
Cardiac Telocytes 16 Years on—What Have We Learned So Far, and How Close Are We to Routine Application of the Knowledge in Cardiovascular Regenerative Medicine?

 , and
, and 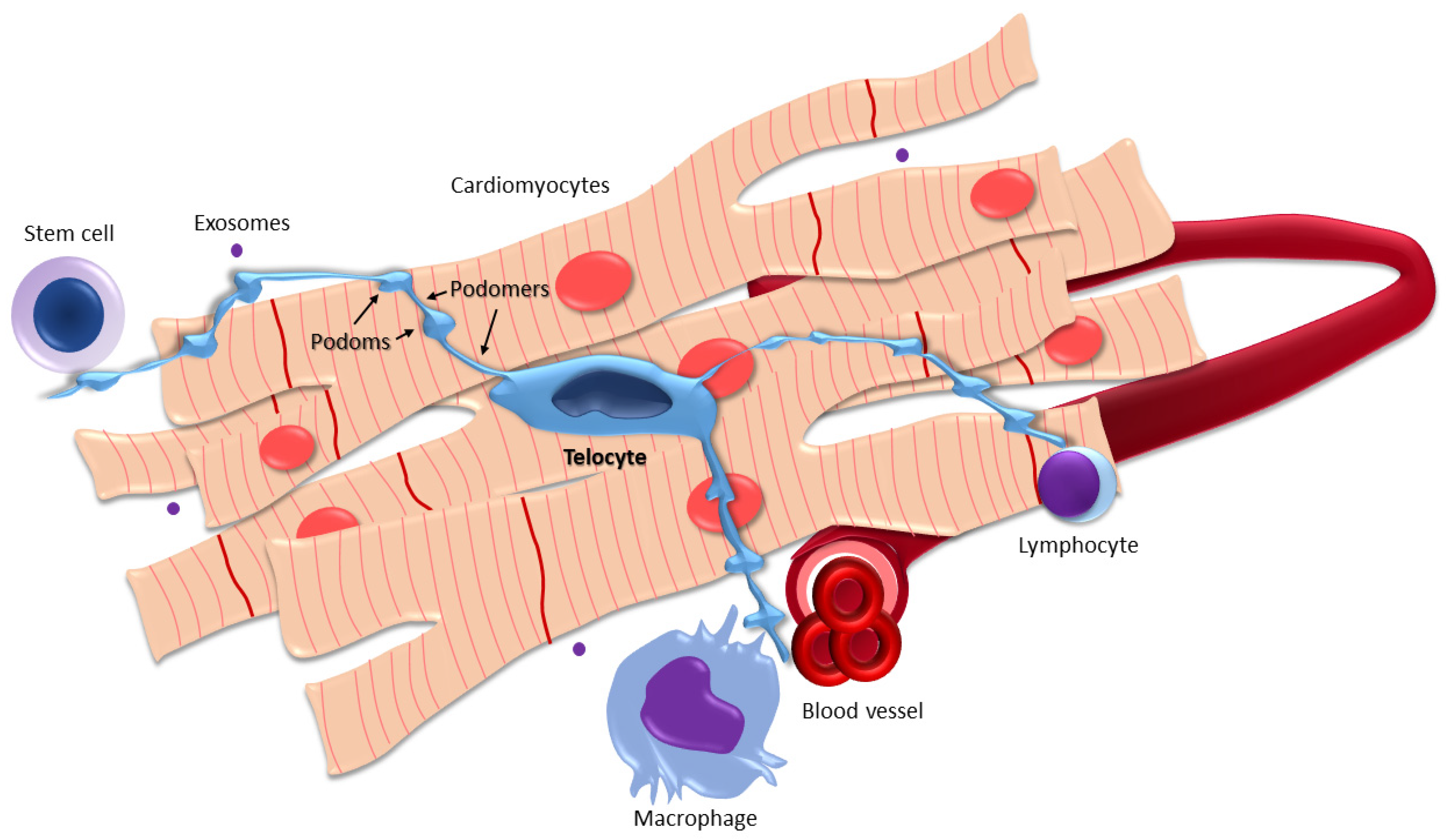{kind=link}
Abstract
:1. Introduction
2. Morphology, Location, Functions, and Topographic Relations of Cardiac Telocytes
3. Telocytes in Heart Diseases
3.1. Myocardial Infarction
3.2. Arrhythmia
4. Cardiac Telocytes in Cardiovascular Regenerative Medicine—Recent Developments
5. Controversies in Telocyte Research
6. Conclusions and Future Perspectives
- The standardization of protocols for TCs isolation, cultivation, and transplantation (although standardized protocols for the isolation of cardiac TCs are yet to be established, Romano et al. previously successfully isolated TCs from human skin using a novel two-step immunomagnetic microbead-based cell separation, taking advantage of the CD34+/CD31− immunophenotype of TCs [81]; this methodology could also be applied for selective purification of cardiac TCs in future research);
- Figuring out the applicability and extrapolation of the results of animal model studies to human medicine;
- Viability assessment of transplanted TCs;
- Quantitative optimization of transplanted cells for each patient;
- Cost-effectiveness analysis.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jensen, R.V.; Hjortbak, M.V.; Bøtker, H.E. Ischemic Heart Disease: An Update. Semin. Nucl. Med. 2020, 50, 195–207. [Google Scholar] [CrossRef]
- Giacca, M. Cardiac Regeneration After Myocardial Infarction: An Approachable Goal. Curr. Cardiol. Rep. 2020, 22, 122. [Google Scholar] [CrossRef]
- Cui, B.; Zheng, Y.; Sun, L.; Shi, T.; Shi, Z.; Wang, L.; Huang, G.; Sun, N. Heart Regeneration in Adult Mammals after Myocardial Damage. Acta Cardiol. Sin. 2018, 34, 115–123. [Google Scholar] [CrossRef]
- Wen, H.; Peng, L.; Chen, Y. The effect of immune cell-derived exosomes in the cardiac tissue repair after myocardial infarction: Molecular mechanisms and pre-clinical evidence. J. Cell. Mol. Med. 2021, 25, 6500–6510. [Google Scholar] [CrossRef] [PubMed]
- Veress, B.; Ohlsson, B. Spatial relationship between telocytes, interstitial cells of Cajal and the enteric nervous system in the human ileum and colon. J. Cell. Mol. Med. 2020, 24, 3399–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, L.M.; Faussone-Pellegrini, M.S. TELOCYTES—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, L.M.; Hinescu, M.E.; Ionescu, N.; Ciontea, S.M.; Cretoiu, D.; Ardelean, C. Interstitial cells of Cajal in pancreas. J. Cell. Mol. Med. 2005, 9, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Hinescu, M.E.; Radu, E.; Ciontea, S.M.; Cretoiu, D.; Leabu, M.; Ardeleanu, C. CD117/c-kit positive interstitial (Cajal-like) cells in human pancreas. J. Cell. Mol. Med. 2005, 9, 738–739. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Kondo, A.; Kaestner, K.H. Emerging diverse roles of telocytes. Development 2019, 146. