
Suppression of Estrogen Receptor Alpha Inhibits Cell Proliferation, Differentiation and Enhances the Chemosensitivity of P53-Positive U2OS Osteosarcoma Cell

 , ,
, ,
Abstract
:1. Introduction
2. Results
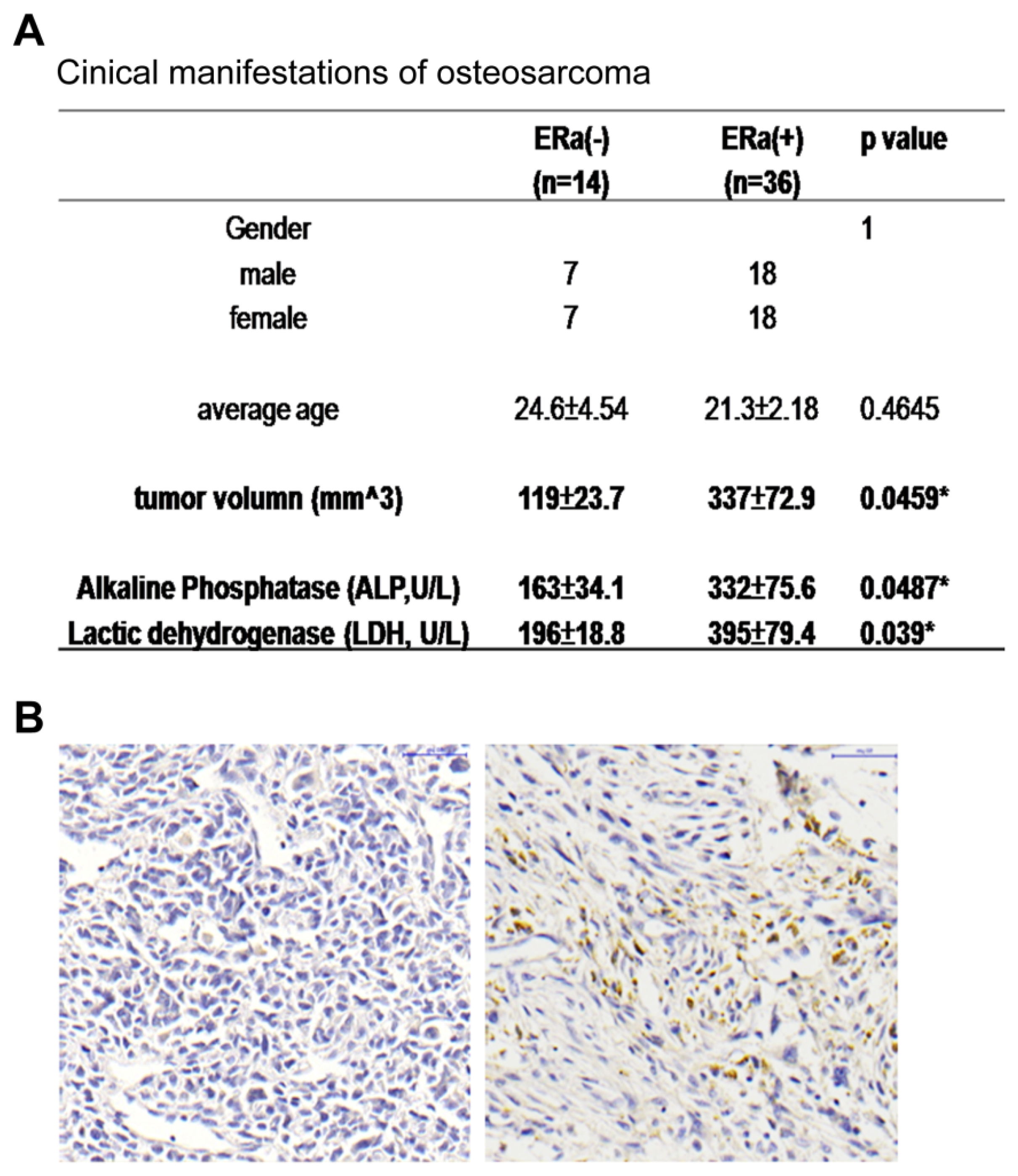
2.1. ERα Positive Expression Pattern in OS Patients Was Correlated with Increased Tumor Size and ALP and LDH Levels
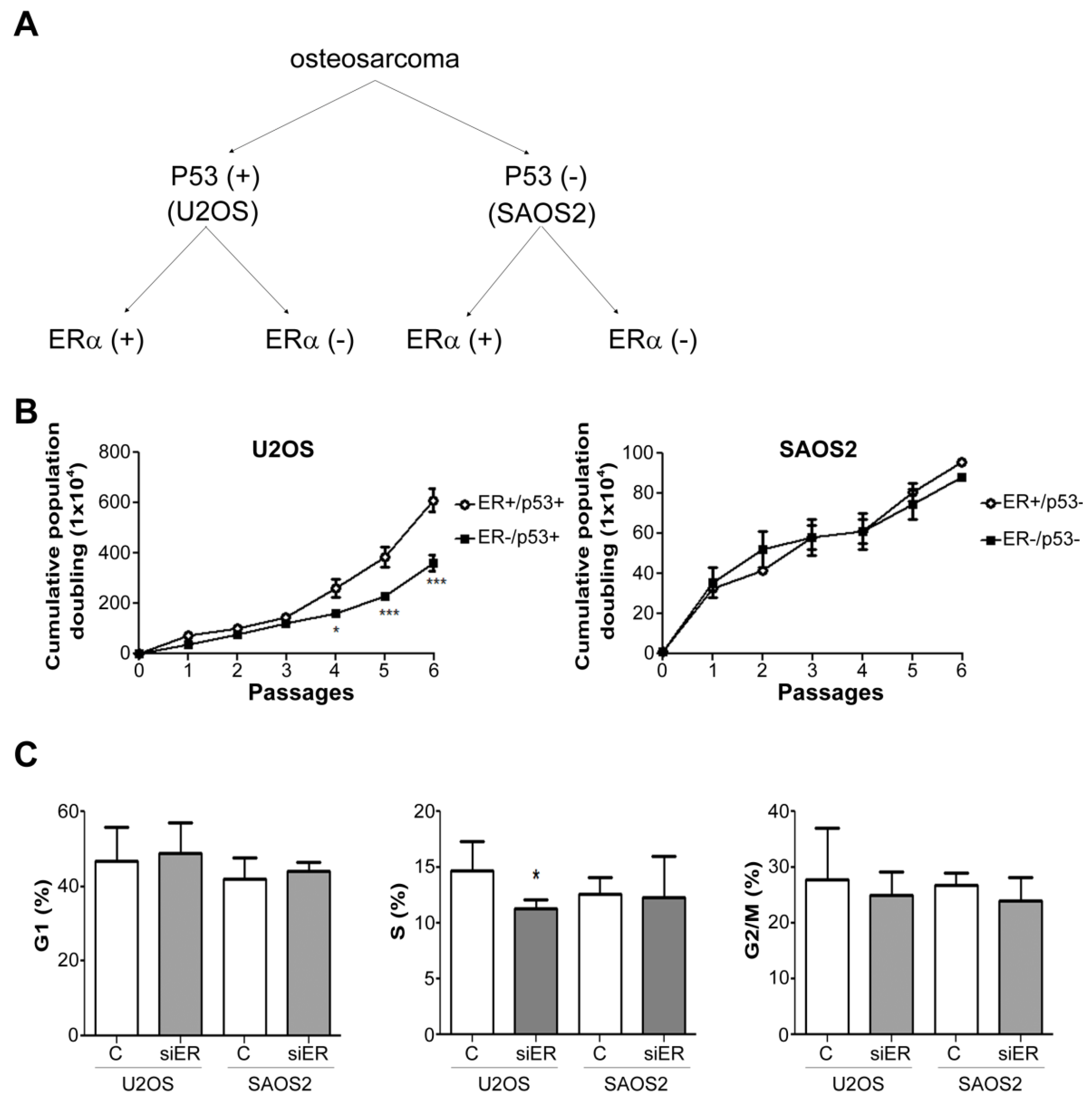
2.2. ERα Knock down Suppressed the Growth Rate of P53-Positive U2OS Cells but Not of P53-Negative SAOS2 Cells
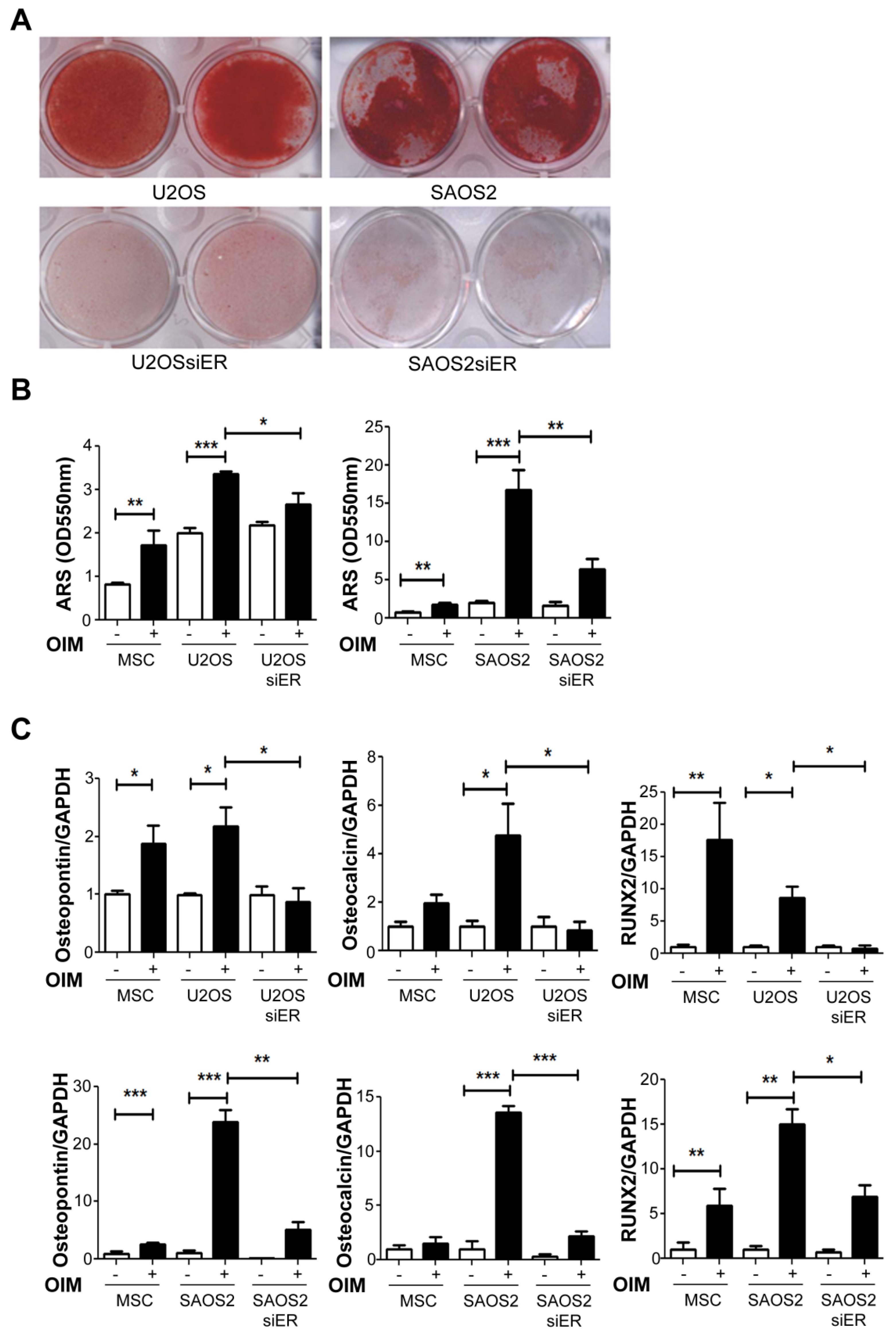
2.3. Knock down of ERα Suppressed the Osteogenesis Ability in Both P53+ U2OS and P53− SAOS2Cells
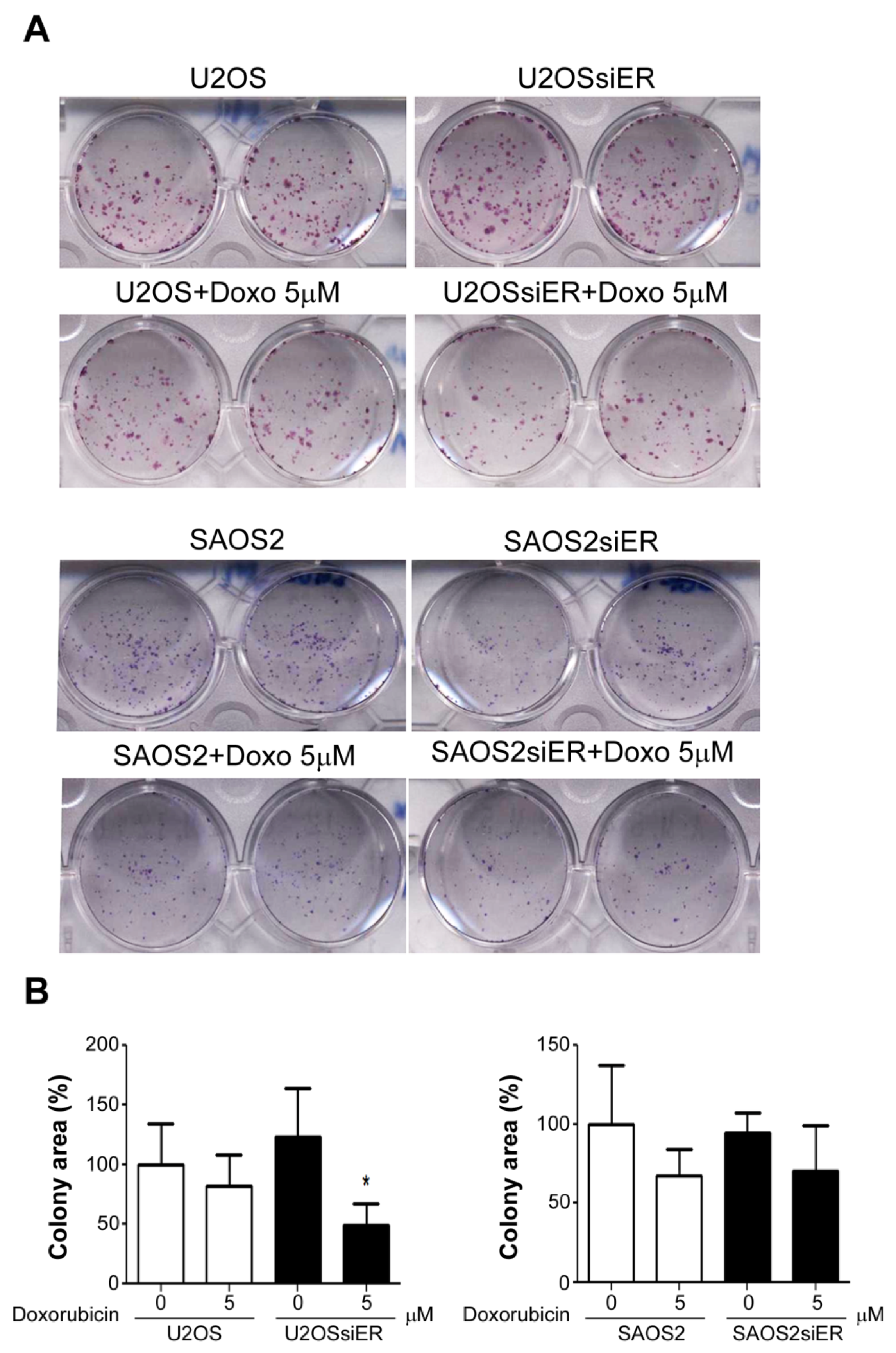
2.4. Silencing of ERα in P53-Positive U2OS Cells Suppressed Colony Formation Ability after Combined Treatment with Doxorubicin
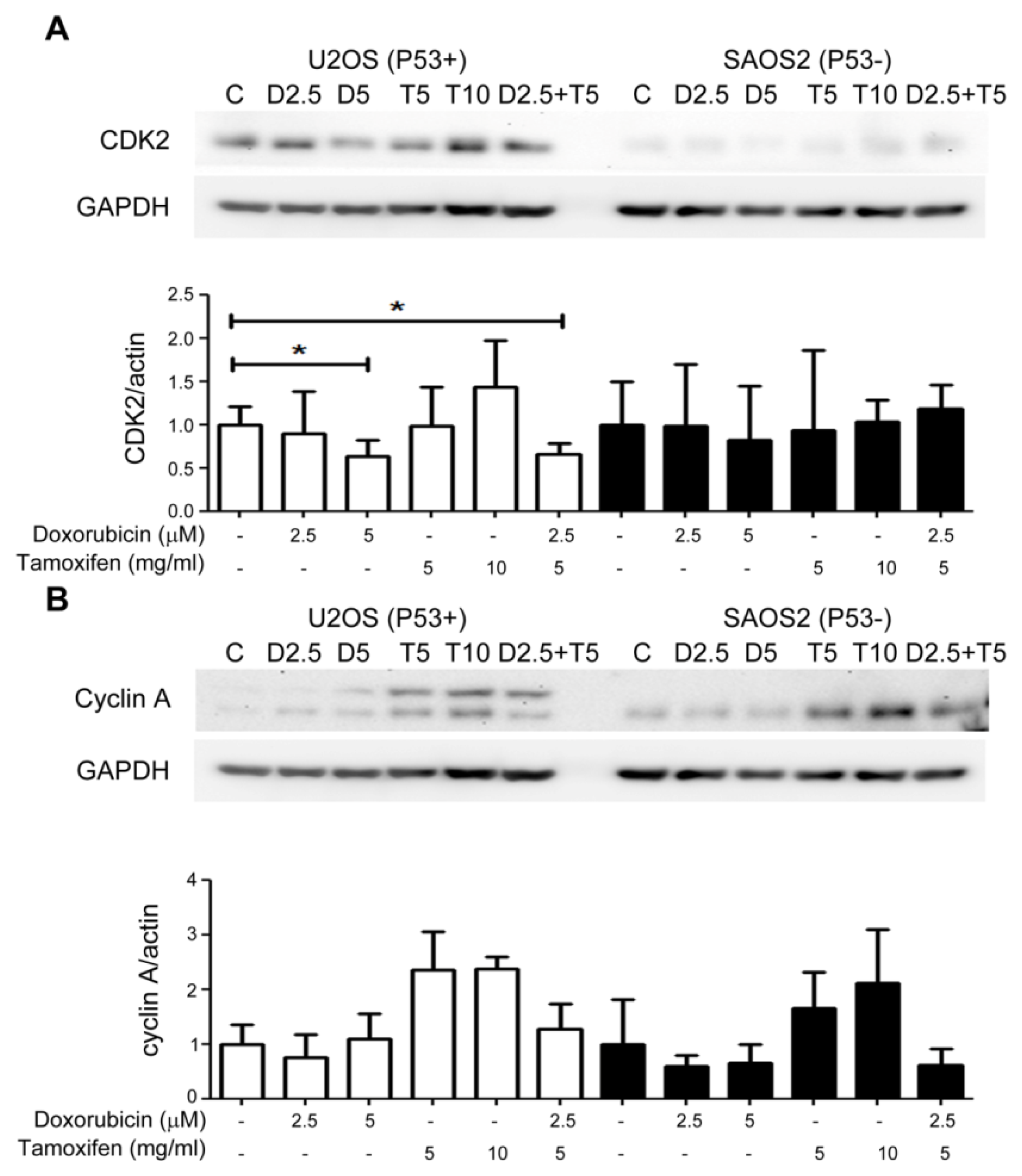
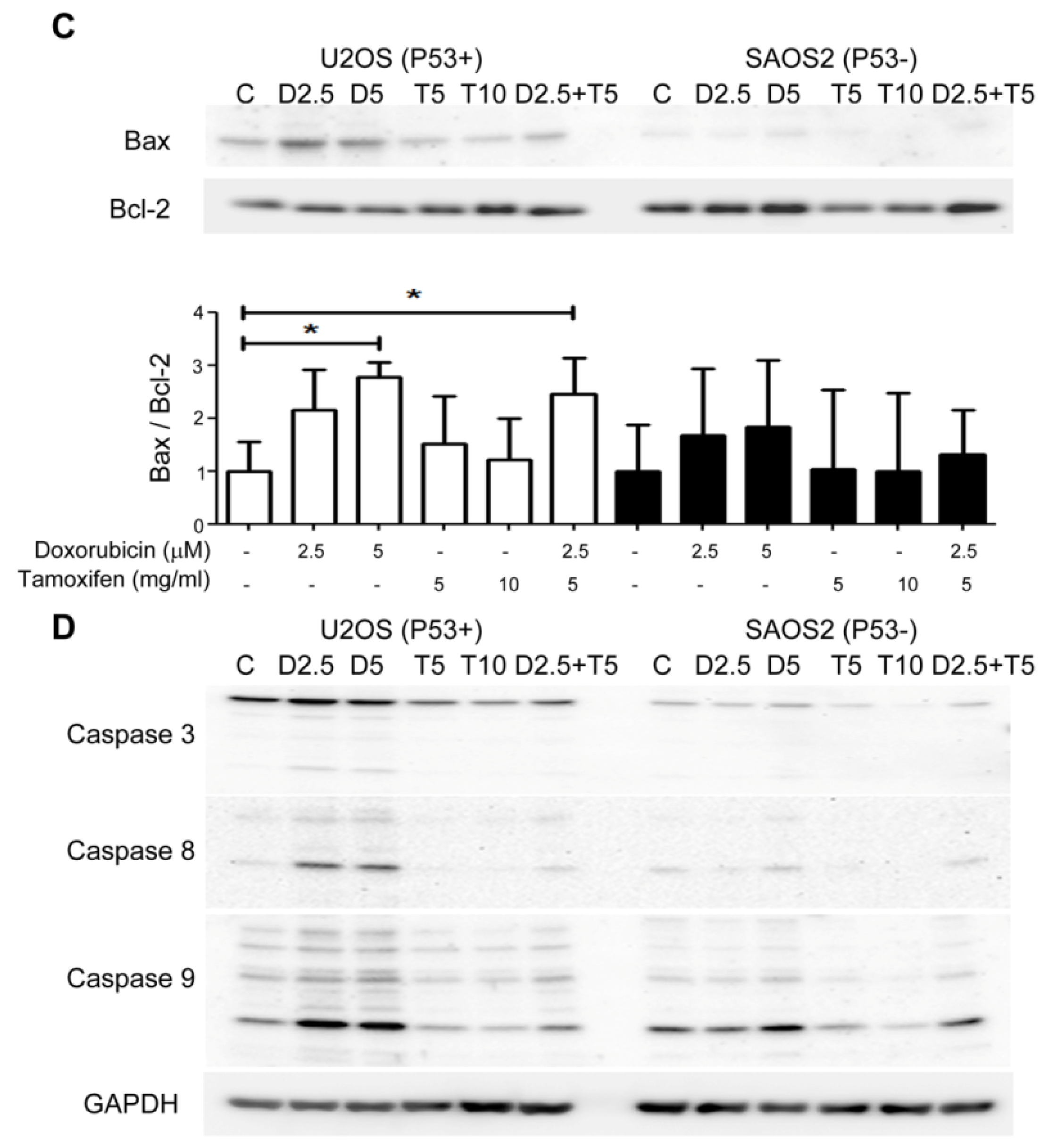
2.5. Combined Treatment with Tamoxifen Enhanced the Growth Inhibition Effects of Doxorubicin on P53(+) U2OS Cell by Suppressing CDK2 and Cyclin A and Inducing Apoptosis
2.6. ERα Low Expression Patterns in P53-Positive OS Patients Were Associated with Better Responses to Chemotherapy and Smaller Tumor Sizes
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
4.2. Transfection and Lentiviral-Mediated Transduction
4.3. Calculation of Cell Growth
4.4. Flow Cytometry for Cell Cycle Analysis
4.5. Colony Formation Assay
4.6. Osteogenesis Induction and Alizarin Red S Staining
4.7. Immunohistochemistry on Tumor Tissue Array
4.8. Western Blotting
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritter, J.; Bielack, S.S. Osteosarcoma. Ann. Oncol. 2010, 21, vii320–vii325. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Chen, J.; Song, W.X.; Tang, N.; Luo, J.; Deng, Z.L.; Sharff, K.A.; He, G.; Bi, Y.; He, B.C.; et al. Osteogenic BMPs promote tumor growth of human osteosarcomas that harbor differentiation defects. Lab. Investig. 2008, 88, 1264–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.W.; Squire, J.A.; Zielenska, M. The genetics of osteosarcoma. Sarcoma 2012, 2012, 627254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.Y.; Wu, P.K.; Chen, P.C.; Lee, C.W.; Chen, W.M.; Hung, S.C. Generation of Osteosarcomas from a Combination of Rb Silencing and c-Myc Overexpression in Human Mesenchymal Stem Cells. Stem Cells Transl. Med. 2017, 6, 512–526. [Google Scholar] [CrossRef] [Green Version]
