
PGRS Domain of Rv0297 of Mycobacterium tuberculosis Functions in A Calcium Dependent Manner

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
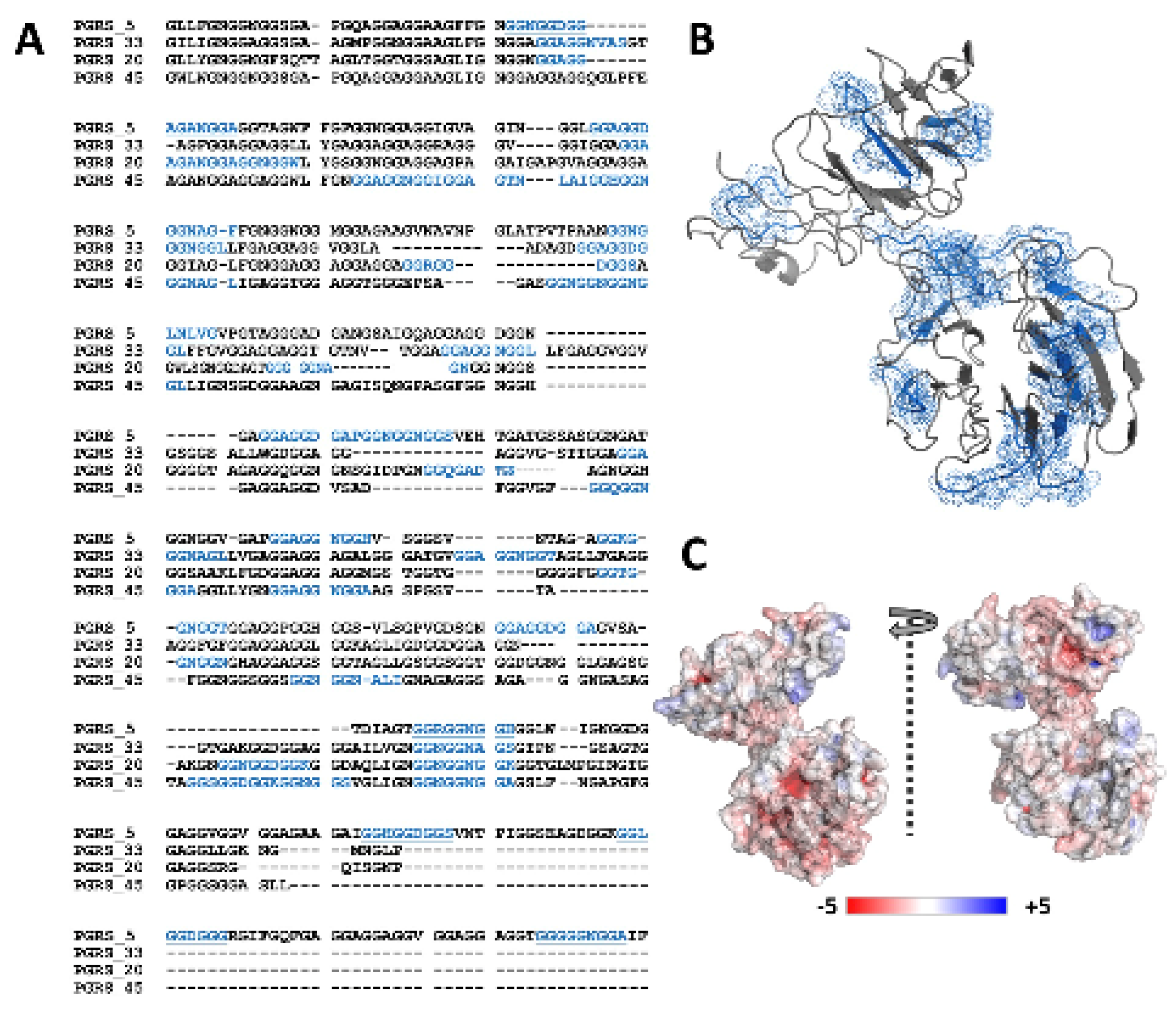
2.1. PGRS Domain of M.tb Rv0297 Contains Putative Calcium-Binding Motifs
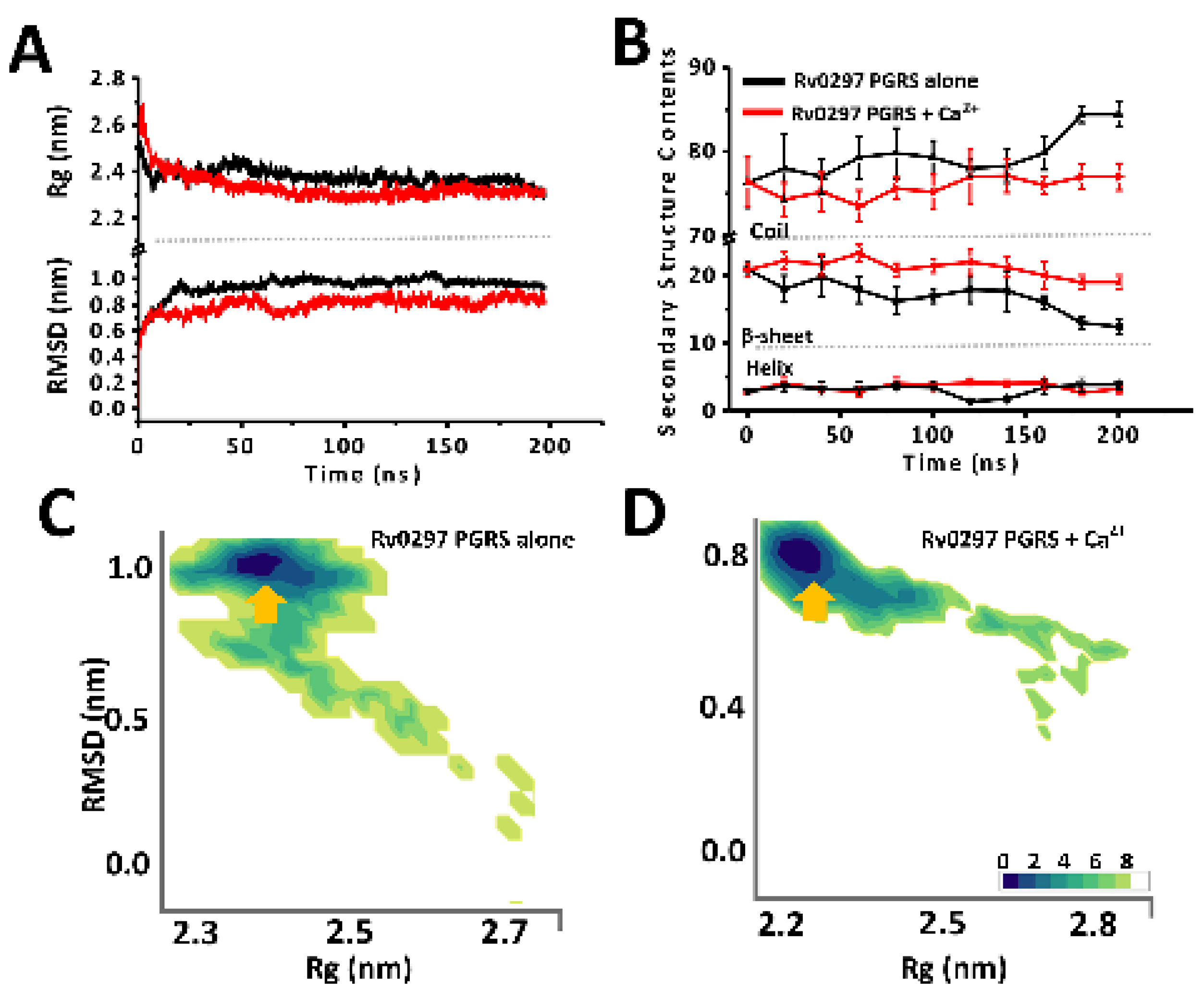
2.2. Ca2+ Binding Stabilizes PGRS Domain Architecture
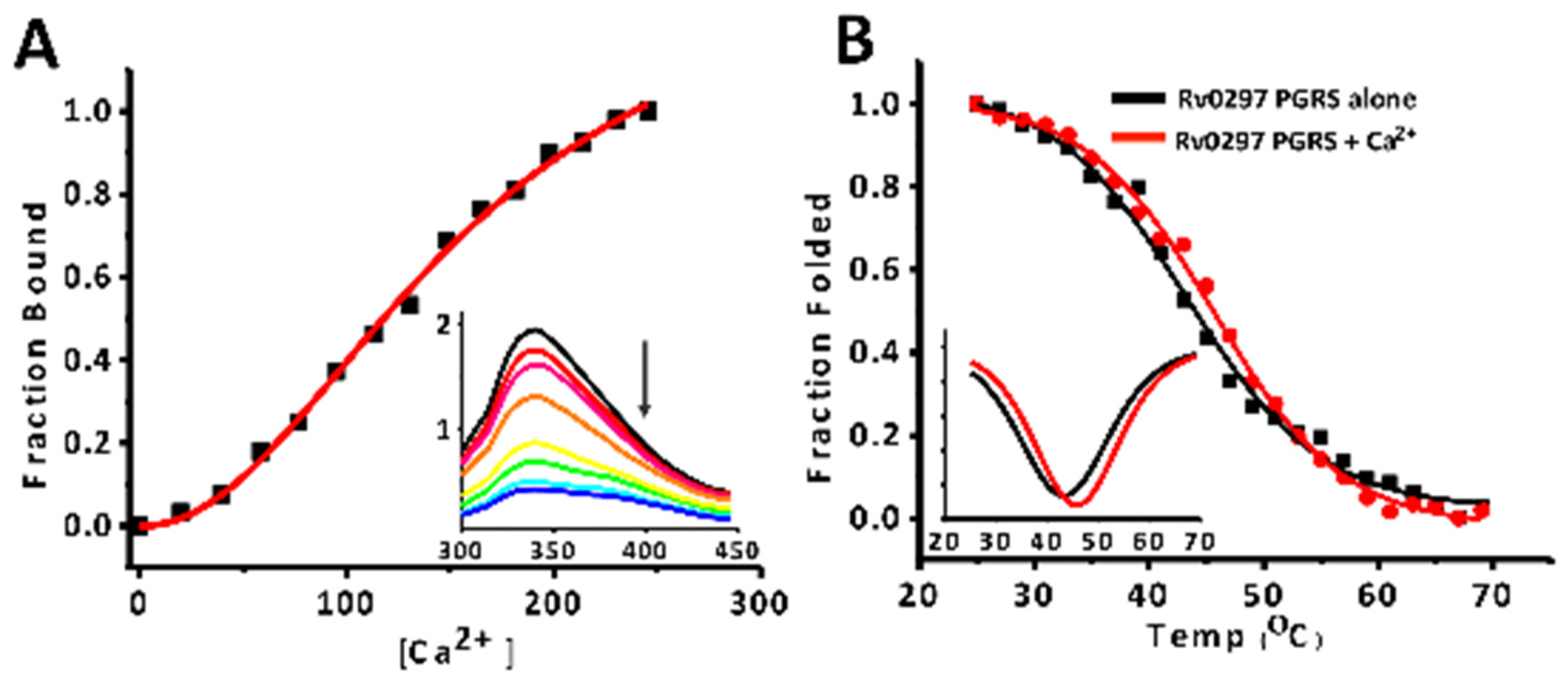
2.3. Calcium Binds PGRS Domain with Micromolar Affinity
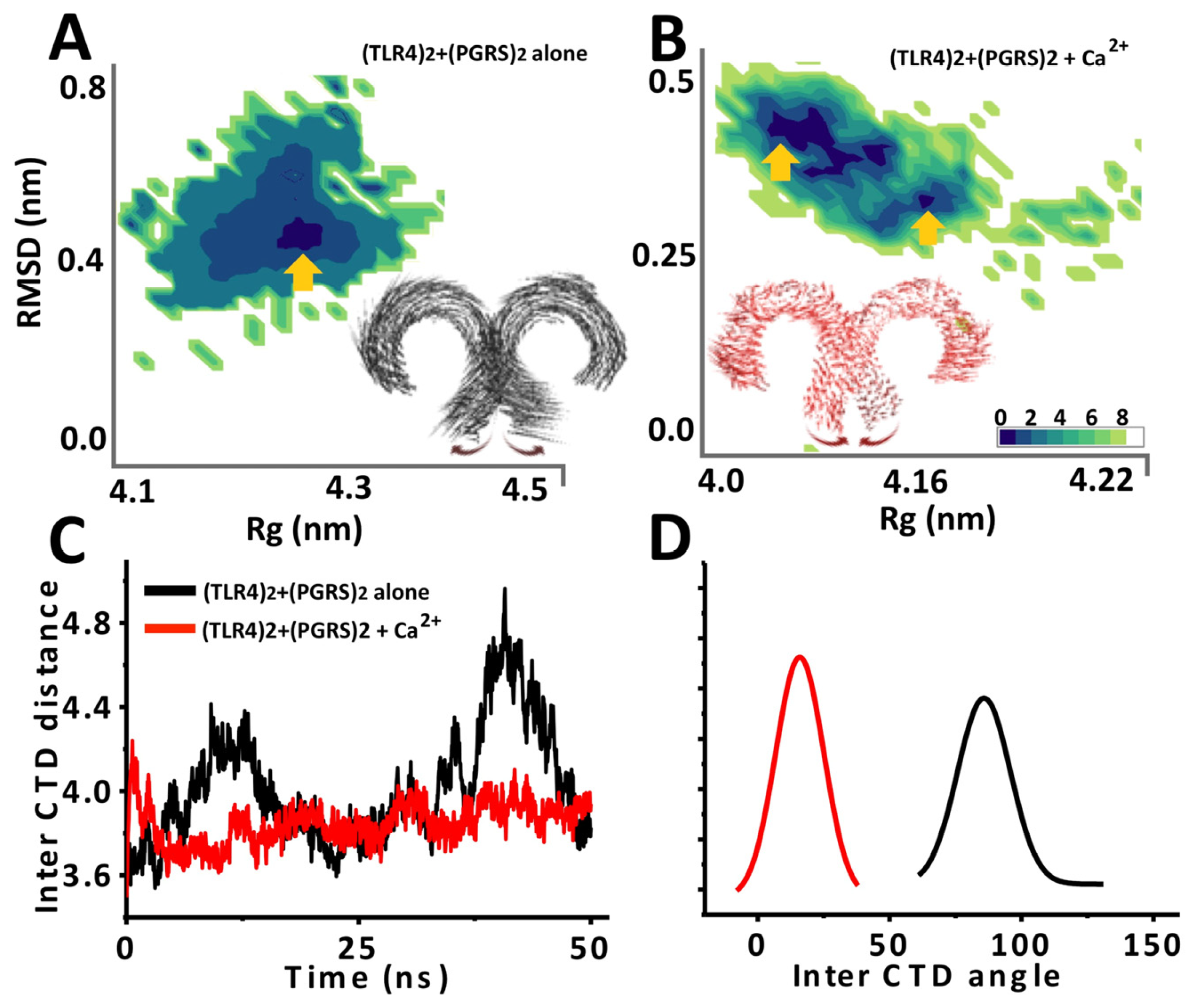
2.4. Ca2+ Bound PGRS also Stabilizes TLR4 Architecture
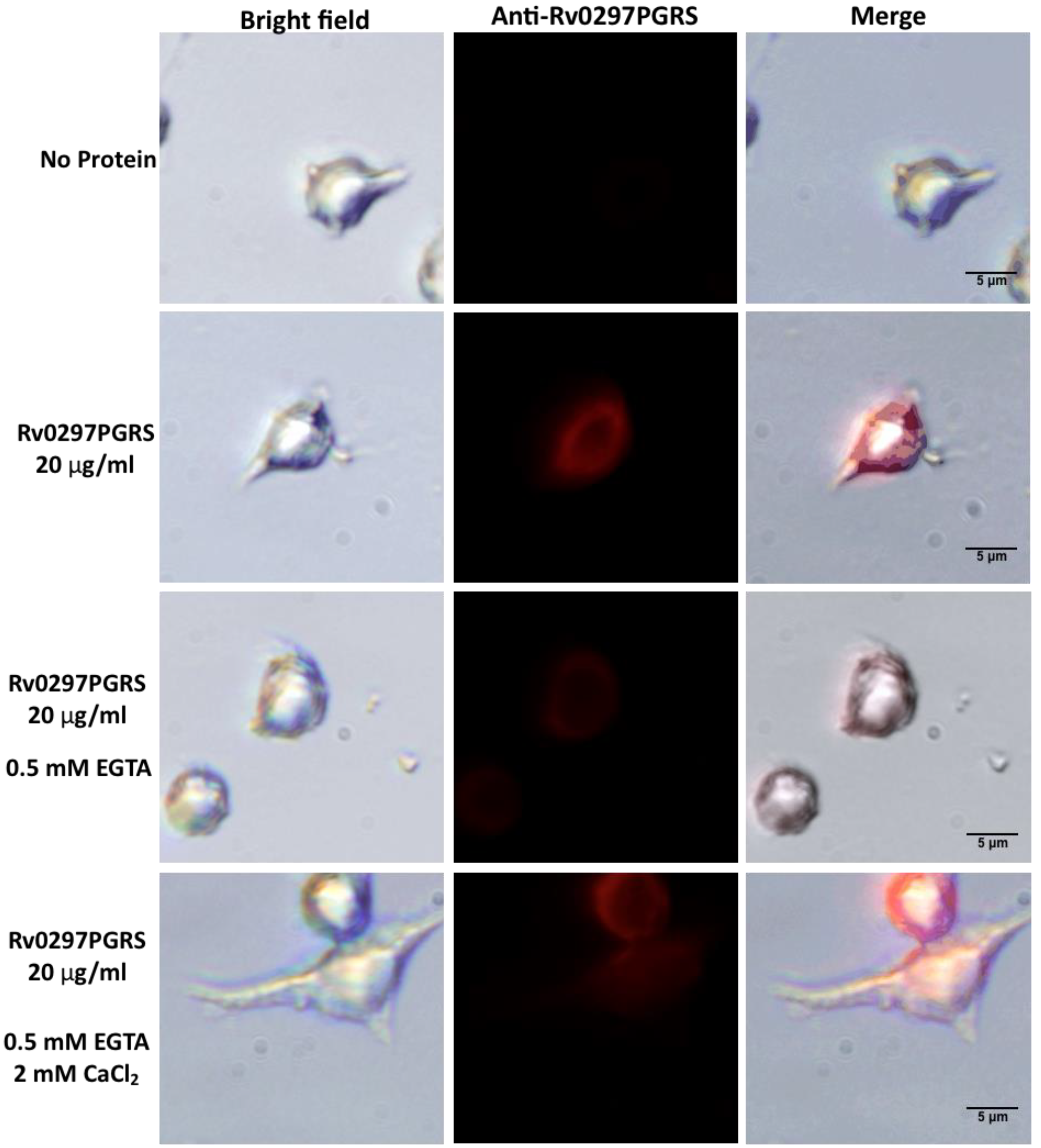
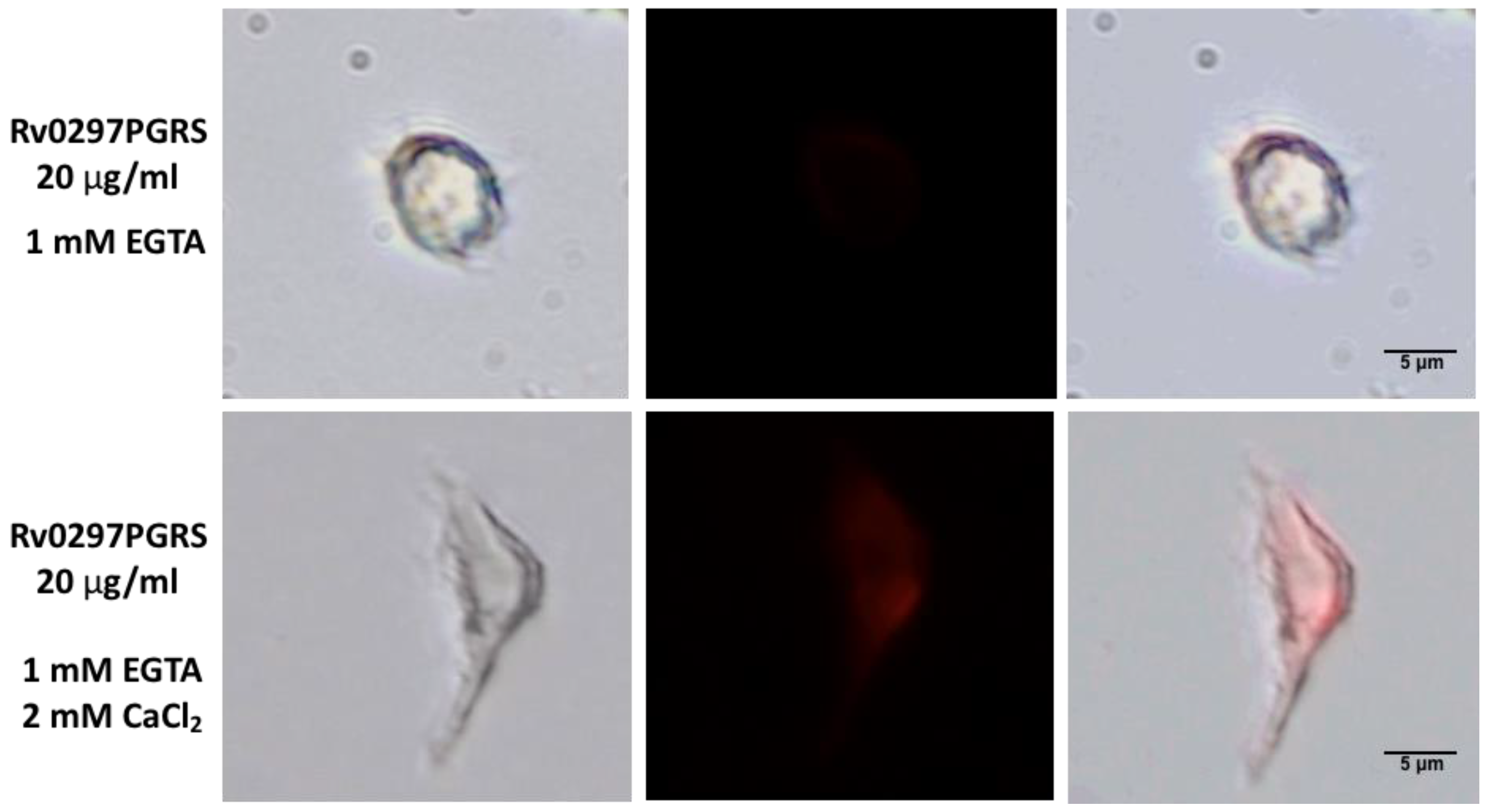
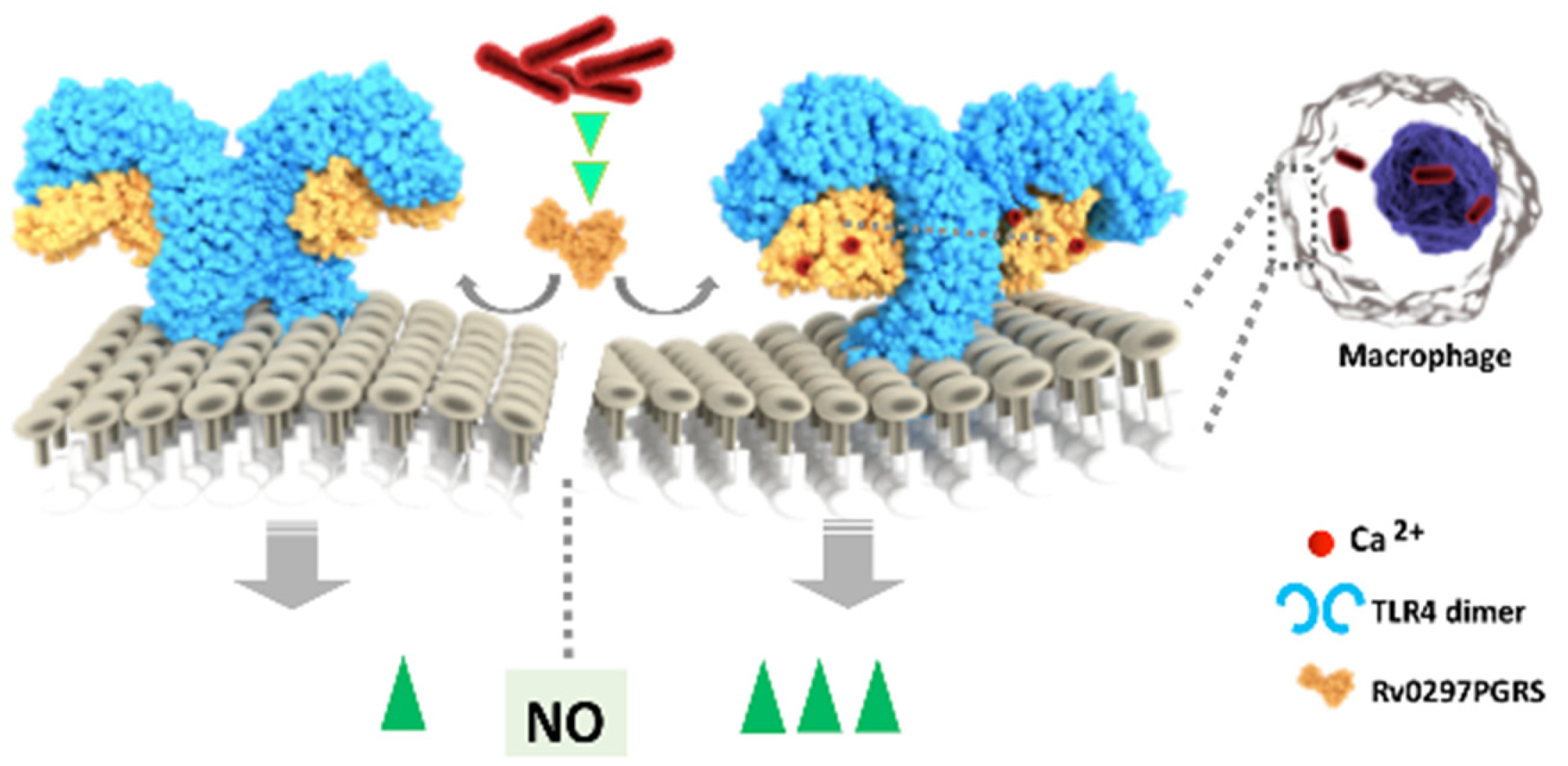
2.5. Interaction of Rv0297PGRS with Macrophage TLR4 Is Stabilized in the Presence of Ca2+ Ions
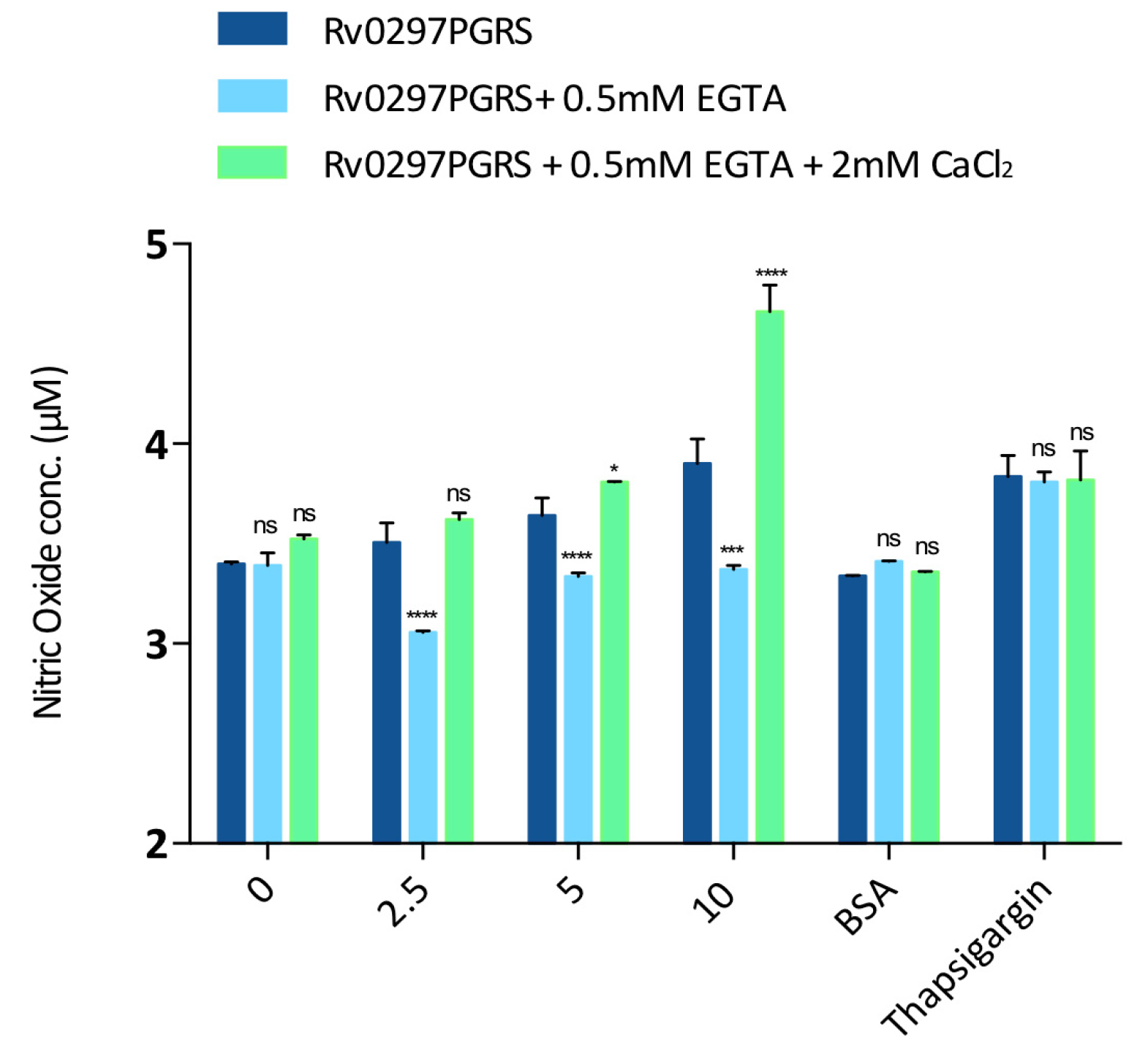
2.6. Downstream Effect of Rv0297PGRS Is Enhanced in the Presence of Calcium
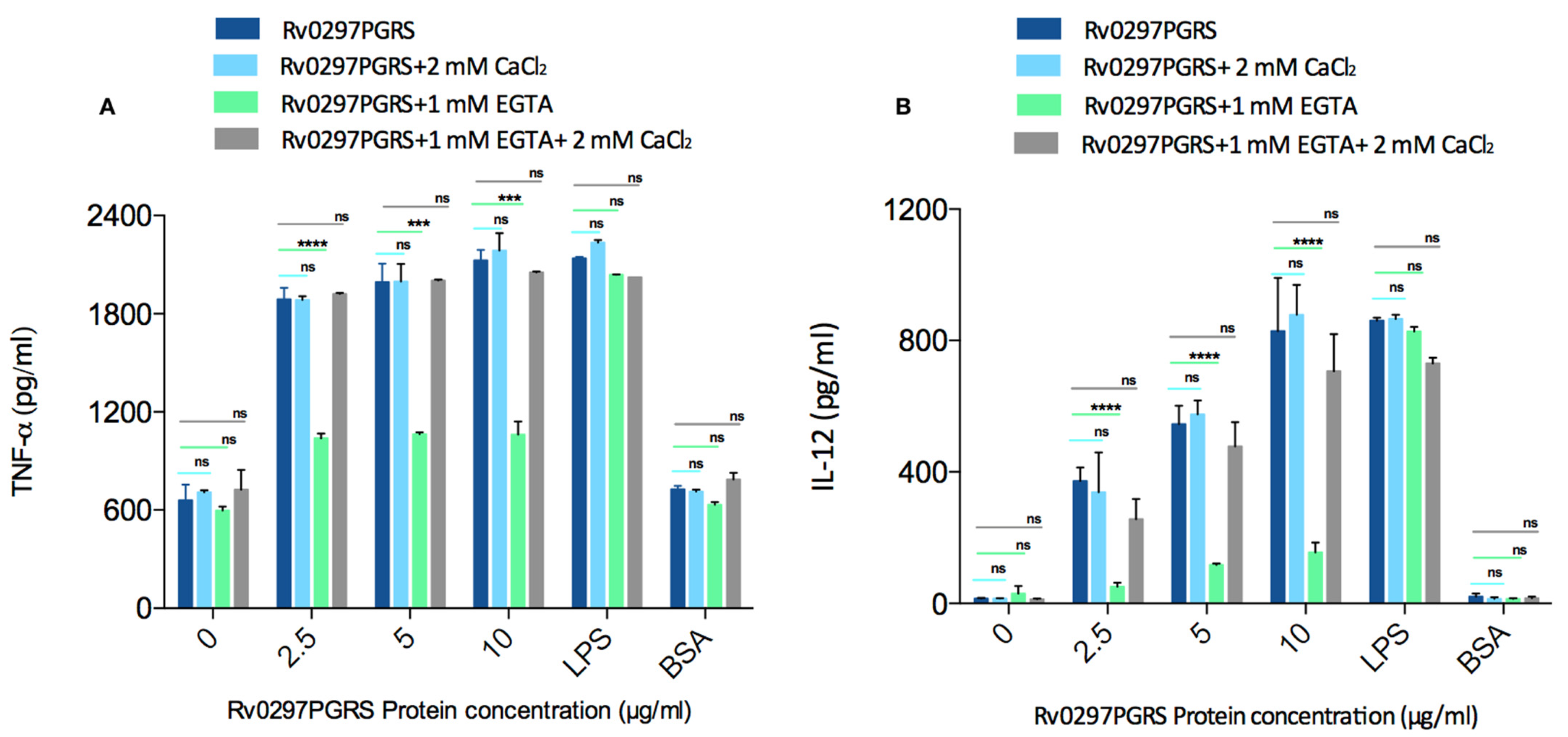
2.7. Rv0297PGRS Induces the Production of Pro-Inflammatory Cytokines in Calcium-Dependent Manner
3. Discussion
4. Material and Methods
4.1. Prediction of Calcium-Binding Motifs in M.tb PE_PGRS Proteins
4.2. Prediction of Ca2+ Binding Sites and Structural Modeling of Rv0297PGRS-TLR4 Complex
4.3. Molecular Dynamics Systems and Simulations
4.4. Binding of Ca2+ with Rv0297PGRS
4.5. Assessment of Rv0297PGRS Stabilization/Destabilization by Ca2+
4.6. Secondary Structure Estimation of Rv0297PGRS
4.7. Cell Culture
4.8. Immunofluorescence Staining
4.9. Nitrite Quantitation in Macrophages
4.10. Cytokine Assessment in Macrophages
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Tuberculosis Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Chakaya, J.M.; Marais, B.; du Cros, P.; Ntoumi, F.; Mfinanga, S.; Kapata, N.; Hasnain, S.E.; Nathavitharana, R.; Zumla, A. Programmatic versus personalised approaches to managing the global epidemic of multidrug-resistant tuberculosis. Lancet Respir. Med. 2020, 8, 334–335. [Google Scholar] [CrossRef]
- WHO. Global Tuberculosis Report 2019 Geneva; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Fenton, M.J.; Vermeulen, M.W. Immunopathology of tuberculosis: Roles of macrophages and monocytes. Infect. Immun. 1996, 64, 683–690. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C., 3rd; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 396, 190–198. [Google Scholar] [CrossRef]
- Sampson, S.L. Mycobacterial PE/PPE proteins at the host-pathogen interface. Clin. Dev. Immunol. 2011, 2011, 497203. [Google Scholar] [CrossRef] [Green Version]
