
The Role of lncRNAs in Rare Tumors with a Focus on HOX Transcript Antisense RNA (HOTAIR)

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. LncRNAs in Rare Tumors

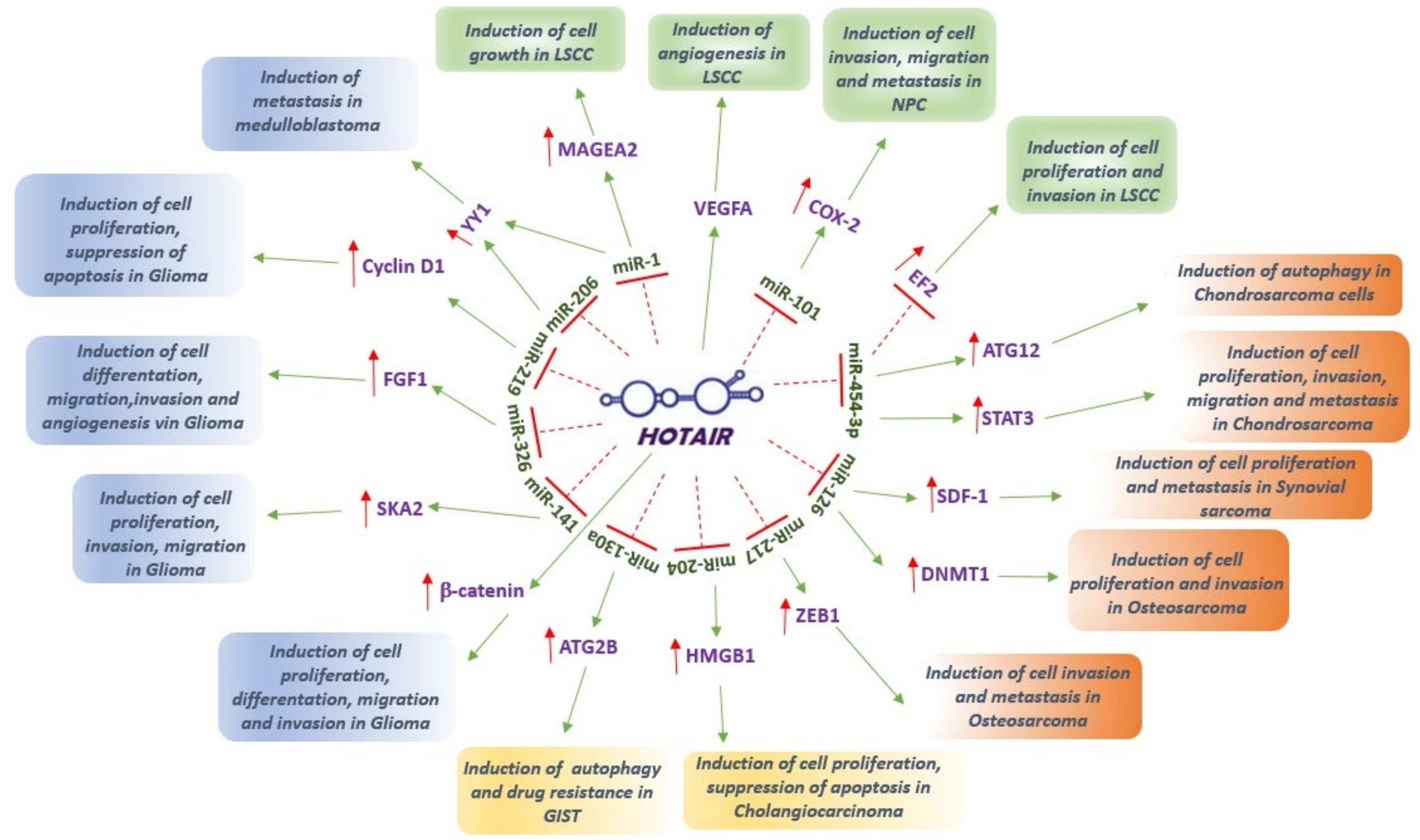{kind=link}
| LncRNAs | Tumor Type | Expression | References |
|---|---|---|---|
| AOC4P | GIST | upregulation | [28] |
| ADAMTS9-AS2 | SACC | upregulation | [29] |
| AFAP1-AS1 | NPC | upregulation | [30] |
| ATB | LSCC | upregulation | [26] |
| BCAR4 | Osteosarcoma | upregulation | [31] |
| CCDC26 | GIST | upregulation | [32,33] |
| DNM3OS | GIST | upregulation | [34] |
| EWSAT1 | Ewing’s sarcoma | upregulation | [35] |
| H19 | LSCC | upregulation | [18] |
| NPC | upregulation | [36] | |
| NEN | upregulation | [37] | |
| GIST | upregulation | [28] | |
| Glioblastoma | upregulation | [38,39] | |
| HNF1A-AS | NPC | upregulation | [36] |
| Osteosarcoma | upregulation | [40] | |
| HOTTIP | Osteosarcoma | upregulation | [41] |
| HOXA11-AS | LSCC | upregulation | [25] |
| NPC | upregulation | [42,43] | |
| KCNQ1OT1 | NPC | upregulation | [44] |
| LINC00668 | LSCC | upregulation | [23] |
| LINC02154 | LSCC | upregulation | [27] |
| MAGI2-AS3 | NPC | upregulation | [45] |
| MALAT1 | Osteosarcoma | upregulation | [46] |
| GEP-NEN | upregulation | [47] | |
| GIST | upregulation | [48] | |
| Glioblastoma | upregulation | [49,50,51,52,53] | |
| MINCR | NPC | upregulation | [54] |
| MRPL23-AS1 | SACC | upregulation | [55] |
| NEAT1 | LSCC | upregulation | [21] |
| Glioblastoma | upregulation | [56] | |
| NONHSAT154433.1 | MEC | upregulation | [57] |
| OIP5-AS1 | NPC | upregulation | [58] |
| PCA3 | Pulmonary NEN | upregulation | [59] |
| PCAT19 | LSCC | upregulation | [22] |
| PILRLS | Liposarcoma | upregulation | [60] |
| SNHG8 | NPC | upregulation | [61] |
| SNHG15 | NPC | upregulation | [62] |
| TP73-AS1 | NPC | upregulation | [63] |
| TUG1 | LSCC | upregulation | [24] |
| Osteosarcoma | upregulation | [64] | |
| TUSC7 | Glioblastoma | upregulation | [65] |
| UCA1 | LSCC | upregulation | [19] |
| Osteosarcoma | upregulation | [66] | |
| LET | NPC | downregulation | [67] |
| MEG3 | Osteosarcoma | downregulation | [68] |
| Pulmonary NEN | downregulation | [59] | |
| TUS7 | Osteosarcoma | downregulation | [68,69] |
3. HOTAIR and Its Role in Human Cancers
4. The Role of HOTAIR in Rare Tumors
4.1. Head and Neck Cancers
4.2. Neuroendocrine Tumors
4.3. Rare Digestive System Tumors
4.4. Central Nervous System Tumors
4.5. Soft Tissue Tumors
5. Conclusions
Funding
Conflicts of Interest
Abbreviations
| HOTAIR | HOX transcript antisense RNA |
| EMT | Epithelial-to-mesenchymal transition |
| LSCC | Laryngeal tumors are squamous cell carcinomas |
| DNMT | DNA methyltransferase enzyme |
| UCA1 | Urothelial Cancer Associated 1 |
| NEAT1 | Nuclear paraspeckle assembly transcript1 |
| PCAT19 | Protocadherin 19 |
| TUG1 | Taurine-upregulated gene 1 |
| HOXA11-AS | HOXA11 antisense RNA |
| MNX1 | Motor Neuron And Pancreas Homeobox 1)-AS1 |
| NPC | Nasopharyngeal carcinoma |
| AFAP1-AS1 | Actin filament-associated protein 1 antisense RNA1 () |
| LET | Low Expression in Tumor |
| SGC | Salivary gland cancer |
| ADAMTS9AS2 | ADAM metallopeptidase thrombospondin type 1, 9 antisense RNA 2 |
| SACC | Salivary adenoid cystic carcinoma |
| MRPL23-AS1 | MRPL23 antisense RNA 1 |
| MEC | Mucoepidermoid carcinoma |
| OS | osteosarcoma |
| MALAT1 | Metastasis Associated Lung Adenocarcinoma Transcript 1 |
| HNF1A | HNF1 Homeobox A-AS1 |
| BCAR4 | Breast Cancer Anti-Estrogen Resistance 4 |
| HULC | Highly upregulated in liver cancer RNA |
| HOTTIP | HOXA Distal Transcript Antisense RNA |
| MEG3 | Maternally expressed 3 |
| EWS-AT1 | EWS RNA Binding Protein 1 |
| PILRLS | Proliferation Interacting LncRNA in Retroperitoneal Liposarcoma |
| NEN | Neuroendocrine neoplasm |
| ISH | In situ hybridization |
| VGF | VGF Nerve Growth Factor Inducible |
| PCA3 | Prostate cancer antigen 3 |
| GISTs | Gastrointestinal stromal tumors |
| TGF-β | Transforming growth factor-beta |
| ZEB1 | Zinc Finger E-Box Binding Homeobox 1 |
| FENDRR | FOXF1 Adjacent Non-Coding Developmental Regulatory RNA |
| CCDC26 | Coiled-coil domain-containing 26 |
| GLUT4 | Glucose transporter member 4 |
| TMZ | Temozolomide |
| NKD1 | NKD Inhibitor of WNT Signaling Pathway 1 |
| TUSC7 | Tumor Suppressor Candidate 7 |
| PRC2 | Polycomb repressive complex |
| LSD1 | Lysine-specific histone demethylase 1A |
| MRE | miRNA recognition element |
| EZH2 | Zeste homolog 2 |
| HCG4 | HLA Complex Group 4 |
| EMX2OS | EMX2 Opposite Strand/Antisense RNA |
| MAGEA2 | Melanoma Antigen A2 gene |
| E2F2 | Eukaryotic elongation factor 2 |
| GRP78 | Glucose regulated protein 78 |
| FASN | Fatty acid synthase |
| COX-2 | Cyclooxygenase-2 |
| TLR4 | Toll Like Receptor 4 |
| STAT3 | Signal transducer and activator of transcription 3 |
| ATG12 | Autophagy-related gene 12 |
| SDF-1 | Stromal cell-derived factor-1 |
| GEP-NET | Gastroenteropancreatic Neuroendocrine Tumors |
| REST | RE1 Silencing Transcription Factor |
| PCDH10 | Protocadherin 10 |
| DPP4 | Dipeptidyl peptidase 4 |
| RASSF1 | Ras Association Domain Family Member 1 |
| ALDH1A3 | Aldehyde Dehydrogenase 1 Family Member A3 |
| ATG2B | Autophagy-related protein 2 homolog B |
| BRD4 | Bromodomain Containing 4 |
| SKA2 | Spindle And Kinetochore Associated Complex Subunit 2 |
| ATRT | Teratoid rhabdoid tumors |
| MPE | Myxopapillary ependymoma |
References
- Botta, L.; Gatta, G.; Trama, A.; Bernasconi, A.; Sharon, E.; Capocaccia, R.; Mariotto, A.B.; RARECARE Net Working Group. Incidence and survival of rare cancers in the US and Europe. Cancer Med. 2020, 9, 5632–5642. [Google Scholar] [CrossRef]
- AIRTUM Working Group; Busco, S.; Buzzoni, C.; Mallone, S.; Trama, A.; Castaing, M.; Bella, F.; Amodio, R.; Bizzoco, S.; Cassetti, T.; et al. Italian cancer figures—Report 2015: The burden of rare cancers in Italy. Epidemiol. Prev. 2016, 40, 1–120. [Google Scholar]
- International Cancer Genome Consortium. International network of cancer genome projects. Nature 2010, 464, 993–998. [Google Scholar] [CrossRef] [Green Version]
- Hutter, C.; Zenklusen, J.C. The Cancer genome atlas: Creating lasting value beyond its data. Cell 2018, 173, 283–285. [Google Scholar] [CrossRef]