- Mirabello, L.J.; Yeager, M.; Mai, P.L.; Gastier-Foster, J.; Gorlick, R.; Khanna, C.; Patino-Garcia, A.; Sierrasesumaga, L.; Lecanda, F.; Andrulis, I.L.; et al. High prevalence of germline TP53 mutations in young osteosarcoma cases. Cancer Res. 2015, 75, 5574. [Google Scholar] [CrossRef]
- Mirabello, L.; Yeager, M.; Mai, P.L.; Gastier-Foster, J.M.; Gorlick, R.; Khanna, C.; Patino-Garcia, A.; Sierrasesumaga, L.; Lecanda, F.; Andrulis, I.L.; et al. Germline TP53 Variants and Susceptibility to Osteosarcoma. JNCI-J. Natl. Cancer Inst. 2015, 107, djv101. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.W.; Aslo, A.; Won, A.; Tan, M.; Lampkin, B.; Koeffler, H.P. Alterations of the p53, Rb and MDM2 genes in osteosarcoma. J. Cancer Res.Clin. 1996, 122, 559–565. [Google Scholar] [CrossRef]
- Schreck, R.R. Tumor Suppressor Gene (Rb and P53) Mutations in Osteosarcoma. Pediatr. Hemat. Oncol. 1992, 9, R9–R10. [Google Scholar] [CrossRef] [PubMed]
- Piperdi, S.; Zhang, W.D.; Ivenitsky, D.; Zhang, Y.D.; Zhang, Y.J.; Geller, D.S.; Hoang, B.; Yang, R.; Gill, J.B.; Roth, M.; et al. Introducing c-Myc into transformed human mesenchymal stem cells and osteoblasts to recapitulate the osteosarcoma phenotype. Cancer Res. 2017, 77. [Google Scholar] [CrossRef]
- Shimizu, T.; Ishikawa, T.; Sugihara, E.; Kuninaka, S.; Miyamoto, T.; Mabuchi, Y.; Matsuzaki, Y.; Tsunoda, T.; Miya, F.; Morioka, H.; et al. c-MYC overexpression with loss of Ink4a/Arf transforms bone marrow stromal cells into osteosarcoma accompanied by loss of adipogenesis. Oncogene 2010, 29, 5687–5699. [Google Scholar] [CrossRef] [Green Version]
- Rickel, K.; Fang, F.; Tao, J. Molecular genetics of osteosarcoma. Bone 2017, 102, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Mohseny, A.B.; Szuhai, K.; Romeo, S.; Buddingh, E.P.; Briaire-de Bruijn, I.; de Jong, D.; van Pel, M.; Cleton-Jansen, A.M.; Hogendoorn, P.C. Osteosarcoma originates from mesenchymal stem cells in consequence of aneuploidization and genomic loss of Cdkn2. J. Pathol. 