- Akhter, Y.; Ehebauer, M.T.; Mukhopadhyay, S.; Hasnain, S.E. The PE/PPE multigene family codes for virulence factors and is a possible source of mycobacterial antigenic variation: Perhaps more? Biochimie 2012, 94, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Meena, L.S. An overview to understand the role of PE_PGRS family proteins in Mycobacterium tuberculosis H37Rv and their potential as new drug targets. Biotechnol. Appl. Biochem. 2015, 62, 145–153. [Google Scholar] [CrossRef]
- Ahmad, J.; Farhana, A.; Pancsa, R.; Arora, S.K.; Srinivasan, A.; Tyagi, A.K.; Babu, M.M.; Ehtesham, N.Z.; Hasnain, S.E. Contrasting function of structured N-terminal and unstructured C-terminal segments of Mycobacterium tuberculosis PPE37 protein. MBio 2018, 9, e01712-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehtesham, N.Z.; Ahmad, J.; Farhan, A.; Khubaib, M.; Kaur, S.; Pancsa, R.; Srinivasan, A.; Kumar, S.; Babu, M.; Hasnain, S.E. Intrinsically Disordered Regions/Proteins Compensate For Genomic Economization In Mycobacterium Tuberculosis. FASEB J. 2018, 31, 526.23. [Google Scholar]
- Ahmad, J.; Khubaib, M.; Sheikh, J.A.; Pancsa, R.; Kumar, S.; Srinivasan, A.; Babu, M.M.; Hasnain, S.E.; Ehtesham, N.Z. Disorder-to-order transition in PE–PPE proteins of Mycobacterium tuberculosis augments the pro-pathogen immune response. FEBS Open Bio 2020, 10, 70–85. [Google Scholar] [CrossRef]
- Brennan, M.J.; Delogu, G. The PE multigene family: A ‘molecular mantra’ for mycobacteria. Trends Microbiol. 2002, 10, 246–249. [Google Scholar] [CrossRef]
- Cohen, I.; Parada, C.; Acosta-Gío, E.; Espitia, C. The PGRS Domain from PE_PGRS33 of Mycobacterium tuberculosis is Target of Humoral Immune Response in Mice and Humans. Front. Immunol. 2014, 5, 236. [Google Scholar] [CrossRef]
- Delogu, G.; Brennan, M.J. Comparative immune response to PE and PE_PGRS antigens of Mycobacterium tuberculosis. Infect. Immun. 2001, 69, 5606–5611. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, B.M.; Kannan, N.; Vemu, L.; Raghunand, T.R. The Mycobacterium tuberculosis PE proteins Rv0285 and Rv1386 modulate innate immunity and mediate bacillary survival in macrophages. PLoS ONE 2012, 7, e51686. [Google Scholar]
- Tiwari, B.; Soory, A.; Raghunand, T.R. An immunomodulatory role for the Mycobacterium tuberculosis region of difference 1 locus proteins PE35 (Rv3872) and PPE68 (Rv3873). FEBS J. 2014, 281, 1556–1570. [Google Scholar] [CrossRef]
- Singh, P.P.; Parra, M.; Cadieux, N.; Brennan, M.J. A comparative study of host response to three Mycobacterium tuberculosis PE_PGRS proteins. Microbiology 2008, 154 Pt 11, 3469–3479. [Google Scholar] [CrossRef] [Green Version]
- Bansal, K.; Elluru, S.R.; Narayana, Y.; Chaturvedi, R.; Patil, S.A.; Kaveri, S.V.; Bayry, J.; Balaji, K.N. PE_PGRS antigens of Mycobacterium tuberculosis induce maturation and activation of human dendritic cells. J. Immunol. 2010, 184, 3495–3504. [Google Scholar] [PubMed] [Green Version]
