
Effects of β-Phenylethylamine on Psychomotor, Rewarding, and Reinforcing Behaviors and Affective State: The Role of Dopamine D1 Receptors

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
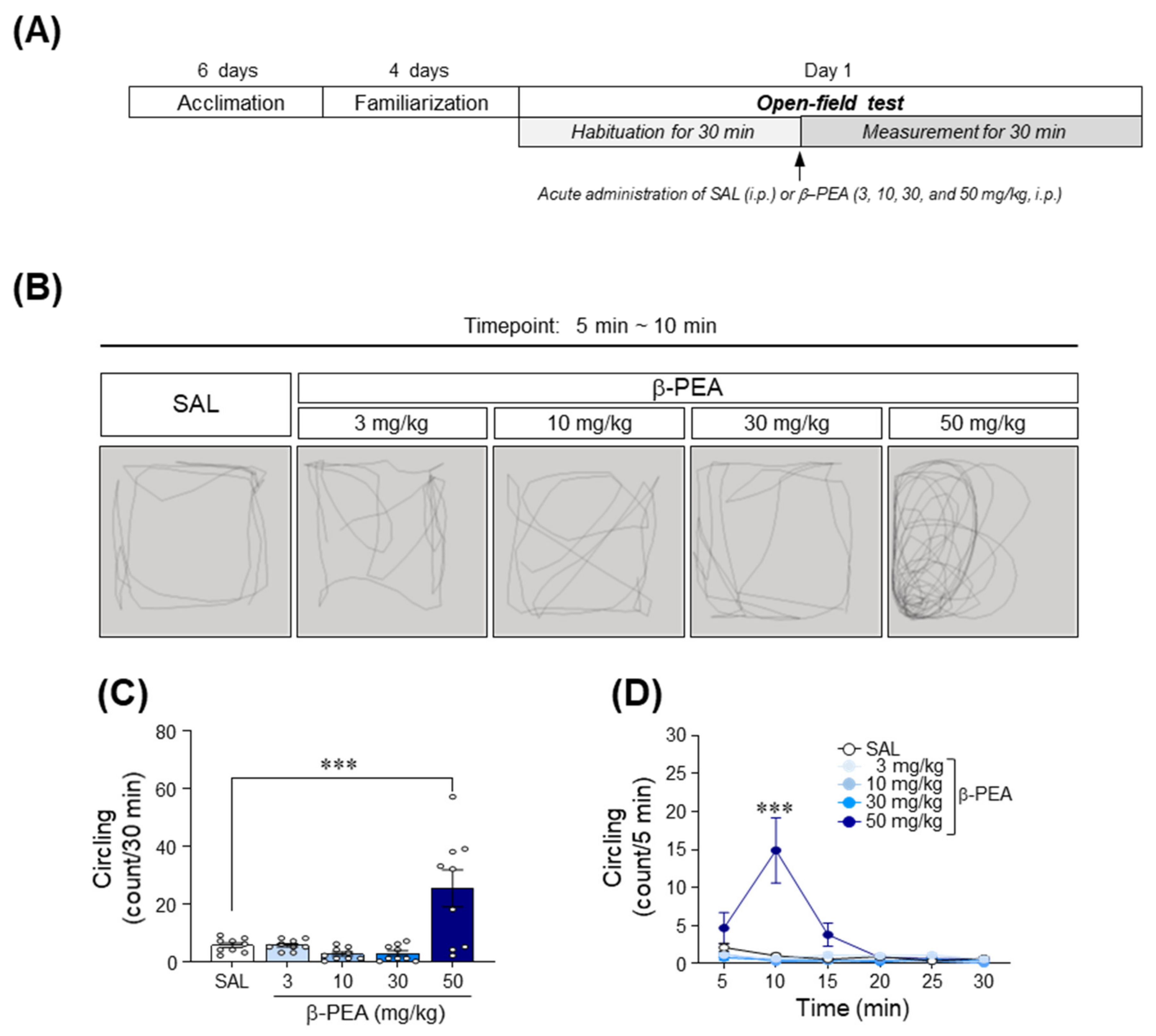
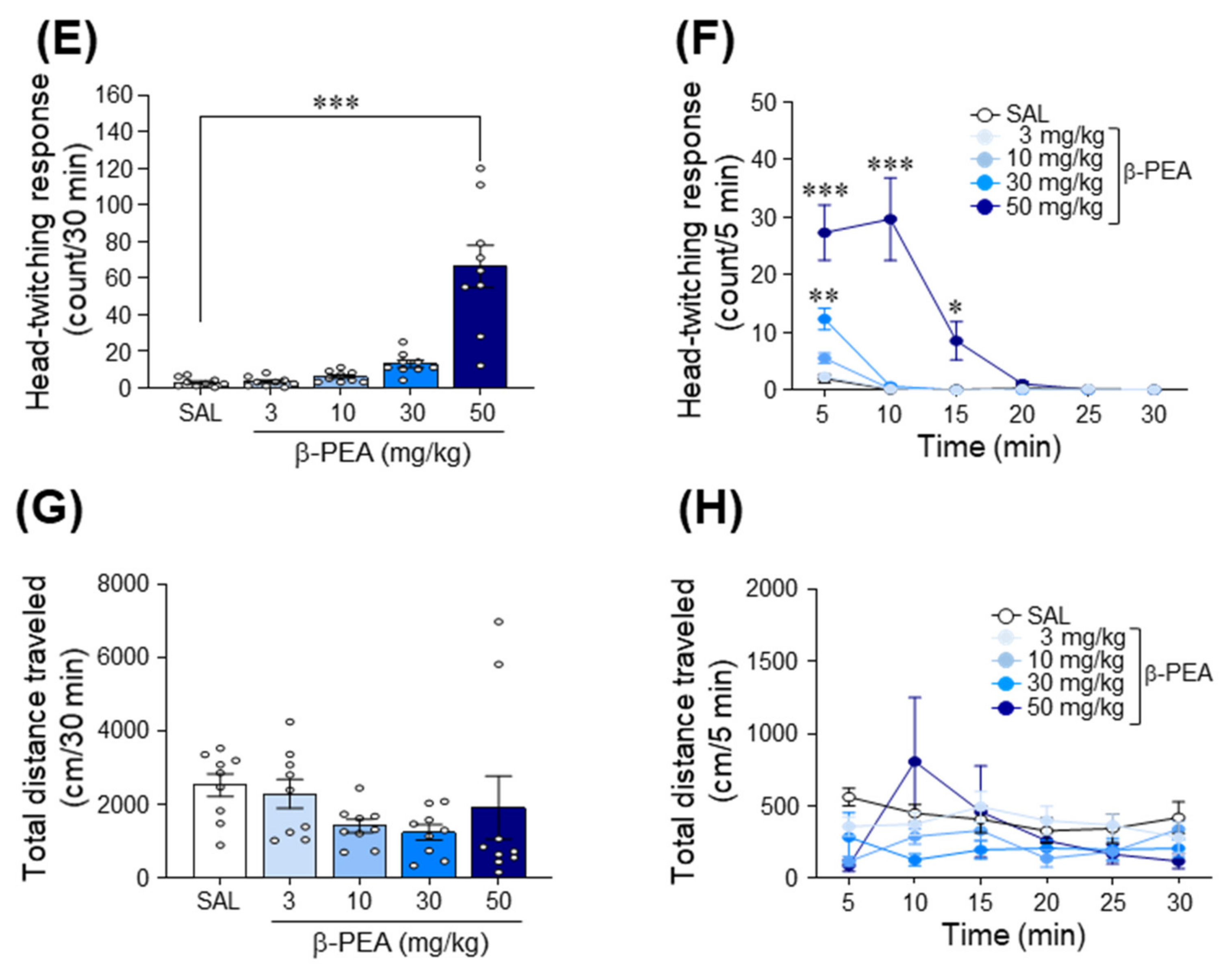
2.1. Acute β-PEA Significantly Increased Circling and Head-Twitching Behaviors in Mice
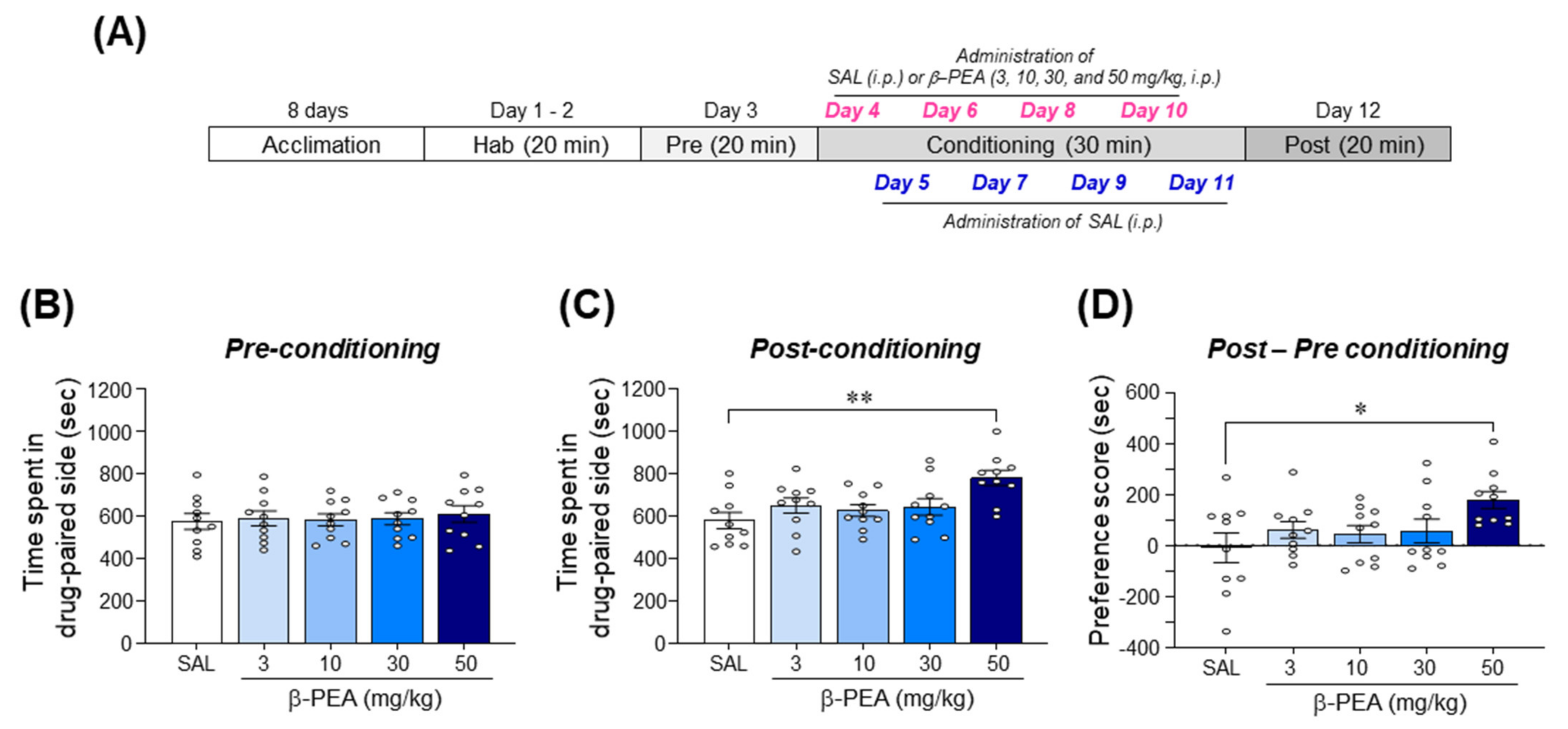
2.2. β-PEA Significantly Produced Conditioned Place Preference in Mice
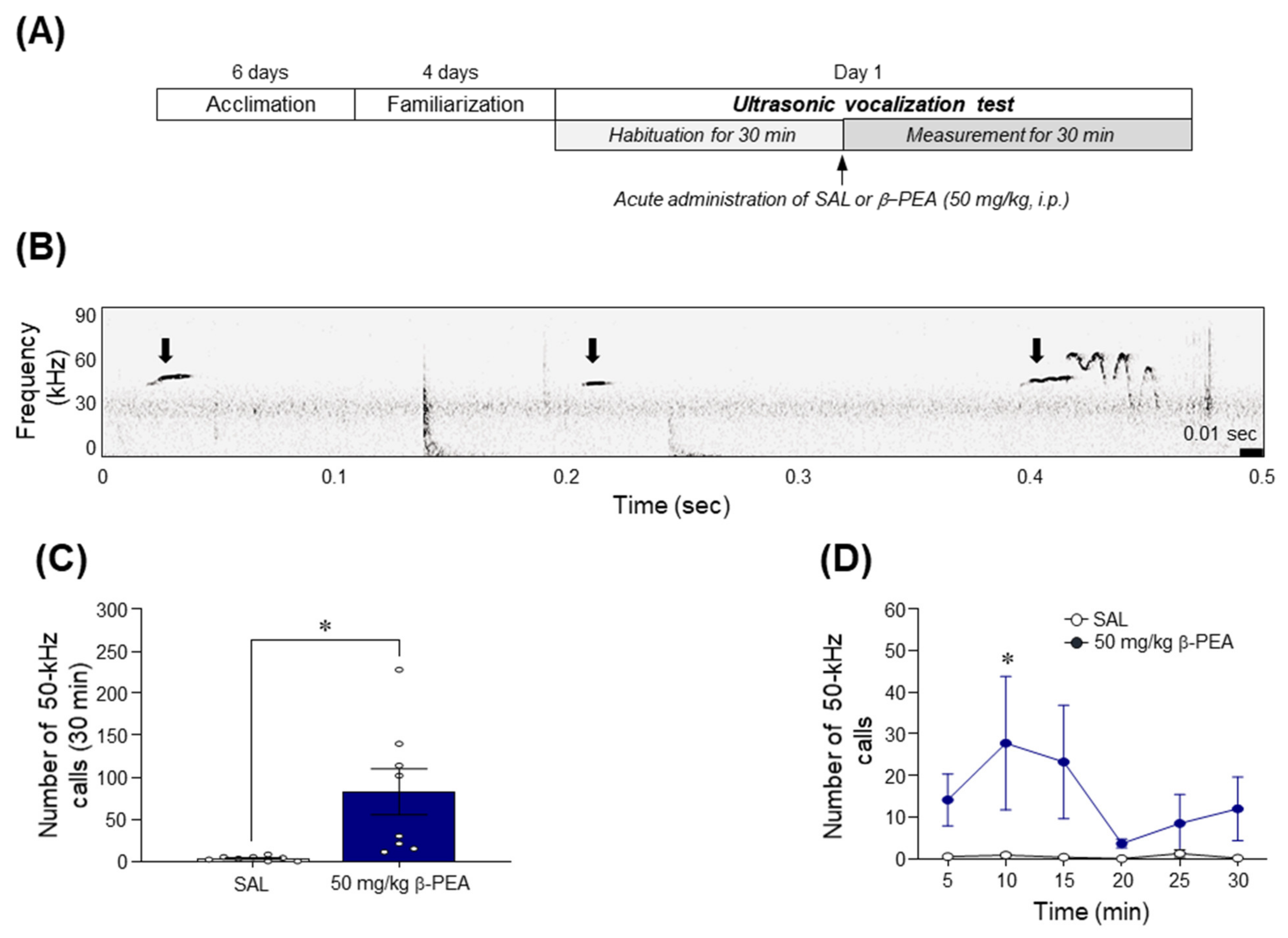
2.3. β-PEA Significantly Increased the 50-kHz Ultrasonic Vocalization Calls in Rats
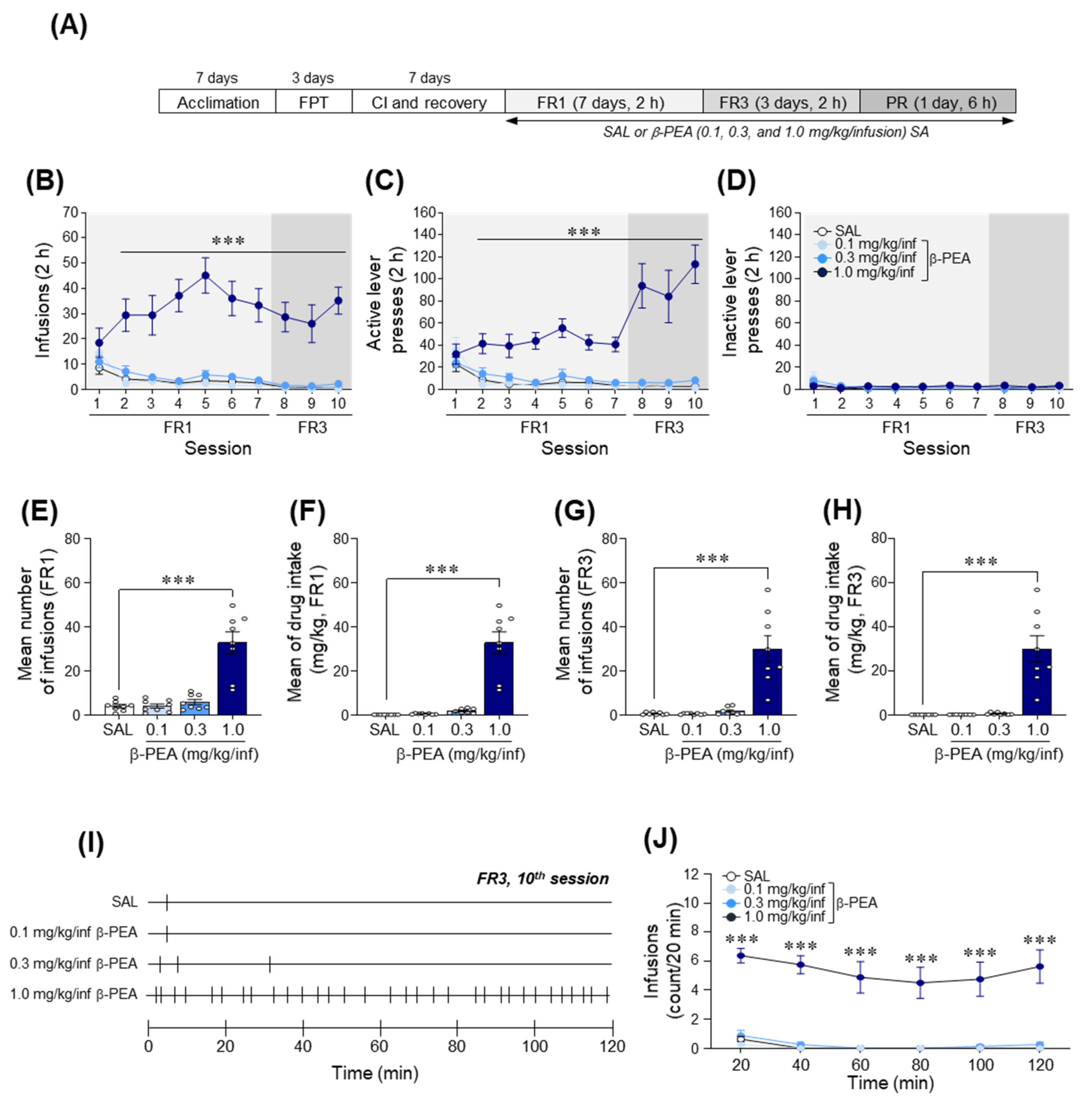
2.4. β-PEA Was Self-Administered under Fixed Ratio Schedules of Reinforcement in Rats
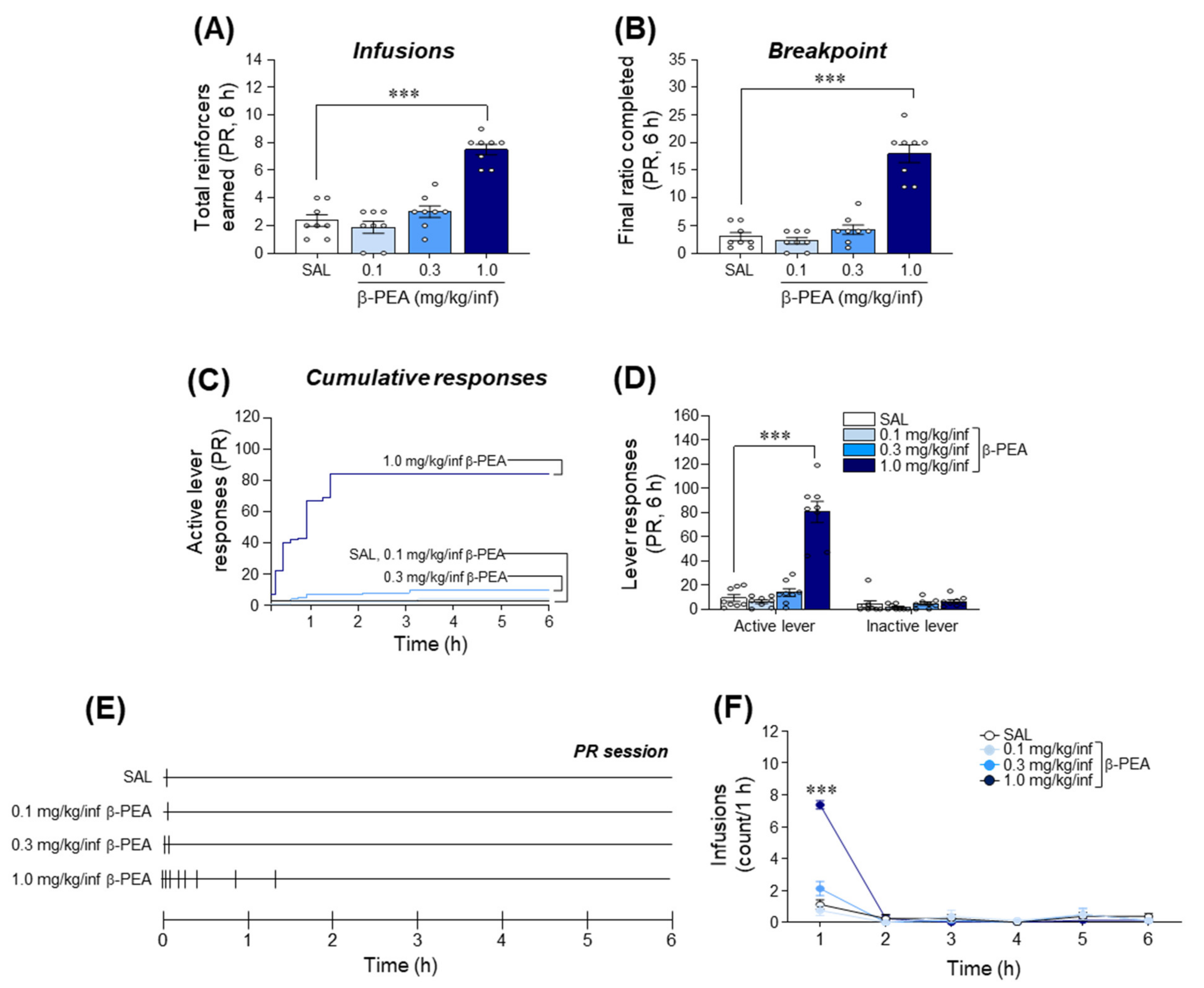
2.5. β-PEA Produced a Higher Breakpoint in Self-Administration under Progressive Ratio Schedules of Reinforcement in Rats
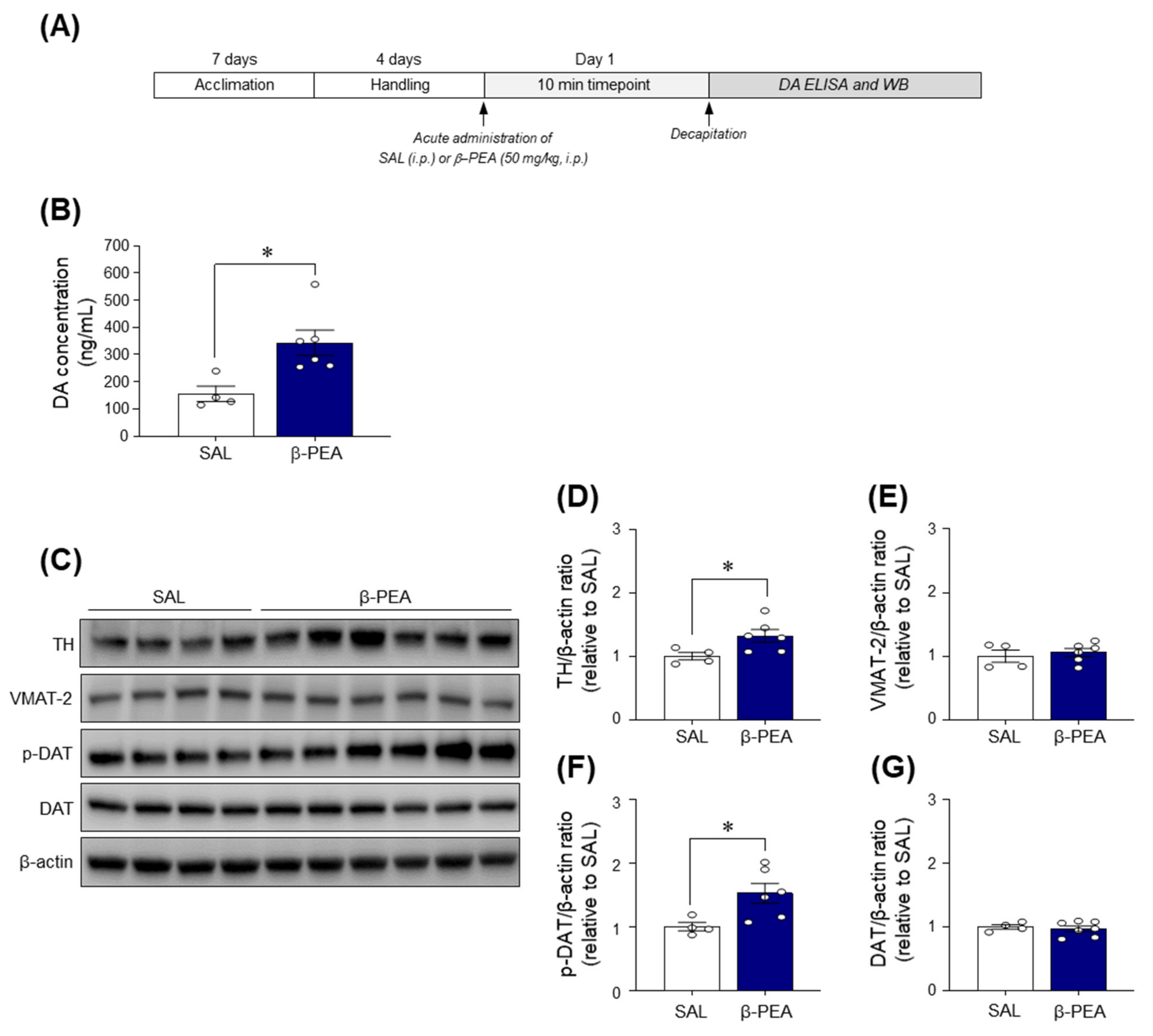
2.6. Acute β-PEA Administration Significantly Increased DA Concentration and p-DAT and TH Expression in the Dorsal Striatum of Mice
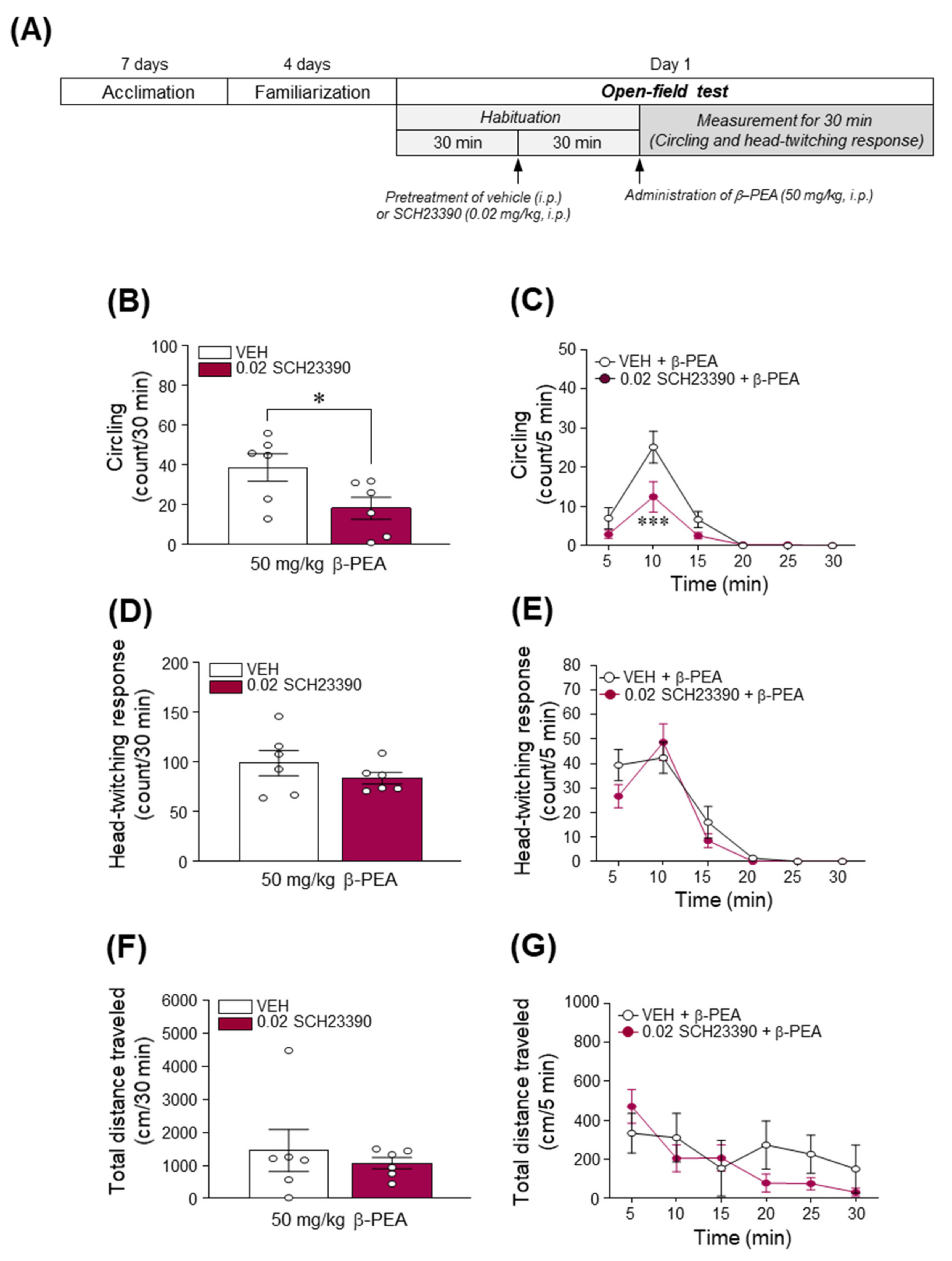
2.7. Blockade of DAD1R Attenuated Acute β-PEA-Induced Circling Behavior in Mice
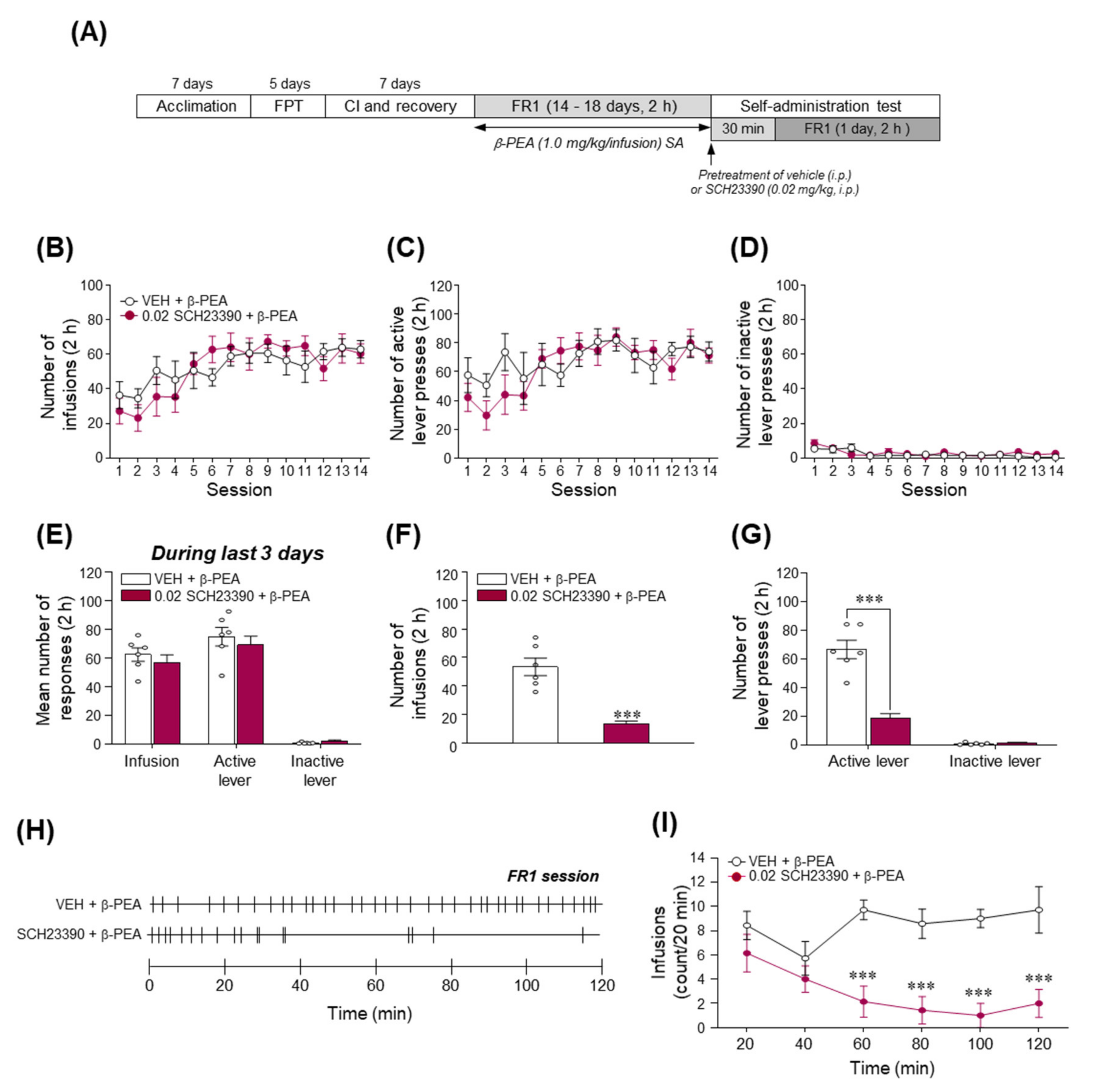
2.8. Blockade of DAD1R Significantly Decreased β-PEA-Taking Behavior
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Open Field Test
4.4. Conditioned Place Preference Test
4.5. Recordings of Ultrasonic Vocalizations
4.6. Food Training and Catheter Implantation Surgery for Self-Administration
4.7. Drug Self-Administration
4.8. Tissue Collection for DA ELISA and Western Blotting
4.9. Dopamine ELISA
4.10. Western Immunoblotting
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Philips, S.R.; Rozdilsky, B.; Boulton, A.A. Evidence for the presence of m-tyramine, p-tyramine, tryptamine, and phenylethylamine in the rat brain and several areas of the human brain. Biol. Psychiatry 1978, 13, 51–57. [Google Scholar]
- Paterson, I.A.; Juorio, A.V.; Boulton, A.A. 2-Phenylethylamine: A modulator of catecholamine transmission in the mammalian central nervous system? J. Neurochem. 1990, 55, 1827–1837. [Google Scholar] [CrossRef]