2009, 219, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Guijarro, M.V. Osteosarcoma: Mouse models, cell of origin and cancer stem cell. Postdoc. J. 2014, 2, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, P.; Lin, C.Y. Characterization of stem cell attributes in human osteosarcoma cell lines. Cancer Biol. Ther. 2009, 8, 543–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, N.; Song, W.X.; Luo, J.; Haydon, R.C.; He, T.C. Osteosarcoma development and stem cell differentiation. Clin. Orthop. Relat.Res. 2008, 466, 2114–2130. [Google Scholar] [CrossRef] [Green Version]
- Mutsaers, A.J.; Walkley, C.R. Cells of origin in osteosarcoma: Mesenchymal stem cells or osteoblast committed cells? Bone 2014, 62, 56–63. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.R.; Luther, G.; Zhu, G.; Luo, Q.; Shi, Q.; Kim, S.H.; Gao, J.L.; Huang, E.; Gao, Y.; Yang, K.; et al. Defective osteogenic differentiation in the development of osteosarcoma. Sarcoma 2011, 2011, 325238. [Google Scholar] [CrossRef]
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An update on anticancer molecular action, toxicity and novel drug delivery systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef]
- Tang, Q.X.; Wang, L.C.; Wang, Y.; Gao, H.D.; Hou, Z.L. Efficacy of methotrexate, doxorubicin, and cisplatin for osteosarcoma: Study protocol for a systematic review of randomized controlled trial. Medicine 2019, 98, e14442. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, Y.S.; Kim, D.K. Doxorubicin exerts cytotoxic effects through cell cycle arrest and Fas-mediated cell death. Pharmacology 2009, 84, 300–309. [Google Scholar] [CrossRef]
- Bar-On, O.; Shapira, M.; Hershko, D.D. Differential effects of doxorubicin treatment on cell cycle arrest and Skp2 expression in breast cancer cells. Anticancer Drugs 2007, 18, 1113–1121. [Google Scholar] [CrossRef]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [Green Version]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin pathways: Pharmacodynamics and adverse effects. Pharm. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Bayat Mokhtari, R.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa, P.; Mendes, R.; Cabral, R.; Martins, L.; Baptista, P.V.; Fernandes, A.R. Combination of chemotherapy and Au-nanoparticle photothermy in the visible light to tackle doxorubicin resistance in cancer cells. Sci. Rep. 2018, 8, 11429. [Google Scholar] [CrossRef] [Green Version]
