
HIV Associated Preeclampsia: A Multifactorial Appraisal


Abstract
:
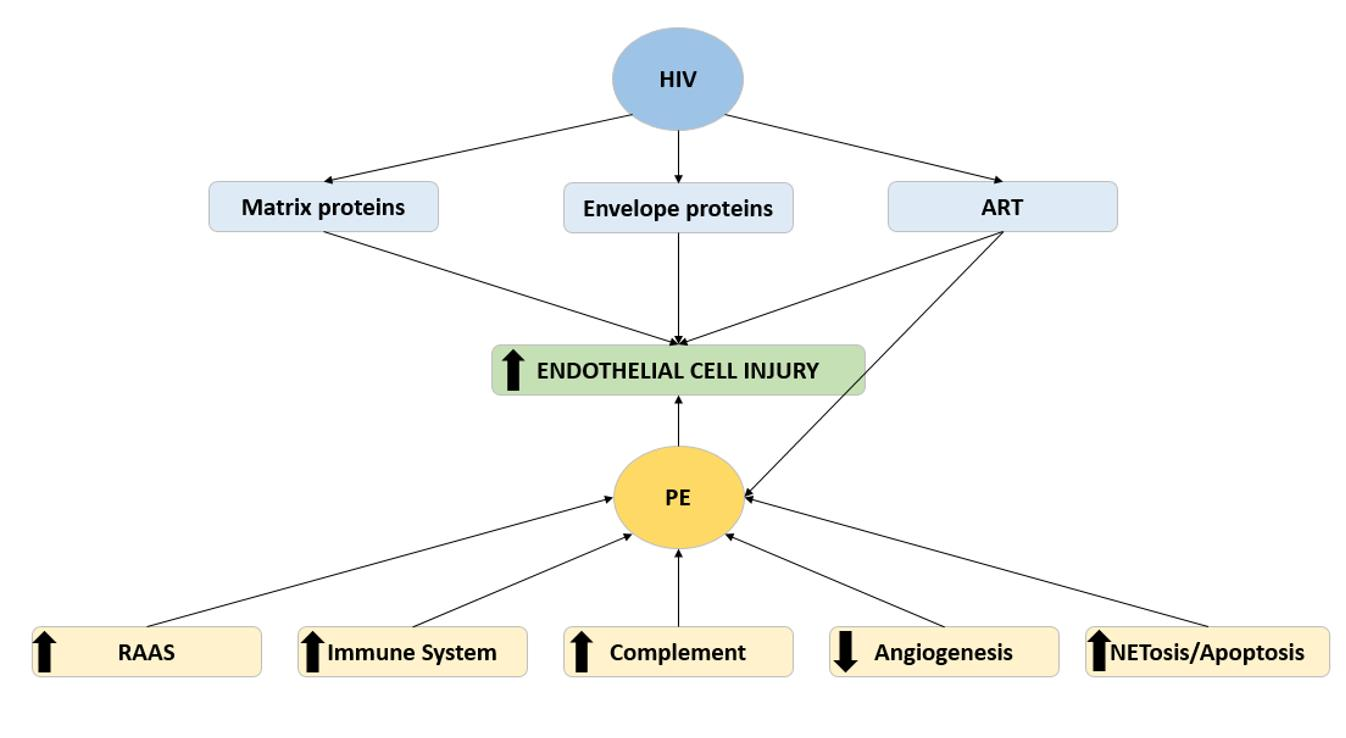
1. Introduction
2. Immune Response and the Complement System
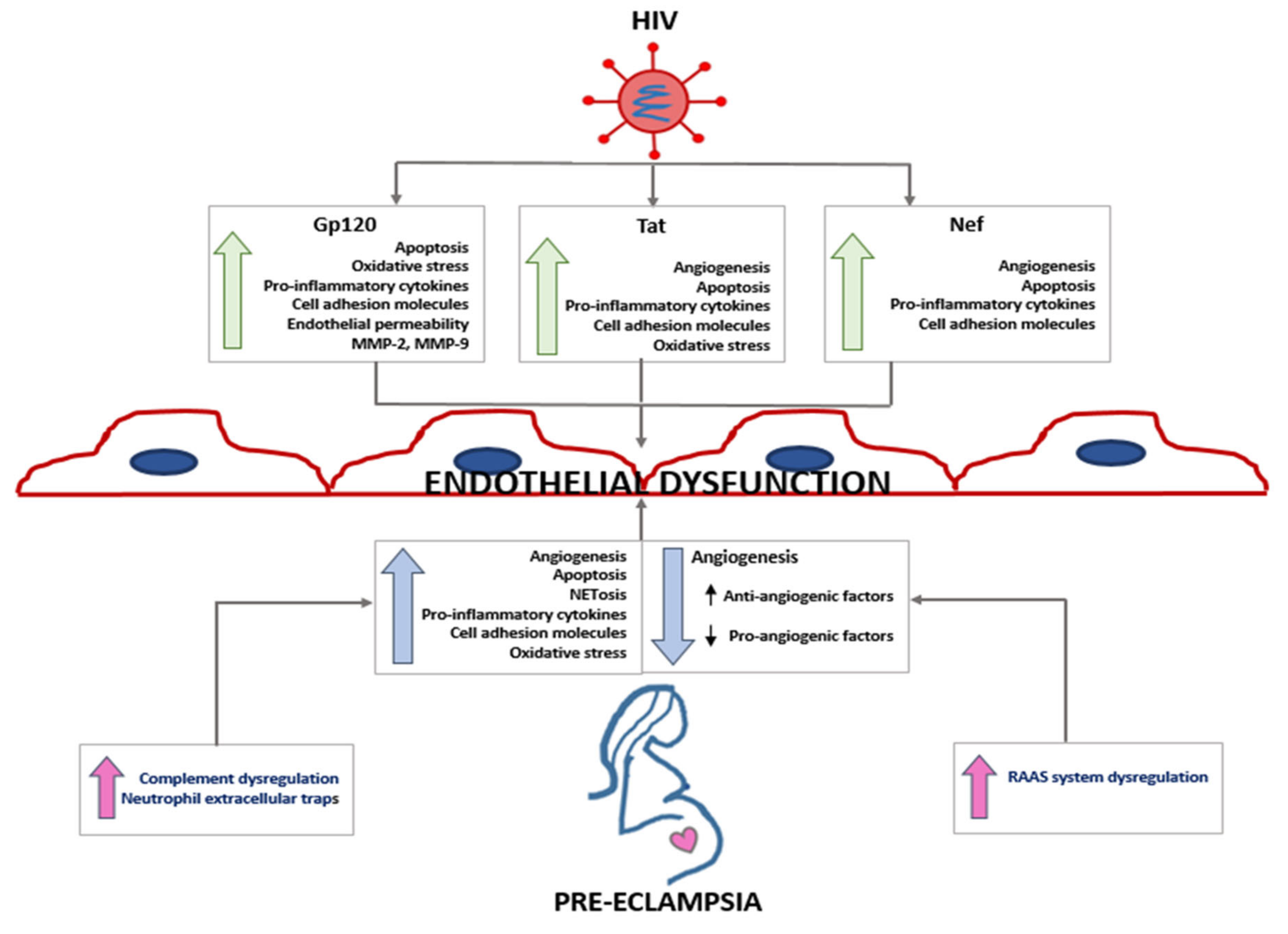
3. HIV Infection and Endothelial Dysfunction
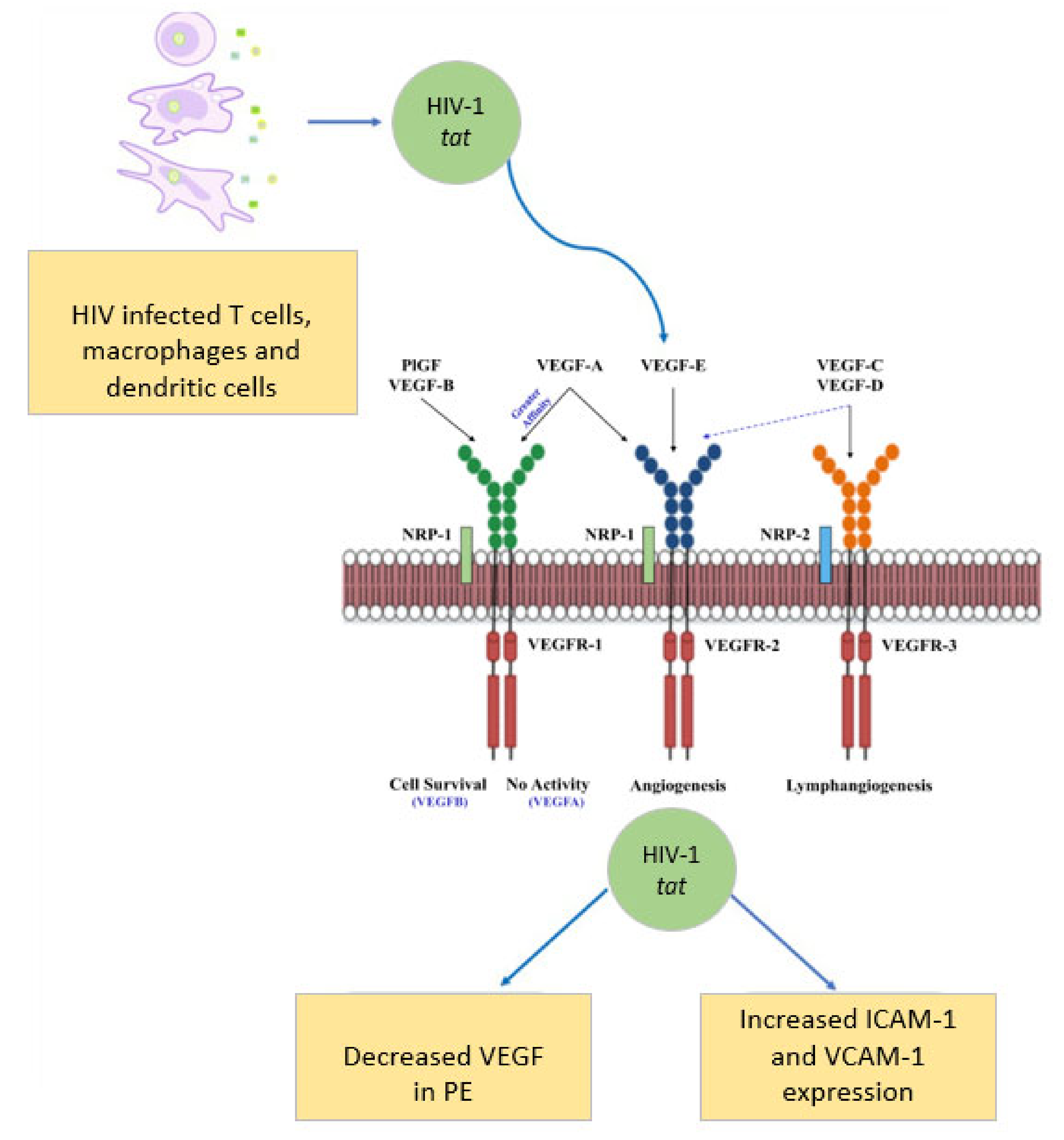
4. Vascular Endothelial Growth Factors: Pivotal Angiogenesis Regulators
5. Angiogenic Imbalance in Preeclampsia

{kind=link}
{kind=link}
{kind=link}
| Angiogenic/ Antiangiogenic/ Lymphangiogenic Factors | Main Findings | References |
|---|---|---|
| VEGFR-2 |
| [70,91,92,93] |
| VEGFR-3 |
| [94,95] |
| VEGF-A |