- Botti, G.; Franco, R.; Cantile, M.; Ciliberto, G.; Ascierto, P.A. Tumor biobanks in translational medicine. J. Transl. Med. 2012, 10, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botti, G.; Di Bonito, M.; Cantile, M. Organoid biobanks as a new tool for pre-clinical validation of candidate drug efficacy and safety. Int. J. Physiol. Pathophysiol. Pharmacol. 2021, 13, 17–21. [Google Scholar]
- Novikova, I.V.; Hennelly, S.P.; Sanbonmatsu, K.Y. Tackling Structures of Long Noncoding RNAs. Int. J. Mol. Sci. 2013, 14, 23672–23684. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martín, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and abundance analysis of human lncRNAs at single-cell and single-molecule resolution. Genome Biol. 2015, 16, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, X.; Hu, X.; Zhou, W.; Zhang, P.; Zhang, J.; Yang, S.; Liu, Y. The effects of lncRNA MALAT1 on proliferation, invasion and migration in colorectal cancer through regulating SOX9. Mol. Med. 2018, 24, 52. [Google Scholar] [CrossRef]
- Peng, F.; Wang, R.; Zhang, Y.; Zhao, Z.; Zhou, W.; Chang, Z.; Liang, H.; Zhao, W.; Qi, L.; Guo, Z.; et al. Differential expression analysis at the individual level reveals a lncRNA prognostic signature for lung adenocarcinoma. Mol. Cancer 2017, 16, 98. [Google Scholar] [CrossRef]
- Cantile, M.; Di Bonito, M.; Cerrone, M.; Collina, F.; De Laurentiis, M.; Botti, G. Long Non-Coding RNA HOTAIR in Breast Cancer Therapy. Cancers 2020, 12, 1197. [Google Scholar] [CrossRef]
- Kupcinskas, J. Small Molecules in Rare Tumors: Emerging Role of MicroRNAs in GIST. Int. J. Mol. Sci. 2018, 19, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarver, A.E.; Subramanian, S. MicroRNAs in the pathobiology of sarcomas. Lab. Investig. 2015, 95, 987–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO/IARC. Classification of Tumours, 4th ed.; El Naggar, A.K., Chan, J.K.C., Grandis, J.R., Takata, T., Slootweg, P., Eds.; WHO: Geneva, Switzerland, 2020; Volume 9. [Google Scholar]
- Li, W.; Chen, Y.; Niem, X. Regulatory Mechanisms of lncRNAs and Their Target Gene Signaling Pathways in Laryngeal Squamous Cell Carcinoma. Front. Pharmacol. 2020, 11, 1140. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Qu, L.; He, G.; Tian, L.; Li, L.; Zhou, H.; Jin, Q.; Ren, J.; Wang, Y.; Wang, J.; et al. Regulation of laryngeal squamous cell cancer progression by the lncRNA H19/miR-148a-3p/DNMT1 axis. Oncotarget 2016, 7, 11553–11566. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Gong, C.; Yuan, K. LncRNA UCA1 promotes cell proliferation, invasion and migration of laryngeal squamous cell carcinoma cells by activating Wnt/β-catenin signaling pathway. Exp. Ther. Med. 2019, 17, 1182–1189. [Google Scholar] [CrossRef] [Green Version]
- Liang, K.; Yang, Y.; Zha, D.; Yue, B.; Qiu, J.; Zhang, C. Overexpression of lncRNAsnaR is correlated with progression and predicts poor survival of laryngeal squamous cell carcinoma. J. Cell Biochem. 2019, 120, 8492–8498. [Google Scholar] [CrossRef]
- Wang, P.; Wu, T.; Zhou, H.; Jin, Q.; He, G.; Yu, H.; Xuan, L.; Wang, X.; Tian, L.; Sun, Y.; et al. Long noncoding RNA NEAT1 promotes laryngeal squamous cell cancer through regulating miR-107/CDK6 pathway. J. Exp. Clin. Cancer Res. 2016, 35, 22. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Guo, J.; Zhang, W. LncRNA PCAT19 promotes the proliferation of laryngocarcinoma cells via modulation of the miR-182/PDK4 axis. J. Cell. Biochem. 2019, 120, 12810–12821. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Cao, H.; Chi, W.; Meng, W.; Cui, W.; Guo, W.; Wang, B. Expression profile analysis identifies the long non-coding RNA landscape and the potential carcinogenic functions of LINC00668 in laryngeal squamous cell carcinoma. Gene 2019, 687, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, X.; Cao, S.; Han, X.; Wang, Z.; Zhao, X.; Liu, X.; Li, G.; Pan, X.; Lei, D. The Long Noncoding RNA TUG1 Promotes Laryngeal Cancer Proliferation and Migration. Cell. Physiol. Biochem. 2018, 49, 2511–2520. [Google Scholar] [CrossRef]
- Qu, L.; Jin, M.; Yang, L.; Sun, C.; Wang, P.; Li, Y.; Tian, L.; Liu, M.; Sun, Y. Expression of long non-coding RNA HOXA11-AS is correlated with progression of laryngeal squamous cell carcinoma. Am. J. Transl. Res. 2018, 10, 573–580. [Google Scholar]
- Huang, D.N.; Liu, H.W.; Li, Z.D. Expression of lncRNA-ATB in laryngeal carcinoma and its relationship with prognosis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11148–11153. [Google Scholar] [PubMed]
- Zhang, G.; Fan, E.; Zhong, Q.; Feng, G.; Shuai, Y.; Wu, M.; Chen, Q.; Gou, X. Identificationand potential mechanisms of a 4-lncRNA signature that predicts prognosis in patients with laryngeal cancer. Hum. Genom. 2019, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.C.; Wang, Q.; Jiang, L.X.; Cai, L.; Zhai, H.Y.; Yao, Z.W.; Zhang, M.L.; Feng, Y. Effect of long non-coding RNA AOC4P on gastrointestinal stromal tumor cells. OncoTargets Ther. 2018, 11, 6259–6269. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Yu, X.; Li, Y.; Ma, H.; Fan, S.; Chen, W.; Pan, G.; Wang, W.; Zhang, H.; Li, J.; et al. Upregulation of lncRNA ADAMTS9-AS2 Promotes Salivary Adenoid Cystic Carcinoma Metastasis via PI3K/Akt and MEK/Erk Signaling. Mol. Ther. 2018, 26, 2766–2778. [Google Scholar] [CrossRef] [Green Version]
- Bo, H.; Gong, Z.; Zhang, W.; Li, X.; Zeng, Y.; Liao, Q.; Chen, P.; Shi, L.; Lian, Y.; Jing, Y.; et al. Upregulated long non-coding RNA ATB expression is associated with progression and poor prognosis of nasopharyngeal carcinoma. Oncotarget 2015, 6, 20404–20418. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Mo, J.; Zhang, L. Long noncoding RNA BCAR4 promotes osteosarcoma progression through activating GLI2-dependent gene transcription. Tumour Biol. 2016, 37, 13403–13412. [Google Scholar] [CrossRef]
- Cao, K.; Li, M.; Miao, J.; Lu, X.; Kang, X.; Zhu, H.; Du, S.; Li, X.; Zhang, Q.; Guan, W.; et al. CCDC26 knockdown enhances resistance of gastrointestinal stromal tumor cells to imatinib by interacting with c-KIT. Am. J. Transl Res. 2018, 10, 274–282. [Google Scholar] [PubMed]
- Li, H.; Guan, C. HOTAIR inhibits the proliferation of glioblastoma cells by targeting miR-219. Cancer Biomark. 2020, 28, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Yin, Y.; Dai, L.; Shen, C.; Chen, N.; Li, J.; Cai, Z.; Jiang, Z.; Wang, J.; Zhao, Z.; et al. Integrated analysis of long non-coding RNAs and mRNAs associated with malignant transformation of gastrointestinal stromal tumors. Cell Death Dis. 2021, 12, 669. [Google Scholar] [CrossRef] [PubMed]