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yu, P.; Zhong, S.; Ge, T.; Peng, S.; Guo, X.; Zhou, Z. Telocytes in pancreas of the Chinese giant salamander (Andrias davidianus). J. Cell. Mol. Med. 2016, 20, 2215–2219. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, M.I.; Popescu, L.M. Telocytes in the interstitium of human exocrine pancreas: Ultrastructural evidence. Pancreas 2012, 41, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Wang, F.; Liu, Z.; Yang, C. Telocytes in liver: Electron microscopic and immunofluorescent evidence. J. Cell. Mol. Med. 2013, 17, 1537–1542. [Google Scholar] [CrossRef]
- Cretoiu, S.M. Immunohistochemistry of Telocytes in the Uterus and Fallopian Tubes. Adv. Exp. Med. Biol. 2016, 913, 335–357. [Google Scholar] [CrossRef]
- Liu, T.; Wang, S.; Li, Q.; Huang, Y.; Chen, C.; Zheng, J. Telocytes as potential targets in a cyclophosphamide-induced animal model of premature ovarian failure. Mol. Med. Rep. 2016, 14, 2415–2422. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elhafeez, H.H.; Soliman, S.A. New Description of Telocyte Sheaths in the Bovine Uterine Tube: An Immunohistochemical and Scanning Microscopic Study. Cells Tissues Organs 2017, 203, 295–315. [Google Scholar] [CrossRef]
- Ceafalan, L.; Gherghiceanu, M.; Popescu, L.M.; Simionescu, O. Telocytes in human skin--are they involved in skin regeneration? J. Cell. Mol. Med. 2012, 16, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Lin, M.; Xu, M.; Manole, C.G.; Wang, X.; Zhu, T. Telocytes in the human kidney cortex. J. Cell. Mol. Med. 2012, 16, 3116–3122. [Google Scholar] [CrossRef]
- Luesma, M.J.; Gherghiceanu, M.; Popescu, L.M. Telocytes and stem cells in limbus and uvea of mouse eye. J. Cell. Mol. Med. 2013, 17, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.M.; Mokhtar, D.M. The roles of telocytes in lung development and angiogenesis: An immunohistochemical, ultrastructural, scanning electron microscopy and morphometrical study. Dev. Biol. 2018, 443, 137–152. [Google Scholar] [CrossRef]
- Pasternak, A.; Gil, K.; Matyja, A. Telocytes: New Players in Gallstone Disease. Adv. Exp. Med. Biol. 2016, 913, 77–103. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G.; Traini, C.; Manetti, M.; Ibba-Manneschi, L.; Faussone-Pellegrini, M.S. Telocytes express PDGFRα in the human gastrointestinal tract. J. Cell. Mol. Med. 2013, 17, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G.; Traini, C. Interstitial cells of Cajal and telocytes in the gut: Twins, related or simply neighbor cells? Biomol. Concepts 2016, 7, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G. The Telocytes: Ten Years after Their Introduction in the Scientific Literature. An Update on Their Morphology, Distribution, and Potential Roles in the Gut. Int. J. Mol. Sci. 2020, 21, 4478. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.G.; Traini, C.; Guasti, D.; Giulio, D.P.; Faussone-Pellegrini, M.S. Telocytes subtypes in human urinary bladder. J. Cell. Mol. Med. 2014, 18, 2000–2008. [Google Scholar] [CrossRef] [PubMed]