- Berry, M.D. Mammalian central nervous system trace amines. Pharmacologic amphetamines, physiologic neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Dyck, L.E.; Yang, C.R.; Boulton, A.A. The biosynthesis of p-tyramine, m-tyramine, and beta-phenylethylamine by rat striatal slices. J. Neurosci. Res. 1983, 10, 211–220. [Google Scholar] [CrossRef]
- Murata, M.; Katagiri, N.; Ishida, K.; Abe, K.; Ishikawa, M.; Utsunomiya, I.; Hoshi, K.; Miyamoto, K.; Taguchi, K. Effect of beta-phenylethylamine on extracellular concentrations of dopamine in the nucleus accumbens and prefrontal cortex. Brain Res. 2009, 1269, 40–46. [Google Scholar] [CrossRef]
- Zsilla, G.; Hegyi, D.E.; Baranyi, M.; Vizi, E.S. 3,4-Methylenedioxymethamphetamine, mephedrone, and beta-phenylethylamine release dopamine from the cytoplasm by means of transporters and keep the concentration high and constant by blocking reuptake. Eur. J. Pharmacol. 2018, 837, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Wickramasekara, R.N.; Carvelli, L. Beta-Phenylethylamine requires the dopamine transporter to increase extracellular dopamine in Caenorhabditis elegans dopaminergic neurons. Neurochem. Int. 2014, 73, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Sotnikova, T.D.; Budygin, E.A.; Jones, S.R.; Dykstra, L.A.; Caron, M.G.; Gainetdinov, R.R. Dopamine transporter-dependent and -independent actions of trace amine beta-phenylethylamine. J. Neurochem. 2004, 91, 362–373. [Google Scholar] [CrossRef]
- Nakamura, M.; Ishii, A.; Nakahara, D. Characterization of beta-phenylethylamine-induced monoamine release in rat nucleus accumbens: A microdialysis study. Eur. J. Pharmacol. 1998, 349, 163–169. [Google Scholar] [CrossRef]
- Safratowich, B.D.; Hossain, M.; Bianchi, L.; Carvelli, L. Amphetamine potentiates the effects of beta-phenylethylamine through activation of an amine-gated chloride channel. J. Neurosci. 2014, 34, 4686–4691. [Google Scholar] [CrossRef] [Green Version]
- Iversen, L.; White, M.; Treble, R. Designer psychostimulants: Pharmacology and differences. Neuropharmacology 2014, 87, 59–65. [Google Scholar] [CrossRef]
- Greenshaw, A.J. β-Phenylethylamine and reinforcement. Prog. Neuropsychopharmacol. Biol. Psychiatry 1984, 8, 615–620. [Google Scholar] [CrossRef]
- Gilbert, D.; Cooper, S.J. Beta-Phenylethylamine-, d-amphetamine-and l-amphetamine-induced place preference conditioning in rats. Eur. J. Pharmacol. 1983, 95, 311–314. [Google Scholar] [CrossRef]
