
Integrative Role of 14-3-3ε in Sleep Regulation


Department of Entomology and MOA Key Lab of Pest Monitoring and Green Management, College of Plant Protection, China Agricultural University, Beijing 100193, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(18), 9748; https://doi.org/10.3390/ijms22189748
Submission received: 23 July 2021
/
Revised: 30 August 2021
/
Accepted: 2 September 2021
/
Published: 9 September 2021
(This article belongs to the Special Issue Genetics of Circadian Rhythms and Sleep)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Sleep is a crucial factor for health and survival in all animals. In this study, we found by proteomic analysis that some cancer related proteins were impacted by the circadian clock. The 14-3-3ε protein, expression of which is activated by the circadian transcription factor Clock, regulates adult sleep of Drosophila independent of circadian rhythm. Detailed analysis of the sleep regulatory mechanism shows that 14-3-3ε directly targets the Ultrabithorax (Ubx) gene to activate transcription of the pigment dispersing factor (PDF). The dopamine receptor (Dop1R1) and the octopamine receptor (Oamb), are also involved in the 14-3-3ε pathway, which in 14-3-3ε mutant flies causes increases in the dopR1 and OAMB, while downregulation of the DopR1 and Oamb can restore the sleep phenotype caused by the 14-3-3ε mutation. In conclusion, 14-3-3ε is necessary for sleep regulation in Drosophila.
1. Introduction
Sleep is very important for the health and survival of animals, and it is regulated mainly by the circadian rhythm and homeostasis [1,2,3]. Sleep has been extensively studied in the model animal Drosophila melanogaster, which is detected by measuring the activity through a Drosophila activity monitoring (DAM) system [4]. Fly immobility for 5 min or longer is defined as sleep [5,6].
In Drosophila, approximately 150 clock neurons in the central nervous system are involved in circadian rhythms, mainly including LNvs (ventral lateral neurons), LNds (dorsal lateral neurons) and DN1s (dorsal neurons) to form a feedback loop to control sleep-activity of Drosophila. The PDF-positive l-LNvs and s-LNvs (M cells) are known as arousal neurons. Loss of PDF neurons or PDF itself increases the amount of daytime sleep, while the CRY-positive LNds and the 5th s-LNv (E cells) control the amount of nighttime sleep; activation of the E cells causes sleep loss [7,8,9,10]. In addition, PDF neurons also modulate the phase of E cell oscillations [8,11]. PDF containing s-LNv dorsal projections exhibit a clock-controlled structural plasticity [12], in which some genes and microRNAs such as the adipokinetic hormone (AKH) [13] and microRNA-263b [14] involved in s-LNvs axonal fasciculation have been shown to impact circadian behavior.
The 14-3-3 family is highly conserved in protein sequence and function from yeast to mammals. They are involved in some biological processes such as cell proliferation, and apoptosis [15,16]. There are seven 14-3-3 members in vertebrates separately named ζ, δ, β, ε, γ, η, and θ according to their amino acid sequences. In Drosophila, there are two paralogs of 14-3-3 protein, ζ and ε, participating in both the Hippo pathway and the Ras/MAPK pathway [17,18,19]. Biochemistry data have shown that the isolated η chain of 14-3-3 protein from bovine brain can activate tyrosine hydroxylase and tryptophan hydroxylase in the presence of Ca2+/calmodulin-dependent protein kinase type II [20]. Inhibition of the 14-3-3 family of proteins results in functional reduction of glutamatergic synapses [21]. However, whether 14-3-3 is related to sleep is still unclear.
The Drosophila CLOCK (CLK) is one of the most important core oscillation proteins in the biological clock for controlling daily circadian rhythms and sleep, and its deficiency may result in a disorder of circadian rhythms and abnormal sleep. Therefore, we used the Clock-deficient mutant (ClkJrk) and the same background of wild-type (w1118) control flies to screen and identify the downstream circadian-related genes, in which the 14-3-3ε is found to be a sleep-regulating factor related to Clock. Thus, we focused on its mechanism of sleep regulation.
2. Materials and Methods
2.1. Fly Stocks
The following stocks were used in this study: Amph26, PssHP31723, 14-3-3εEP3578, 14-3-3εj2b10/+, Ubx-gal4/tm6b, UAS-mRFP, UAS-14-3-3εRNAi, pdf-gal4, 14-3-3εG00082, Dop1R1KO and OambMI11478. Amph26 (BS6498), PssHP31723 (BS22115), 14-3-3εEP3578 (BS17142), 14-3-3εj2b10/+ (BS12142), pdf-gal4 (BS41286), OambMI11478 (BS56423), and 14-3-3εG00082 (BS51385) were purchased from the Bloomington Drosophila Stock Center. UAS-14-3-3εRNAi (v15884) was purchased from the Vienna Drosophila Resource Center. Dop1R1KO was a gift from Dr. Yi Rao’s lab [22]. 14-3-3εEP3578 and 14-3-3εj2b10/+ mutants were derived by insertional mutagenesis using the different P-element constructs. They were backcrossed by w1118 for six generations.
All flies were reared at 25 °C and 65% relative humidity with standard corn flour/yeast/agar food supplemented with CaCl2 in a 12 h light/12 h dark cycle.
2.2. Behavioral Assays
Three to five day-old male adults were housed in monitor tubes (5[W] × 65[L] mm) with fly food. Experiments were performed in a Climate box at 25 ± 1 °C with 50% relative humidity. Light was turned on at ZT0 (06:30) and off at ZT12 (18:30). The activity data were recorded using the Drosophila Activity Monitoring System (Tri-kinetics, Waltham, MA, USA). The protocol and data analysis are described in Chen et al. (2013) [23].
2.3. Immunofluorescence
The flies were immobilized in 4% paraformaldehyde for 12 h at 4˚C and were then dissected in phosphate-buffered saline (PBS). The brains were blocked in blocking buffer (10% Normal Goat Serum diluted with 2% PBST) at RT for 2 h. The tissue was incubated in primary antibody for 24 h at 4 °C before being incubated with secondary antibodies overnight at 4 °C. The primary antibodies were as follows: mouse anti-PDF (DSHB UAS Cat# C7 monoclonal antibody; 1:400), rabbit anti-GFP (Invitrogen UAS Cat# PA1-980A polyclonal antibody; 1:400), and mouse anti-RFP (Abclonal China Cat# AE020 monoclonal antibody; 1:50). Fluorescent secondary antibodies conjugated to Goat anti-Rabbit FITC (Abclonal China Cat# AS011; 1:100) and Goat anti-Mouse TRITC (Abclonal China Cat# AS026; 1:100). The immunofluorescence assay was carried out on a Leica system (Leica SP8, Wetzlar, Germany).
2.4. Total RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR
Total RNA was isolated from heads of five to seven-day-old flies using RNAiso plus (TaKaRa Japan Cat# 9109). Each sample contained 30 individual flies with three biological repeats, which were reversely transcribed and measured by real-time PCR, respectively. There were three technical repeats in each biological repeat in the real-time PCR experiment. The total RNA quality was checked by Agilent Bioanalyser. A total of 1 μg RNA was added in each reverse transcription system. The RNA was reversely transcribed with a PrimeScripTM RT reagent Kit with gDNA Eraser (TaKaRa Japan Cat# RR047A). A total of 1μL cDNA was added in each real-time PCR reaction system. SYBR Green method was used for Real-time PCR with SuperReal PreMix Plus kit (TIANGEN China Cat# FP205-02). The PDF gene real-time PCR program: holding stage 95 °C 10mins; cycling stage 95 °C 15 s, 57 °C 25 s, 68 °C 35 s, 40 cycles; melt curve stage 95 °C 15 s, 60 °C 1 min, temperature increment +0.3 °C, 95 °C 15 s. The Ubx gene real-time PCR program: holding stage 95 °C 10 min; cycling stage 95 °C 15 s, 60 °C 20 s, 72 °C 30 s, 40cycles; melt curve stage 95 °C 15 s, 60 °C 1 min, temperature increment +0.3 °C, 95 °C 15 s. RP49 (also named as RpL32) was regarded as reference gene. w1118 control was used for normalization. The △△CT method was used for quantification. The real-time PCR data analysis is described in Livak et al. (2001) [24]. The real-time PCR assay was carried out on an Applied Biosystem Step One Real-Time PCR system (Applied Biosystem, Foster, CA, USA). The primers were designed by Beacon Designer 8. The sequences of the primers are shown in Table S4.