- Khubaib, M.; Sheikh, J.A.; Pandey, S.; Srikanth, B.; Bhuwan, M.; Khan, N.; Hasnain, S.E.; Ehtesham, N.Z. Mycobacterium tuberculosis Co-operonic PE32/PPE65 Proteins Alter Host Immune Responses by Hampering Th1 Response. Front. Microbiol. 2016, 7, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Pathak, S.K.; Banerjee, A.; Pathak, S.; Bhattacharyya, A.; Yang, Z.; Talarico, S.; Kundu, M.; Basu, J. Execution of macrophage apoptosis by PE_PGRS33 of Mycobacterium tuberculosis is mediated by Toll-like receptor 2-dependent release of tumor necrosis factor-alpha. J. Biol. Chem. 2007, 282, 1039–1050. [Google Scholar] [CrossRef] [Green Version]
- Grover, S.; Sharma, T.; Singh, Y.; Kohli, S.; Manjunath, P.; Singh, A.; Semmler, T.; Wieler, L.H.; Tedin, K.; Ehtesham, N.Z.; et al. The PGRS domain of Mycobacterium tuberculosis PE_PGRS protein Rv0297 Is involved in endoplasmic eeticulum stress-mediated apoptosis through Toll-Like receptor 4. MBio 2018, 9, e01017-18. [Google Scholar] [CrossRef] [Green Version]
- Iantomasi, R.; Sali, M.; Cascioferro, A.; Palucci, I.; Zumbo, A.; Soldini, S.; Rocca, S.; Greco, E.; Maulucci, G.; De Spirito, M.; et al. PE_PGRS30 is required for the full virulence of Mycobacterium tuberculosis. Cell. Microbiol. 2012, 14, 356–367. [Google Scholar] [CrossRef]
- Ramakrishnan, L.; Federspiel, N.A.; Falkow, S. Granuloma-specific expression of Mycobacterium virulence proteins from the glycine-rich PE-PGRS family. Science 2000, 288, 1436–1439. [Google Scholar] [CrossRef] [Green Version]
- Thi, E.P.; Hong, C.J.; Sanghera, G.; Reiner, N.E. Identification of the Mycobacterium tuberculosis protein PE-PGRS62 as a novel effector that functions to block phagosome maturation and inhibit iNOS expression. Cell Microbiol. 2013, 15, 795–808. [Google Scholar] [CrossRef]
- Kohli, S.; Singh, Y.; Sharma, K.; Mittal, A.; Ehtesham, N.Z.; Hasnain, S.E. Comparative genomic and proteomic analyses of PE/PPE multigene family of Mycobacterium tuberculosis H(3)(7)Rv and H(3)(7)Ra reveal novel and interesting differences with implications in virulence. Nucleic Acids Res. 2012, 40, 7113–7122. [Google Scholar] [CrossRef]
- Palucci, I.; Camassa, S.; Cascioferro, A.; Sali, M.; Anoosheh, S.; Zumbo, A.; Minerva, M.; Iantomasi, R.; De Maio, F.; Di Sante, G.; et al. PE_PGRS33 contributes to Mycobacterium tuberculosis entry in macrophages through interaction with TLR2. PLoS ONE 2016, 11, e0150800. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.J.; Delogu, G.; Chen, Y.; Bardarov, S.; Kriakov, J.; Alavi, M.; Jacobs, W.R. Evidence that mycobacterial PE_PGRS proteins are cell surface constituents that influence interactions with other cells. Infect. Immun. 