossRef] [Green Version]
- Ferrera, L.; Barbieri, R.; Picco, C.; Zuccolini, P.; Remigante, A.; Bertelli, S.; Fumagalli, M.; Zifarelli, G.; La Porta, C.; Gavazzo, P.; et al. TRPM2 Oxidation Activates Two Distinct Potassium Channels in Melanoma Cells through Intracellular Calcium Increase. Int. J. Mol. Sci. 2021, 22, 8359. [Google Scholar] [CrossRef] [PubMed]
- Obrador, E.; Salvador, R.; López-Blanch, R.; Jihad-Jebbar, A.; Alcácer, J.; Benlloch, M.; Pellicer, J.A.; Estrela, J.M. Melanoma in the liver: Oxidative stress and the mechanisms of metastatic cell survival. Semin. Cancer Biol. 2021, 71, 109–121. [Google Scholar] [CrossRef]
- Klimas, K.; Fischer, A. Phototoxic reaction and factors causing it. Publ. Health Forum 2018, 4, 83–88. [Google Scholar]
- Kim, J.H.; Kim, H.J. Fast and simple method for screening of single-stranded DNA breaking photosensitizers using graphene oxide. Nano Converg. 2018, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Latosinski, G.; Amzalak, M.; Pantoja, J. Efficacy of ketoprofen for treatment of spontaneous, culture-negative, mild cases of clinical mastitis: A randomized, controlled superiority trial. J. Dairy Sci. 2020, 103, 2624–2635. [Google Scholar] [CrossRef]
- Paunescu, A.; Ponepal, C.M.; Zagardan, M.C.; Plesa, C.F.; Nemes, R.M.; Nicolae, C.; Bisoc, A.; Diaconu, M.; Fierascu, I.; Fierascu, R.C.; et al. Evaluation of histophysiological alterations associated with ketoprofen administration in albino NMRI mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1033–1039. [Google Scholar] [CrossRef]
- Bignon, E.; Marazzi, M.; Besancenot, V.; Gattuso, H.; Drouot, G.; Morell, C.; Eriksson, L.A.; Grandemange, S.; Dumont, E.; Monari, A. Ibuprofen and ketoprofen potentiate UVA-induced cell death by a photosensitization process. Sci. Rep. 2017, 7, 8885. [Google Scholar] [CrossRef] [Green Version]
- Banach, K.; Kowalska, J.; Rzepka, Z.; Beberok, A.; Rok, J.; Wrześniok, D. The role of UVA radiation in ketoprofen-mediated BRAF-mutant amelanotic melanoma cells death—A study at the cellular and molecular level. Toxicol. Vitr. 2021, 72, 105108. [Google Scholar] [CrossRef] [PubMed]
- Miliński, M.; Delijewski, M.; Rok, J.; Wrześniok, D.; Beberok, A.; Chełmecka, E.; Buszman, E. The pivotal effect of sulindac on melanin-containing cancer cells. Acta Pol. Pharm. 2017, 74, 1681–1689. [Google Scholar]
- Ibbotson, S.H. A Perspective on the Use of NB-UVB Phototherapy vs. PUVA Photochemotherapy. Front. Med. 2018, 5, 184. [Google Scholar] [CrossRef] [Green Version]
- Farahnik, B.; Nakamura, M.; Singh, R.K.; Abrouk, M.; Zhu, T.H.; Lee, K.M.; Jose, M.V.; Dalovisio, R.; Koo, J.; Bhutani, T.; et al. The Patient’s Guide to Psoriasis Treatment. Part 2: PUVA Phototherapy. Dermatol. Ther. 2016, 6, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juarez, M.C.; Grossberg, A.L. Phototherapy in the Pediatric Population. Dermatol. Clin. 2020, 38, 91–108. [Google Scholar] [CrossRef]
- Da Veiga, G.P.; Pérez-Feal, P.; Moreiras-Arias, N.; Peteiro-García, C.; Suárez-Peñaranda, J.M.; Vázquez-Veiga, H.; Rodríguez-Granados, M.T. Treatment of nail lichen planus with localized bath-PUVA. Photodermatol. Photoimmunol. Photomed. 2019, 36, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.S.; Elmariah, S.B. Phototherapy for Itch. Dermatol. Clin. 2020, 38, 145–155. [Google Scholar] [CrossRef]