- Marques Howarth, M.; Simpson, D.; Ngok, S.P.; Nieves, B.; Chen, R.; Siprashvili, Z.; Vaka, D.; Breese, M.R.; Crompton, B.D.; Alexe, G.; et al. Long noncoding RNA ewsat1-mediated gene repression facilitates ewing sarcoma oncogenesis. J. Clin. Investig. 2014, 124, 5275–5290. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, K.; Wu, Q.; Jin, C.S.; Yuan, H.J.; Cheng, J.Z. Long non-coding RNA HNF1A-AS is upregulated and promotes cell proliferation and metastasis in nasopharyngeal carcinoma. Cancer Biomark. 2016, 16, 291–300. [Google Scholar] [CrossRef]
- Ji, M.; Yao, Y.; Liu, A.; Shi, L.; Chen, D.; Tang, L.; Yang, G.; Liang, X.; Peng, J.; Shao, C. lncRNA H19 binds VGF and promotes pNEN progression via PI3K/AKT/CREB signalling. Endocr. Relat. Cancer 2019, 26, 643–658. [Google Scholar] [CrossRef]
- Fazi, B.; Garbo, S.; Toschi, N.; Mangiola, A.; Lombari, M.; Sicari, D. The Lncrna H19 Positively Affects the Tumorigenic Properties of Glioblastoma Cells and Contributes to NKD1 Repression Through the Recruitment of EZH2 on Its Promoter. Oncotarget 2018, 9, 15512. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Li, M.; Wang, Z.; Wang, L.; Liu, Y. H19 Induced by Oxidative Stress Confers Temozolomide Resistance in Human Glioma Cells Via Activating NF-κb Signaling. OncoTargets Ther. 2018, 11, 6395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Hou, W.; Tao, J.; Zhao, Y.; Wan, G.; Ma, C.; Xu, H. Upregulation of lncRNA HNF1A-AS1 promotes cell proliferation and metastasis in osteosarcoma through activation of the Wnt/β-catenin signaling pathway. Am. J. Transl. Res. 2016, 8, 3503–3512. [Google Scholar]
- Li, F.; Cao, L.; Hang, D.; Wang, F.; Wang, Q. Long non-coding RNA HOTTIP is upregulated and associated with poor prognosis in patients with osteosarcoma. Int. J. Clin. Exp. Pathol. 2015, 8, 11414–11420. [Google Scholar]
- Lin, F.J.; Lin, X.D.; Xu, L.Y.; Zhu, S.Q. Long Noncoding RNA HOXA11-AS Modulates the Resistance of Nasopharyngeal Carcinoma Cells to Cisplatin via miR-454-3p/c-Met. Mol. Cells 2020, 43, 856–869. [Google Scholar]
- Li, H.; Huang, J.; Yu, S.; Li, H.; Zhou, Y.; Wu, Q. HOXA11-AS induces cisplatin resistance by modulating the microRNA-98/PBX3 axis in nasopharyngeal carcinoma. Oncol. Lett. 2021, 21, 493. [Google Scholar] [CrossRef]
- Yuan, F.; Lou, Z.; Zhou, Z.; Yan, X. Long non-coding RNA LET promotes nasopharyngeal carcinoma cell cisplatin resistance via the miR-454/USP47 axis. Int. J. Mol. Med. 2021, 47, 54. [Google Scholar] [CrossRef]
- Cao, C.; Zhou, S.; Hu, J. Long noncoding RNA MAGI2-AS3/miR-218-5p/GDPD5/SEC61A1 axis drives cellular proliferation and migration and confers cisplatin resistance in nasopharyngeal carcinoma. Int. Forum Allergy Rhinol. 2020, 10, 1012–1023. [Google Scholar] [CrossRef]
- Dong, Y.; Liang, G.; Yuan, B.; Yang, C.; Gao, R.; Zhou, X. MALAT1 promotes the proliferation and metastasis of osteosarcoma cells by activating the PI3k/Akt pathway. Tumour Biol. 2015, 36, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.H.; Hardin, H.; Jens Eickhoff, J.; Lloyd, R.V. In Situ Hybridization Analysis of Long Non-coding RNAs MALAT1 and HOTAIR in Gastroenteropancreatic Neuroendocrine Neoplasms. Endocr. Pathol. 2019, 30, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, G.; Barraco, N.; Incorvaia, L.; Galvano, A.; Fanale, D.; Cabibi, D.; Calò, V.; Currò, G.; Bazan, V.; Russo, A. Are Long Noncoding RNAs New Potential Biomarkers in Gastrointestinal Stromal Tumors (GISTs)? The Role of H19 and MALAT1. J. Oncol. 2019, 2019, 5458717. [Google Scholar] [CrossRef] [Green Version]
- Vassallo, I.; Zinn, P.; Lai, M.; Rajakannu, P.; Hamou, M.F.; Hegi, M.E. WIF1 re-expression in glioblastoma inhibits migration through attenuation of non-canonical WNT signaling by downregulating the lncRNA MALAT1. Oncogene 2016, 35, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yuan, X.; Yan, D.; Li, D.; Guan, F.; Dong, Y. Long Non-Coding RNA MALAT1 Decreases the Sensitivity of Resistant Glioblastoma Cell Lines to Temozolomide. Cell Physiol. Biochem. 2017, 42, 1192–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Xu, X.-K.; Li, J.-L.; Kong, K.-K.; Li, H.; Chen, C. MALAT1 is a Prognostic Factor in Glioblastoma Multiforme and Induces Chemoresistance to Temozolomide Through Suppressing miR-203 and Promoting Thymidylate Synthase Expression. Oncotarget 2017, 8, 22783. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.; Liu, Y.; Xiao, J. Long Noncoding RNA MALAT 1 Knockdown Reverses Chemoresistance to Temozolomide Via Promoting Micro RNA-101 in Glioblastoma. Cancer Med. 2018, 7, 1404–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voce, D.J.; Bernal, G.M.; Wu, L.; Crawley, C.D.; Zhang, W.; Mansour, N.M. Temozolomide Treatment Induces Lncrna MALAT1 in an NF-κb and p53 Codependent Manner in Glioblastoma. Cancer Res. 2019, 79, 2536–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Q.; Chen, Y.; Chen, Z. LncRNA MINCR regulates irradiation resistance in nasopharyngeal carcinoma cells via the microRNA-223/ZEB1 axis. Cell Cycle 2020, 19, 53–66. [Google Scholar] [CrossRef]
- Chen, C.W.; Fu, M.; Du, Z.H.; Zhao, F.; Yang, W.W.; Xu, L.H.; Li, S.L.; Ge, X.Y. Long Noncoding RNA NEAT1 Promotes Adenoid Cystic Carcinoma Lung Metastasis. Cancer Res. 2020, 80, 2273–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.-L.; Xu, Q.-S.; Wu, Z.-X.; Xu, S.-J.; Shen, J. Lncrna NEAT1 is Involved in Temozolomide Resistance by Regulating MGMT in Glioblastoma Multiforme. Clin. Surg. Res. Commun. 2018, 2, 24–30. [Google Scholar]
- Lu, H.; Han, N.; Xu, W.; Zhu, Y.; Liu, L.; Liu, S.; Yang, W. Screening and bioinformatics analysis of mRNA, long non-coding RNA and circular RNA expression profiles in mucoepidermoid carcinoma of salivary gland. Biochem. Biophys. Res. Commun. 2019, 508, 66–71. [Google Scholar] [CrossRef]
- Tang, J.; Fu, C.; Li, Y.; Chen, S.; Jiang, X.; Xu, W.; Xie, H. Long Noncoding RNA PCA3 Promotes the Disease Progression in Nasopharyngeal Carcinoma by Targeting miR-203. Biomed. Res. Int. 2021, 2021, 9850928. [Google Scholar] [CrossRef]
- Narayanan, D.; Mandal, R.; Hardin, H.; Chanana, V.; Schwalbe, M.; Rosenbaum, J.; Buehler, D.; Lloyd, R.V. Long Non-coding RNAs in Pulmonary Neuroendocrine Neoplasms. Endocr. Pathol. 2020, 31, 254–263. [Google Scholar] [CrossRef]