| [71,96,97,98] |
| VEGF-C |
| [99,100] |
| PlGF |
| [70,71,96,98] |
| sFlt-1 |
| [71,96,101,102,103,104] |
| sEng |
| [69,70,73,80,105,106,107,108,109] |
6. Angiogenesis and Lymphangiogenesis
7. Antiretroviral Therapy and Preeclampsia
8. Renin-Angiotensin-Aldosterone System in HIV Associated Preeclamptic Women on ART
9. Cell Death-Neutrophil Extracellular Traps (NETs): Immune Defence
10. NETs in HIV-Associated Preeclampsia
11. Recommendations and Therapy Guidelines: HIV Infection and Preeclampsia
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization Maternal Mortality Ratio. Available online: https://www.who.int/news-room/fact-sheets/detail/maternal-mortality (accessed on 16 August 2021).
- Say, L.; Chou, D.; Gemmill, A.; Tunçalp, Ö.; Moller, A.B.; Daniels, J.; Gülmezoglu, A.M.; Temmerman, M.; Alkema, L. Global causes of maternal death: A WHO systematic analysis. Lancet Glob. Health 2014, 2, e323–e333. [Google Scholar] [CrossRef] [Green Version]
- National Committee for Confidential Enquiry into Maternal Deaths. Saving Mothers Report 2017; Department of Health: Pretoria, South Africa, 2017. [Google Scholar]
- UNAIDS Fact Sheet—Global AIDS Update. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 21 December 2019).