2001, 69, 7326–7333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delogu, G.; Pusceddu, C.; Bua, A.; Fadda, G.; Brennan, M.J.; Zanetti, S. Rv1818c-encoded PE_PGRS protein of Mycobacterium tuberculosis is surface exposed and influences bacterial cell structure. Mol. Microbiol. 2004, 52, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Dheenadhayalan, V.; Delogu, G.; Brennan, M.J. Expression of the PE_PGRS 33 protein in Mycobacterium smegmatis triggers necrosis in macrophages and enhanced mycobacterial survival. Microbes Infect. 2006, 8, 262–272. [Google Scholar] [CrossRef]
- Balaji, K.N.; Goyal, G.; Narayana, Y.; Srinivas, M.; Chaturvedi, R.; Mohammad, S. Apoptosis triggered by Rv1818c, a PE family gene from Mycobacterium tuberculosis is regulated by mitochondrial intermediates in T cells. Microbes Infect. 2007, 9, 271–281. [Google Scholar] [CrossRef]
- Cadieux, N.; Parra, M.; Cohen, H.; Maric, D.; Morris, S.L.; Brennan, M.J. Induction of cell death after localization to the host cell mitochondria by the Mycobacterium tuberculosis PE_PGRS33 protein. Microbiology 2011, 157, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Chatrath, S.; Gupta, V.K.; Dixit, A.; Garg, L.C. The Rv1651c-encoded PE-PGRS30 protein expressed in Mycobacterium smegmatis exhibits polar localization and modulates its growth profile. FEMS Microbiol. Lett. 2011, 322, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Chatrath, S.; Gupta, V.K.; Dixit, A.; Garg, L.C. PE_PGRS30 of Mycobacterium tuberculosis mediates suppression of proinflammatory immune response in macrophages through its PGRS and PE domains. Microbes Infect. 2016, 18, 536–542. [Google Scholar] [CrossRef]
- Bhuwan, M.; Arora, N.; Sharma, A.; Khubaib, M.; Pandey, S.; Chaudhuri, T.K.; Hasnain, S.E.; Ehtesham, N.Z. Interaction of Mycobacterium tuberculosis Virulence Factor RipA with Chaperone MoxR1 Is Required for Transport through the TAT Secretion System. MBio 2016, 7, e02259. [Google Scholar] [CrossRef] [Green Version]
- Gupta, M.N.; Alam, A.; Hasnain, S.E. Protein promiscuity in drug discovery, drug-repurposing and antibiotic resistance. Biochimie 2020, 175, 50–57. [Google Scholar] [CrossRef]
- Fayyazi, A.; Eichmeyer, B.; Soruri, A.; Schweyer, S.; Herms, J.; Schwarz, P.; Radzun, H.J. Apoptosis of macrophages and T cells in tuberculosis associated caseous necrosis. J. Pathol. 2000, 191, 417–425. [Google Scholar] [CrossRef]