- Wagener, M.G.; Leonhard-Marek, S.; Häger, J.D.; Pfarrer, C. CD117- and vimentin-positive telocytes in the bovine teat sphincter. Anat. Histol. Embryol. 2018, 47, 268–270. [Google Scholar] [CrossRef] [Green Version]
- Popescu, L.M.; Curici, A.; Wang, E.; Zhang, H.; Hu, S.; Gherghiceanu, M. Telocytes and putative stem cells in ageing human heart. J. Cell. Mol. Med. 2015, 19, 31–45. [Google Scholar] [CrossRef]
- Shim, W. Myocardial Telocytes: A New Player in Electric Circuitry of the Heart. Adv. Exp. Med. Biol. 2016, 913, 241–251. [Google Scholar] [CrossRef]
- Tay, H.; Vandecasteele, T.; Van den Broeck, W. Identification of telocytes in the porcine heart. Anat. Histol. Embryol. 2017, 46, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Gherghiceanu, M.; Suciu, L.C.; Manole, C.G.; Hinescu, M.E. Telocytes and putative stem cells in the lungs: Electron microscopy, electron tomography and laser scanning microscopy. Cell Tissue Res. 2011, 345, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Cretoiu, D.; Hummel, E.; Zimmermann, H.; Gherghiceanu, M.; Popescu, L.M. Human cardiac telocytes: 3D imaging by FIB-SEM tomography. J. Cell. Mol. Med. 2014, 18, 2157–2164. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Cretoiu, D.; Marin, A.; Radu, B.M.; Popescu, L.M. Telocytes: Ultrastructural, immunohistochemical and electrophysiological characteristics in human myometrium. Reproduction 2013, 145, 357–370. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Valladares, F.; Madrid, J.F. CD34+ stromal cells/fibroblasts/fibrocytes/telocytes as a tissue reserve and a principal source of mesenchymal cells. Location, morphology, function and role in pathology. Histol. Histopathol. 2014, 29, 831–870. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wei, L.; Zhong, C.; Fu, S.; Bei, Y.; Huică, R.I.; Wang, F.; Xiao, J. Cardiac telocytes are double positive for CD34/PDGFR-α. J. Cell. Mol. Med. 2015, 19, 2036–2042. [Google Scholar] [CrossRef] [PubMed]
- Lis, G.J.; Dubrowski, A.; Lis, M.; Solewski, B.; Witkowska, K.; Aleksandrovych, V.; Jasek-Gajda, E.; Hołda, M.K.; Gil, K.; Litwin, J.A. Identification of CD34+/PGDFRα+ Valve Interstitial Cells (VICs) in Human Aortic Valves: Association of Their Abundance, Morphology and Spatial Organization with Early Calcific Remodeling. Int. J. Mol. Sci. 2020, 21, 6330. [Google Scholar] [CrossRef] [PubMed]
- Gherghiceanu, M.; Manole, C.G.; Popescu, L.M. Telocytes in endocardium: Electron microscope evidence. J. Cell. Mol. Med. 2010, 14, 2330–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherghiceanu, M.; Popescu, L.M. Cardiomyocyte precursors and telocytes in epicardial stem cell niche: Electron microscope images. J. Cell. Mol. Med. 2010, 14, 871–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, L.M.; Gherghiceanu, M.; Manole, C.G.; Faussone-Pellegrini, M.S. Cardiac renewing: Interstitial Cajal-like cells nurse cardiomyocyte progenitors in epicardial stem cell niches. J. Cell. Mol. Med. 2009, 13, 866–886. [Google Scholar] [CrossRef] [Green Version]