- Buondonno, I.; Gazzano, E.; Jean, S.R.; Audrito, V.; Kopecka, J.; Fanelli, M.; Salaroglio, I.C.; Costamagna, C.; Roato, I.; Mungo, E.; et al. Mitochondria-Targeted Doxorubicin: A New Therapeutic Strategy against Doxorubicin-Resistant Osteosarcoma. Mol. Cancer Ther. 2016, 15, 2640–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, A.S.; Sable, R.B.; Kothari, R.M. An update on transforming growth factor-beta (TGF-beta): Sources, types, functions and clinical applicability for cartilage/bone healing. J. Cell Physiol. 2011, 226, 3094–3103. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Komori, T.; Suda, T. Regulation of osteoblast differentiation mediated by bone morphogenetic proteins, hedgehogs, and Cbfa1. Endocr. Rev. 2000, 21, 393–411. [Google Scholar] [CrossRef]
- Windahl, S.H.; Saxon, L.; Borjesson, A.E.; Lagerquist, M.K.; Frenkel, B.; Henning, P.; Lerner, U.H.; Galea, G.L.; Meakin, L.B.; Engdahl, C.; et al. Estrogen receptor-alpha is required for the osteogenic response to mechanical loading in a ligand-independent manner involving its activation function 1 but not 2. J. Bone Miner. Res. 2013, 28, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Auld, K.L.; Berasi, S.P.; Liu, Y.; Cain, M.; Zhang, Y.; Huard, C.; Fukayama, S.; Zhang, J.; Choe, S.; Zhong, W.; et al. Estrogen-related receptor alpha regulates osteoblast differentiation via Wnt/beta-catenin signaling. J. Mol. Endocrinol. 2012, 48, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Kallio, A.; Guo, T.; Lamminen, E.; Seppanen, J.; Kangas, L.; Vaananen, H.K.; Harkonen, P. Estrogen and the selective estrogen receptor modulator (SERM) protection against cell death in estrogen receptor alpha and beta expressing U2OS cells. Mol. Cell Endocrinol. 2008, 289, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Liu, B.; Jin, L.; Tao, H.; Yang, Z. Estrogen receptor beta exhibited anti-tumor effects on osteosarcoma cells by regulating integrin, IAP, NF-kB/BCL-2 and PI3K/Akt signal pathway. J. Bone Oncol. 2017, 9, 15–20. [Google Scholar] [CrossRef]
