
Characterization of Human Medullary Thyroid Carcinoma Glycosphingolipids Identifies Potential Cancer Markers

 , ,
, , 
Abstract
:1. Introduction
2. Results
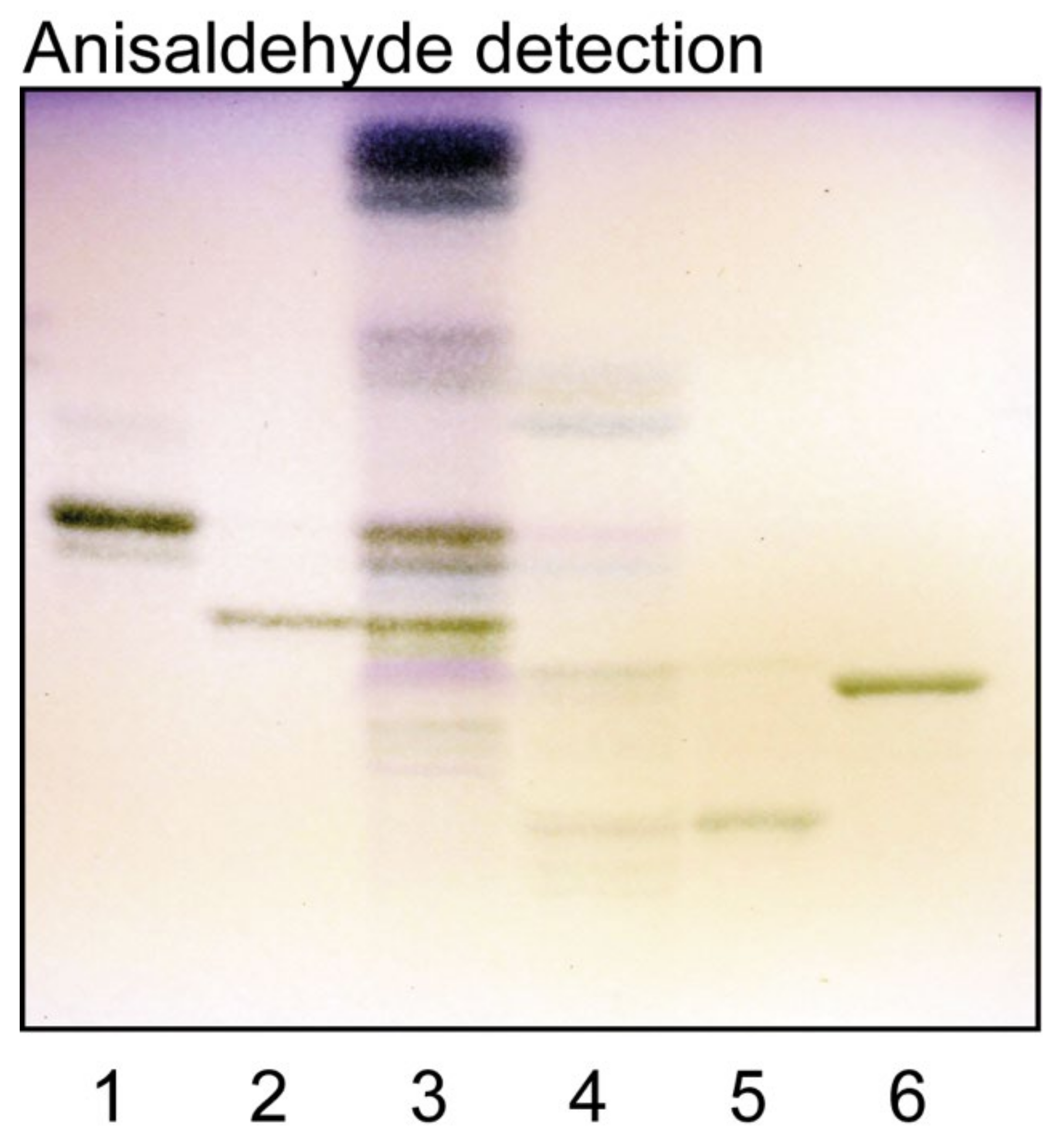
2.1. Isolation of Human Medullary Thyroid Cancer Glycosphingolipids
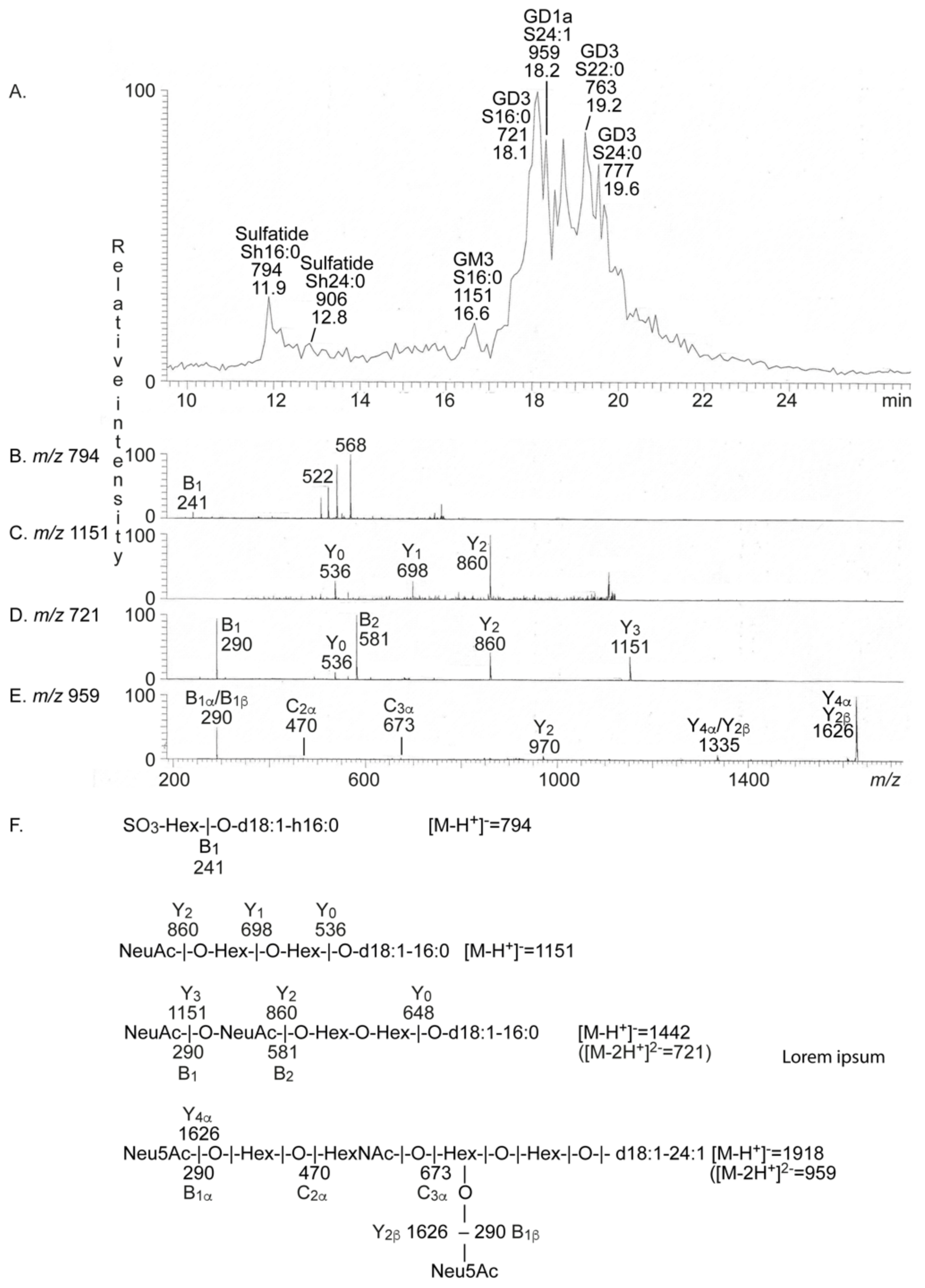
2.2. Characterization of the Acid Glycosphingolipids from Human Medullary Thyroid Cancer
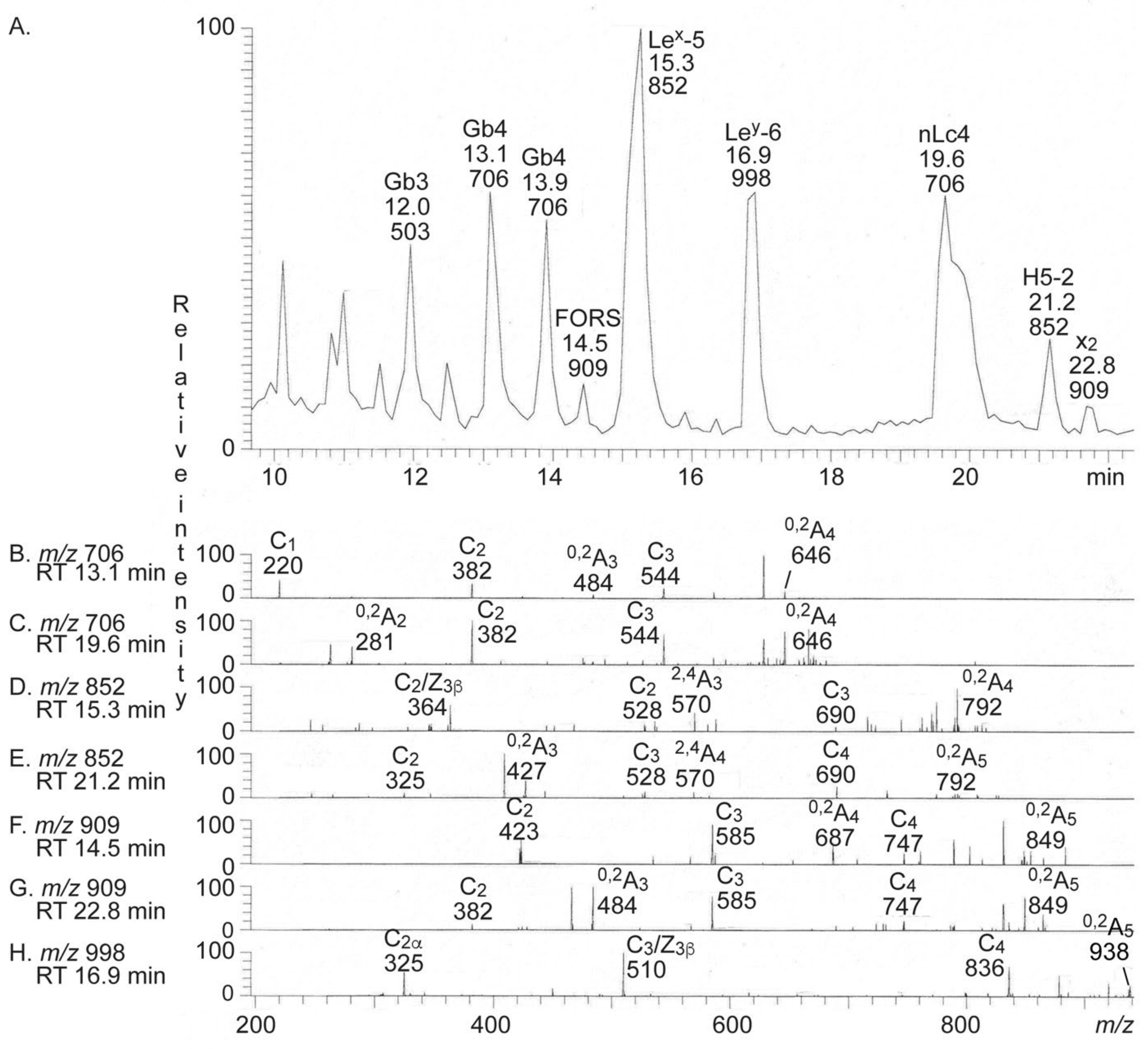
2.3. Characterization of the Non-Acid Glycosphingolipids from Human Medullary Thyroid Cancer
2.3.1. LC-ESI/MS of Glycosphingolipid-Derived Oligosaccharides
α-N-Acetylgalactosaminidase Hydrolysis
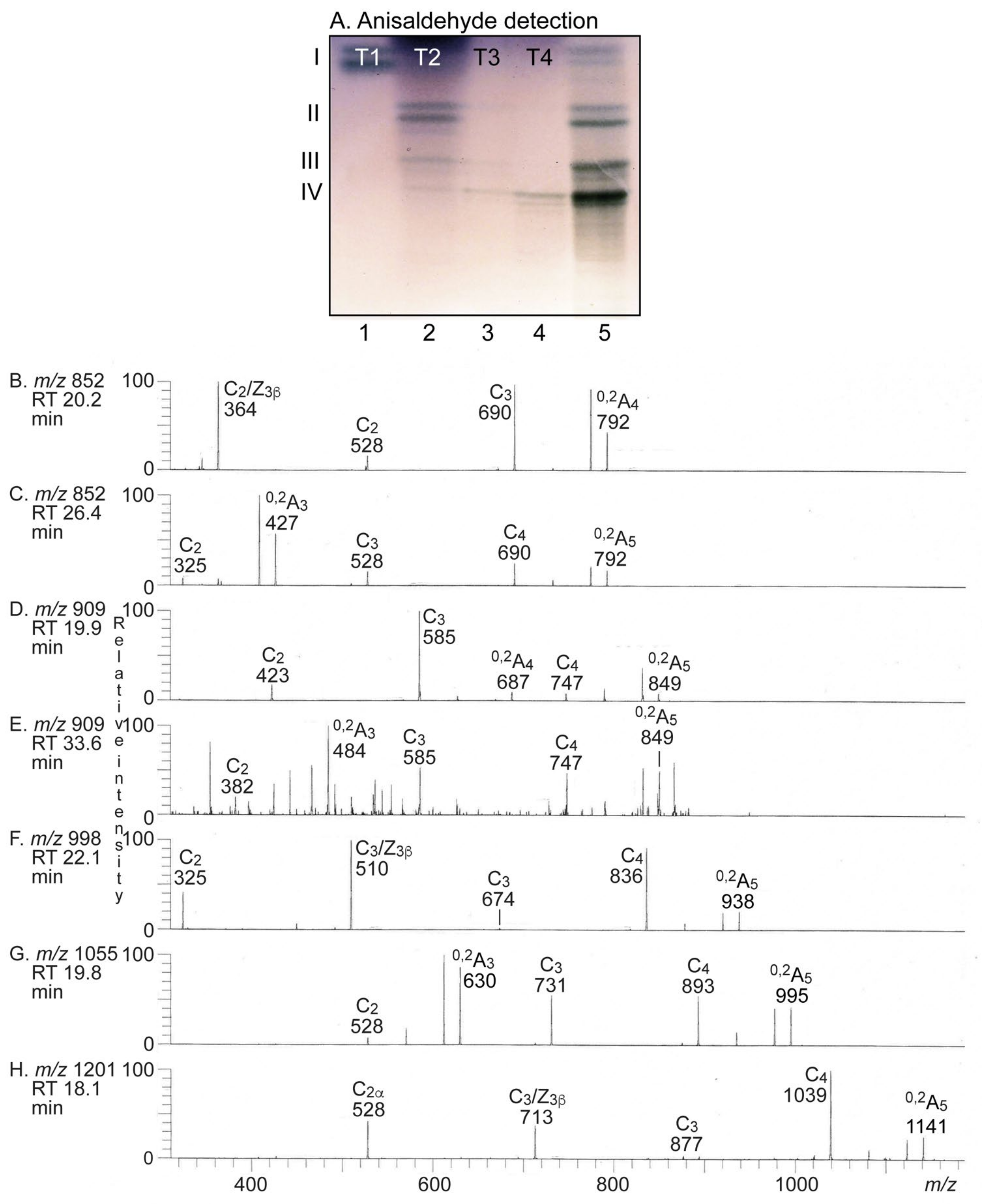
2.3.2. Separation of the Non-Acid Glycosphingolipids
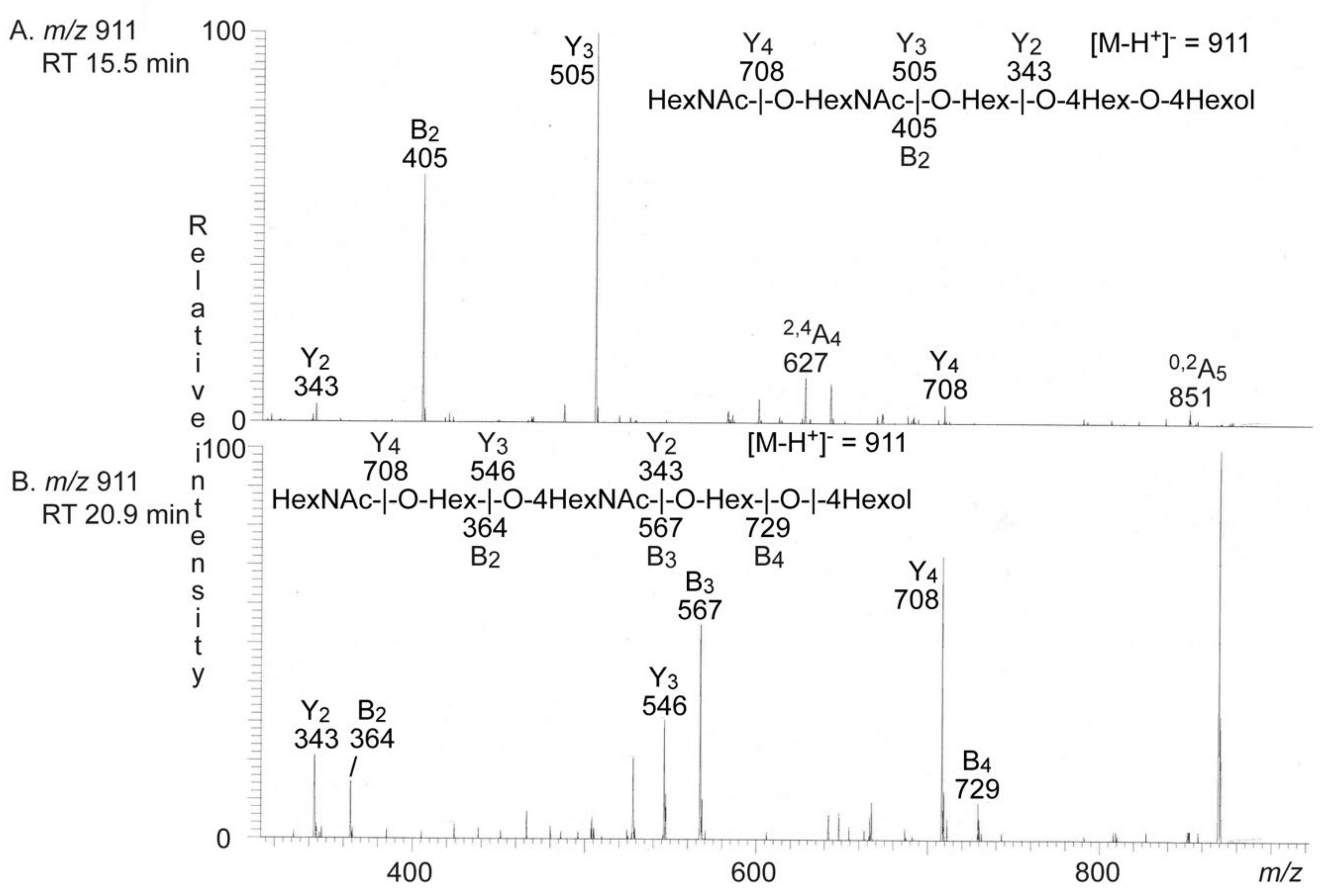
2.3.3. LC-ESI/MS of Fraction T-1
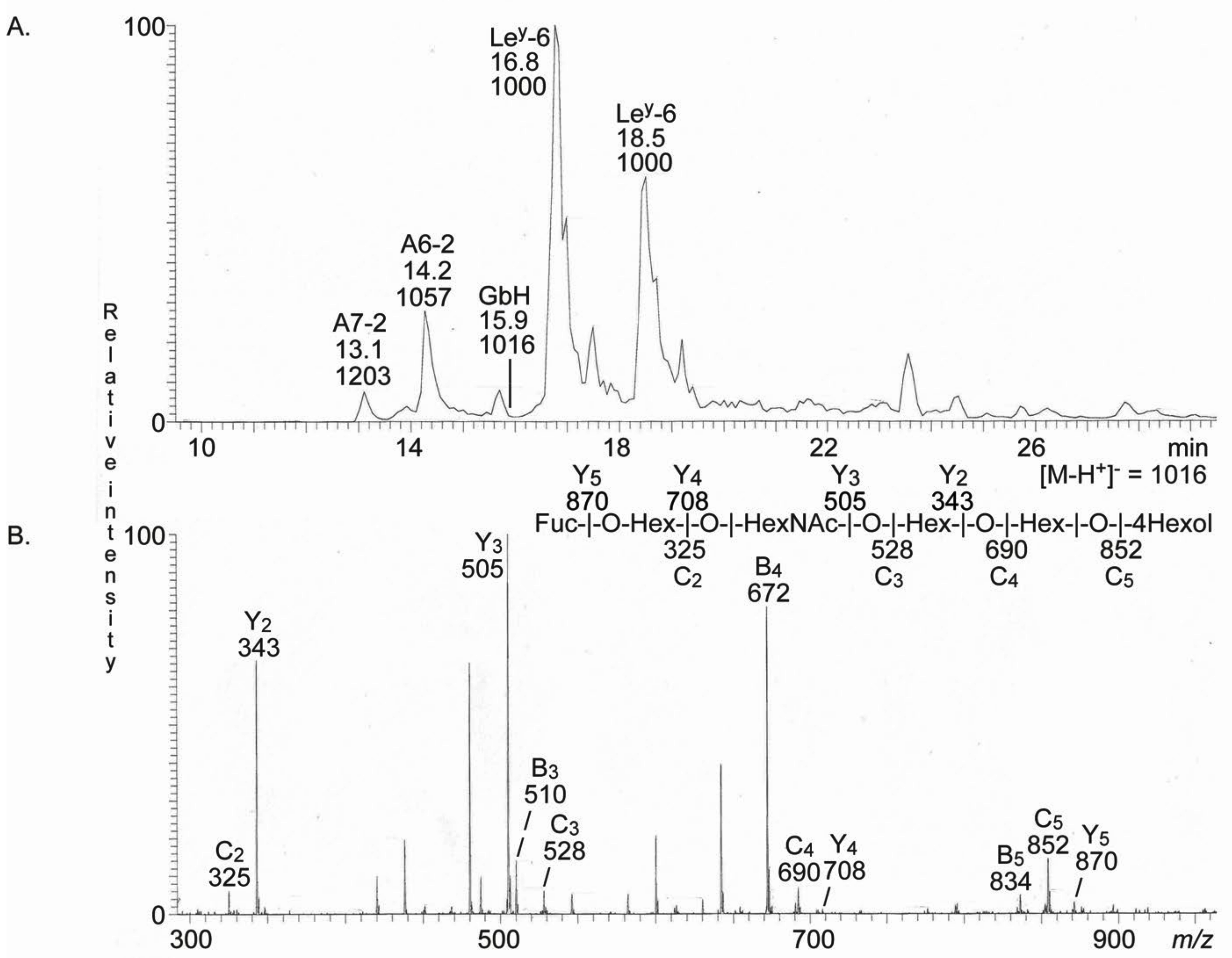
2.3.4. LC-ESI/MS of Fractions T2–T4
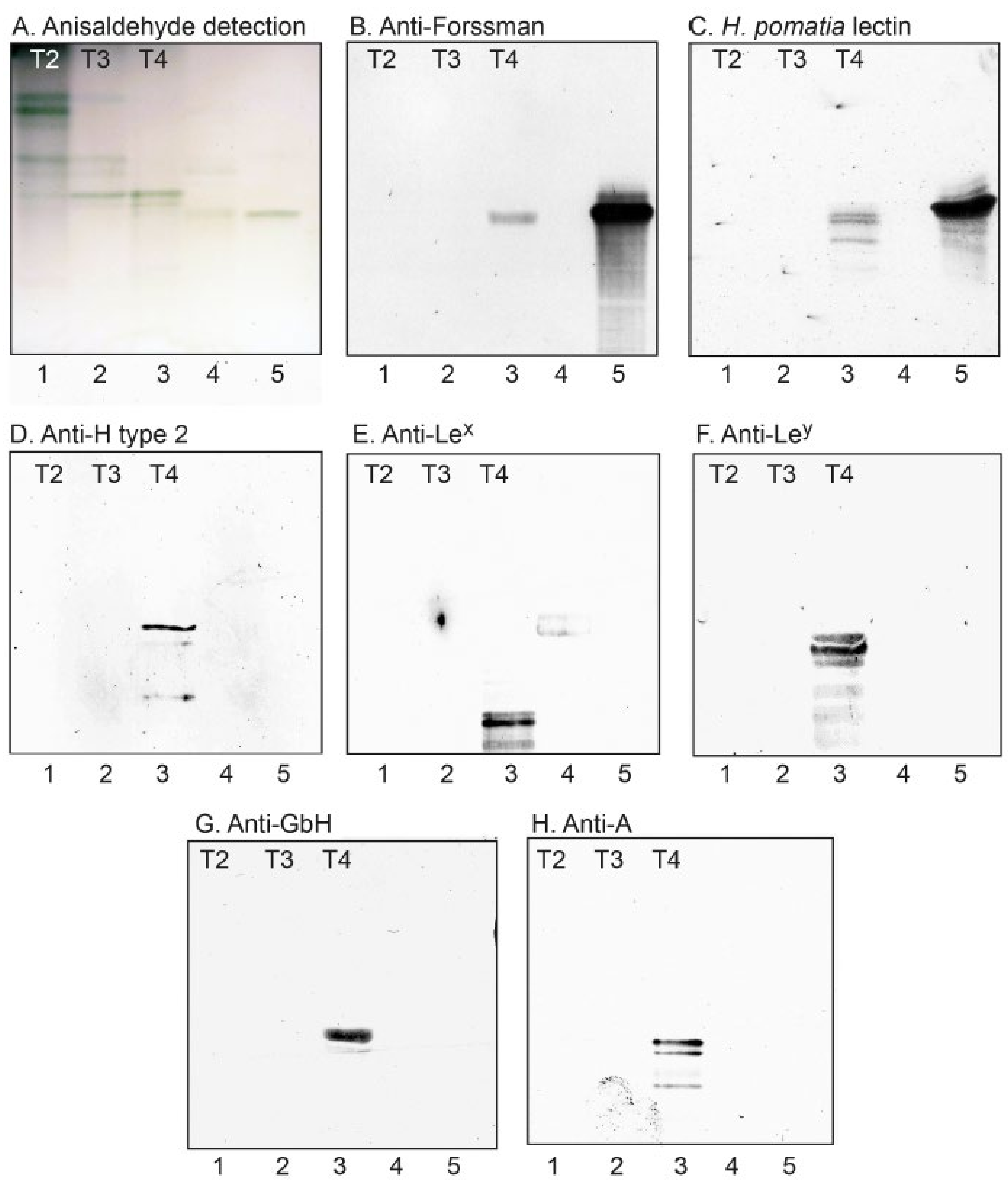
2.3.5. Binding of Carbohydrate Recognizing Ligands to the Non-Acid Glycosphingolipids
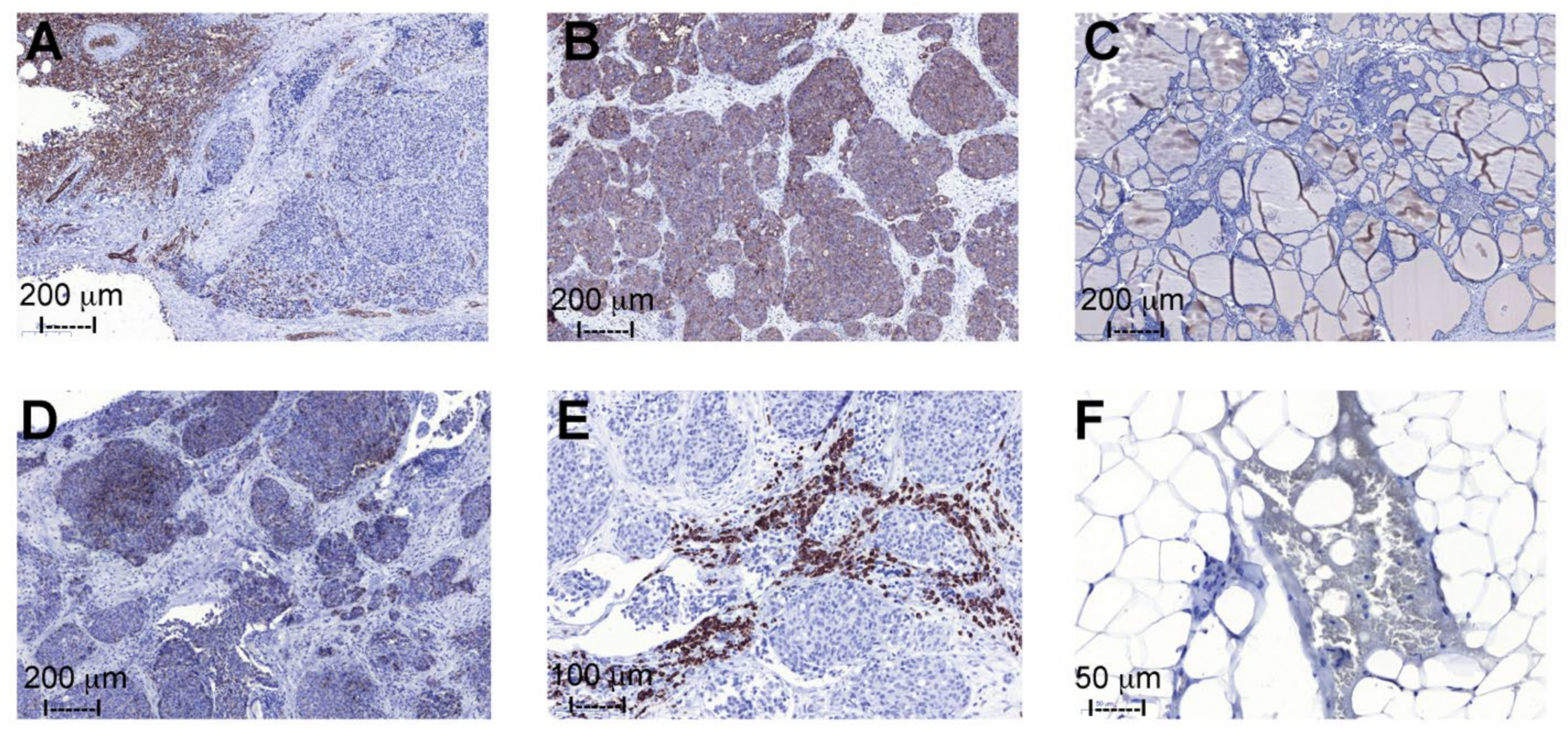
2.4. Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Glycosphingolipid Preparations
4.2. Reference Glycosphingolipids
4.3. Thin-Layer Chromatography
4.4. Chromatogram Binding Assays
4.5. LC-ESI/MS of Native Acid Glycosphingolipids
4.6. Endoglycoceramidase Digestion and LC-ESI/MS
α-N-Acetylgalactosaminidase Hydrolysis
4.7. Histology and Immunohistochemistry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Alexander, E.K.; Pearce, E.N.; Brent, G.A.; Brown, P.S.; Chen, H.; Dosiou, C.; Grobman, W.A.; Laurberg, P.; Lazarus, J.H.; Mandel, S.J.; et al. Guidelines of the American Thyroid Association for the diagnosis and management of thyroid disease during pregnancy and the postpartum. Thyroid 2017, 27, 315–389. [Google Scholar] [CrossRef] [Green Version]