2.5. Western Blot Analysis and Co-Immunoprecipitation
D. melanogaster heads were collected and lysed with strong lysis buffer (CWBIO China Cat# CW2333) and protease inhibitor (CWBIO China Cat# CW2200). Whole tissue lysates were subjected to SDS-PAGE and immunoblotting as described (REF). The molecular weights of 14-3-3ε, Ubx and β-tubulin protein are 30KDa, 40KDa and 50KDa, respectively. The used primary antibodies were as follows: guinea pig anti-14-3-3ε (1:1000, from Aurelio A. Teleman as gift), mouse anti-Ubx (DSHB UAS Cat# FP3.38 monoclonal antibody; 1:50), and mouse anti-β-tubulin (Abclonal China Cat# AC010 monoclonal antibody; 1:1000). The used secondary antibodies were as follows: HRP Goat anti-Guinea Pig IgG (Abclonal China Cat# AS025; 1:2000), HRP Goat anti-Mouse IgG (Abclonal China Cat# AS003; 1:2000). Co-immunoprecipitation was conducted as previously described [25].
2.6. Chromatin Immunoprecipitation Assay
Chromatin immunoprecipitation was conducted as previously described [26]. w1118 strain fly heads were collected and fixed by shaking in 1% formaldehyde for 10 min at RT, and cross-linking reactions were stopped by adding glycine at a final concentration of 125 mM at RT for 5 min. The cross-linked chromatin was cut by sonication to approximately 200–500 bp fragments. A 120 μL sample of protein was used for immunoprecipitation, and 10 μL was maintained as the input DNA. The chromatin immunoprecipitation reaction was performed with 20 μL of antibody (mouse anti-Ubx). Immunoprecipitated DNA was quantified by real-time PCR. The ChIP-qPCR data were normalized by the input DNA, and the results were presented as the enrichment fold DNA. The sequences of the primers are shown in Table S4. Each experiment was independently performed three times.
2.7. Calculation of Axonal Cross
Axonal cross was used to quantify the morphology complexity. The data analysis is described in Fernández et al. (2008) [12]. Six evenly spaced (10 μm) concentric rings centered at the point where the first dorsal ramification opens up were drawn on each brain hemisphere. The number of intersections of each projection with a particular ring were counted. The total number of intersections were compared using nonparametric statistical methods.
2.8. Proteomic Screen and RNA-Seq
Total proteins from whole heads in both wild-type (w1118) and ClkJrk mutant flies were sampled at ZT2, ZT8, ZT14 and ZT20 and analyzed using the iTRAQ-MS method. Ingenuity Pathway Analysis (IPA) was used for protein screening. For the detailed methods of experiment and data analysis of proteomic screen, refer to our previous publication [27]. The total RNA extracted from Drosophila heads of homozygote 14-3-3εEP3578 and w1118 at ZT2 and ZT14 was used for RNA-seq. The RNA-seq was completed by Beijing Biomics Biotech Co. Ltd. (Beijing, China). Each sample contained 70 individual flies.
2.9. Statistical Analysis
Statistical analysis was performed with SPSS statistics 18.0. p values were obtained with One-way ANOVA, Two-way ANOVA and unpaired Student’s t-test and were considered to indicate significance; n.s. no significant difference, * p < 0.05, ** p < 0.01, *** p < 0.001, and **** p < 0.0001.
3. Results
3.1. 14-3-3ε Regulates Sleep Independent of the Circadian Rhythm
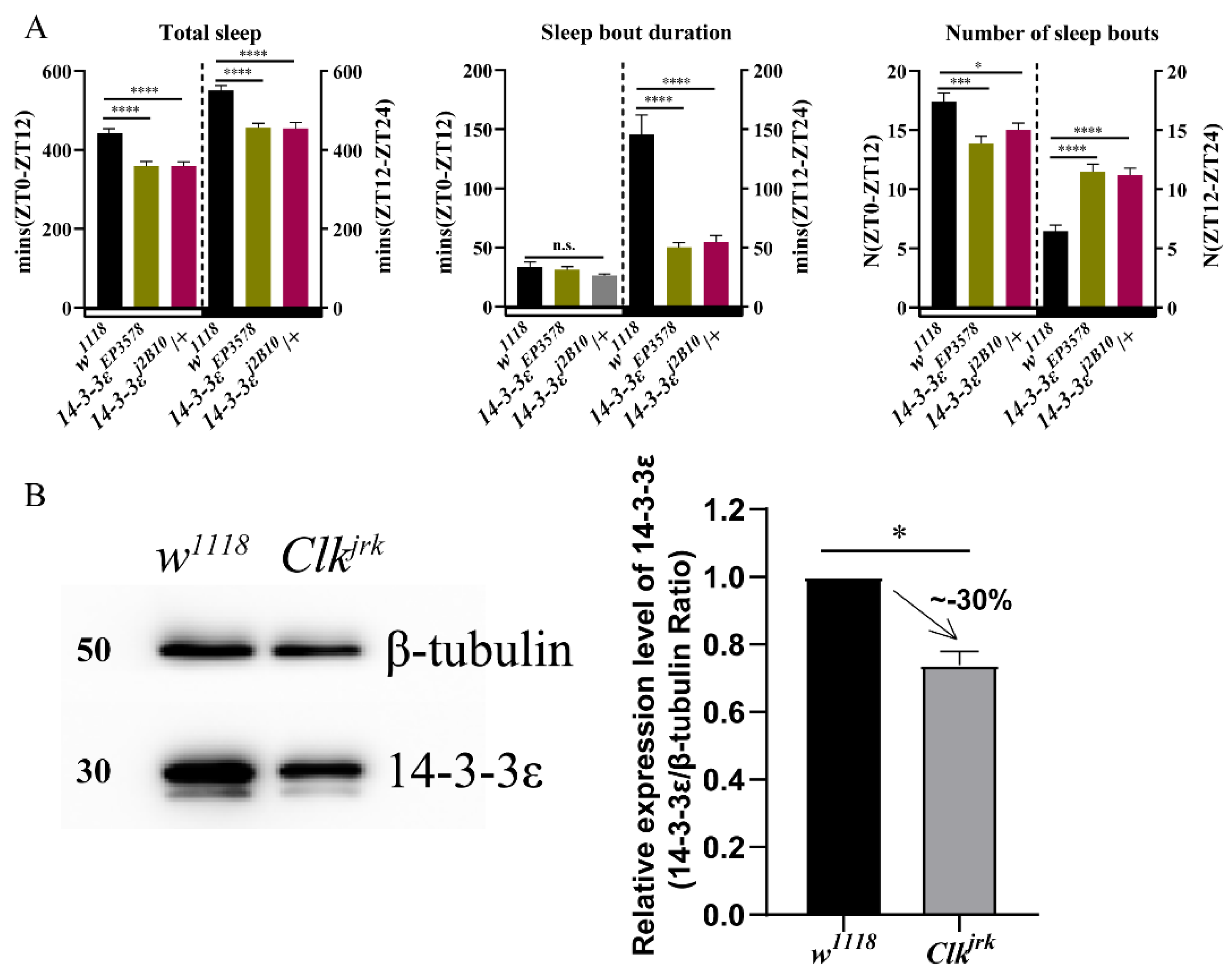
In order to identify potential circadian regulators, we conducted a proteomic screen for oscillating proteins that are differentially expressed in the Drosophila head between the wild-type w1118 and Clock-deficient mutant ClkJrk [27]. Surprisingly, we found that some cancer-related proteins were controlled by Clock, in which some genes were selected from non-phosphorylated and phosphorylated differential proteins between w1118 and ClkJrk. Behavioral analysis of the circadian rhythm and sleep from the mutants of some candidate genes showed that they had anomalous sleep phenotypes (Figure S1), in which we found that the 14-3-3ε regulates sleep independent of the circadian rhythm, with a normal rhythmic percentage when compared to that of control (Figure 1A, Table S1). It is significantly decreased in the ClkJrk mutant detected by the proteomic screen, further verified by using Western blotting (p = 0.023) (Figure 1B). Thus, we focused on its mechanism of sleep regulation in this study.