- Chen, P.; Wang, H.; Duan, Z.; Zou, J.X.; Chen, H.; He, W.; Wang, J. Estrogen-related receptor alpha confers methotrexate resistance via attenuation of reactive oxygen species production and P53 mediated apoptosis in osteosarcoma cells. Biomed. Res. Int. 2014, 2014, 616025. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.X.; Li, X.A. Inhibitory effects of tamoxifen and doxorubicin, alone and in combination, on the proliferation of the MG63 human osteosarcoma cell line. Oncol. Lett. 2013, 6, 970–976. [Google Scholar] [CrossRef]
- Robl, B.; Pauli, C.; Botter, S.M.; Bode-Lesniewska, B.; Fuchs, B. Prognostic value of tumor suppressors in osteosarcoma before and after neoadjuvant chemotherapy. BMC Cancer 2015, 15, 379. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Guo, J.; Zhang, K.; Guo, Y. TP53 Mutations and Survival in Osteosarcoma Patients: A Meta-Analysis of Published Data. Dis. Markers 2016, 2016, 4639575. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Jun, Y.; Lv, Y.; Li, Y.; Zhang, Z.; Tao, M.; Chen, X.; He, J.; Zhang, L.; Wang, Q.L. Aptamer-conjugated and doxorubicin-loaded grapefruit-derived nanovectors for targeted therapy against HER2(+) breast cancer. J. Drug Target. 2019, 28, 186–294. [Google Scholar] [CrossRef] [PubMed]
- Herber, C.B.; Krause, W.C.; Wang, L.; Bayrer, J.R.; Li, A.; Schmitz, M.; Fields, A.; Ford, B.; Zhang, Z.; Reid, M.S.; et al. Estrogen signaling in arcuate Kiss1 neurons suppresses a sex-dependent female circuit promoting dense strong bones. Nat. Commun. 2019, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Hung, G.Y.; Yen, H.J.; Yen, C.C.; Wu, P.K.; Chen, C.F.; Chen, P.C.; Wu, H.T.; Chiou, H.J.; Chen, W.M. Improvement in High-Grade Osteosarcoma Survival: Results from 202 Patients Treated at a Single Institution in Taiwan. Medicine 2016, 95, e3420. [Google Scholar] [CrossRef] [PubMed]
- Kovac, M.; Blattmann, C.; Ribi, S.; Smida, J.; Mueller, N.S.; Engert, F.; Castro-Giner, F.; Weischenfeldt, J.; Kovacova, M.; Krieg, A.; et al. Exome sequencing of osteosarcoma reveals mutation signatures reminiscent of BRCA deficiency. Nat. Commun. 2015, 6, 8940. [Google Scholar] [CrossRef]
- Overholtzer, M.; Rao, P.H.; Favis, R.; Lu, X.Y.; Elowitz, M.B.; Barany, F.; Ladanyi, M.; Gorlick, R.; Levine, A.J. The presence of p53 mutations in human osteosarcomas correlates with high levels of genomic instability. Proc. Natl. Acad. Sci. USA 2003, 100, 11547–11552. [Google Scholar] [CrossRef] [Green Version]