- Williams, E.D. Histogenesis of medullary carcinoma of the thyroid. J. Clin. Pathol. 1966, 19, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Kurzrock, R.; Sherman, S.I.; Ball, D.W.; Forastiere, A.A.; Cohen, R.B.; Mehra, R.; Pfister, D.G.; Cohen, E.E.W.; Janisch, L.; Nauling, F.; et al. Activity of XL184 (Cabozantinib), an oral tyrosine kinase inhibitor, in patients with medullary thyroid cancer. J. Clin. Oncol. 2011, 29, 2660–2666. [Google Scholar] [CrossRef]
- Wells, S.A., Jr.; Robinson, B.G.; Gagel, R.F.; Dralle, H.; Fagin, J.A.; Santoro, M.; Baudin, E.; Elisei, R.; Jarzab, B.; Vasselli, J.R.; et al. Vandetanib in patients with locally advanced or metastatic medullary thyroid cancer: A randomized, double-blind phase III trial. J. Clin. Oncol. 2012, 30, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakomori, S. The general concept of tumor-associated carbohydrate antigens: Their chemical, physical and enzymatic basis. In Gangliosides and Cancer; Oettgen, H.F., Ed.; VCH Verlagsgesellschaft mbH: Weinheim, Germany, 1989; pp. 57–68. [Google Scholar]
- Fuster, M.M.; Esko, J.D. The sweet and sour of cancer: Glycans as novel therapeutic targets. Nat. Rev. Cancer 2005, 5, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Kannagi, R.; Toole, B.P. Glycosylation changes in cancer. In Essentials of Glycobiology; Varki, A., Cummings, R.D., Esko, J.D., Freeze, H.H., Stanley, P., Bertozzi, C.R., Hart, G.W., Etzler, M.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009; pp. 617–632. [Google Scholar]
- Pearce, O.M.T. Cancer glycan epitopes: Biosynthesis, structure and function. Glycobiology 2018, 28, 670–696. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, E.; Ito, Y.; Miyoshi, Y. Involvement of aberrant glycosylation in thyroid cancer. J. Oncol. 