- Sánchez, D.; Rojas, M.; Hernández, I.; Radzioch, D.; García, L.F.; Barrera, L.F. Role of TLR2- and TLR4-mediated signaling in Mycobacterium tuberculosis-induced macrophage death. Cell. Immunol. 2010, 260, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Shariq, M.; Quadir, N.; Sheikh, J.A.; Singh, A.K.; Bishai, W.R.; Ehtesham, N.Z.; Hasnain, S.E. Post translational modifications in tuberculosis: Ubiquitination paradox. Autophagy 2020, 1–4. [Google Scholar] [CrossRef]
- Wojtas, M.; Hołubowicz, R.; Poznar, M.; Maciejewska, M.; Ożyhar, A.; Dobryszycki, P. Calcium ion binding properties and the effect of phosphorylation on the intrinsically disordered starmaker protein. Biochemistry 2015, 54, 6525–6534. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chenal, A.; Karst, J.C.; Pérez, A.C.; Wozniak, A.K.; Baron, B.; England, P.; Ladant, D. Calcium-induced folding and stabilization of the intrinsically disordered RTX domain of the CyaA toxin. Biophys. J. 2010, 99, 3744–3753. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Conway, J.F.; Thibodeau, P.H. Calcium-induced folding and stabilization of the Pseudomonas aeruginosa alkaline protease. J. Biol. Chem. 2012, 287, 4311–4322. [Google Scholar] [CrossRef] [Green Version]
- Malik, Z.A.; Thompson, C.R.; Hashimi, S.; Porter, B.; Iyer, S.S.; Kusner, D.J. Cutting edge: Mycobacterium tuberculosis blocks Ca2+ signaling and phagosome maturation in human macrophages via specific inhibition of sphingosine kinase. J. Immunol. 2003, 170, 2811–2815. [Google Scholar] [CrossRef] [Green Version]
- Meena, L.S. Interrelation of Ca2+ and PE_PGRS proteins during Mycobacterium tuberculosis pathogenesis. J. Biosci. 2019, 44, 24. [Google Scholar] [CrossRef]
- Sharma, T.; Grover, S.; Arora, N.; Ehtesham, N.Z.; Hasnain, S.E. PGRS Domain of Rv0297 of Mycobacterium tuberculosis Is Involved in Modulation of Macrophage Functions to Favor Bacterial Persistence. Front. Cell. Infect. Microbiol. 2020, 10, 451. [Google Scholar] [CrossRef]
- Bachhawat, N.; Singh, B. Mycobacterial PE_PGRS proteins contain calcium-binding motifs with parallel beta-roll folds. Genom. Proteom. Bioinform. 2007, 5, 236–241. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Rao, H.; Jin, C.E.; Liu, J. Involvement of the Toll-like receptor/nitric oxide signaling pathway in the pathogenesis of cervical cancer caused by high-risk human papillomavirus infection. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Lipsitch, M.; Moxon, E.R. Virulence and transmissibility of pathogens: What is the relationship? Trends Microbiol. 1997, 5, 31–37. [Google Scholar] [CrossRef]