3.2. 14-3-3ε Regulates Sleep Factor PDF
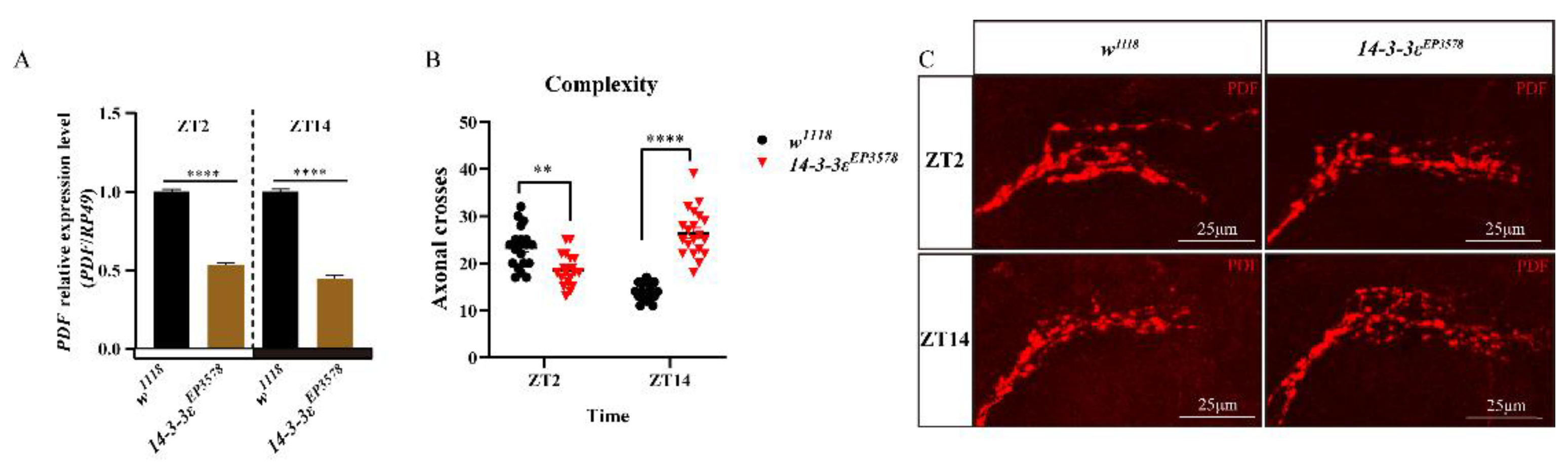
The pigment dispersing factor (PDF), expressed in the LNvs of clock neurons, is a regulatory factor of sleep in Drosophila. We found that the PDF transcription level decreased in the 14-3-3ε mutant by transcriptomic analysis (Table S2), which was further verified by the real-time PCR, with significant decreases of 47% (p < 0.0001) at ZT2 and 55% (p < 0.0001) at ZT14 in the 14-3-3ε mutant flies (14-3-3εEP3578) compared to those in control flies (Figure 2A). Then, we detected the morphology of the PDF-containing sLNv dorsal projections in 14-3-3εEP3578 and w1118 flies, by which PDF signal transmits to the central complexes. The results showed that the morphology of dorsal projections changed greatly, in which the sLNv dorsal termini axonal cross at ZT2 was significantly decreased by 20% (p = 0.0011, n = 20) compared to that of the control, but it was significantly increased by 89% (p < 0.0001, n = 20) at ZT14 (Figure 2B,C). Furthermore, we used the 14-3-3ε protein trap fly line (14-3-3εG00082) fusing 14-3-3ε with GFP (14-3-3ε-GFP) to co-locate the 14-3-3ε and PDF (green for 14-3-3ε by rabbit anti-GFP and red for PDF by mouse anti-PDF). The results showed that both 14-3-3ε and PDF co-expressed in the sLNvs (Figure 3A–C).
Because 14-3-3ε is expressed in the PDF neurons, we specifically downregulated its expression with an RNAi driven by pdf-gal4. The results showed that downregulation of 14-3-3ε recapitulated the sleep phenotype caused by the 14-3-3ε mutant, with significant decreases in the total sleep at daytime compared to that of controls (p < 0.0001 and p = 0.003, respectively) (Figure 3D). These results indicate that 14-3-3ε regulates sleep via PDF in the PDF neurons.
3.3. 14-3-3ε Directly Acts on Ubx to Regulate PDF Transcription
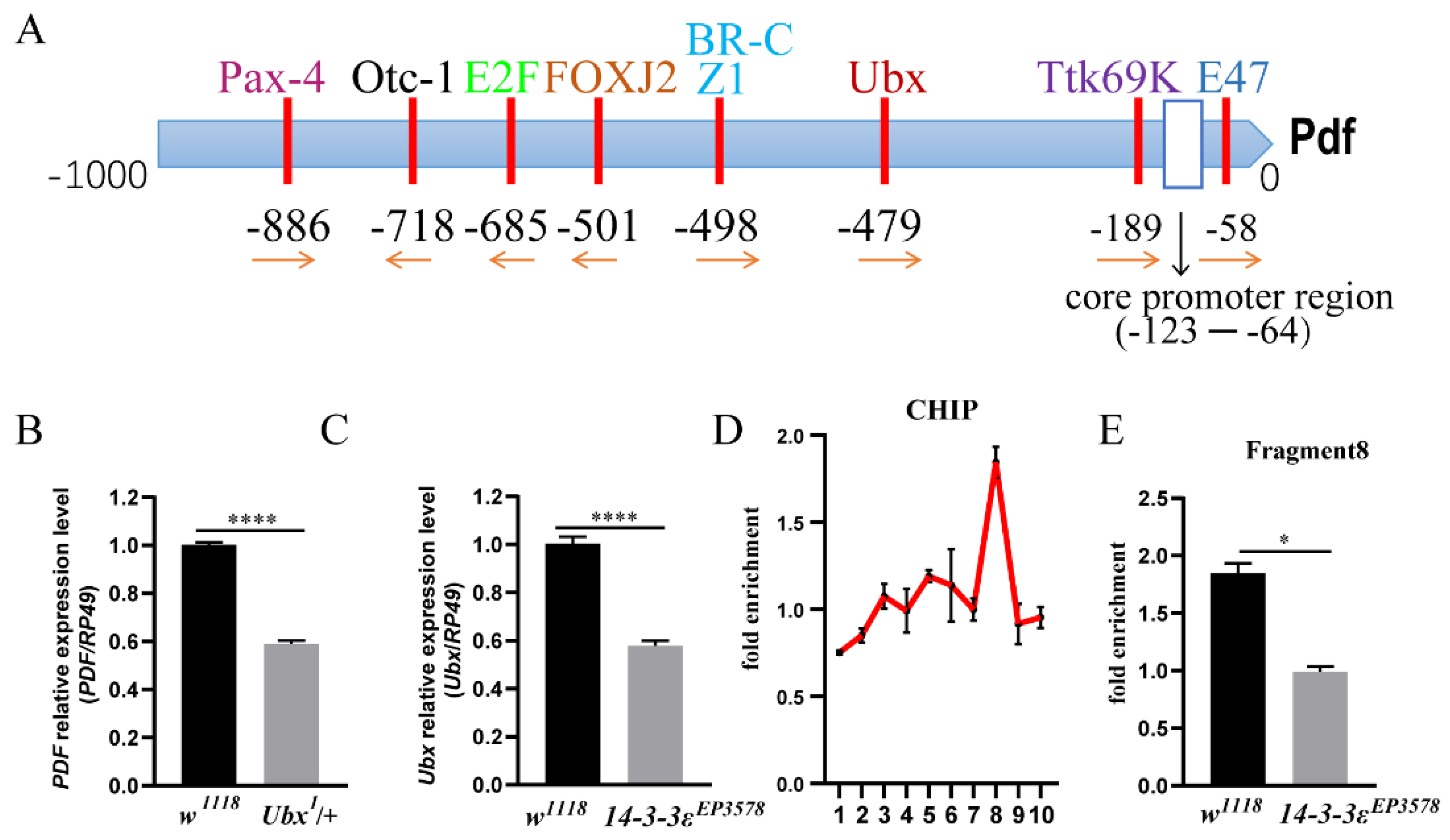
To further determine how 14-3-3ε regulates PDF, we predicted the transcription factors in the promotor of PDF using the website http://gene-regulation.com/index2.html (accessed on 1 October 2020), in which the Ultrabithorax (Ubx) is one of the transcription factors (Figure 4A). When Ubx was downregulated in the Ubx heterozygous mutant (Ubx1/+), the pdf mRNA level significantly decreased by 41% compared to that in the control (p < 0.0001) (Figure 4B). From the transcriptomic data (Table S2), the Ubx level decreased in the 14-3-3ε-deficient mutant, which was further verified by real-time PCR (p < 0.0001) (Figure 4C).
In order to determine the binding sites of Ubx, we designed 10 pairs of PCR primers spanning 2 kb upstream of the translational start sites of PDF (each fragment of which was around 200 bp) for analysis of DNA from ChIP (chromatin immunoprecipitation) with anti-Ubx. The results showed that there was an active peak in fragment 8 (Figure 4D), and the enrichment of this fragment was significantly decreased in the 14-3-3ε mutant (0.99 vs. 1.85 times, p = 0.013) (Figure 4E).