2010, 2010, 816595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ząbczyńska, M.; Kozłowska, K.; Pocheć, E. Glycosylation in the thyroid gland: Vital aspects of glycoprotein function in thyrocyte physiology and thyroid disorders. Int. J. Mol. Sci. 2018, 19, 2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchon, B.; Portoukalian, J.; Bornet, H. Major gangliosides in normal and pathological human thyroids. Biochem. Int. 1985, 10, 531–538. [Google Scholar] [PubMed]
- Mariano, A.; Di Carlo, A.; Macchia, P.E.; Pisano, G.; Parmeggiani, U.; Macchia, V. Gangliosides and phospholipids in human thyroids responsive and unresponsive to thyrotropin. J. Endocrinol. Invest. 1990, 13, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, K.A. Preparation of total non-acid glycolipids for overlay analysis of receptors for bacteria and viruses and for other studies. Meth. Enzymol. 1987, 138, 212–220. [Google Scholar] [CrossRef]
- Säljö, K.; Thornell, A.; Jin, C.; Stålberg, P.; Norlén, O.; Teneberg, S. Characterization of glycosphingolipids in the human parathyroid and thyroid glands. Int. J. Mol. Sci. 2021, 22, 7044. [Google Scholar] [CrossRef]
- Karlsson, H.; Halim, A.; Teneberg, S. Differentiation of glycosphingolipid-derived glycan structural isomers by liquid chromatography-mass spectrometry. Glycobiology 2010, 20, 1103–1116. [Google Scholar] [CrossRef] [Green Version]
- Chai, W.; Piskarev, V.; Lawson, A.M. Negative-ion electrospray mass spectrometry of neutral underivatized oligosaccharides. Anal. Chem. 2001, 73, 651–657. [Google Scholar] [CrossRef]
- Svensson, L.; Hult, A.K.; Stamps, R.; Ångström, J.; Teneberg, S.; Storry, J.R.; Jørgensen, R.; Rydberg, L.; Henry, S.M.; Olsson, M.L. Forssman expression on human erythrocytes: Biochemical and genetic evidence of a new histo-blood group system. Blood 2013, 121, 1459–1468. [Google Scholar] [CrossRef] [Green Version]