- Popescu, L.M.; Manole, C.G.; Gherghiceanu, M.; Ardelean, A.; Nicolescu, M.I.; Hinescu, M.E.; Kostin, S. Telocytes in human epicardium. J. Cell. Mol. Med. 2010, 14, 2085–2093. [Google Scholar] [CrossRef] [Green Version]
- Rosa, I.; Marini, M.; Manetti, M. Telocytes: An Emerging Component of Stem Cell Niche Microenvironment. J. Histochem. Cytochem. 2021, 221554211025489. [Google Scholar] [CrossRef]
- Nagy, E.; Jermendy, A.L.; Merkely, B.; Maurovich-Horvat, P. Clinical importance of epicardial adipose tissue. Arch. Med. Sci. 2017, 13, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Nerlekar, N.; Thakur, U.; Lin, A.; Koh, J.Q.S.; Potter, E.; Liu, D.; Muthalaly, R.G.; Rashid, H.N.; Cameron, J.D.; Dey, D.; et al. The Natural history of Epicardial Adipose Tissue Volume and Attenuation: A long-term prospective cohort follow-up study. Sci. Rep. 2020, 10, 7109. [Google Scholar] [CrossRef] [PubMed]
- Christensen, R.H.; von Scholten, B.J.; Lehrskov, L.L.; Rossing, P.; Jørgensen, P.G. Epicardial adipose tissue: An emerging biomarker of cardiovascular complications in type 2 diabetes? Ther. Adv. Endocrinol. Metab. 2020, 11, 2042018820928824. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Carrasco, J.L.; Alvarez-Argüelles, H.; Díaz-Flores, L., Jr. Telocytes/CD34+ Stromal Cells in Pathologically Affected White Adipose Tissue. Int. J. Mol. Sci. 2020, 21, 9694. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Chen, S.; Liu, J.; Yuan, Z.; Qi, X.; Qin, J.; Zheng, X.; Shen, X.; Yu, Y.; Qnin, T.J.; et al. Cardiac telocytes were decreased during myocardial infarction and their therapeutic effects for ischaemic heart in rat. J. Cell. Mol. Med. 2013, 17, 123–133. [Google Scholar] [CrossRef]
- Liskova, Y.V.; Stadnikov, A.A.; Salikova, S.P. The role of telocytes in myocardial remodeling and the development of cardiovascular complications in patients with chronic heart failure after coronary artery bypass grafting. Kardiologiia 2018, 58, 29–37. [Google Scholar] [CrossRef]
- Lin, Y.K.; Chen, Y.J. Telocytes: Supporting cells participating in ventricular arrhythmogenesis? J. Arrhythm. 2019, 35, 522–523. [Google Scholar] [CrossRef] [Green Version]
- Mandache, E.; Gherghiceanu, M.; Macarie, C.; Kostin, S.; Popescu, L.M. Telocytes in human isolated atrial amyloidosis:ultrastructural remodelling. J. Cell. Mol. Med. 2010, 14, 2739–2747. [Google Scholar] [CrossRef] [Green Version]
- Nour, M.S.; Sarhan, N.R.; Mazroa, S.A.; Gawish, S.A. Histological and immunohistochemical study of cardiac telocytes in a rat model of isoproterenol-induced myocardial infarction with a reference to the effect of grape seed extract. Acta Histochem. 2017, 119, 747–758. [Google Scholar] [CrossRef]