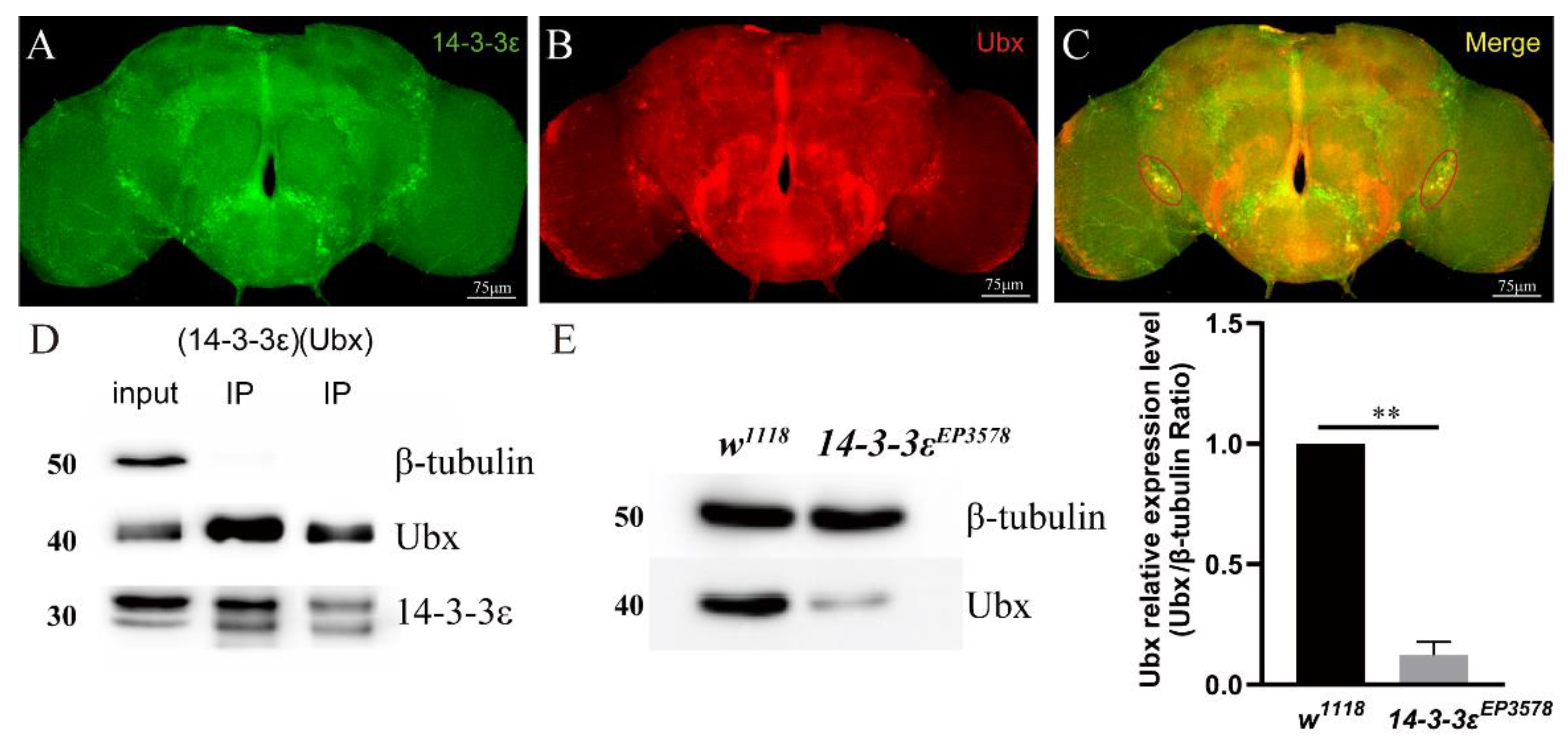
To gain more relationship between 14-3-3ε and Ubx, we co-localized Ubx and 14-3-3ε by using the 14-3-3ε-GFP/+; Ubx-gal4/+ fly lines. The brains were stained with immunofluorescence using anti-GFP and anti-Ubx antibodies. The results showed that 14-3-3ε and Ubx were merged together in PDF neurons (Figure 5A–C). Then, we employed co-immunoprecipitation experiments using anti-Ubx antibody and anti-14-3-3ε antibody, and results revealed that 14-3-3ε was able to directly combine with Ubx (Figure 5D). Furthermore, we quantified the Ubx protein in 14-3-3EP3578 by Western blotting. The evidence showed that Ubx significantly decreased by 88% when compared to control (p = 0.004) (Figure 5E). All these data indicate that Ubx is a direct target of 14-3-3ε, which regulates PDF through activating the fragment 8 of PDF.
3.4. 14-3-3ε Regulates Sleep by Impacting Neurotransmitters
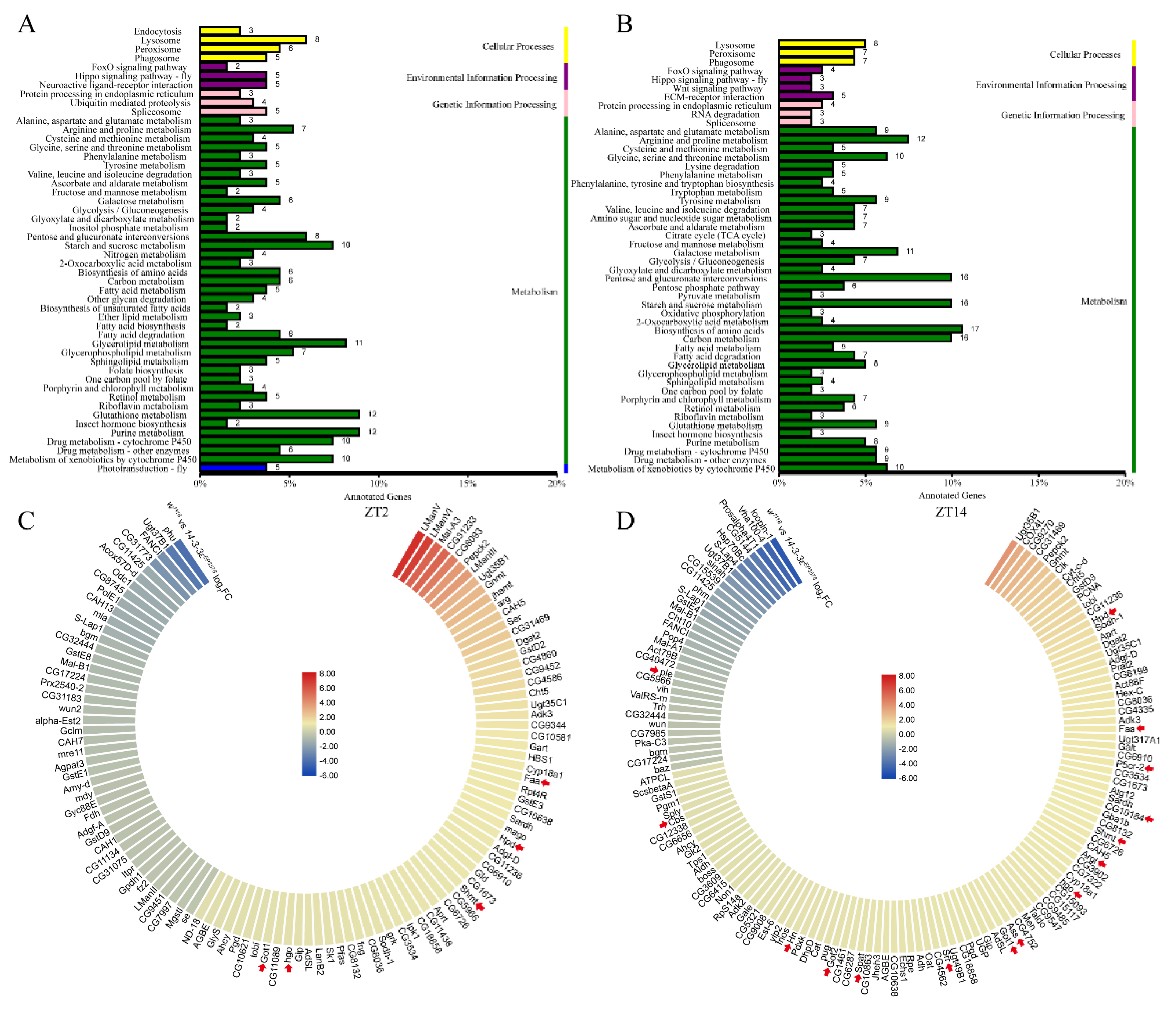
To identify the molecular mechanism of 14-3-3ε on sleep regulation, we performed RNA-seq of the head tissue at ZT2 and ZT14 in both the 14-3-3ε deficient mutant (14-3-3εEP3578) and w1118 control flies (Table S2). The results showed that a number of differentially expressed genes between the mutant and control flies were related to metabolism, including glucometabolism, lipid metabolism, and amino acid metabolism (Figure 6A,B). These were classified into categories in which some of the genes are involved in the tyrosine metabolic process, the amino acid biosynthetic process of the glutamine family, and the amino acid metabolic process of the serine family (arrows in Figure 6C,D). Tyrosine, glutamate, and serine are important precursors for the synthesis of neurotransmitters. Most of them were upregulated at both ZT2 and ZT14 in 14-3-3εEP3578 flies (Table S3). Specially, the differential genes between w1118 and 14-3-3ε mutant flies in tyrosine metabolism were involved in dopamine and octopamine synthesis process, in which 14-3-3ε inhibits the production of dopamine and octopamine.