- Westman, J.S.; Benktander, J.; Storry, J.R.; Peyrard, T.; Hult, A.K.; Hellberg, Å.; Teneberg, S.; Olsson, M.L. Identification of the molecular and genetic basis of PX2, a glycosphingolipid blood group antigen lacking on globoside-deficient erythrocytes. J. Biol. Chem. 2015, 290, 18505–18518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.; Barone, A.; Borén, T.; Teneberg, S. Helicobacter pylori binding non-acid glycosphingolipids in the human stomach. J. Biol. Chem. 2018, 293, 17248–17266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, J.F.; Lescar, J.; Chazalet, V.; Audfray, A.; Gagnon, J.; Alvarez, R.; Breton, C.; Imberty, A.; Mitchell, E.P. Biochemical and structural analysis of Helix pomatia agglutinin. A hexameric lectin with a novel fold. J. Biol. Chem. 2006, 281, 20171–20180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babál, P.; Janega, P.; Cerná, A.; Kholová, I.; Brabencová, E. Neoplastic transformation of the thyroid gland is accompanied by changes in cellular sialylation. Acta Histochem. 2006, 108, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Vierbuchen, M.; Schröder, S.; Larena, A.; Uhlenbruck, G.; Fischer, R. Native and sialic acid masked Lewis(a) antigen reactivity in medullary thyroid carcinoma. Distinct tumour-associated and prognostic relevant antigens. Virchows Arch. 1994, 424, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Krzeslak, A.; Gaj, Z.; Pomorski, L.; Lipinska, A. Sialylation of intracellular proteins of thyroid lesions. Oncol. Rep. 2007, 17, 1237–1242. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Yamagata, T. Purification and characterization of glycosphingolipid-specific endoglycosidases (endoglycoceramidases) from a mutant strain of Rhodococcus sp. Evidence for three molecular species of endoglycoceramidase with different specificities. J. Biol. Chem. 1989, 264, 9510–9519. [Google Scholar] [CrossRef]