- Calvert, C.; Marston, M.; Slaymaker, E.; Crampin, A.C.; Price, A.J.; Klein, N.; Herbst, K.; Michael, D.; Urassa, M.; Clark, S.J.; et al. Direct maternal deaths attributable to HIV in the era of antiretroviral therapy: Evidence from three population-based HIV cohorts with verbal autopsy. Aids 2020, 34, 1397–1405. [Google Scholar] [CrossRef]
- Zaba, B.; Calvert, C.; Marston, M.; Isingo, R.; Nakiyingi-Miiro, J.; Lutalo, T.; Crampin, A.; Robertson, L.; Herbst, K.; Newell, M.L.; et al. Effect of HIV infection on pregnancy-related mortality in sub-Saharan Africa: Secondary analyses of pooled community-based data from the network for Analysing Longitudinal Population-based HIV/AIDS data on Africa (ALPHA). Lancet 2013, 381, 1763–1771. [Google Scholar] [CrossRef] [Green Version]
- Clouse, K.; Malope-Kgokong, B.; Bor, J.; Nattey, C.; Mudau, M.; Maskew, M. The South African National HIV Pregnancy Cohort: Evaluating continuity of care among women living with HIV. BMC Public Health 2020, 20, 1662. [Google Scholar] [CrossRef] [PubMed]
- Kalumba, V.M.; Moodley, J.; Naidoo, T.D. Is the prevalence of pre-eclampsia affected by HIV/AIDS? A retrospective case-control study. Cardiovasc. J. Afr. 2013, 24, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.G.; Rabkin, M.; El-Sadr, W.M. Achieving the fourth 90: Healthy aging for people living with HIV. Aids 2018, 32, 1563–1569. [Google Scholar] [CrossRef]
- The Antiretroviral Therapy Cohort Collaboration. Survival of HIV-positive patients starting antiretroviral therapy between 1996 and 2013: A collaborative analysis of cohort studies. Lancet HIV 2017, 4, e349–e356. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.; Yudin, M.H. HIV Infection in Pregnant Women: A 2020 Update. Prenat. Diagn. 2020, 40, 1715–1721. [Google Scholar] [CrossRef]
- Fauci, A.S.; Lane, H.C.; Redfield, R.R. Covid-19—Navigating the Uncharted. N. Engl. J. Med. 2020, 382, 1268–1269. [Google Scholar] [CrossRef]
- Villar, J.; Ariff, S.; Gunier, R.B.; Thiruvengadam, R.; Rauch, S.; Kholin, A.; Roggero, P.; Prefumo, F.; do Vale, M.S.; Cardona-Perez, J.A.; et al. Maternal and Neonatal Morbidity and Mortality among Pregnant Women with and without COVID-19 Infection: The INTERCOVID Multinational Cohort Study. JAMA Pediatr. 2021, 175, 817–826. [Google Scholar] [CrossRef]
- Wei, S.Q.; Bilodeau-Bertrand, M.; Liu, S.; Auger, N. The impact of COVID-19 on pregnancy outcomes: A systematic review and meta-analysis. Cmaj 2021, 193, E540–E548. [Google Scholar] [CrossRef] [PubMed]
- Govender, R.; Moodley, J.; Naicker, T. The COVID-19 Pandemic: An Appraisal of its Impact on Human Immunodeficiency Virus Infection and Pre-Eclampsia. Curr. Hypertens. Rep. 2021, 23, 9. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, N.; Moodley, J.; Naicker, T. Maternal endothelial dysfunction in HIV-associated preeclampsia comorbid with COVID-19: A review. Hypertens. Res. 2021, 44, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. Hypertensive Disorders of Pregnancy: ISSHP Classification, Diagnosis, and Management Recommendations for International Practice. Hypertension 2018, 72, 24–43. [Google Scholar] [CrossRef] [Green Version]
- Jena, M.K.; Sharma, N.R.; Petitt, M.; Maulik, D.; Nayak, N.R. Pathogenesis of Preeclampsia and Therapeutic Approaches Targeting the Placenta. Biomolecules 2020, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Arullampalam, P.; Yang, Z.; Ming, X.F. Hypoxia Enhances Endothelial Intercellular Adhesion Molecule 1 Protein Level through Upregulation of Arginase Type II and Mitochondrial Oxidative Stress. Front. Physiol. 2019, 10, 1003. [Google Scholar] [CrossRef] [Green Version]
- Burwick, R.M.; Feinberg, B.B. Complement activation and regulation in preeclampsia and hemolysis, elevated liver enzymes, and low platelet count syndrome. Am. J. Obstet. Gynecol. 2020. [Google Scholar] [CrossRef]