- Varga, I.; Kyselovic, J.; Danihel, L.; Klein, M.; Barczi, T.; Galfiova, P.; Danisovic, L. Cardiac telocytes as principal interstitial cells for myocardial reparation and regeneration after infarction—Our hope. Bratisl. Lek. Listy 2017, 118, 721–723. [Google Scholar] [CrossRef]
- Jenča, D.; Melenovský, V.; Stehlik, J.; Staněk, V.; Kettner, J.; Kautzner, J.; Adámková, V.; Wohlfahrt, P. Heart failure after myocardial infarction: Incidence and predictors. ESC Heart Failure 2021, 8, 222–237. [Google Scholar] [CrossRef]
- Richter, M.; Kostin, S. The failing human heart is characterized by decreased numbers of telocytes as result of apoptosis and altered extracellular matrix composition. J. Cell. Mol. Med. 2015, 19, 2597–2606. [Google Scholar] [CrossRef]
- Manetti, M.; Rosa, I.; Messerini, L.; Ibba-Manneschi, L. Telocytes are reduced during fibrotic remodelling of the colonic wall in ulcerative colitis. J. Cell. Mol. Med. 2015, 19, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Czubryt, M.P. Cardiac Fibroblast to Myofibroblast Phenotype Conversion-An Unexploited Therapeutic Target. J. Cardiovasc. Dev. Dis. 2019, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manetti, M.; Rosa, I.; Messerini, L.; Guiducci, S.; Matucci-Cerinic, M.; Ibba-Manneschi, L. A loss of telocytes accompanies fibrosis of multiple organs in systemic sclerosis. J. Cell. Mol. Med. 2014, 18, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Banciu, D.D.; Banciu, A.; Radu, B.M. Electrophysiological Features of Telocytes. Adv. Exp. Med. Biol. 2016, 913, 287–302. [Google Scholar] [CrossRef] [PubMed]
- DeSimone, C.V.; McLeod, C.J.; Gomez Pinilla, P.J.; Beyder, A.; Farrugia, G.; Asirvatham, S.J.; Kapa, S. Telocytes express ANO-1-encoded chloride channels in canine ventricular myocardium. J. Arrhythm. 2019, 35, 515–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitrofanova, L.B.; Gorshkov, A.N.; Konovalov, P.V.; Krylova, J.S. Telocytes in the human sinoatrial node. J. Cell. Mol. Med. 2018, 22, 521–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherghiceanu, M.; Hinescu, M.E.; Andrei, F.; Mandache, E.; Macarie, C.E.; Faussone-Pellegrini, M.S.; Popescu, L.M. Interstitial Cajal-like cells (ICLC) in myocardial sleeves of human pulmonary veins. J. Cell. Mol. Med. 2008, 12, 1777–1781. [Google Scholar] [CrossRef] [Green Version]
- Bei, Y.; Zhou, Q.; Sun, Q.; Xiao, J. Telocytes in cardiac regeneration and repair. Semin. Cell Dev. Biol. 2016, 55, 14–21. [Google Scholar] [CrossRef]
- Faussone-Pellegrini, M.S.; Bani, D. Relationships between telocytes and cardiomyocytes during pre- and post-natal life. J. Cell. Mol. Med. 2010, 14, 1061–1063. [Google Scholar] [CrossRef] [Green Version]
- Sukhacheva, T.V.; Nizyaeva, N.V.; Samsonova, M.V.; Chernyaev, A.L.; Shchegolev, A.I.; Serov, R.A. Telocytes in the Myocardium of Children with Congenital Heart Disease Tetralogy of Fallot. Bull. Exp. Biol. Med. 2020, 169, 137–146. [Google Scholar] [CrossRef]