- Walkley, C.R.; Qudsi, R.; Sankaran, V.G.; Perry, J.A.; Gostissa, M.; Roth, S.I.; Rodda, S.J.; Snay, E.; Dunning, P.; Fahey, F.H.; et al. Conditional mouse osteosarcoma, dependent on p53 loss and potentiated by loss of Rb, mimics the human disease. Genes Dev. 2008, 22, 1662–1676. [Google Scholar] [CrossRef] [Green Version]
- Hientz, K.; Mohr, A.; Bhakta-Guha, D.; Efferth, T. The role of p53 in cancer drug resistance and targeted chemotherapy. Oncotarget 2017, 8, 8921–8946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.M.; Shih, L.H.; Lee, J.Y.; Shen, Y.J.; Lee, H.H. Estrogen enhances activity of Wnt signaling during osteogenesis by inducing Fhl1 expression. J. Cell Biochem. 2015, 116, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- McDougall, K.E.; Perry, M.J.; Gibson, R.L.; Bright, J.M.; Colley, S.M.; Hodgin, J.B.; Smithies, O.; Tobias, J.H. Estrogen-induced osteogenesis in intact female mice lacking ERbeta. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E817–E823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, M.J.; Samuels, A.; Bird, D.; Tobias, J.H. Effects of high-dose estrogen on murine hematopoietic bone marrow precede those on osteogenesis. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E1159–E1165. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Li, S.; Chen, C.; Zhu, M.; Chen, Y.; Zhao, Z. 17betaEstradiol treatment drives Sp1 to upregulate MALAT1 expression and epigenetically affects physiological processes in U2OS cells. Mol. Med. Rep. 2017, 15, 1335–1342. [Google Scholar] [CrossRef] [Green Version]
- Fang, D.; Yang, H.; Lin, J.; Teng, Y.; Jiang, Y.; Chen, J.; Li, Y. 17beta-estradiol regulates cell proliferation, colony formation, migration, invasion and promotes apoptosis by upregulating miR-9 and thus degrades MALAT-1 in osteosarcoma cell MG-63 in an estrogen receptor-independent manner. Biochem. Biophys. Res. Commun. 2015, 457, 500–506. [Google Scholar] [CrossRef]
- Ruza, E.; Sotillo, E.; Sierrasesumaga, L.; Azcona, C.; Patino-Garcia, A. Analysis of polymorphisms of the vitamin D receptor, estrogen receptor, and collagen Ialpha1 genes and their relationship with height in children with bone cancer. J. Pediatr. Hematol. Oncol. 2003, 25, 780–786. [Google Scholar] [CrossRef]