- Shao, Y.; Zhang, Y.; Hou, Y.; Tong, H.; Zhuang, R.; Ji, Z.; Wang, B.; Zhou, Y.; Lu, W. A novel long noncoding RNA PILRLS promote proliferation through TCL1A by activing MDM2 in retroperitoneal liposarcoma. Oncotarget 2017, 8, 13971–13978. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Liu, Y.; Wang, Z.; Wu, S. lncRNA SNHG8 promotes aggressive behaviors of nasopharyngeal carcinoma via regulating miR-656-3p/SATB1 axis. Biomed. Pharmacother. 2020, 131, 110564. [Google Scholar] [CrossRef]
- Yi, T.W.; Lv, X.X.; Fan, H.; Zan, N.; Su, X.D. LncRNA SNHG15 promotes the proliferation of nasopharyngeal carcinoma via sponging miR-141-3p to upregulate KLF9. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6744–6751. [Google Scholar]
- Hu, J.; Pan, J.; Luo, Z.; Duan, Q.; Wang, D. Long non-coding RNA FOXD3-AS1 silencing exerts tumor suppressive effects in nasopharyngeal carcinoma by downregulating FOXD3 expression via microRNA-185-3p upregulation. Cancer Gene Ther. 2021, 28, 602–618. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Li, M.; Zhang, L.; Huang, M.; Lei, J.B.; Fu, G.H.; Liu, C.X.; Lai, Q.W.; Chen, Q.Q.; Wang, Y.L. Upregulation of long non-coding RNA TUG1 correlates with poor prognosis and disease status in osteosarcoma. Tumour Biol. 2016, 37, 4445–4455. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.; Tang, W.; Pan, C.; Hu, X.; Hong, Y. Long Non-Coding RNA TUSC7 Inhibits Temozolomide Resistance by Targeting miR-10a in Glioblastoma. Cancer Chemother. Pharmacol. 2018, 81, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xie, P.; Ruan, W.H. Overexpression of lncRNA UCA1 promotes osteosarcoma progression and correlates with poor prognosis. J. Bone Oncol. 2016, 5, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Liu, H.; Li, L.; Zhang, S.; Liu, K.; Liu, Y.; Yang, C. Long noncoding RNA-LET, which is repressed by EZH2, inhibits cell proliferation and induces apoptosis of nasopharyngeal carcinoma cell. Med. Oncol. 2015, 32, 226. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.Z.; Guo, X.J.; Zhao, Y.M.; Fang, Y. Decreased expression of long non-coding RNA MEG3 acts as a potential predictor biomarker in progression and poor prognosis of osteosarcoma. Int. J. Clin. Exp. Pathol. 2015, 8, 15138–15142. [Google Scholar] [PubMed]
- Cong, M.; Li, J.; Jing, R.; Li, Z. Long non-coding RNA tumor suppressor candidate 7 functions as a tumor suppressor and inhibits proliferation in osteosarcoma. Tumour Biol. 2016, 37, 9441–9450. [Google Scholar] [CrossRef]
- Li, X.; Lin, Y.; Yang, X.; Wu, X.; He, X. Long noncoding RNA H19 regulates EZH2 expression by interacting with miR-630 and promotes cell invasion in nasopharyngeal carcinoma. Biochem. Biophys. Res. Commun. 2016, 13, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.E.; Tao, J.M.; Wang, S.X.; Ju, X.; Song, Z.Y. Long non-coding RNA HEIH modulates CDK8 expression by inhibiting miR-193a-5p to accelerate nasopharyngeal carcinoma progression. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 770–778. [Google Scholar]
- Dai, B.Q.; Jiang, X.; Feng, L.C. LncRNA TUG1 regulates miR-495 expression to promote migration and invasion of nasopharyngeal carcinoma cells through junctional adhesion molecule A. Kaohsiung J. Med. Sci. 2021, 37, 361–370. [Google Scholar] [CrossRef]
- Feng, M.B.; Li, G.H.; Dou, F.F. Long-chain non-coding RNA LOC554202 promotes proliferation, migration, and invasion of nasopharyngeal carcinoma cells by binding to microRNA-31 expression and regulating RhoA expression. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10550–10556. [Google Scholar] [PubMed]
- Yue, W.J.; Wang, Y.; Li, W.Y.; Wang, Z.D. LINC00887 regulates the proliferation of nasopharyngeal carcinoma via targeting miRNA-203b-3p to upregulate NUP205. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8863–8870. [Google Scholar]
- Li, H.; Huang, J.; Yu, S.; Lou, Z. Long Non-Coding RNA DLEU1 Up-Regulates BIRC6 Expression by Competitively Sponging miR-381-3p to Promote Cisplatin Resistance in Nasopharyngeal Carcinoma. OncoTargets Ther. 2020, 13, 2037–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Pu, T.; Zhang, Y.; Wang, J.; Zhao, Y. Long non-coding RNA LINC00346 contributes to cisplatin resistance in nasopharyngeal carcinoma by repressing miR-342-5p. Open Biol. 2020, 10, 190286. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.Q.; Li, Z.X.; Guan, J.L.; Liu, X.; Li, J.Y.; Chen, Y.; Lin, L.; Kou, J.; Lv, J.W.; Zhang, L.L.; et al. Long Noncoding RNA TINCR-Mediated Regulation of Acetyl-CoA Metabolism Promotes Nasopharyngeal Carcinoma Progression and Chemoresistance. Cancer Res. 2020, 80, 5174–5188. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, L.; Wang, Y.; Cong, J.; Lin, Z.; Wang, Y.; Liu, Q.; Wang, L.; Yang, B.; Li, T. lncRNA MIAT/HMGB1 Axis Is Involved in Cisplatin Resistance via Regulating IL6-Mediated Activation of the JAK2/STAT3 Pathway in Nasopharyngeal Carcinoma. Front. Oncol. 2021, 11, 651693. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zheng, W.; Chen, Q.; Zhou, Y.; Pan, Y.; Zhang, J.; Bai, Y.; Shao, C. lncRNA CASC19 Contributes to Radioresistance of Nasopharyngeal Carcinoma by Promoting Autophagy via AMPK-mTOR Pathway. Int. J. Mol. Sci. 2021, 22, 1407. [Google Scholar] [CrossRef]
- Peng, J.; Liu, F.; Zheng, H.; Wu, Q.; Liu, S. IncRNA ZFAS1 contributes to the radioresistance of nasopharyngeal carcinoma cells by sponging hsa-miR-7-5p to upregulate ENO2. Cell Cycle 2021, 20, 126–141. [Google Scholar] [CrossRef]
- Speight, P.M.; Barrett, A.W. Salivary gland tumours: Diagnostic challenges and an update on the latest WHO classification. Diagn. Histopathol. 2020, 26, 147–158. [Google Scholar] [CrossRef]
- Kallen, M.E.; Hornick, J.L. The 2020 WHO Classification: What’s New in Soft Tissue Tumor Pathology? Am. J. Surg. Pathol. 2021, 45, e1–e23. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.H.; Yang, L.B.; Geng, X.L.; Wang, R.; Zhang, Z.C. Increased expression of lncRNA HULC indicates a poor prognosis and promotes cell metastasis in osteosarcoma. Int. J. Clin. Exp. Pathol. 2015, 8, 2994–3000. [Google Scholar] [PubMed]
- Taal, B.G.; Visser, O. Epidemiology of neuroendocrine tumours. Neuroendocrinology 2004, 8, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Oronsky, B.; Ma, P.C.; Morgensztern, D.; Carter, C.A. Nothing But NET: A Review of Neuroendocrine Tumors and Carcinomas. Neoplasia 2017, 19, 991–1002. [Google Scholar] [CrossRef]