- Li, Y.T.; Chou, C.W.; Li, S.C.; Kobayashi, U.; Ishibashi, Y.h.; Ito, M. Preparation of homogenous oligosaccharide chains from glycosphingolipids. Glycoconj. J. 2009, 26, 929–933. [Google Scholar] [CrossRef]
- Yu, J.; Hung, J.T.; Wang, S.H.; Cheng, J.Y.; Yu, A.L. Targeting glycosphingolipids for cancer immunotherapy. FEBS Lett. 2020, 594, 3602–3618. [Google Scholar] [CrossRef]
- Cheng, S.P.; Yang, P.S.; Chien, M.N.; Chen, M.J.; Lee, J.J.; Liu, C.L. Aberrant expression of tumor-associated carbohydrate antigen Globo H in thyroid carcinoma. J. Surg. Oncol. 2016, 114, 853–858. [Google Scholar] [CrossRef]
- Hult, A.K.; McSherry, E.; Moller, M.; Olsson, M.L. GBGT1 is allelically diverse but dispensable in humans and naturally occurring anti-FORS1 shows an ABO-restricted pattern. Transfusion 2018, 58, 2036–2045. [Google Scholar] [CrossRef]
- Luetscher, R.N.D.; McKitrick, T.R.; Gao, C.; Mehta, A.Y.; Mcquillan, A.M.; Kardish, R.; Boligan, K.F.; Song, X.; Lu, L.; Heimburg-Molinaro, J.; et al. Unique repertoire of anti-carbohydrate antibodies in individual human serum. Sci. Rep. 2020, 10, 15436. [Google Scholar] [CrossRef]
- Hakomori, S.; Wang, S.M.; Young, W.W., Jr. Isoantigenic expression of Forssman glycolipid in human gastric and colonic mucosa: Its possible identity with A-like “antigen” in human cancer. Proc. Natl. Acad. Sci. USA 1977, 74, 3023–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breimer, M.E. Chemical and immunological identification of the forssman pentaglycosylceramide in human kidney. Glycoconj. J. 1985, 2, 375–385. [Google Scholar]