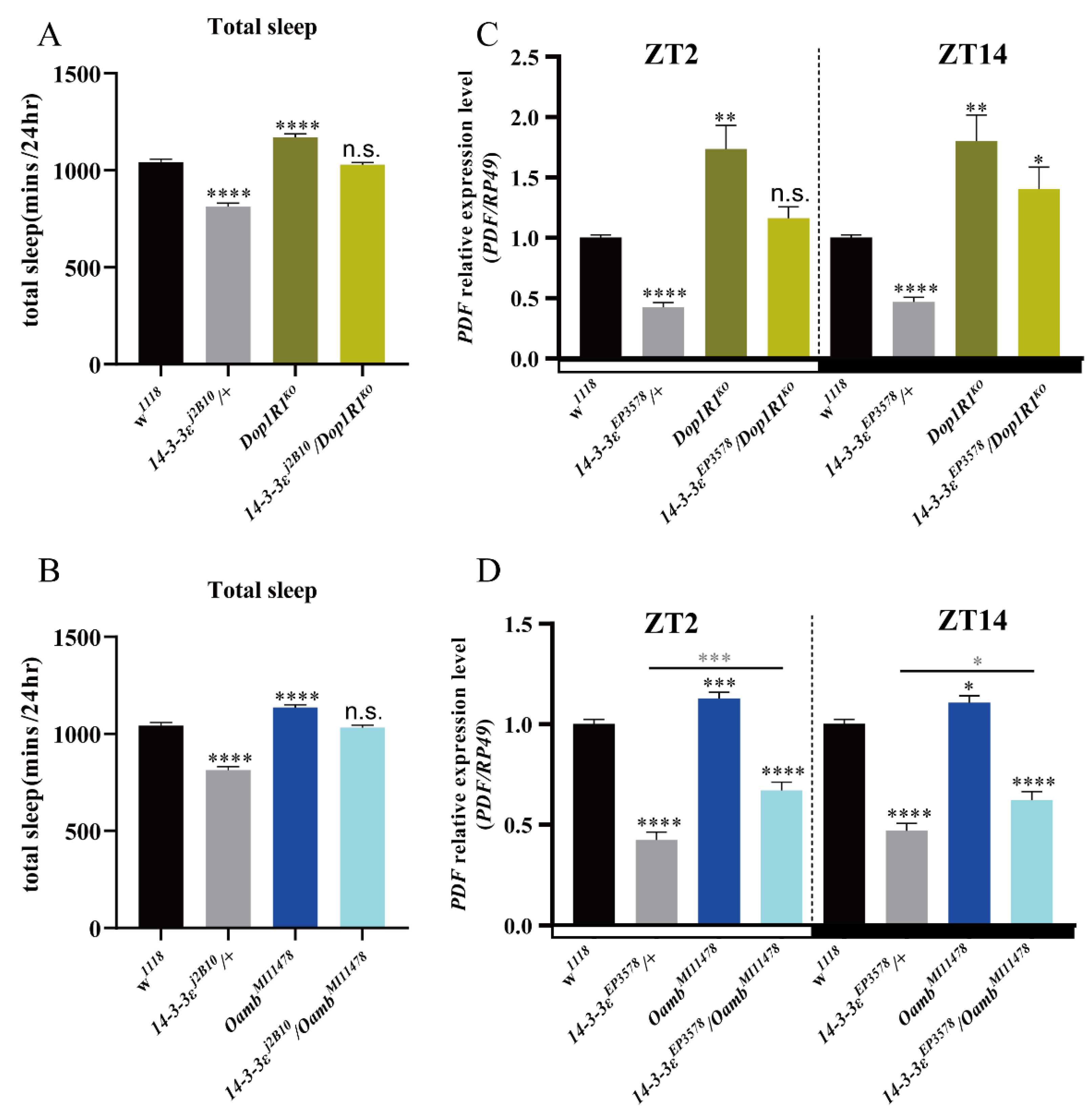
In order to identify whether the sleep phenotypes of the 14-3-3ε mutant are related to these genes, we first measured sleep phenotypes of the 14-3-3ε mutant and receptor mutants of the neurotransmitters Dop1R1 and Oamb, respectively. The results showed that sleep decreased in the 14-3-3ε mutant but increased in the Dop1R1 and Oamb receptor mutants compared to their controls (Figure 7A,B). Furthermore, we examined the genetic interactions between 14-3-3ε and the Dop1R1 or Oamb receptor by using the flies of simultaneously mutating 14-3-3ε and Dop1R1 (Dop1R1KO /14-3-3εj2B10) or 14-3-3ε and Oamb (OambM111578/14-3-3εj2B10). The results showed that decreases in sleep phenotype caused by 14-3-3εj2B10/+ could be partially recovered in these double-mutant flies (Figure 7A,B).
Does 14-3-3ε regulate PDF by these neurotransmitters? To answer this question, we analyzed the relationship by detecting PDF levels in the double-mutant flies of 14-3-3ε and Dop1R1 (Dop1R1KO/14-3-3εEP3578) or 14-3-3ε and Oamb (OambM111578/14-3-3εEP3578). The results showed that decreases in PDF level caused by 14-3-3ε mutant flies could be partially recovered in these double-mutant flies (Figure 7C,D), which is a similar finding to that regarding the sleep behaviors presented in Figure 7A,B. These results indicate that the sleep phenotypes of the 14-3-3ε mutant are related to these neurotransmitters.
From all of the above data, we propose a model for 14-3-3ε sleep regulation. The 14-3-3ε protein regulates sleep through two pathways: one is achieved by regulating PDF pathway through interacting with Ubx, which results in a negative regulation of sleep; on the other hand, 14-3-3ε regulates the synthesis enzymes of the neurotransmitters, which results in positive regulation of sleep. As a result, 14-3-3ε integrates these factors to maintain a sleep balance (Figure 8).
4. Discussion
The pigment dispersing factor (PDF), a neuropeptide secreted from the LNv neurons of the brain, is a wake-promoting factor. When flies are stimulated by light, LNvs respond to light and promote arousal by releasing PDF [28,29]. Loss of PDF leads to an increase in the amount of sleep in Drosophila [30]. Functionally, it is analogous to vertebrate orexin/hypocretin [30,31,32]. In mammals, the neuropeptide vasoactive intestinal peptide (VIP) functions to synchronize the oscillations of clock neurons and transfer circadian signals to downstream neurons [33,34].
Ultrabithorax (Ubx) encodes a homeodomain transcription factor involved in cell fate decisions, cell proliferation, and organ identity, and it belongs to the Hox gene family. Hox genes, including Sex-combs reduced (Scr), Antennapedia (Antp), Ultrabithorax (Ubx) and abdominal-A (abd-A), play a conserved role in establishing the thoracic and abdominal segments during insect embryogenesis [35]. Singh et al. reported that Ubx regulates the Fat/Hippo and IIS/dAkt pathways in specifying haltere development, including organ decision and size, sensory bristle repression, trichome morphology, and arrangement. The Ubx-mediated Fat/Hippo pathway is key for the transformation of wing identity to haltere [36]. Ubx functions as a tumor inhibitor in its respective endogenous domains [37]. When interacting with Pho, Ubx can stabilize lineage choice through suppressing the multipotency encoded in the genome [38]. Regulated by polycomb complex, Ubx is a repressor of alternative cell fates within the mesoderm, and it also maintains normal muscle differentiation by repressing Twi [39]. In this study, Ubx takes part in regulating fly sleep by cooperating with 14-3-3ε.
In this study, we identified the role of 14-3-3ε in sleep regulation. 14-3-3ε, controlled by Clock, regulates both PDF and metabolic factors that are important for neurotransmitter biogenesis. Previous studies showed that multiple types of neurotransmitters had been identified, including acetylcholine (Ach), noradrenaline (NA), histamine, 5-hydroxytryptophan (5-HT), dopamine (DA), glutamate (Glu), and γ-aminobutyric acid (GABA) [22]. The production of many neurotransmitters is closely related to amino acid production. Glutamate (glutamic acid) is a natural amino acid, while GABA (γ-amino butyric acid) is derived from glutamate. Serotonin (also called 5-HT), dopamine, noradrenaline, and histamine are derived from aromatic amino acids like tyrosine and belong to the monoamine neurotransmitters. Our data from the RNA-seq indicate that multiple factors in the tyrosine and glutamate metabolic process are affected in the 14-3-3ε mutant, among which dopamine and octopamine have been proved to be sleep regulators by previous studies in Drosophila. Thus, 14-3-3ε was found to be a novel regulator of neurotransmitters in this study, in which the mutants from the Dop1R1 and Oamb can restore the 14-3-3ε phenotype to different degrees. In this study, a new sleep regulation pathway, i.e., 14-3-3ε, that regulates sleep through the dopamine and octopamine signal pathway, was identified.