- Banu, S.; Honoré, N.; Saint-Joanis, B.; Philpott, D.; Prévost, M.C.; Cole, S.T. Are the PE-PGRS proteins of Mycobacterium tuberculosis variable surface antigens? Mol. Microbiol. 2002, 44, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Talarico, S.; Cave, M.D.; Marrs, C.F.; Foxman, B.; Zhang, L.; Yang, Z. Variation of the Mycobacterium tuberculosis PE_PGRS33 Gene among Clinical Isolates. J. Clin. Microbiol. 2005, 43, 4954. [Google Scholar] [PubMed] [Green Version]
- Abdallah, A.M.; Verboom, T.; Weerdenburg, E.M.; Gey van Pittius, N.C.; Mahasha, P.W.; Jiménez, C.; Parra, M.; Cadieux, N.; Brennan, M.J.; Appelmelk, B.J. PPE and PE_PGRS proteins of Mycobacterium marinum are transported via the type VII secretion system ESX-5. Mol. Microbiol. 2009, 73, 329–340. [Google Scholar] [CrossRef]
- Ates, L.S.; Dippenaar, A.; Ummels, R.; Piersma, S.R.; van der Woude, A.D.; van der Kuij, K.; Le Chevalier, F.; Mata-Espinosa, D.; Barrios-Payán, J.; Marquina-Castillo, B.; et al. Mutations in ppe38 block PE-PGRS secretion and increase virulence of Mycobacterium tuberculosis. Nat. Microbiol. 2018, 3, 181–188. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, Y.; Bai, Y.; Wang, Z.G.; Yang, L.; Zhao, D. Expression of PE_PGRS 62 protein in Mycobacterium smegmatis decrease mRNA expression of proinflammatory cytokines IL-1beta, IL-6 in macrophages. Mol. Cell. Biochem. 2010, 340, 223–229. [Google Scholar] [CrossRef]
- Yang, W.; Deng, W.; Zeng, J.; Ren, S.; Ali, M.K.; Gu, Y.; Li, Y.; Xie, J. Mycobacterium tuberculosis PE_PGRS18 enhances the intracellular survival of M. smegmatis via altering host macrophage cytokine profiling and attenuating the cell apoptosis. Apoptosis 2017, 22, 502–509. [Google Scholar] [CrossRef]
- Deng, W.; Long, Q.; Zeng, J.; Li, P.; Yang, W.; Chen, X.; Xie, J. Mycobacterium tuberculosis PE_PGRS41 Enhances the Intracellular Survival of M. smegmatis within Macrophages Via Blocking Innate Immunity and Inhibition of Host Defense. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Xie, Y.; Luo, W.; Dou, Y.; Xiong, H.; Xiao, Z.; Zhang, X.L. PE_PGRS31-S100A9 Interaction Promotes Mycobacterial Survival in Macrophages Through the Regulation of NF-κB-TNF-α Signaling and Arachidonic Acid Metabolism. Front. Microbiol. 2020, 11, 845. [Google Scholar] [CrossRef]
- Baumann, U.; Wu, S.; Flaherty, K.M.; McKay, D.B. Three-dimensional structure of the alkaline protease of Pseudomonas aeruginosa: A two-domain protein with a calcium binding parallel beta roll motif. EMBO J. 1993, 12, 3357–3364. [Google Scholar] [CrossRef] [PubMed]
- Tompa, P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef]
- Via, A.; Uyar, B.; Brun, C.; Zanzoni, A. How pathogens use linear motifs to perturb host cell networks. Trends Biochem. Sci. 2015, 40, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.N.; Pandey, S.; Ehtesham, N.Z.; Hasnain, S.E. Medical implications of protein moonlighting. Indian J. Med. Res. 2019, 149, 322–325. [Google Scholar] [PubMed]