- Wrześniok, D.; Beberok, A.; Rok, J.; Delijewski, M.; Hechmann, A.; Oprzondek, M.; Rzepka, Z.; Bacler-Żbikowska, B.; Buszman, E. UVA radiation augments cytotoxic activity of psoralens in melanoma cells. Int. J. Radiat. Biol. 2017, 93, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Shin, J.-W.; Pham, T.-H.; Choi, Y.-J.; Ryu, H.-W.; Oh, S.-R.; Oh, J.-W.; Yoon, D.-Y. Methyl lucidone induces apoptosis and G2/M phase arrest via the PI3K/Akt/NF-κB pathway in ovarian cancer cells. Pharm. Biol. 2019, 58, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.-H.; Ko, H.-M.; Han, G.-D.; Lee, S.-Y.; Moon, J.-S.; Kim, M.-S.; Koh, J.-T.; Kim, S.-H. Dual role of phosphatidylserine and its receptors in osteoclastogenesis. Cell Death Dis. 2020, 11, 497. [Google Scholar] [CrossRef]
- Rok, J.; Buszman, E.; Delijewski, M.; Otręba, M.; Beberok, A.; Wrześniok, D. Effect of tetracycline and UV radiation on melanization and antioxidant status of melanocytes. J. Photochem. Photobiol. B: Biol. 2015, 148, 168–173. [Google Scholar] [CrossRef]
- Rok, J.; Wrześniok, D.; Beberok, A.; Otręba, M.; Delijewski, M.; Buszman, E. Phototoxic effect of oxytetracycline on normal human melanocytes. Toxicol. In Vitro 2018, 48, 26–32. [Google Scholar] [CrossRef]
- Xu, W.-T.; Li, T.-Z.; Li, S.-M.; Wang, C.; Wang, H.; Luo, Y.-H.; Piao, X.-J.; Wang, J.-R.; Zhang, Y.; Zhang, T.; et al. Cytisine exerts anti-tumour effects on lung cancer cells by modulating reactive oxygen species-mediated signalling pathways. Artif. Cells Nanomed. Biotechnol. 2019, 48, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Barnum, K.J.; O’Connell, M.J. Cell Cycle Regulation by Checkpoints. Methods Mol. Biol. 2014, 1170, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Jingwen, B.; Yaochen, L.; Guojun, Z. Cell cycle regulation and anticancer drug discovery. Cancer Biol. Med. 2017, 14, 348–362. [Google Scholar] [CrossRef]
- Diaz-Moralli, S.; Tarrado-Castellarnau, M.; Miranda, A.; Cascante, M. Targeting cell cycle regulation in cancer therapy. Pharmacol. Ther. 2013, 138, 255–271. [Google Scholar] [CrossRef]
- Conklin, K.A. Chemotherapy-Associated Oxidative Stress: Impact on Chemotherapeutic Effectiveness. Integr. Cancer Ther. 2004, 3, 294–300. [Google Scholar] [CrossRef]
- Huang, X.; Halicka, H.D.; Traganos, F.; Tanaka, T.; Kurose, A.; Darzynkiewicz, Z. Cytometric assessment of DNA damage in relation to cell cycle phase and apoptosis. Cell Prolif. 2005, 38, 223–243. [Google Scholar] [CrossRef] [PubMed]
- Beberok, A.; Wrześniok, D.; Minecka, A.; Rok, J.; Delijewski, M.; Rzepka, Z.; Respondek, M.; Buszman, E. Ciprofloxacin-mediated induction of S-phase cell cycle arrest and apoptosis in COLO829 melanoma cells. Pharmacol. Rep. 2018, 70, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Guimarais, V.; Gil-Guiñon, E.; Sánchez-Osuna, M.; Casanelles, E.; García-Belinchón, M.; Comella, J.X.; Yuste, V.J. Chromatin Collapse during Caspase-dependent Apoptotic Cell Death Requires DNA Fragmentation Factor, 40-kDa Subunit-/Caspase-activated Deoxyribonuclease-mediated 3′-OH Single-strand DNA Breaks. J. Biol. Chem. 2013, 288, 9200–9215. [Google Scholar] [CrossRef] [Green Version]