- Hult, A.K.; Olsson, M.L. The FORS awakens: Review of a blood group system reborn. Immunohematology 2017, 33, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.T.; Gulley, J.L.; Oyelaran, O.; Hodge, J.W.; Schlom, J.; Gildersleeve, J.C. Humoral response to a viral glycan correlates with survival on PROSTVAC-VF. Proc. Natl. Acad. Sci. USA 2014, 111, E1749–E1758. [Google Scholar] [CrossRef] [Green Version]
- Vierbuchen, M.; Larena, A.; Schröder, S.; Hanisch, F.G.; Ortmann, M.; Larena, A.; Uhlenbruck, G.; Fischer, R. Blood group antigen expression in medullary carcinoma of the thyroid. An immunohistochemical study on the occurrence of type 1 chain-derived antigens. Virchows Arch. B Cell. Pathol. Incl. Mol. Pathol. 1992, 62, 79–88. [Google Scholar] [CrossRef]
- Larena, A.; Vierbuchen, M.; Fischer, R. Blood group antigen expression in malignant tumors of the thyroid: A parallel between medullary and nonmedullary carcinomas. Langenbecks Arch. Chir. 1995, 380, 269–272. [Google Scholar] [CrossRef]
- González-Cámpora, R.; García-Sanatana, J.A.; i Heras, M.M.J.; Salaverri, C.O.; Vázquez-Ramírez, F.J.; Argueta-Manzano, O.E.; Galera-Davidson, H. Blood group antigens in differentiated thyroid neoplasms. Arch. Pathol. Lab. Med. 1998, 122, 957–965. [Google Scholar] [PubMed]
- Manimala, J.C.; Roach, T.A.; Li, Z.; Gildersleeve, J.C. High-throughput carbohydrate microarray profiling of 27 antibodies demonstrates widespread specificity problems. Glycobiology 2007, 17, 17C–23C. [Google Scholar] [CrossRef]
- Samuelsson, B.E.; Pimlott, W.; Karlsson, K.A. Mass spectrometry of mixtures of intact glycosphingolipids. Meth. Enzymol. 1990, 193, 623–646. [Google Scholar] [CrossRef]
- Koerner, T.A.W., Jr.; Prestegard, J.H.; Demou, P.C.; Yu, R.K. High-resolution proton NMR studies of gangliosides. 1. Use of homonuclear spin-echo J-correlated spectroscopy for determination of residue composition and anomeric configurations. Biochemistry 1983, 22, 2676–2687. [Google Scholar] [CrossRef]