- Bergman, J.; Yasar, S.; Winger, G. Psychomotor stimulant effects of beta-phenylethylamine in monkeys treated with MAO-B inhibitors. Psychopharmacology 2001, 159, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.; Simmons, S.J.; West, M.O. Ultrasonic Vocalizations as a Measure of Affect in Preclinical Models of Drug Abuse: A Review of Current Findings. Curr. Neuropharmacol. 2015, 13, 193–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F. Drug Addiction: The Yin and Yang of Hedonic Homeostasis. Neuron 1996, 16, 893–896. [Google Scholar] [CrossRef] [Green Version]
- Koo, J.W.; Han, J.-S.; Kim, J.J. Selective Neurotoxic Lesions of Basolateral and Central Nuclei of the Amygdala Produce Differential Effects on Fear Conditioning. J. Neurosci. 2004, 24, 7654–7662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covington, H.E., 3rd; Miczek, K.A. Vocalizations during withdrawal from opiates and cocaine: Possible expressions of affective distress. Eur. J. Pharmacol. 2003, 467, 1–13. [Google Scholar] [CrossRef]
- Burgdorf, J.; Knutson, B.; Panksepp, J. Anticipation of rewarding electrical brain stimulation evokes ultrasonic vocalization in rats. Behav. Neurosci. 2000, 114, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.M.; Gourdon, J.C.; Clarke, P.B.S. Identification of multiple call categories within the rich repertoire of adult rat 50-kHz ultrasonic vocalizations: Effects of amphetamine and social context. Psychopharmacology 2010, 211, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kuchniak, K.; Wyszogrodzka, E.; Chrapusta, S.J.; Czarna, M.; Michalak, M.; Płaźnik, A.; Krząścik, P.; Mierzejewski, P.; Taracha, E. Using anticipatory and drug-evoked appetitive ultrasonic vocalization for monitoring the rewarding effect of amphetamine in a rat model of drug self-administration. Behav. Brain Res. 2019, 376, 112187. [Google Scholar] [CrossRef] [PubMed]
- Mahler, S.V.; Moorman, D.E.; Feltenstein, M.W.; Cox, B.M.; Ogburn, K.B.; Bachar, M.; McGonigal, J.T.; Ghee, S.M.; See, R.E. A rodent “self-report” measure of methamphetamine craving? Rat ultrasonic vocalizations during methamphetamine self-administration, extinction, and reinstatement. Behav. Brain. Res. 2013, 236, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Costa, G.; Morelli, M.; Simola, N. Involvement of Glutamate NMDA Receptors in the Acute, Long-Term, and Conditioned Effects of Amphetamine on Rat 50 kHz Ultrasonic Vocalizations. Int. J. Neuropsychopharmacol. 