14-3-3 proteins are found to be important in both cancer- and age-related neurodegenerative disease [40], which can directly interact with yki [17], an important cancer factor in the Hippo pathway [19,40]. Previous studies indicated that 14-3-3ε is involved in gastric cancer and colorectal cancer [41,42,43,44,45,46], and disrupted sleep is a risk factor that contributes to cancer [47]. Many current papers showed a link between molecules upregulated in cancer patients and selected sleep disturbances. The obstructive sleep apnea (OSA) patients have less sleep and worse sleep quality, in which the serum hypoxia-inducible factor 1α (HIF-1α) protein level as a key factor of cellular oxygen metabolism is significantly higher [48,49,50,51,52]. Hypoxia is regarded as a feature of rapidly proliferating tissues, such as cancer [53]. HIF-1αof dysregulation/overexpression have been connected to both obstructive sleep apnea and cancer biology, specifically in areas of vascularization and angiogenesis, energy metabolism, cell survival, tumor invasion, and so on [48,49,50,51]. This link is interesting because disruption of HIF-1α expression may lead to a developing circadian clock disruption, as its increased protein level is associated with overexpression of circadian clock proteins [54].
As is well known, the clock genes regulate the circadian rhythm or/and sleep [2]. In this current study, we found that 14-3-3ε, controlled by Clock, regulates sleep through pathways of both the 14-3-3ε/Ubx/PDF and neurotransmitters. In addition, there are previous reports that 14-3-3ε is also related to cancers [40,41,42,43,44,45,46], indicating that 14-3-3ε is a multifunctional gene in Drosophila regulating different physiological activities. Currently, the direct relationship between cancer and sleep is still unclear, which need to be carefully designed for systematical investigation in future.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ijms22189748/s1.
Author Contributions
Z.Z. designed the research; Y.W. performed the research; Y.W. and J.D. analyzed the data; Y.W., J.D. and Z.Z. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 31970458, 31572317 to Z.Z. and 31772535 to J.D.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The RNA-seq data from this publication have been deposited in the NCBI bioproject database (https://www.ncbi.nlm.nih.gov/bioproject/) (accessed on 1 May 2021) and assigned the identifier PRJNA680635.
Acknowledgments
We thank Jeffrey Price (University of Missouri at Kansas City) for revision of this manuscript.
Conflicts of Interest
The authors declare no conflict of interests.
References
- Deboer, T. Sleep homeostasis and the circadian clock: Do the circadian pacemaker and the sleep homeostat influence each other’s functioning? Neurobiol. Sleep Circadian Rhythms 2018, 5, 68–77. [Google Scholar] [CrossRef]
- Franken, P.; Dijk, D.-J. Circadian clock genes and sleep homeostasis. Eur. J. Neurosci. 2009, 29, 1820–1829. [Google Scholar] [CrossRef]
- Szmyd, B.; Rogut, M.; Białasiewicz, P.; Gabryelska, A. The impact of glucocorticoids and statins on sleep quality. Sleep Med. Rev. 2020, 55, 101380. [Google Scholar] [CrossRef]
- Allada, R.; Siegel, J.M. Unearthing the Phylogenetic Roots of Sleep. Curr. Biol. 2008, 18, R670–R679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendricks, J.C.; Finn, S.M.; Panckeri, K.A.; Chavkin, J.; Williams, J.A.; Sehgal, A.; Pack, A. Rest in Drosophila Is a Sleep-like State. Neuron 2000, 25, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Shaw, P.J.; Cirelli, C.; Greenspan, R.J.; Tononi, G. Correlates of Sleep and Waking in Drosophila melanogaster. Science 2000, 287, 1834–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, B.Y.; Kilman, V.L.; Keath, J.R.; Pitman, J.L.; Allada, R. The GABA(A) receptor RDL acts in peptidergic PDF neurons to promote sleep in Drosophila. Curr. Biol. 2009, 19, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Cerullo, I.; Chen, X.; Rosbash, M. PDF neuron firing phase-shifts key circadian activity neurons in Drosophila. Elife 2014, 3, e02780. [Google Scholar] [CrossRef] [PubMed]
- Heekeren, H.; Marrett, S.; Bandettini, P.A.; Ungerleider, L.G. A general mechanism for perceptual decision-making in the human brain. Nature 2004, 431, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Shafer, O.T. The Drosophila Circadian Clock Is a Variably Coupled Network of Multiple Peptidergic Units. Science 2014, 343, 1516–1520. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Yu, J.; Jung, H.J.; Abruzzi, K.C.; Luo, W.; Griffith, L.C.; Rosbash, M. Circadian neuron feedback controls the Dro-sophila sleep-activity profile. Nature 2016, 536, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.D.L.P.; Berni, J.; Ceriani, M.F. Circadian Remodeling of Neuronal Circuits Involved in Rhythmic Behavior. PLoS Biol. 2008, 6, e69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Du, J.; Wei, L.; Zhao, Z. AKH-FOXO pathway regulates starvation-induced sleep loss through remodeling of the small ventral lateral neuron dorsal projections. PLoS Genet. 2020, 16, e1009181. [Google Scholar] [CrossRef]
- Nian, X.; Chen, W.; Bai, W.; Zhao, Z.; Zhang, Y. miR-263b Controls Circadian Behavior and the Structural Plasticity of Pacemaker Neurons by Regulating the LIM-Only Protein Beadex. Cells 2019, 8, 923. [Google Scholar] [CrossRef] [Green Version]
- Morrison, D.K. The 14–3-3 proteins: Integrators of diverse signaling cues that impact cell fate and cancer development. Trends Cell Biol. 2009, 19, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14–3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Pan, D. The Hippo Signaling Pathway in Development and Cancer. Dev. Cell 2010, 19, 491–505. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Li, L.; Lei, Q.-Y.; Guan, K.-L. The Hippo-YAP pathway in organ size control and tumorigenesis: An updated version. Genes. Dev. 2010, 24, 862–874. [Google Scholar] [CrossRef] [Green Version]
- Ren, F.; Zhang, L.; Jiang, J. Hippo signaling regulates Yorkie nuclear localization and activity through 14–3-3 dependent and independent mechanisms. Dev. Biol. 2010, 337, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, T.; Isobe, T.; Okuyama, T.; Yamauchi, T.; Fujisawa, H. Brain 14–3-3 Protein Is an Activator Protein That Activates Tryptophan 5-Monooxygenase and Tyrosine 3-Monooxygenase in the Presence of Ca-2+, Calmodulin-Dependent Protein Ki-nase-Ii. FEBS Lett. 1987, 219, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, Y. 14–3-3 Proteins in Glutamatergic Synapses. Neural Plast. 2018, 2018, 1–6. [Google Scholar] [CrossRef]
- Deng, B.; Li, Q.; Liu, X.; Cao, Y.; Li, B.; Qian, Y.; Xu, R.; Mao, R.; Zhou, E.; Zhang, W.; et al. Chemoconnectomics: Mapping Chemical Transmission in Drosophila. Neuron 2019, 101, 876–893.e4. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Shi, W.; Li, L.; Zheng, Z.; Li, T.; Bai, W.; Zhao, Z. Regulation of sleep by the short neuropeptide F (sNPF) in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2013, 43, 809–819. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, F.; Liu, J.; Wang, D.; Yan, Y.; Ji, S.; Zan, J.; Zhou, J. The co-chaperone Cdc37 regulates the rabies virus phospho-protein stability by targeting to Hsp90AA1 machinery. Sci. Rep. 2016, 6, 27123. [Google Scholar] [CrossRef]
- Liu, X.; Li, H.; Liu, Q.; Niu, Y.; Hu, Q.; Deng, H.; Cha, J.; Wang, Y.; Liu, Y.; He, Q. Role for Protein Kinase A in the Neurospora Circadian Clock by Regulating White Collar-Independent frequency Transcription through Phosphorylation of RCM-1. Mol. Cell. Biol. 2015, 35, 2088–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Zhang, Y.F.; Xue, Y.B.; Zhao, X.Y.; Zhao, X.G.; Wei, Y.; Li, Z.; Zhang, Y.; Zhao, Z.W. Diurnal protein oscillation profiles in Drosophila head. FEBS Lett. 2018, 592, 3736–3749. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Haynes, P.; Pírez, N.; Harrington, K.; Guo, F.; Pollack, J.; Hong, P.; Griffith, L.; Rosbash, M. Imaging analysis of clock neurons reveals light buffers the wake-promoting effect of dopamine. Nat. Neurosci. 2011, 14, 889–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Griffith, L.; Rosbash, M. Light-arousal and circadian photoreception circuits intersect at the large PDF cells of the Drosophila brain. Proc. Natl. Acad. Sci. USA 2008, 105, 19587–19594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisky, K.M.; Agosto, J.; Pulver, S.R.; Shang, Y.; Kuklin, E.; Hodge, J.J.; Kang, K.; Liu, X.; Garrity, P.A.; Rosbash, M.; et al. PDF cells are a GABA-responsive wake-promoting component of the Drosophila sleep circuit. Neuron 2008, 60, 672–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Mignot, E. Genetics of Sleep and Sleep Disorders. Cell 2011, 146, 194–207. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, T.; Crocker, A.; McCormack, S.; Yanagisawa, M.; Sakurai, T.; Scammell, T.E. Behavioral State Instability in Orexin Knock-Out Mice. J. Neurosci. 2004, 24, 6291–6300. [Google Scholar] [CrossRef]