- Luo, J.; Wang, K.; Yeh, S.; Sun, Y.; Liang, L.; Xiao, Y.; Xu, W.; Niu, Y.; Liang Cheng, L.; Maity, S.N.; et al. LncRNA-p21 alters the antiandrogen enzalutamide-induced prostate cancer neuroendocrine differentiation via modulating the EZH2/STAT3 signaling. Nat. Commun. 2019, 10, 2571. [Google Scholar] [CrossRef] [Green Version]
- Al-Share, B.; Alloghbi, A.; Al Hallak, M.N.; Uddin, H.; Azmi, A.; Mohammad, R.M.; Kim, S.H.; Shields, A.F.; Philip, P.A. Gastrointestinal stromal tumor: A review of current and emerging therapies. Cancer Metastasis Rev. 2021, 40, 625–641. [Google Scholar] [CrossRef]
- Gyvyte, U.; Kupcinskas, J.; Juzenas, S.; Inciuraite, R.; Poskiene, L.; Salteniene, V.; Link, A.; Fassan, M.; Franke, A.; Kupcinskas, L.; et al. Identification of long intergenic non-coding RNAs (lincRNAs) deregulated in gastrointestinal stromal tumors (GISTs). PLoS ONE 2018, 13, 0209342. [Google Scholar]
- Reni, M.; Mazza, E.; Zanon, S.; Gatta, G.; Vecht, C.J. Central nervous system gliomas. Crit. Rev. Oncol. Hematol. 2017, 113, 213–234. [Google Scholar] [CrossRef]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [Green Version]
- Ding, A.; Mandal, S.S. LncRNA HOTAIR: A master regulator of chromatin dynamics and cancer. Biochim. Biophys. Acta 2015, 1856, 151–164. [Google Scholar]
- Majello, B.; Gorini, F.; Saccà, C.D.; Amente, S. Expanding the role of the histone lysine-specific demethylase LSD1 in cancer. Cancers 2019, 11, 324. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Abdelmohsen, K.; Kim, J.; Yang, X.; Martindale, J.L.; Tominaga-Yamanaka, K.; White, E.J.; Orjalo, A.V.; Rinn, J.L.; Kreft, S.G. Scaffold function of long non-coding RNA HOTAIR in protein ubiquitination. Nat. Commun. 2013, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, H.; Li, Y.; Wang, R.; Li, Y.; Zhang, H.; Ren, D.; Liu, H.; Kang, C.; Chen, J. HOTAIR, a long noncoding RNA, is a marker of abnormal cell cycle regulation in lung cancer. Cancer Sci. 2018, 109, 2717–2733. [Google Scholar] [CrossRef] [Green Version]
- Ding, W.; Ren, J.; Ren, H.; Wang, D. Long Noncoding RNA HOTAIR Modulates MiR-206-mediated Bcl-w Signaling to Facilitate Cell Proliferation in Breast Cancer. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Peng, X.; Cheng, D.; Zhu, Y.; Du, J.; Li, J.; Yu, X. Delphinidin suppresses breast carcinogenesis through the HOTAIR/microRNA-34a axis. Cancer Sci. 2019, 110, 3089–3097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantile, M.; Di Bonito, M.; Tracey De Bellis, M.; Botti, G. Functional Interaction among lncRNA HOTAIR and MicroRNAs in Cancer and Other Human Diseases. Cancers 2021, 13, 570. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Kong, D.; Chen, Q.; Ping, Y.; Pang, D. Oncogenic long noncoding RNA landscape in breast cancer. Mol. Cancer 2017, 16, 129. [Google Scholar] [CrossRef] [Green Version]
- Botti, G.; Collina, F.; Scognamiglio, G.; Aquino, G.; Cerrone, M.; Liguori, G.; Gigantino, V.; Malzone, M.G.; Cantile, M. LncRNA HOTAIR Polymorphisms Association with Cancer Susceptibility in Different Tumor Types. Curr. Drug Targets 2018, 19, 1220–1226. [Google Scholar] [CrossRef]
- Qu, X.; Alsager, S.; Zhuo, Y.; Shan, B. HOX transcript antisense RNA (HOTAIR) in cancer. Cancer Lett. 2019, 454, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Gökmen-Polar, Y.; Vladislav, I.T.; Neelamraju, Y.; Janga, S.C.; Badve, S. Prognostic impact of HOTAIR expression is restricted to ER-negative breast cancers. Sci. Rep. 2015, 5, 8765. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, K.P.; Thomassen, M.; Tan, Q.; Bak, M.; Cold, S.; Burton, M.; Larsen, M.J.; Kruse, T.A. Long non-coding RNA HOTAIR is an independent prognostic marker of metastasis in estrogen receptor-positive primary breast cancer. Breast Cancer Res. Treat. 2013, 142, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Nguyen, H.T.; Burow, M.E.; Zhuo, Y.; El-Dahr, S.S.; Yao, X.; Cao, S.; Flemington, E.K.; Nephew, K.P.; Fang, F. Elevated expression of long intergenic non-coding RNA HOTAIR in a basal-like variant of MCF-7 breast cancer cells. Mol. Carcinog. 2015, 54, 1656–1667. [Google Scholar] [CrossRef] [Green Version]
- Collina, F.; Aquino, G.; Brogna, M.; Cipolletta, S.; Buonfanti, G.; De Laurentiis, M.; Di Bonito, M.; Cantile, M.; Botti, G. LncRNA HOTAIR upregulation is strongly related with lymph nodes metastasis and LAR subtype of Triple Negative Breast Cancer. J. Cancer 2019, 10, 2018–2024. [Google Scholar] [CrossRef] [Green Version]
- Pádua Alves, C.; Fonseca, A.S.; Muys, B.R.; de Barros, E.; Lima Bueno, R.; Bürger, M.C.; de Souza, J.E.; Valente, V.; Zago, M.A.; Silva, W.A., Jr. Brief report: The lincRNA Hotair is required for epithelial-to-mesenchymal transition and stemness maintenance of cancer cell lines. Stem Cells 2013, 31, 2827–2832. [Google Scholar] [CrossRef]
- Ren, Y.; Jia, H.H.; Xu, Y.Q.; Zhou, X.; Zhao, X.H.; Wang, Y.F.; Song, X.; Zhu, Z.Y.; Sun, T.; Dou, Y. Paracrine and epigenetic control of CAF-induced metastasis: The role of HOTAIR stimulated by TGF-ß1 secretion. Mol. Cancer 2018, 17, 5. [Google Scholar] [CrossRef]
- Zhang, H.; Cai, K.; Wang, J.; Wang, X.; Cheng, K.; Shi, F.; Jiang, L.; Zhang, Y.; Dou, J. MiR-7, inhibited indirectly by lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of breast cancer stem cells by downregulating the STAT3 pathway. Stem Cells 2014, 32, 2858–2868. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yang, M.; Jiang, R.; An, N.; Wang, X.; Liu, B. Long Non-Coding RNA HOTAIR Regulates the Proliferation, Self-Renewal Capacity, Tumor Formation and Migration of the Cancer Stem-Like Cell (CSC) Subpopulation Enriched from Breast Cancer Cells. PLoS ONE 2017, 12, e0170860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Song, X.; Wang, X.; Xie, Y.; Wang, Z.; Xu, Y.; You, X.; Liang, Z.; Cao, H. Circulating DNA of HOTAIR in serum is a novel biomarker for breast cancer. Breast Cancer Res. Treat. 2015, 152, 199–208. [Google Scholar] [CrossRef]
- Tang, S.; Zheng, K.; Tang, Y.; Li, Z.; Zou, T.; Liu, D. Overexpression of serum exosomal HOTAIR is correlated with poor survival and poor response to chemotherapy in breast cancer patients. J. Biosci. 2019, 44, 37. [Google Scholar] [CrossRef]