2015, 18, pyv057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.N.; Undieh, A.S. Dopamine-sensitive signaling mediators modulate psychostimulant-induced ultrasonic vocalization behavior in rats. Behav. Brain Res. 2015, 296, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Yager, L.; Garcia, A.; Wunsch, A.; Ferguson, S. The ins and outs of the striatum: Role in drug addiction. Neuroscience 2015, 301, 529–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, K.; Robinson, T.E. What is the role of dopamine in reward: Hedonic impact, reward learning, or incentive salience? Brain Res. Rev. 1998, 28, 309–369. [Google Scholar] [CrossRef]
- Spanagel, R.; Weiss, F. The dopamine hypothesis of reward: Past and current status. Trends Neurosci. 1999, 22, 521–527. [Google Scholar] [CrossRef]
- Vezina, P. D1 dopamine receptor activation is necessary for the induction of sensitization by amphetamine in the ventral tegmental area. J. Neurosci. 1996, 16, 2411–2420. [Google Scholar] [CrossRef]
- Aguirre, N.; Ramos, M.; Goñi-Allo, B. Studies on the role of dopamine D1 receptors in the development and expression of MDMA-induced behavioral sensitization in rats. Psychopharmacology 2004, 177, 100–110. [Google Scholar] [CrossRef]
- Crawford, C.; Drago, J.; Watson, J.B.; Levine, M.S. Effects of repeated amphetamine treatment on the locomotor activity of the dopamine D1A-deficient mouse. NeuroReport 1997, 8, 2523–2527. [Google Scholar] [CrossRef] [PubMed]
- Nazari-Serenjeh, F.; Jamali, S.; Rezaee, L.; Zarrabian, S.; Haghparast, A. D1- but not D2-like dopamine receptor antagonist in the CA1 region of the hippocampus reduced stress-induced reinstatement in extinguished morphine-conditioning place preference in the food-deprived rats. Behav. Pharmacol. 2020, 31, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Bari, A.; Pierce, R. D1-like and D2 dopamine receptor antagonists administered into the shell subregion of the rat nucleus accumbens decrease cocaine, but not food, reinforcement. Neuroscience 2005, 135, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Corominas-Roso, M.; Roncero, C.; Bruguera, E.; Casas, M. The dopaminergic system and addictions. Rev. Neurol. 2007, 44, 23–31. [Google Scholar] [PubMed]