- Waldi, D. Sprühreagentien für die dünnschicht-chromatographie. In Dünnschicht-Chromatographie; Stahl, E., Ed.; Springer: Berlin, Germany, 1962; pp. 496–515. [Google Scholar]
- Barone, A.; Benktander, J.; Ångström, J.; Aspegren, A.; Björquist, P.; Teneberg, S.; Breimer, M.E. Structural complexity of non-acid glycosphingolipids in human embryonic stem cells grown under feeder-free conditions. J. Biol. Chem. 2013, 288, 10035–10050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barone, A.; Säljö, K.; Benktander, J.; Blomqvist, M.; Månsson, J.E.; Johansson, B.R.; Mölne, J.; Aspegren, A.; Björquist, P.; Breimer, M.E.; et al. Sialyl-lactotetra: A novel cell surface marker of undifferentiated human pluripotent stem cells. J. Biol. Chem. 2014, 289, 18846–18859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teneberg, S.; Jovall, P.Å.; Ångström, J.; Karlsson, K.A. Characterization of binding of Galβ4GlcNAc-specific lectins from Erythrina christagalli and Erythrina corallodendron to glycosphingolipids. Detection, isolation and characteriztion of a novel glycosphingolipid of bovine buttermilk. J. Biol. Chem. 1994, 269, 8554–8563. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.M.; Dedic, B.; Lundholm, K.; Branzell, F.B.; Barone, A.; Benktander, J.; Teneberg, S. Characterization of moose intestinal glycosphingolipids. Glycoconj. J. 2015, 32, 393–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceroni, A.; Maass, K.; Geyer, H.; Geyer, R.; Dell, A.; Haslam, S.M. GlycoWorkbench: A Tool for the Computer-Assisted Annotation of Mass Spectra of Glycans. J. Proteome Res. 2008, 7, 1650–1659. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| m/z | Trivial Name | Structure |