- Savage, S.A.; Mirabello, L. Using epidemiology and genomics to understand osteosarcoma etiology. Sarcoma 2011, 2011, 548151. [Google Scholar] [CrossRef] [Green Version]
- Lillo Osuna, M.A.; Garcia-Lopez, J.; El Ayachi, I.; Fatima, I.; Khalid, A.B.; Kumpati, J.; Slayden, A.V.; Seagroves, T.N.; Miranda-Carboni, G.A.; Krum, S.A. Activation of Estrogen Receptor Alpha by Decitabine Inhibits Osteosarcoma Growth and Metastasis. Cancer Res. 2019, 79, 1054–1068. [Google Scholar] [CrossRef] [PubMed]
- Dohi, O.; Hatori, M.; Suzuki, T.; Ono, K.; Hosaka, M.; Akahira, J.; Miki, Y.; Nagasaki, S.; Itoi, E.; Sasano, H. Sex steroid receptors expression and hormone-induced cell proliferation in human osteosarcoma. Cancer Sci. 2008, 99, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Chandar, N.; Billig, B.; McMaster, J.; Novak, J. Inactivation of p53 gene in human and murine osteosarcoma cells. Br.J. Cancer 1992, 65, 208–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solakidi, S.; Psarra, A.M.; Sekeris, C.E. Differential subcellular distribution of estrogen receptor isoforms: Localization of ERalpha in the nucleoli and ERbeta in the mitochondria of human osteosarcoma SaOS-2 and hepatocarcinoma HepG2 cell lines. Biochim. Biophys. Acta. 2005, 1745, 382–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johmura, Y.; Maeda, I.; Suzuki, N.; Wu, W.; Goda, A.; Morita, M.; Yamaguchi, K.; Yamamoto, M.; Nagasawa, S.; Kojima, Y.; et al. Fbxo22-mediated KDM4B degradation determines selective estrogen receptor modulator activity in breast cancer. J. Clin. Investig. 2018, 128, 5603–5619. [Google Scholar] [CrossRef]
- Lin, P.I.; Tai, Y.T.; Chan, W.P.; Lin, Y.L.; Liao, M.H.; Chen, R.M. Estrogen/ERalpha signaling axis participates in osteoblast maturation via upregulating chromosomal and mitochondrial complex gene expressions. Oncotarget 2018, 9, 1169–1186. [Google Scholar] [CrossRef] [Green Version]
- Allan, L.A.; Fried, M. p53-dependent apoptosis or growth arrest induced by different forms of radiation in U2OS cells: p21WAF1/CIP1 repression in UV induced apoptosis. Oncogene 1999, 18, 5403–5412. [Google Scholar] [CrossRef] [Green Version]