- Robinson, T.E.; Berridge, K.C. The neural basis of drug craving: An incentive-sensitization theory of addiction. Brain Res. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar] [CrossRef]
- Foster, J.D.; Yang, J.W.; Moritz, A.E.; Challasivakanaka, S.; Smith, M.A.; Holy, M.; Wilebski, K.; Sitte, H.H.; Vaughan, R.A. Dopamine transporter phosphorylation site threonine 53 regulates substrate reuptake and amphetamine-stimulated efflux. J. Biol. Chem. 2012, 287, 29702–29712. [Google Scholar] [CrossRef] [Green Version]
- Challasivakanaka, S.; Zhen, J.; Smith, M.E.; Reith, M.E.; Foster, J.D.; Vaughan, R.A. Dopamine transporter phosphorylation site threonine 53 is stimulated by amphetamines and regulates dopamine transport, efflux, and cocaine analog binding. J. Biol. Chem. 2017, 292, 19066–19075. [Google Scholar] [CrossRef] [Green Version]
- Wojtas, A.; Herian, M.; Skawski, M.; Sobocińska, M.; González-Marín, A.; Noworyta-Sokołowska, K.; Gołembiowska, K. Neurochemical and Behavioral Effects of a New Hallucinogenic Compound 25B-NBOMe in Rats. Neurotox. Res. 2020, 39, 305–326. [Google Scholar] [CrossRef]
- Herian, M.; Wojtas, A.; Kamińska, K.; Świt, P.; Wach, A.; Gołembiowska, K. Hallucinogen-Like Action of the Novel Designer Drug 25I-NBOMe and Its Effect on Cortical Neurotransmitters in Rats. Neurotox. Res. 2019, 36, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Gatch, M.; Dolan, S.; Forster, M.J. Locomotor and discriminative stimulus effects of four novel hallucinogens in rodents. Behav. Pharmacol. 2017, 28, 375–385. [Google Scholar] [CrossRef]
- Morency, M.A.; Stewart, R.J.; Beninger, R.J. Circling behavior following unilateral microinjections of cocaine into the medial prefrontal cortex: Dopaminergic or local anesthetic effect? J. Neurosci. 1987, 7, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, K.