- Kitazumi, I.; Tsukahara, M. Regulation of DNA fragmentation: The role of caspases and phosphorylation. FEBS J. 2011, 278, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Czarnomysy, R.; Radomska, D.; Muszyńska, A.; Hermanowicz, J.M.; Prokop, I.; Bielawska, A.; Bielawski, K. Evaluation of the Anticancer Activities of Novel Transition Metal Complexes with Berenil and Nitroimidazole. Molecules 2020, 25, 2860. [Google Scholar] [CrossRef]
- Minich, D.M.; Brown, B.I. A Review of Dietary (Phyto)Nutrients for Glutathione Support. Nutrition 2019, 11, 2073. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Zhen, C.; Liu, J.; Yang, P.; Hu, L.; Shang, P. Unraveling the Potential Role of Glutathione in Multiple Forms of Cell Death in Cancer Therapy. Oxidative Med. Cell. Longev. 2019, 2019, 3150145. [Google Scholar] [CrossRef] [Green Version]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hyde, A.S.; Simpson, M.A.; Barycki, J.J. Emerging Regulatory Paradigms in Glutathione Metabolism. Adv. Cancer Res. 2014, 122, 69–101. [Google Scholar]
- Franco, R.; Cidlowski, J.A. Apoptosis and glutathione: Beyond an antioxidant. Cell Death Differ. 2009, 16, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Harris, I.; Endress, J.E.; Coloff, J.L.; Selfors, L.; McBrayer, S.K.; Rosenbluth, J.M.; Takahashi, N.; Dhakal, S.; Koduri, V.; Oser, M.G.; et al. Deubiquitinases Maintain Protein Homeostasis and Survival of Cancer Cells upon Glutathione Depletion. Cell Metab. 2019, 29, 1166–1181.e6. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Manero, F.; Gonin, S.; Kretz-Remy, C.; Virot, S.; Arrigo, A.-P. Hsp27 as a Negative Regulator of Cytochrome c Release. Mol. Cell. Biol. 2002, 22, 816–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.R.; Trush, M.A. Defining ROS in Biology and Medicine. React. Oxyg. Species 2016, 1, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Lian, G. ROS and diseases: Role in metabolism and energy supply. Mol. Cell. Biochem. 2019, 467, 349–361. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Beberok, A.; Wrześniok, D.; Szlachta, M.; Rok, J.; Rzepka, Z.; Respondek, M.; Buszman, E. Lomefloxacin Induces Oxidative Stress and Apoptosis in COLO829 Melanoma Cells. Int. J. Mol. Sci. 2017, 18, 2194. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS Using Oxidized DCFDA and Flow-Cytometry. Methods Mol. Biol. 2010, 594, 57–72. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banach, K.; Kowalska, J.; Rzepka, Z.; Beberok, A.; Rok, J.; Wrześniok, D. Ketoprofen Combined with UVA Irradiation Exerts Higher Selectivity in the Mode of Action against Melanotic Melanoma Cells than against Normal Human Melanocytes. Int. J. Mol. Sci. 2021, 22, 11966. https://doi.org/10.3390/ijms222111966
Banach K, Kowalska J, Rzepka Z, Beberok A, Rok J, Wrześniok D. Ketoprofen Combined with UVA Irradiation Exerts Higher Selectivity in the Mode of Action against Melanotic Melanoma Cells than against Normal Human Melanocytes. International Journal of Molecular Sciences. 2021; 22(21):11966. https://doi.org/10.3390/ijms222111966
Chicago/Turabian StyleBanach, Klaudia, Justyna Kowalska, Zuzanna Rzepka, Artur Beberok, Jakub Rok, and Dorota Wrześniok. 2021. "Ketoprofen Combined with UVA Irradiation Exerts Higher Selectivity in the Mode of Action against Melanotic Melanoma Cells than against Normal Human Melanocytes" International Journal of Molecular Sciences 22, no. 21: 11966. https://doi.org/10.3390/ijms222111966