- Luo, Z.F.; Zhao, D.; Li, X.Q.; Cui, Y.X.; Ma, N.; Lu, C.X.; Liu, M.Y.; Zhou, Y. Clinical significance of HOTAIR expression in colon cancer. World J. Gastroenterol. 2016, 22, 5254–5259. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Dong, S.; Duan, B.; Chen, P.; Shi, L.; Gao, H.; Qi, H. HOTAIR is a predictive and prognostic biomarker for patients with advanced gastric adenocarcinoma receiving fluorouracil and platinum combination chemotherapy. Am. J. Transl. Res. 2015, 7, 1295–1302. [Google Scholar] [PubMed]
- Ishibashi, M.; Kogo, R.; Shibata, K.; Sawada, G.; Takahashi, Y.; Kurashige, J.; Akiyoshi, S.; Sasaki, S.; Iwaya, T. Clinical significance of the expression of long non-coding RNA HOTAIR in primary hepatocellular carcinoma. Oncol. Rep. 2013, 29, 946–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Zhou, L.; Wu, L.M.; Lai, M.C.; Xie, H.Y.; Zhang, F.; Zheng, S.S. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann. Surg. Oncol. 2011, 18, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zhao, J.C.; Kim, J.; Fong, K.W.; Yang, Y.A.; Chakravarti, D.; Mo, Y.Y.; Yu, J. LncRNA HOTAIR Enhances the Androgen-Receptor-Mediated Transcriptional Program and Drives Castration-Resistant Prostate Cancer. Cell Rep. 2015, 13, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Yan, T.H.; Lu, S.W.; Huang, Y.Q.; Que, G.B.; Chen, J.H.; Chen, Y.P.; Zhang, H.B.; Liang, X.L.; Jiang, J.H. Upregulation of the long noncoding RNA HOTAIR predicts recurrence in stage Ta/T1 bladder cancer. Tumour Biol. 2014, 35, 10249–10257. [Google Scholar] [CrossRef]
- Shang, C.; Guo, Y.; Zhang, H.; Xue, Y.X. Long noncoding RNA HOTAIR is a prognostic biomarker and inhibits chemosensitivity to doxorubicin in bladder transitional cell carcinoma. Cancer Chemother. Pharmacol. 2016, 77, 507–513. [Google Scholar] [CrossRef]
- Berrondo, C.; Flax, J.; Kucherov, V.; Siebert, A.; Osinski, T.; Rosenberg, A.; Fucile, C.; Richheimer, S.; Beckham, C.J. Expression of the Long Non-Coding RNA HOTAIR Correlates with Disease Progression in Bladder Cancer and Is Contained in Bladder Cancer Patient Urinary Exosomes. PLoS ONE 2016, 11, e0147236. [Google Scholar]
- Qiu, J.J.; Lin, Y.Y.; Ye, L.C.; Ding, J.X.; Feng, W.W.; Jin, H.Y.; Zhang, Y.; Li, Q.; Hua, K.Q. Overexpression of long non-coding RNA HOTAIR predicts poor patient prognosis and promotes tumor metastasis in epithelial ovarian cancer. Gynecol. Oncol. 2014, 134, 121–128. [Google Scholar] [CrossRef]
- Huang, L.; Liao, L.M.; Liu, A.W.; Wu, J.B.; Cheng, X.L.; Lin, J.X.; Zheng, M. Overexpression of long noncoding RNA HOTAIR predicts a poor prognosis in patients with cervical cancer. Arch. Gynecol. Obstet. 2014, 290, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Yu, J.; Dong, R.; Qiu, H. A high level of circulating HOTAIR is associated with progression and poor prognosis of cervical cancer. Tumour Biol. 2015, 36, 1661–1665. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Bao, W.; Li, X.; Chen, Z.; Che, Q.; Wang, H.; Wan, X.P. The long non-coding RNA HOTAIR is upregulated in endometrial carcinoma and correlates with poor prognosis. Int. J. Mol. Med. 2014, 33, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.Y.; Zhu, J.Y.; Zhang, C.Y.; Zhang, M.; Song, Y.N.; Rahman, K.; Zhang, L.J.; Zhang, H. Autophagy regulated by lncRNA HOTAIR contributes to the cisplatin-induced resistance in endometrial cancer cells. Biotechnol. Lett. 2017, 39, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Endo, H.; Yokoyama, M.; Abe, J.; Tamai, K.; Tanaka, N.; Sato, I.; Takahashi, S.; Kondo, T.; Satoh, K. Large noncoding RNA HOTAIR enhances aggressive biological behavior and is associated with short disease-free survival in human non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2013, 436, 319–324. [Google Scholar] [CrossRef]
- Liu, M.Y.; Li, X.Q.; Gao, T.H.; Cui, Y.; Ma, N.; Zhou, Y.; Zhang, G.J. Elevated HOTAIR expression associated with cisplatin resistance in non-small cell lung cancer patients. J. Thorac. Dis. 2016, 8, 3314–3322. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Xiao, X.; Wu, C.; Huang, J.; Zhang, Y.; Xie, M.; Zhang, M.; Zhou, L. The role of long non-coding RNA HOTAIR in the progression and development of laryngeal squamous cell carcinoma interacting with EZH2. Acta Otolaryngol. 2017, 137, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xin, Y.; Zhou, L.; Huang, J.M.; Tao, L.; Cheng, L.; Tian, J. Cisplatin and paclitaxel target significant long noncoding RNAs in laryngeal squamous cell carcinoma. Med. Oncol. 2014, 31, 246. [Google Scholar] [CrossRef]
- Nie, Y.; Liu, X.; Qu, S.; Song, E.; Zou, H.; Gong, C. Long non-coding RNA HOTAIR is an independent prognostic marker for nasopharyngeal carcinoma progression and survival. Cancer Sci. 2013, 104, 458–464. [Google Scholar] [CrossRef]
- Fu, W.M.; Lu, Y.F.; Hu, B.G.; Liang, W.C.; Zhu, X.; Yang, H.D.; Li, G.; Zhang, J.F. Long noncoding RNA Hotair mediated angiogenesis in nasopharyngeal carcinoma by direct and indirect signaling pathways. Oncotarget 2016, 7, 4712–4723. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.B.; Luo, L. Expression of HOX transcript antisense intergenic RNA in salivary adenoid cystic carcinoma and its influence on prognosis. Hua Xi Kou Qiang Yi Xue Za Zhi West China J. Stomatol. 2020, 38, 509–512. [Google Scholar]
- Zhang, J.; Chen, K.; Tang, Y.; Luan, X.; Zheng, X.; Lu, X.; Mao, J.; Hu, L.; Zhang, S.; Zhang, X.; et al. LncRNA-HOTAIR activates autophagy and promotes the imatinib resistance of gastrointestinal stromal tumor cells through a mechanism involving the miR-130a/ATG2B pathway. Cell Death Dis. 2021, 12, 367. [Google Scholar] [CrossRef]
- Pastori, C.; Kapranov, P.; Penas, C.; Peschansky, V.; Volmar, C.H.; Sarkaria, J.N.; Bregy, A.; Komotar, R.; St Laurent, G.; Ayad, N.G.; et al. The Bromodomain protein BRD4 controls HOTAIR, a long noncoding RNA essential for glioblastoma proliferation. Proc. Natl. Acad. Sci. USA 2015, 112, 8326–8331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.K.; Pastori, C.; Penas, C.; Komotar, R.J.; Ivan, M.E.; Wahlestedt, C.; Ayad, N.G. Serum long noncoding RNA HOTAIR as a novel diagnostic and prognostic biomarker in glioblastoma multiforme. Mol. Cancer 2018, 17, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Lu, H.; Fan, G.; He, M.; Sun, Y.; Xu, K.; Shi, F. A novel interplay between HOTAIRn and DNA methylation in osteosarcoma cells indicates a new therapeutic strategy. J. Cancer Res. Clin. Oncol. 2017, 143, 2189–2200. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.T.; Lin, T.P.; Tang, J.T.; Campbell, M.; Luo, Y.L.; Lu, S.Y.; Yang, C.P.; Cheng, T.Y.; Chang, C.H.; Liu, T.T.; et al. HOTAIR is a REST-regulated lncRNA that promotes neuroendocrine differentiation in castration resistant prostate cancer. Cancer Lett. 2018, 433, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulou, D.I.; Laschos, K.; Aravantinos, G.; Georgiou, K.; Papiris, K.; Theodoropoulos, G.; Gazouli, M.; Filippou, D. Association between homeobox protein transcript antisense intergenic ribonucleic acid genetic polymorphisms and cholangiocarcinoma. World J. Clin. Cases 2021, 9, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Niinuma, T.; Suzuki, H.; Nojima, M.; Nosho, K.; Yamamoto, H.; Takamaru, H.; Yamamoto, E.; Maruyama, R. Upregulation of miR-196a and HOTAIR drive malignant character in gastrointestinal stromal tumors. Cancer Res. 2012, 72, 1126–1136. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Feng, J.; Wu, T.; Wang, Y.; Sun, Y.; Ren, J.; Liu, M. Long intergenic noncoding RNA HOTAIR is overexpressed and regulates PTEN methylation in laryngeal squamous cell carcinoma. Am. J. Pathol. 2013, 182, 64–70. [Google Scholar] [CrossRef]
- Tang, Z.; Wei, G.; Zhang, L.; Xu, Z. Signature microRNAs and long noncoding RNAs in laryngeal cancer recurrence identified using a competing endogenous RNA network. Mol. Med. Rep. 2019, 19, 4806–4818. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, Y.; Lu, J.; Sun, Y.; Xiao, H.; Liu, M.; Tian, L. Combined detection of serum exosomal miR-21 and HOTAIR as diagnostic and prognostic biomarkers for laryngeal squamous cell carcinoma. Med. Oncol. 2014, 31, 148. [Google Scholar] [CrossRef]
- Cui, X.; Xiao, D.; Cui, Y.; Wang, X. Exosomes-Derived Long Non-Coding RNA HOTAIR Reduces Laryngeal Cancer Radiosensitivity by Regulating microRNA-454-3p/E2F2 Axis. OncoTargets Ther. 2019, 12, 10827–10839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.D.; Yuan, L.L.; Lin, L.Q. LncRNA HOTAIR contributes to the tumorigenesis of nasopharyngeal carcinoma via up-regulating FASN. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5143–5152. [Google Scholar] [PubMed]
- Hu, W.; Xu, W.; Shi, Y.; Dai, W. lncRNA HOTAIR upregulates COX-2 expression to promote invasion and migration of nasopharyngeal carcinoma by interacting with miR-101. Biochem. Biophys. Res. Commun. 2018, 505, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.L.; Wei, Y.; Liao, B.Y.; Wei, G.J.; Qin, H.M.; Pang, X.X.; Wang, J.L. LncRNA HOTAIR regulates the expression of E-cadherin to affect nasopharyngeal carcinoma progression by recruiting histone methylase EZH2 to mediate H3K27 trimethylation. Genomics 2021, 113, 2276–2289. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, A.; Scognamiglio, G.; Aquino, G.; Cerrone, M.; Liguori, G.; Clemente, O.; Di Bonito, M.; Cantile, M.; Botti, G.; Tafuto, S.; et al. Aberrant Expression of Long Non Coding RNA HOTAIR and De-Regulation of the Paralogous 13 HOX Genes Are Strongly Associated with Aggressive Behavior of Gastro-Entero-Pancreatic Neuroendocrine Tumors. Int. J. Mol. Sci. 2021, 22, 7049. [Google Scholar] [CrossRef] [PubMed]
- Mather, R.L.; Wang, Y.; Crea, F. Is HOTAIR really involved in neuroendocrine prostate cancer differentiation? Epigenomics 2018, 10, 1259–1261. [Google Scholar] [CrossRef] [Green Version]
- Blechacz, B.; Komuta, M.; Roskams, T.; Gores, G.J. Clinical diagnosis and staging of cholangiocarcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Kang, P.; Xu, Y.; Leng, K.; Li, Z.; Huang, L.; Gao, J.; Cui, Y.; Zhong, X. Long non-coding RNA HOTAIR promotes tumorigenesis and forecasts a poor prognosis in cholangiocarcinoma. Sci. Rep. 2018, 8, 12176. [Google Scholar] [CrossRef]
- Lu, M.; Qin, X.; Zhou, Y.; Li, G.; Liu, Z.; Yue, H.; Geng, X. LncRNA HOTAIR suppresses cell apoptosis, autophagy and induces cell proliferation in cholangiocarcinoma by modulating the miR-204-5p/HMGB1 axis. Biomed. Pharmacother. 2020, 130, 110566. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Lee, J.H.; Kim, W.K.; Yun, S.; Youn, Y.H.; Park, C.H.; Choi, Y.Y.; Kim, H.; Lee, S.K. Promoter methylation of PCDH10 by HOTAIR regulates the progression of gastrointestinal stromal tumors. Oncotarget 2016, 7, 75307–75318. [Google Scholar] [CrossRef]
- Bure, I.; Geer, S.; Knopf, J.; Roas, M.; Henze, S.; Ströbel, P.; Agaimy, A.; Wiemann, S.; Hoheisel, J.D.; Hartmann, A.; et al. Long noncoding RNA HOTAIR is upregulated in an aggressive subgroup of gastrointestinal stromal tumors (GIST) and mediates the establishment of gene-specific DNA methylation patterns. Genes Chromosomes Cancer 2018, 57, 584–597. [Google Scholar] [CrossRef]
- Yan, J.; Chen, D.; Chen, X.; Sun, X.; Dong, Q.; Hu, C.; Zhou, F.; Chen, W. Downregulation of lncRNA CCDC26 contributes to imatinib resistance in human gastrointestinal stromal tumors through IGF-1R upregulation. Braz. J. Med. Biol. Res. 2019, 52, e8399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Han, L.; Bao, Z.S.; Wang, Y.Y.; Chen, L.Y.; Yan, W.; Yu, S.Z.; Pu, P.Y.; Liu, N.; You, Y.P.; et al. Chinese Glioma Cooperative Group. HOTAIR, a cell cycle-associated long noncoding RNA and a strong predictor of survival, is preferentially expressed in classical and mesenchymal glioma. Neuro-Oncology 2013, 15, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Sun, X.; Zhou, X.; Han, L.; Chen, L.; Shi, Z.; Zhang, A.; Ye, M.; Wang, Q.; Liu, C.; et al. Long non-coding RNA HOTAIR promotes glioblastoma cell cycle progression in an EZH2 dependent manner. Oncotarget 2015, 6, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Ren, Y.; Zhang, J.; Zhang, C.; Zhang, K.; Han, L.; Kong, L.; Wei, J.; Chen, L.; Yang, J.; et al. HOTAIR is a therapeutic target in glioblastoma. Oncotarget 2015, 6, 8353–8365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, J.; Yao, Y.L.; Zheng, J.; Wang, P.; Liu, Y.H.; Ma, J.; Li, Z.; Liu, X.B.; Li, Z.Q.; Wang, Z.H.; et al. Knockdown of long non-coding RNA HOTAIR inhibits malignant biological behaviors of human glioma cells via modulation of miR-326. Oncotarget 2015, 6, 21934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, E.B.; Ma, C.C.; He, X.J.; Wang, C.; Zong, G.; Wang, H.L.; Zhao, B. Epigenetic modification of miR-141 regulates SKA2 by an endogenous ‘sponge’ HOTAIR in glioma. Oncotarget 2016, 7, 30610–30625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Hodges, T.R.; Song, R.; Gong, Y.; Calin, G.A.; Heimberger, A.B.; Zhao, H. Serum HOTAIR and GAS5 levels as predictors of survival in patients with glioblastoma. Mol. Carcinog. 2018, 57, 137–141. [Google Scholar] [CrossRef]