-Y.; Lewis, B.L.; Lipska, B.K.; O’Donnell, P. Post-Pubertal Disruption of Medial Prefrontal Cortical Dopamine–Glutamate Interactions in a Developmental Animal Model of Schizophrenia. Biol. Psychiatry 2007, 62, 730–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardo, M.T.; Bevins, R.A. Conditioned place preference: What does it add to our preclinical understanding of drug reward? Psychopharmacology 2000, 153, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Huskinson, S.L.; Naylor, J.E.; Rowlett, J.K.; Freeman, K.B. Predicting abuse potential of stimulants and other dopaminergic drugs: Overview and recommendations. Neuropharmacology 2014, 87, 66–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.-Y.; Hur, K.-H.; Ko, Y.-H.; Kim, K.; Lee, B.-R.; Kim, Y.-J.; Kim, S.-K.; Lee, Y.-S.; Kim, H.-C.; Lee, S.-Y.; et al. A novel designer drug, 25N-NBOMe, exhibits abuse potential via the dopaminergic system in rodents. Brain Res. Bull. 2019, 152, 19–26. [Google Scholar] [CrossRef]
- Custodio, R.J.P.; Sayson, L.V.; Botanas, C.J.; Abiero, A.; You, K.Y.; Kim, M.; Lee, H.J.; Yoo, S.Y.; Lee, K.W.; Lee, Y.S.; et al. 25B-NBOMe, a novel N -2-methoxybenzyl-phenethylamine (NBOMe) derivative, may induce rewarding and reinforcing effects via a dopaminergic mechanism: Evidence of abuse potential. Addict. Biol. 2019, 25, e12850. [Google Scholar] [CrossRef] [PubMed]
- Richardson, N.R.; Roberts, D. Progressive ratio schedules in drug self-administration studies in rats: A method to evaluate reinforcing efficacy. J. Neurosci. Methods 1996, 66, 1–11. [Google Scholar] [CrossRef]
- Abiero, A.; Botanas, C.J.; Custodio, R.J.; Sayson, L.V.; Kim, M.; Lee, H.J.; Kim, H.J.; Lee, K.W.; Jeong, Y.; Seo, J.-W.; et al. 4-MeO-PCP and 3-MeO-PCMo, new dissociative drugs, produce rewarding and reinforcing effects through activation of mesolimbic dopamine pathway and alteration of accumbal CREB, deltaFosB, and BDNF levels. Psychopharmacology 2019, 237, 757–772. [Google Scholar] [CrossRef]
- Ryu, I.S.; Yoon, S.S.; Lee, Y.E.; Kim, W.H.; Kim, J.S.; Cheong, J.H.; Kim, H.J.; Jang, C.-G.; Lee, Y.S.; Ka, M.; et al. The potent psychomotor, rewarding and reinforcing properties of 3-fluoromethamphetamine in rodents. IBRO Rep. 2019, 6, S236. [Google Scholar] [CrossRef]
- Mu, P.; Fuchs, T.; Saal, D.B.; Sorg, B.; Dong, Y.; Panksepp, J. Repeated cocaine exposure induces sensitization of ultrasonic vocalization in rats. Neurosci. Lett. 2009, 453, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.J.; Ryu, Y.; Lee, B.H.; Chang, S.; Fan, Y.; Gwak, Y.S.; Yang, C.H.; Bills, K.B.; Steffensen, S.C.; Koo, J.S.; et al. Acupuncture inhibition of methamphetamine-induced behaviors, dopamine release and hyperthermia in the nucleus accumbens: Mediation of group II mGluR. Addict. Biol. 2018, 24, 206–217. [Google Scholar] [CrossRef]