- Blundell, T.L.; Gupta, M.N.; Hasnain, S.E. Intrinsic disorder in proteins: Relevance to protein assemblies, drug design and host-pathogen interactions. Prog. Biophys. Mol. Biol. 2020, 156, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Yeruva, V.C.; Kulkarni, A.; Khandelwal, R.; Sharma, Y.; Raghunand, T.R. The PE_PGRS proteins of Mycobacterium tuberculosis are Ca2+ binding mediators of host-pathogen interaction. Biochemistry 2016, 55, 4675–4687. [Google Scholar] [CrossRef] [PubMed]
- Trimble, W.S.; Grinstein, S. TB or not TB: Calcium regulation in mycobacterial survival. Cell 2007, 130, 12–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, A.N.; Powis, S.J. Pathogen evasion strategies for the major histocompatibility complex class I assembly pathway. Immunology 2008, 124, 1–12. [Google Scholar] [CrossRef]
- Niu, W.; Sun, B.; Li, M.; Cui, J.; Huang, J.; Zhang, L. TLR-4/microRNA-125a/NF-κB signaling modulates the immune response to Mycobacterium tuberculosis infection. Cell Cycle 2018, 17, 1931–1945. [Google Scholar] [CrossRef] [Green Version]
- Stamm, C.E.; Collins, A.C.; Shiloh, M.U. Sensing of Mycobacterium tuberculosis and consequences to both host and bacillus. Immunol. Rev. 2015, 264, 204–219. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.V.; Boom, W.H. Regulation of antigen presentation by Mycobacterium tuberculosis: A role for Toll-like receptors. Nat. Rev. Microbiol. 2010, 8, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, S.; Dkhar, H.K.; Chandra, V.; Dave, S.; Nanduri, R.; Janmeja, A.K.; Agrewala, J.N.; Gupta, P. Mycobacterium tuberculosis modulates macrophage lipid-sensing nuclear receptors PPARγ and TR4 for survival. J. Immunol. 2012, 188, 5593–5603. [Google Scholar] [CrossRef] [Green Version]
- Kleinnijenhuis, J.; Oosting, M.; Joosten, L.A.; Netea, M.G.; Van Crevel, R. Innate Immune Recognition of Mycobacterium tuberculosis. Clin. Dev. Immunol. 2011, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortaz, E.; Adcock, I.M.; Tabarsi, P.; Masjedi, M.R.; Mansouri, D.; Velayati, A.A.; Casanova, J.L.; Barnes, P.J. Interaction of pattern recognition receptors with Mycobacterium tuberculosis. J. Clin. Immunol. 2015, 35, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gilleron, M.; Quesniaux, V.F.; Puzo, G. Acylation State of the Phosphatidylinositol Hexamannosides from Mycobacterium bovis Bacillus Calmette Guérin and Mycobacterium tuberculosis H37Rv and Its Implication in Toll-like Receptor Response. J. Biol. Chem. 2003, 278, 29880–29889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latty, S.L.; Sakai, J.; Hopkins, L.; Verstak, B.; Paramo, T.; Berglund, N.A.; Cammarota, E.; Cicuta, P.; Gay, N.J.; Bond, P.J.; et al. Activation of Toll-like receptors nucleates assembly of the MyDDosome signaling hub. Elife 2018, 7, e31377. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [Green Version]