- Aton, S.J.; Colwell, C.S.; Harmar, A.J.; Waschek, J.; Herzog, E.D. Vasoactive intestinal polypeptide mediates circadian rhyth-micity and synchrony in mammalian clock neurons. Nat. Neurosci. 2005, 8, 476–483. [Google Scholar] [CrossRef]
- Vosko, A.M.; Schroeder, A.; Loh, D.; Colwell, C.S. Vasoactive intestinal peptide and the mammalian circadian system. Gen. Comp. Endocrinol. 2007, 152, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.L.; Kaufman, T.C. Hox genes and the evolution of the arthropod body plan1. Evol. Dev. 2002, 4, 459–499. [Google Scholar] [CrossRef]
- Singh, S.; Sanchez-Herrero, E.; Shashidhara, L.S. Critical role for Fat/Hippo and IIS/Akt pathways downstream of Ultrabi-thorax during haltere specification in Drosophila. Mech. Dev. 2015, 138, 198–209. [Google Scholar] [CrossRef]
- Gupta, R.P.; Bajpai, A.; Sinha, P. Selector genes display tumor cooperation and inhibition in Drosophila epithelium in a de-velopmental context-dependent manner. Biol. Open 2017, 6, 1581–1591. [Google Scholar] [CrossRef] [Green Version]
- Domsch, K.; Carnesecchi, J.; Disela, V.; Friedrich, J.; Trost, N.; Ermakova, O.; Polychronidou, M.; Lohmann, I. The Hox tran-scription factor Ubx stabilizes lineage commitment by suppressing cellular plasticity in Drosophila. Elife 2019, 8, e42675. [Google Scholar] [CrossRef] [PubMed]
- Domsch, K.; Schroder, J.; Janeschik, M.; Schaub, C.; Lohmann, I. The Hox Transcription Factor Ubx Ensures Somatic Myo-genesis by Suppressing the Mesodermal Master Regulator Twist. Cell Rep. 2021, 34, 108577. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Cui, L.; Zeng, Y.; Song, W.; Gaur, U.; Yang, M. 14–3-3 Proteins Are on the Crossroads of Cancer, Aging, and Age-Related Neurodegenerative Disease. Int. J. Mol. Sci. 2019, 20, 3518. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Huang, H.R.; Li, W.D.; Jin, X.J.; Zeng, J.; Liu, Y.W.; Gu, Y.; Sun, X.G.; Wen, G.; Ding, Y.Q.; et al. Nuclear local-ization of 14–3-3epsilon inversely correlates with poor long-term survival of patients with colorectal cancer. J. Surg. Oncol. 2012, 106, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Gu, H.; Li, J.; Xu, M.; Liu, T.; Shen, Y.; Chen, B.; Zhang, G. RKIP and 14–3-3ε Exert an Opposite Effect on Human Gastric Cancer Cells SGC7901 by Regulating the ERK/MAPK Pathway Differently. Dig. Dis. Sci. 2012, 58, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Yan, L.; Gu, H.; Mu, Y.; Tong, G.; Zhang, G. 14-3-3ε functions as an oncogene in SGC7901 gastric cancer cells through involvement of cyclin E and p27kip1. Mol. Med. Rep. 2014, 10, 3145–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Fang, X.; Fang, H.; Feng, Y.; Chen, F.; Xia, Q. ATPR-induced G(0)/G(1) phase arrest in gastric cancer cells by regulating the binding of 14-3-3ε and filamin A. Cancer Med. 2018, 7, 3373–3384. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.Y.; Ghelani, D.; Yeh, S.; Wu, K.K. Nonsteroidal anti-inflammatory drugs induce colorectal cancer cell apoptosis by suppressing 14-3-3epsilon. Cancer Res. 2007, 67, 3185–3191. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.K.; Liou, J.Y. Cyclooxygenase Inhibitors Induce Colon Cancer Cell Apoptosis Via PPAR delta -> 14-3-3 epsilon Pathway. Methods Mol. Biol. 2009, 512, 295–307. [Google Scholar]
- Soucise, A.; Vaughn, C.; Thompson, C.L.; Millen, A.E.; Freudenheim, J.L.; Wactawski-Wende, J.; Phipps, A.I.; Hale, L.; Qi, L.H.; Ochs-Balcom, H.M. Sleep quality, duration, and breast cancer aggressiveness. Breast Cancer Res. Tr. 2017, 164, 169–178. [Google Scholar] [CrossRef]
- Gabryelska, A.; Szmyd, B.; Szemraj, J.; Stawski, R.; Sochal, M.; Białasiewicz, P. Patients with obstructive sleep apnea present with chronic upregulation of serum HIF-1α protein. J. Clin. Sleep Med. 2020, 16, 1761–1768. [Google Scholar] [CrossRef]
- Gabryelska, A.; Szmyd, B.; Panek, M.; Szemraj, J.; Kuna, P.; Białasiewicz, P. Serum Hypoxia-Inducible Factor-1α protein level as a diagnostic marker of obstructive sleep apnea. Pol. Arch. Intern. Med. 2019, 130, 158–160. [Google Scholar] [CrossRef] [Green Version]
- Gabryelska, A.; Stawski, R.; Sochal, M.; Szmyd, B.; Białasiewicz, P. Influence of one-night CPAP therapy on the changes of HIF-1α protein in OSA patients: A pilot study. J. Sleep Res. 2020, 29, e12995. [Google Scholar] [CrossRef]
- Gabryelska, A.; Karuga, F.F.; Szmyd, B.; Białasiewicz, P. HIF-1α as a Mediator of Insulin Resistance, T2DM, and Its Compli-cations: Potential Links With Obstructive Sleep Apnea. Front. Physiol. 2020, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Li, N.; Yao, X.; Zhou, L. Potential inflammatory markers in obstructive sleep apnea-hypopnea syndrome. Bosn. J. Basic Med. Sci. 2016, 17, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Gabryelska, A.; Sochal, M.; Turkiewicz, S.; Białasiewicz, P. Relationship between HIF-1 and Circadian Clock Proteins in Ob-structive Sleep Apnea Patients-Preliminary Study. J. Clin. Med. 2020, 9, 1599. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Sleep and the circadian locomotor rhythm in 14-3-3ε mutants: (A), sleep pattern of 14-3-3ε mutants. (B), 14-3-3ε protein expression level in Clock mutant (ClkJrk) and w1118 control flies by Western blotting. Bar graphs are presented as mean ± SEM. Statistical differences were measured using unpaired Student’s t-test; n.s. indicates no significant difference, * p < 0.05, *** p < 0.001, **** p < 0.0001. Each experiment was conducted in triplicate.
Figure 1.
Sleep and the circadian locomotor rhythm in 14-3-3ε mutants: (A), sleep pattern of 14-3-3ε mutants. (B), 14-3-3ε protein expression level in Clock mutant (ClkJrk) and w1118 control flies by Western blotting. Bar graphs are presented as mean ± SEM. Statistical differences were measured using unpaired Student’s t-test; n.s. indicates no significant difference, * p < 0.05, *** p < 0.001, **** p < 0.0001. Each experiment was conducted in triplicate.

Figure 2.
Effects of the 14-3-3ε mutant on PDF expression and sLNv dorsal projections: (A), PDF mRNA expression level in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies at ZT2 and ZT14 by real-time PCR. (B), sLNv dorsal termini axonal cross in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies at ZT2 and ZT14. (C), sLNv dorsal projections in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies at ZT2 and ZT14 by immunofluorescence using PDF antibody (red). The scale bar indicates 25 μm. Bar graphs are presented as mean ± SEM. A, statistical differences were measured using unpaired Student’s t-test. (B), statistical differences were measured using Two-way ANOVA and Tukey’s multiple comparison test; ** p < 0.01, **** p < 0.0001. Each experiment was conducted in triplicate.
Figure 2.
Effects of the 14-3-3ε mutant on PDF expression and sLNv dorsal projections: (A), PDF mRNA expression level in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies at ZT2 and ZT14 by real-time PCR. (B), sLNv dorsal termini axonal cross in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies at ZT2 and ZT14. (C), sLNv dorsal projections in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies at ZT2 and ZT14 by immunofluorescence using PDF antibody (red). The scale bar indicates 25 μm. Bar graphs are presented as mean ± SEM. A, statistical differences were measured using unpaired Student’s t-test. (B), statistical differences were measured using Two-way ANOVA and Tukey’s multiple comparison test; ** p < 0.01, **** p < 0.0001. Each experiment was conducted in triplicate.