- Simola, N.; Brudzynski, S.M. Rat 50-kHz ultrasonic vocalizations as a tool in studying neurochemical mechanisms that regulate positive emotional states. J. Neurosci. Methods 2018, 310, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.M.; Dobosiewicz, M.; Clarke, P.B.S. The role of dopaminergic transmission through D1-like and D2-like receptors in amphetamine-induced rat ultrasonic vocalizations. Psychopharmacology 2012, 225, 853–868. [Google Scholar] [CrossRef]
- London, E.D. Impulsivity, Stimulant Abuse, and Dopamine Receptor Signaling. Adv. Pharmacol. 2016, 76, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Serra, M.; Marongiu, J.; Morelli, M.; Simola, N. Influence of dopamine transmission in the medial prefrontal cortex and dorsal striatum on the emission of 50-kHz ultrasonic vocalizations in rats treated with amphetamine: Effects on drug-stimulated and conditioned calls. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 97, 109797. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, I.; Carino, M.; Horita, A. Involvement of D1 and D2 dopamine systems in the behavioral effects of cocaine in rats. Pharmacol. Biochem. Behav. 1995, 52, 737–741. [Google Scholar] [CrossRef]
- Kita, K.; Shiratani, T.; Takenouchi, K.; Fukuzako, H.; Takigawa, M. Effects of D1 and D2 dopamine receptor antagonists on cocaine-induced self-stimulation and locomotor activity in rats. Eur. Neuropsychopharmacol. 1999, 9, 1–7. [Google Scholar] [CrossRef]
- Liao, R.-M.; Chang, Y.H.; Wang, S.H. Influence of SCH23390 and spiperone on the expression of conditioned place preference induced by d-amphetamine or cocaine in the rat. Chin. J. Physiol. 1998, 41, 85–92. [Google Scholar] [PubMed]
- Caine, S.B.; Thomsen, M.; Gabriel, K.; Berkowitz, J.S.; Gold, L.H.; Koob, G.F.; Tonegawa, S.; Zhang, J.; Xu, M. Lack of Self-Administration of Cocaine in Dopamine D1 Receptor Knock-Out Mice. J. Neurosci. 2007, 27, 13140–13150. [Google Scholar] [CrossRef]
- Awasaki, Y.; Nishida, N.; Sasaki, S.; Sato, S. Dopamine D1 antagonist SCH23390 attenuates self-administration of both cocaine and fentanyl in rats. Environ. Toxicol. Pharmacol. 1997, 3, 115–122. [Google Scholar] [CrossRef]