- Xavier-Magalhães, A.; Oliveira, A.I.; de Castro, J.V.; Pojo, M.; Gonçalves, C.S.; Lourenço, T.; Viana-Pereira, M.; Costa, S.; Linhares, P.; Vaz, R.; et al. Effects of the functional HOTAIR rs920778 and rs12826786 genetic variants in glioma susceptibility and patient prognosis. J. Neuro-Oncol. 2017, 132, 27–34. [Google Scholar] [CrossRef]
- Chakravadhanula, M.; Ozols, V.V.; Hampton, C.N.; Zhou, L.; Catchpoole, D.; Bhardwaj, R.D. Expression of the HOX genes and HOTAIR in atypical teratoid rhabdoid tumors and other pediatric brain tumors. Cancer Genet. 2014, 207, 425–428. [Google Scholar] [CrossRef]
- Zheng, H.; Baranova, K.; Song, J.; Yan, L.; Biswas, S.; Chakrabarti, S.; Zhang, Q. Overexpression of Long Noncoding RNA HOTAIR Is a Unique Epigenetic Characteristic of Myxopapillary Ependymoma. J. Neuropathol. Exp. Neurol. 2020, 79, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, N.; Fu, J.; Zhou, W. Long noncoding RNA HOTAIR promotes medulloblastoma growth, migration and invasion by sponging miR-1/miR-206 and targeting YY1. Biomed. Pharmacother. 2020, 124, 109887. [Google Scholar] [CrossRef] [PubMed]
- Milhem, M.M.; Knutson, T.; Yang, S.; Zhu, D.; Wang, X.; Leslie, K.K.; Meng, X. Correlation of MTDH/AEG-1 and HOTAIR Expression with Metastasis and Response to Treatment in Sarcoma Patients. J. Cancer Sci. Ther. 2011, 5, 004. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Su, Y.; Yang, Q.; Lv, D.; Zhang, W.; Tang, K.; Wang, H.; Zhang, R.; Liu, Y. Overexpression of Long Non-Coding RNA HOTAIR Promotes Tumor Growth and Metastasis in Human Osteosarcoma. Mol. Cells 2015, 38, 432–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Qu, X.L.; Liu, J.; Lu, J.; Zhou, Z.Y. HOTAIR promotes osteosarcoma development by sponging miR-217 and targeting ZEB1. J. Cell Physiol. 2019, 234, 6173–6181. [Google Scholar] [CrossRef]
- Wang, N.; Meng, X.; Liu, Y.; Chen, Y.; Liang, Q. LPS promote Osteosarcoma invasion and migration through TLR4/HOTAIR. Gene 2019, 680, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Chen, F.; Fei, Z.; Zhao, J.; Liang, Y.; Pan, W.; Liu, X.; Zheng, D. Genetic variants of lncRNA HOTAIR contribute to the risk of osteosarcoma. Oncotarget 2016, 7, 19928–19934. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Ren, T.; Huang, Y.; Sun, K.; Wang, S.; Liu, K.; Zheng, B.; Guo, W. Knockdown of long non-coding RNA HOTAIR increases miR-454-3p by targeting Stat3 and Atg12 to inhibit chondrosarcoma growth. Cell Death Dis. 2017, 8, e2605. [Google Scholar] [CrossRef]
- Feng, Q.; Wang, D.; Guo, P.; Zhang, Z.; Feng, J. Long non-coding RNA HOTAIR promotes the progression of synovial sarcoma through microRNA-126/stromal cell-derived factor-1 regulation. Oncol. Lett. 2021, 21, 444. [Google Scholar] [CrossRef]
- Yang, G.; Lu, X.; Yuan, L. LncRNA: A link between RNA and cancer. Biochim. Biophys. Acta 2014, 1839, 1097–1109. [Google Scholar] [CrossRef]
- Peng, W.X.; Koirala, P.; Mo, Y.Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef] [PubMed]
- Botti, G.; Marra, L.; Malzone, M.G.; Anniciello, A.; Botti, C.; Franco, R.; Cantile, M. LncRNA HOTAIR as Prognostic Circulating Marker and Potential Therapeutic Target in Patients with Tumor Diseases. Curr. Drug Targets 2017, 18, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wang, Y.F.; Zhang, J.; Wang, Q.X.; Han, L.; Mei, M.; Kang, C.S. Targeted design and identification of AC1NOD4Q to block activity of HOTAIR by abrogating the scaffold interaction with EZH2. Clin. Epigenet. 2019, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ren, Y.; Wang, Y.; Tan, Y.; Wang, Q.; Cai, J.; Zhou, J.; Yang, C.; Zhao, K.; Yi, K. A Compound AC1Q3QWB Selectively Disrupts HOTAIR-Mediated Recruitment of PRC2 and Enhances Cancer Therapy of DZNep. Theranostics 2019, 9, 4608–4623. [Google Scholar] [CrossRef]

| Tumor Types | Clinical Studies | References | |
|---|---|---|---|
| Head and Neck rare cancers | LSCC | HOTAIR overexpression correlates with poor differentiation, pathological grade, metastatic risk and chemotherapy sensibility. HOTAIR is a circulating markers in LSCC patients. | [127,128] |
| NPC | HOTAIR overexpression correlates with clinical stage and poor prognosis. | [129,130] | |
| SACC | HOTAIR overexpression correlates with clinical stage, nerve invasion, metastatic risk and poor survival. | [131] | |
| Neuroendocrine tumors | GEP NEN | HOTAIR overexpression correlates with tumor grade and tumor stage. | [47] |
| Rare digestive system tumors | Cholangiocarcinoma | HOTAIR overexpression correlates with tumor size, TNM stage and metastatic risk. | [132] |
| GIST | HOTAIR overexpression correlates with tumor grade, recurrence and metastatic risk, poor survival and drug resistance. | [133] | |
| Central Nervous System tumors | Glioma/Glioblastoma | HOTAIR overexpression correlates with tumor grade, molecular subtype and poor prognosis. HOTAIR is a circulating markers in glioma patients. | [134,135] |
| Soft tissue tumors | Osteosarcoma | HOTAIR overexpression correlates with advanced tumor stage, high histological grade and shorter overall survival. | [136] |
| Chondrosarcoma | HOTAIR overexpression correlates with tumor grade and shorter overall survival. | [137] | |
| Synovial sarcoma | HOTAIR overexpression correlates with histological grade, advanced tumor stage and metastatic risk. | [138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liguori, G.; Cerrone, M.; De Chiara, A.; Tafuto, S.; Tracey de Bellis, M.; Botti, G.; Di Bonito, M.; Cantile, M. The Role of lncRNAs in Rare Tumors with a Focus on HOX Transcript Antisense RNA (HOTAIR). Int. J. Mol. Sci. 2021, 22, 10160. https://doi.org/10.3390/ijms221810160
Liguori G, Cerrone M, De Chiara A, Tafuto S, Tracey de Bellis M, Botti G, Di Bonito M, Cantile M. The Role of lncRNAs in Rare Tumors with a Focus on HOX Transcript Antisense RNA (HOTAIR). International Journal of Molecular Sciences. 2021; 22(18):10160. https://doi.org/10.3390/ijms221810160
Chicago/Turabian StyleLiguori, Giuseppina, Margherita Cerrone, Annarosaria De Chiara, Salvatore Tafuto, Maura Tracey de Bellis, Gerardo Botti, Maurizio Di Bonito, and Monica Cantile. 2021. "The Role of lncRNAs in Rare Tumors with a Focus on HOX Transcript Antisense RNA (HOTAIR)" International Journal of Molecular Sciences 22, no. 18: 10160. https://doi.org/10.3390/ijms221810160