|---|---|---|
| 794 | Sulfatide | SO3-3Galβ1Cer |
| 1151 | Neu5Ac-GM3 | Neu5Acα3Galβ4Glcβ1Cer |
| 721 | Neu5Ac-GD3 | Neu5Acα8Neu5Acα3Galβ4Glcβ1Cer |
| 959 | Neu5Ac-GD1a | Neu5Acα3Galβ3GalNAcβ4(Neu5Acα3)Galβ4Glcβ1Cer |
| m/z | Trivial Name | Structure |
|---|---|---|
| 503 | Globotri (Gb3) | Galα4Galβ4Glc |
| 706 | Neolactotetra (nLc4) | Galβ4GlcNAcβ3Galβ4Glc |
| 706 | Globotetra (Gb4) | GalNAcβ3Galα4Galβ4Glc |
| 909 | Forssman | GalNAcα3GalNAcβ3Galα4Galβ4Glc |
| 909 | x2 penta (x2) | GalNAcβ3Galβ4GlcNAcβ3Galβ4Glc |
| 852 | H type 2 penta (H5-2) | Fucα2Galβ4GlcNAcβ3Galβ4Glc |
| 852 | Lex penta (Lex-5) | Galβ4(Fucα3)GlcNAcβ3Galβ4Glc |
| 998 | Ley hexa (Ley-6) | Fucα2Galβ4(Fucα3)GlcNAcβ3Galβ4Glc |
| 1016 (red) * | Globo H | Fucα2Galβ3GalNAcβ3Galα4Galβ4Glc |
| 1055 | A hexa type 2 (A6-2) | GalNAcα3(Fucα2)Galβ4GlcNAcβ3Galβ4Glc |
| 1201 | A hepta type 2 (A7-2) | GalNAcα3(Fucα2)Galβ4(Fucα3)GlcNAcβ3Galβ4Glc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Säljö, K.; Thornell, A.; Jin, C.; Norlén, O.; Teneberg, S. Characterization of Human Medullary Thyroid Carcinoma Glycosphingolipids Identifies Potential Cancer Markers. Int. J. Mol. Sci. 2021, 22, 10463. https://doi.org/10.3390/ijms221910463
Säljö K, Thornell A, Jin C, Norlén O, Teneberg S. Characterization of Human Medullary Thyroid Carcinoma Glycosphingolipids Identifies Potential Cancer Markers. International Journal of Molecular Sciences. 2021; 22(19):10463. https://doi.org/10.3390/ijms221910463
Chicago/Turabian StyleSäljö, Karin, Anders Thornell, Chunsheng Jin, Olov Norlén, and Susann Teneberg. 2021. "Characterization of Human Medullary Thyroid Carcinoma Glycosphingolipids Identifies Potential Cancer Markers" International Journal of Molecular Sciences 22, no. 19: 10463. https://doi.org/10.3390/ijms221910463