- Avchalumov, Y.; Trenet, W.; Piña-Crespo, J.; Mandyam, C. SCH23390 Reduces Methamphetamine Self-Administration and Prevents Methamphetamine-Induced Striatal LTD. Int. J. Mol. Sci. 2020, 21, 6491. [Google Scholar] [CrossRef]
- Brennan, K.A.; Carati, C.; Lea, R.; Fitzmaurice, P.; Schenk, S. Effect of D1-like and D2-like receptor antagonists on methamphetamine and 3,4-methylenedioxymethamphetamine self-administration in rats. Behav. Pharmacol. 2009, 20, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Ryu, I.S.; Kim, O.-H.; Lee, Y.E.; Kim, J.S.; Li, Z.-H.; Kim, T.W.; Lim, R.-N.; Lee, Y.J.; Cheong, J.H.; Kim, H.J.; et al. The Abuse Potential of Novel Synthetic Phencyclidine Derivative 1-(1-(4-Fluorophenyl)Cyclohexyl)Piperidine (4′-F-PCP) in Rodents. Int. J. Mol. Sci. 2020, 21, 4631. [Google Scholar] [CrossRef] [PubMed]
- DiPalma, D.; Rezvani, A.H.; Willette, B.; Wells, C.; Slade, S.; Hall, B.J.; Levin, E.D. Persistent attenuation of nicotine self-administration in rats by co-administration of chronic nicotine infusion with the dopamine D1 receptor antagonist SCH-23390 or the serotonin 5-HT2C agonist lorcaserin. Pharmacol. Biochem. Behav. 2018, 176, 16–22. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, I.S.; Kim, O.-H.; Kim, J.S.; Sohn, S.; Choe, E.S.; Lim, R.-N.; Kim, T.W.; Seo, J.-W.; Jang, E.Y. Effects of β-Phenylethylamine on Psychomotor, Rewarding, and Reinforcing Behaviors and Affective State: The Role of Dopamine D1 Receptors. Int. J. Mol. Sci. 2021, 22, 9485. https://doi.org/10.3390/ijms22179485
Ryu IS, Kim O-H, Kim JS, Sohn S, Choe ES, Lim R-N, Kim TW, Seo J-W, Jang EY. Effects of β-Phenylethylamine on Psychomotor, Rewarding, and Reinforcing Behaviors and Affective State: The Role of Dopamine D1 Receptors. International Journal of Molecular Sciences. 2021; 22(17):9485. https://doi.org/10.3390/ijms22179485
Chicago/Turabian StyleRyu, In Soo, Oc-Hee Kim, Ji Sun Kim, Sumin Sohn, Eun Sang Choe, Ri-Na Lim, Tae Wan Kim, Joung-Wook Seo, and Eun Young Jang. 2021. "Effects of β-Phenylethylamine on Psychomotor, Rewarding, and Reinforcing Behaviors and Affective State: The Role of Dopamine D1 Receptors" International Journal of Molecular Sciences 22, no. 17: 9485. https://doi.org/10.3390/ijms22179485