- Marcellus, R.C.; Teodoro, J.G.; Charbonneau, R.; Shore, G.C.; Branton, P.E. Expression of p53 in Saos-2 osteosarcoma cells induces apoptosis which can be inhibited by Bcl-2 or the adenovirus E1B-55 kDa protein. Cell Growth Differ. 1996, 7, 1643–1650. [Google Scholar]
- Sun, C.; Liu, Z.; Li, S.; Yang, C.; Xue, R.; Xi, Y.; Wang, L.; Wang, S.; He, Q.; Huang, J.; et al. Down-regulation of c-Met and Bcl2 by microRNA-206, activates apoptosis, and inhibits tumor cell proliferation, migration and colony formation. Oncotarget 2015, 6, 25533–25574. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ERα(+)/P53(+) | ERα(−)/P53(+) | ERα(+)/P53(−) | ERα(−)/P53(−) | p-Value | |
|---|---|---|---|---|---|
| (n = 29) | (n = 11) | (n = 7) | (n = 3) | ||
| Gender | 0.6961 | ||||
| Male | 16 | 7 | 2 | 0 | |
| Female | 17 | 6 | 1 | 1 | |
| Average age | 21.5 ± 2.3 | 25.3 ± 4.8 | 19.3 ± 4.9 | 13.5 ± 0.5 | 0.6874 |
| Lung metastasis | 0.7016 | ||||
| No metastasis | 10 | 6 | 3 | 1 | |
| Metastasis | 19 | 5 | 4 | 2 | |
| Chemoresponse rate | 0.0358 * | ||||
| Good response (necrosis rate > 90%) | 13 | 8 | 1 | 0 | |
| Poor response (necrosis rate < 90%) | 16 | 3 | 6 | 3 | |
| Tumor volume (mm3) | 315 ± 84.7 | 113 ± 30.3 | 433 ± 63.9 | 140 ± 23.2 | 0.0151 * |
| 5-year survival | 21 (72.4%) | 9 (81.8%) | 3 (42.8%) | 0 (0%) | 0.0028 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.-Y.; Chen, C.-M.; Chen, C.-F.; Wu, P.-K.; Chen, W.-M. Suppression of Estrogen Receptor Alpha Inhibits Cell Proliferation, Differentiation and Enhances the Chemosensitivity of P53-Positive U2OS Osteosarcoma Cell. Int. J. Mol. Sci. 2021, 22, 11238. https://doi.org/10.3390/ijms222011238
Wang J-Y, Chen C-M, Chen C-F, Wu P-K, Chen W-M. Suppression of Estrogen Receptor Alpha Inhibits Cell Proliferation, Differentiation and Enhances the Chemosensitivity of P53-Positive U2OS Osteosarcoma Cell. International Journal of Molecular Sciences. 2021; 22(20):11238. https://doi.org/10.3390/ijms222011238
Chicago/Turabian StyleWang, Jir-You, Chao-Ming Chen, Cheng-Fong Chen, Po-Kuei Wu, and Wei-Ming Chen. 2021. "Suppression of Estrogen Receptor Alpha Inhibits Cell Proliferation, Differentiation and Enhances the Chemosensitivity of P53-Positive U2OS Osteosarcoma Cell" International Journal of Molecular Sciences 22, no. 20: 11238. https://doi.org/10.3390/ijms222011238