- Liao, Z.; Chen, Y.; Duan, C.; Zhu, K.; Huang, R.; Zhao, H.; Hintze, M.; Pu, Q.; Yuan, Z.; Lv, L.; et al. Cardiac telocytes inhibit cardiac microvascular endothelial cell apoptosis through exosomal miRNA-21-5p-targeted cdip1 silencing to improve angiogenesis following myocardial infarction. Theranostics 2021, 11, 268–291. [Google Scholar] [CrossRef]
- Cismaşiu, V.B.; Popescu, L.M. Telocytes transfer extracellular vesicles loaded with microRNAs to stem cells. J. Cell. Mol. Med. 2015, 19, 351–358. [Google Scholar] [CrossRef]
- Marini, M.; Ibba-Manneschi, L.; Manetti, M. Cardiac Telocyte-Derived Exosomes and Their Possible Implications in Cardiovascular Pathophysiology. Adv. Exp. Med. Biol. 2017, 998, 237–254. [Google Scholar] [CrossRef]
- Popescu, L.M.; Fertig, E.T.; Gherghiceanu, M. Reaching out: Junctions between cardiac telocytes and cardiac stem cells in culture. J. Cell. Mol. Med. 2016, 20, 370–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.; Liao, Z.; Luo, J.; Chen, H.; Guo, H.; Yang, J.; Huang, R.; Pu, Q.; Zhao, H.; Yuan, Z.; et al. Cardiac telocytes exist in the adult Xenopus tropicalis heart. J. Cell. Mol. Med. 2020, 24, 2531–2541. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Li, L.; Qi, G.; Hu, M.; Hu, C.; Wang, S.; Li, J.; Zhang, M.; Zhang, W.; Zeng, Y.; et al. Transplantation of Telocytes Attenuates Unilateral Ureter Obstruction-Induced Renal Fibrosis in Rats. Cell. Physiol. Biochem. 2018, 46, 2056–2071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Song, D.; Shi, L.; Sun, X.; Zheng, Y.; Zeng, Y.; Wang, X. Mechanisms of interactions between lung-origin telocytes and mesenchymal stem cells to treat experimental acute lung injury. Clin. Transl. Med. 2020, 10, e231. [Google Scholar] [CrossRef] [PubMed]
- Ibba-Manneschi, L.; Rosa, I.; Manetti, M. Telocytes in Chronic Inflammatory and Fibrotic Diseases. Adv. Exp. Med. Biol. 2016, 913, 51–76. [Google Scholar] [CrossRef] [PubMed]
- Miao, Q.; Shim, W.; Tee, N.; Lim, S.Y.; Chung, Y.Y.; Ja, K.P.; Ooi, T.H.; Tan, G.; Kong, G.; Wei, H.; et al. iPSC-derived human mesenchymal stem cells improve myocardial strain of infarcted myocardium. J. Cell. Mol. Med. 2014, 18, 1644–1654. [Google Scholar] [CrossRef]
- Ja, K.P.; Miao, Q.; Zhen Tee, N.G.; Lim, S.Y.; Nandihalli, M.; Ramachandra, C.J.A.; Mehta, A.; Shim, W. iPSC-derived human cardiac progenitor cells improve ventricular remodelling via angiogenesis and interstitial networking of infarcted myocardium. J. Cell. Mol. Med. 2016, 20, 323–332. [Google Scholar] [CrossRef] [Green Version]
- FIPAT. Terminologia Histologica: International Terms for Human Cytology and Histology; Wolters Kluwer; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008; p. 300. [Google Scholar]