Figure 3.
14-3-3ε function in the PDF neuron for sleep regulation: (A–C), expression pattern of 14-3-3ε in the adult fly brain by immunofluorescence. 14-3-3ε protein trap strain (14-3-3εG00082, BS51385) fused with GFP (14-3-3ε-GFP), rabbit anti-GFP (14-3-3ε, 1:400, green), and mouse anti-PDF (PDF, 1:200, red). (D), sleep pattern with 14-3-3ε downregulation in PDF neuron. The scale bar indicates 75 μm. Statistical differences were measured using One-way ANOVA and Tukey’s multiple comparison test and unpaired Student’s t-test; n.s. indicates no significant difference, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001. Each experiment was conducted in triplicate.
Figure 3.
14-3-3ε function in the PDF neuron for sleep regulation: (A–C), expression pattern of 14-3-3ε in the adult fly brain by immunofluorescence. 14-3-3ε protein trap strain (14-3-3εG00082, BS51385) fused with GFP (14-3-3ε-GFP), rabbit anti-GFP (14-3-3ε, 1:400, green), and mouse anti-PDF (PDF, 1:200, red). (D), sleep pattern with 14-3-3ε downregulation in PDF neuron. The scale bar indicates 75 μm. Statistical differences were measured using One-way ANOVA and Tukey’s multiple comparison test and unpaired Student’s t-test; n.s. indicates no significant difference, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001. Each experiment was conducted in triplicate.

Figure 4.
Ubx is a transcription factor of PDF: (A), Pdf transcription factor prediction. (B), PDF mRNA expression level in Ubx mutant (Ubx1/+) and w1118 control flies by real-time PCR. (C), Ubx mRNA expression level in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies by real-time PCR. (D), chromatin immunoprecipitation with Ubx antibody and w1118 adult heads as the sample. X-axis shows fragment numbers. (E), chromatin immunoprecipitation used the Ubx antibody, the 14-3-3ε mutant (14-3-3εEP3578), and w1118 control adult heads as samples. Fragment 8 was tested by CHIP-qPCR. Bar graphs are presented as mean ± SEM. Statistical differences were measured using unpaired Student’s t-test; n.s. indicates no significant difference, * p < 0.05, **** p < 0.0001. Each experiment was conducted in triplicate.
Figure 4.
Ubx is a transcription factor of PDF: (A), Pdf transcription factor prediction. (B), PDF mRNA expression level in Ubx mutant (Ubx1/+) and w1118 control flies by real-time PCR. (C), Ubx mRNA expression level in 14-3-3ε mutant (14-3-3εEP3578) and w1118 control flies by real-time PCR. (D), chromatin immunoprecipitation with Ubx antibody and w1118 adult heads as the sample. X-axis shows fragment numbers. (E), chromatin immunoprecipitation used the Ubx antibody, the 14-3-3ε mutant (14-3-3εEP3578), and w1118 control adult heads as samples. Fragment 8 was tested by CHIP-qPCR. Bar graphs are presented as mean ± SEM. Statistical differences were measured using unpaired Student’s t-test; n.s. indicates no significant difference, * p < 0.05, **** p < 0.0001. Each experiment was conducted in triplicate.

Figure 5.
14-3-3ε interacts with Ubx: (A–C), immunofluorescence colocalization between 14-3-3ε and Ubx with mouse anti-RFP (Ubx, 1:100, red) and rabbit anti-GFP (14-3-3ε, 1:200, green) in UAS-mRFP/+; Ubx-gal4/+, 14-3-3ε-GFP/+ adult fly brain. (D), co-immunoprecipitation between 14-3-3ε and Ubx. First well, input. Second well, anti-14-3-3ε antibody for IP. Third well, anti-Ubx antibody for IP. w1118 adult fly heads were used as the sample. (E), Ubx protein expression level in 14-3-3εEP3578 and w1118 by Western blotting. The scale bar (right bottom white line) indicates 75 μm. Bar graphs are presented as mean ± SEM. Statistical differences were measured using unpaired Student’s t-test; n.s. indicates no significant difference, ** p < 0.01. Each experiment was conducted in triplicate.
Figure 5.
14-3-3ε interacts with Ubx: (A–C), immunofluorescence colocalization between 14-3-3ε and Ubx with mouse anti-RFP (Ubx, 1:100, red) and rabbit anti-GFP (14-3-3ε, 1:200, green) in UAS-mRFP/+; Ubx-gal4/+, 14-3-3ε-GFP/+ adult fly brain. (D), co-immunoprecipitation between 14-3-3ε and Ubx. First well, input. Second well, anti-14-3-3ε antibody for IP. Third well, anti-Ubx antibody for IP. w1118 adult fly heads were used as the sample. (E), Ubx protein expression level in 14-3-3εEP3578 and w1118 by Western blotting. The scale bar (right bottom white line) indicates 75 μm. Bar graphs are presented as mean ± SEM. Statistical differences were measured using unpaired Student’s t-test; n.s. indicates no significant difference, ** p < 0.01. Each experiment was conducted in triplicate.

Figure 6.
Neurotransmitter metabolism is changed when 14-3-3ε gene is mutated: (A,B), KEGG enrichment analysis of different genes in 14-3-3ε mutant (14-3-3εEP3578) transcriptome at ZT2 and ZT14. (C,D), the heatmaps of different genes relevant to metabolism from the transcriptome at ZT2 and ZT14. Red arrow displays differences for neurotransmitter metabolism genes.
Figure 6.
Neurotransmitter metabolism is changed when 14-3-3ε gene is mutated: (A,B), KEGG enrichment analysis of different genes in 14-3-3ε mutant (14-3-3εEP3578) transcriptome at ZT2 and ZT14. (C,D), the heatmaps of different genes relevant to metabolism from the transcriptome at ZT2 and ZT14. Red arrow displays differences for neurotransmitter metabolism genes.

Figure 7.
Neurotransmitter mutants rescue sleep loss induced by 14-3-3ε: (A), total sleep of 14-3-3ε mutant as a double mutant with Dop1R1. (B), total sleep of 14-3-3ε mutant as a double mutant with Oamb. (C), PDF mRNA expression level of 14-3-3ε mutant as a double mutant with Dop1R1 by real-time PCR. (D), PDF mRNA expression level of 14-3-3ε mutant as a double mutant with Oamb by real-time PCR. Black star, compared with w1118 control; gray star, compared with 14-3-3εEP3578/+. Bar graphs are presented as mean ± SEM. (A,B): statistical differences were measured using One-way ANOVA and Tukey’s multiple comparison test. (C,D): statistical differences were measured using unpaired Student’s t-test. n.s. indicates no significant difference, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p< 0.0001. Each experiment was conducted in triplicate.
Figure 7.
Neurotransmitter mutants rescue sleep loss induced by 14-3-3ε: (A), total sleep of 14-3-3ε mutant as a double mutant with Dop1R1. (B), total sleep of 14-3-3ε mutant as a double mutant with Oamb. (C), PDF mRNA expression level of 14-3-3ε mutant as a double mutant with Dop1R1 by real-time PCR. (D), PDF mRNA expression level of 14-3-3ε mutant as a double mutant with Oamb by real-time PCR. Black star, compared with w1118 control; gray star, compared with 14-3-3εEP3578/+. Bar graphs are presented as mean ± SEM. (A,B): statistical differences were measured using One-way ANOVA and Tukey’s multiple comparison test. (C,D): statistical differences were measured using unpaired Student’s t-test. n.s. indicates no significant difference, * p < 0.05, ** p < 0.01, *** p < 0.001, **** p< 0.0001. Each experiment was conducted in triplicate.

Figure 8.
The 14-3-3ε mediated sleep regulation in Drosophila: A model shows that 14-3-3ε regulates sleep through Ubx-PDF pathways and neurotransmitters.
Figure 8.
The 14-3-3ε mediated sleep regulation in Drosophila: A model shows that 14-3-3ε regulates sleep through Ubx-PDF pathways and neurotransmitters.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wei, Y.; Du, J.; Zhao, Z. Integrative Role of 14-3-3ε in Sleep Regulation. Int. J. Mol. Sci. 2021, 22, 9748. https://doi.org/10.3390/ijms22189748
AMA Style
Wei Y, Du J, Zhao Z. Integrative Role of 14-3-3ε in Sleep Regulation. International Journal of Molecular Sciences. 2021; 22(18):9748. https://doi.org/10.3390/ijms22189748
Chicago/Turabian StyleWei, Yu, Juan Du, and Zhangwu Zhao. 2021. "Integrative Role of 14-3-3ε in Sleep Regulation" International Journal of Molecular Sciences 22, no. 18: 9748. https://doi.org/10.3390/ijms22189748
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.