- Schrodinger, LLC. The PyMOL Molecular Graphics System; Version 1.8.; Schrodinger, LLC: New York, NY, USA, 2015. [Google Scholar]
- Hu, X.; Dong, Q.; Yang, J.; Zhang, Y. Recognizing metal and acid radical ion-binding sites by integrating ab initio modeling with template-based transferals. Bioinformatics 2016, 32, 3260–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2017, 35 (Suppl. S2), W407–W410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein-protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef]
- van Gunsteren, W.F.; Billeter, S.R.; Eising, A.A.; Hünenberger, P.H.; Krüger, P.K.; Mark, A.E.; Scott, W.R.; Tironi, I.G. Biomolecular simulation: The {GROMOS96} manual and user guide. Vdf Hochsch. AG ETH Zür. Zür. 1996, 86, 1–1044. [Google Scholar]
- van Gunsteren, W.F.; Daura, X.; Mark, A.E. GROMOS force field. Encycl. Comput. Chem. 2002, 2. [Google Scholar]
- Micsonai, A.; Bulyáki, É.; Kardos, J. BeStSel: From Secondary Structure Analysis to Protein Fold Prediction by Circular Dichroism Spectroscopy. Methods Mol. Biol. 2021, 2199, 175–189. [Google Scholar]
- Micsonai, A.; Wien, F.; Bulyáki, É.; Kun, J.; Moussong, É.; Lee, Y.H.; Goto, Y.; Réfrégiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucleic Acids Res. 2018, 46, W315–W322. [Google Scholar] [CrossRef]
- Grover, S.; Arya, R. Role of UDP-N-acetylglucosamine2-epimerase/N-acetylmannosamine kinase (GNE) in beta1-integrin-mediated cell adhesion. Mol. Neurobiol. 2014, 50, 257–273. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, T.; Singh, J.; Grover, S.; P., M.; Firdos, F.; Alam, A.; Ehtesham, N.Z.; Hasnain, S.E. PGRS Domain of Rv0297 of Mycobacterium tuberculosis Functions in A Calcium Dependent Manner. Int. J. Mol. Sci. 2021, 22, 9390. https://doi.org/10.3390/ijms22179390
Sharma T, Singh J, Grover S, P. M, Firdos F, Alam A, Ehtesham NZ, Hasnain SE. PGRS Domain of Rv0297 of Mycobacterium tuberculosis Functions in A Calcium Dependent Manner. International Journal of Molecular Sciences. 2021; 22(17):9390. https://doi.org/10.3390/ijms22179390
Chicago/Turabian StyleSharma, Tarina, Jasdeep Singh, Sonam Grover, Manjunath P., Firdos Firdos, Anwar Alam, Nasreen Z. Ehtesham, and Seyed E. Hasnain. 2021. "PGRS Domain of Rv0297 of Mycobacterium tuberculosis Functions in A Calcium Dependent Manner" International Journal of Molecular Sciences 22, no. 17: 9390. https://doi.org/10.3390/ijms22179390