- Varga, I.; Gálfiová, P.; Blanková, A.; Konarik, M.; Báča, V.; Dvořákova, V.; Musil, V.; Turyna, R.; Klein, M. Terminologia Histologica 10 years on: Some disputable terms in need of discussion and recent developments. Ann. Anat. 2019, 226, 16–22. [Google Scholar] [CrossRef]
- Rusu, M.C.; Hostiuc, S. Critical review: Cardiac telocytes vs cardiac lymphatic endothelial cells. Ann. Anat. 2019, 222, 40–54. [Google Scholar] [CrossRef]
- Rosa, I.; Marini, M.; Sgambati, E.; Ibba-Manneschi, L.; Manetti, M. Telocytes and lymphatic endothelial cells: Two immunophenotypically distinct and spatially close cell entities. Acta Histochem. 2020, 122, 151530. [Google Scholar] [CrossRef] [PubMed]
- Rosa, I.; Taverna, C.; Novelli, L.; Marini, M.; Ibba-Manneschi, L.; Manetti, M. Telocytes constitute a widespread interstitial meshwork in the lamina propria and underlying striated muscle of human tongue. Sci. Rep. 2019, 9, 5858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostiuc, S.; Marinescu, M.; Costescu, M.; Aluaș, M.; Negoi, I. Cardiac telocytes. From basic science to cardiac diseases. II. Acute myocardial infarction. Ann. Anat. 2018, 218, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Kyselovič, J.; Danišovič, Ľ.; Gálfiová, P.; Kachlík, D.; Polák, Š.; Klein, M. Recently discovered interstitial cells termed telocytes: Distinguishing cell-biological and histological facts from fictions. Biologia 2019, 74, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Bei, Y.; Zhou, Q.; Fu, S.; Lv, D.; Chen, P.; Chen, Y.; Wang, F.; Xiao, J. Cardiac telocytes and fibroblasts in primary culture: Different morphologies and immunophenotypes. PLoS ONE 2015, 10, e0115991. [Google Scholar] [CrossRef] [PubMed]
- Romano, E.; Rosa, I.; Fioretto, B.S.; Lucattelli, E.; Innocenti, M.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Manetti, M. A Two-Step Immunomagnetic Microbead-Based Method for the Isolation of Human Primary Skin Telocytes/CD34+ Stromal Cells. Int. J. Mol. Sci. 2020, 21, 5877. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klein, M.; Csöbönyeiová, M.; Žiaran, S.; Danišovič, Ľ.; Varga, I. Cardiac Telocytes 16 Years on—What Have We Learned So Far, and How Close Are We to Routine Application of the Knowledge in Cardiovascular Regenerative Medicine? Int. J. Mol. Sci. 2021, 22, 10942. https://doi.org/10.3390/ijms222010942
Klein M, Csöbönyeiová M, Žiaran S, Danišovič Ľ, Varga I. Cardiac Telocytes 16 Years on—What Have We Learned So Far, and How Close Are We to Routine Application of the Knowledge in Cardiovascular Regenerative Medicine? International Journal of Molecular Sciences. 2021; 22(20):10942. https://doi.org/10.3390/ijms222010942
Chicago/Turabian StyleKlein, Martin, Mária Csöbönyeiová, Stanislav Žiaran, Ľuboš Danišovič, and Ivan Varga. 2021. "Cardiac Telocytes 16 Years on—What Have We Learned So Far, and How Close Are We to Routine Application of the Knowledge in Cardiovascular Regenerative Medicine?" International Journal of Molecular Sciences 22, no. 20: 10942. https://doi.org/10.3390/ijms222010942