- Colucci, F. The immunological code of pregnancy. Science 2019, 365, 862–863. [Google Scholar] [CrossRef]
- Lokki, A.I.; Heikkinen-Eloranta, J.K.; Laivuori, H. The Immunogenetic Conundrum of Preeclampsia. Science 2018, 9, 2630. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Wang, H.; Wang, Z.; Huang, H.; Dong, M. Elevated serum levels of interleukin-15 and interleukin-16 in preeclampsia. J. Reprod. Immunol. 2007, 73, 166–171. [Google Scholar] [CrossRef]
- Maharaj, N.R.; Phulukdaree, A.; Nagiah, S.; Ramkaran, P.; Tiloke, C.; Chuturgoon, A.A. Pro-Inflammatory Cytokine Levels in HIV Infected and Uninfected Pregnant Women with and without Preeclampsia. PLoS ONE 2017, 12, e0170063. [Google Scholar] [CrossRef] [Green Version]
- Noris, M.; Remuzzi, G. Overview of Complement Activation and Regulation. Semin. Nephrol. 2013, 33, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Girardi, G.; Lingo, J.J.; Fleming, S.D.; Regal, J.F. Essential Role of Complement in Pregnancy: From Implantation to Parturition and Beyond. Front. Immunol. 2020, 11, 1681. [Google Scholar] [CrossRef] [PubMed]
- Teirilä, L.; Heikkinen-Eloranta, J.; Kotimaa, J.; Meri, S.; Lokki, A.I. Regulation of the complement system and immunological tolerance in pregnancy. Semin. Immunol. 2019, 45, 101337. [Google Scholar] [CrossRef] [PubMed]
- Derzsy, Z.; Prohaszka, Z.; Rigo, J., Jr.; Fust, G.; Molvarec, A. Activation of the complement system in normal pregnancy and preeclampsia. Mol. Immunol. 2010, 47, 1500–1506. [Google Scholar] [CrossRef] [PubMed]
- Lokki, A.I.; Heikkinen-Eloranta, J.; Jarva, H.; Saisto, T.; Lokki, M.-L.; Laivuori, H.; Meri, S. Complement Activation and Regulation in Preeclamptic Placenta. Front. Immunol. 2014, 5, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, A.M.; Wagner, B.D.; Giclas, P.C.; West, N.A.; Gibbs, R.S.; Holers, V.M. The relationship of longitudinal levels of complement bb during pregnancy with preeclampsia. Am. J. Reprod. Immunol. 2016, 75, 104–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burwick, R.M.; Fichorova, R.N.; Dawood, H.Y.; Yamamoto, H.S.; Feinberg, B.B. Urinary excretion of C5b-9 in severe preeclampsia: Tipping the balance of complement activation in pregnancy. Hypertension 2013, 62, 1040–1045. [Google Scholar] [CrossRef] [Green Version]
- Mazzuca, P.; Caruso, A.; Caccuri, F. HIV-1 infection, microenvironment and endothelial cell dysfunction. New Microbiol. 2016, 39, 163–173. [Google Scholar]
- Pillay, Y.; Moodley, J.; Naicker, T. The role of the complement system in HIV infection and preeclampsia. Inflamm. Res. 2019, 68, 459–469. [Google Scholar] [CrossRef]
- Yu, Q.; Yu, R.; Qin, X. The good and evil of complement activation in HIV-1 infection. Cell. Mol. Immunol. 2010, 7, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Eisen, S.; Dzwonek, A.; Klein, N.J. Mannose-binding lectin in HIV infection. Future Virol. 2008, 3, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Kacani, L.; Bánki, Z.; Zwirner, J.; Schennach, H.; Bajtay, Z.; Erdei, A.; Stoiber, H.; Dierich, M.P. C5a and C5a(desArg) enhance the susceptibility of monocyte-derived macrophages to HIV infection. J. Immunol. 2001, 166, 3410–3415. [Google Scholar] [CrossRef] [Green Version]
- Bouhlal, H.; Chomont, N.; Haeffner-Cavaillon, N.; Kazatchkine, M.D.; Belec, L.; Hocini, H. Opsonization of HIV-1 by Semen Complement Enhances Infection of Human Epithelial Cells. J. Immunol. 2002, 169, 3301. [Google Scholar] [CrossRef] [Green Version]
- Harmon, A.C.; Cornelius, D.C.; Amaral, L.M.; Faulkner, J.L.; Cunningham, M.W., Jr.; Wallace, K.; LaMarca, B. The role of inflammation in the pathology of preeclampsia. Clin. Sci. 2016, 130, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Rossheim, A.E.B.; Cunningham, T.D.; Hair, P.S.; Shah, T.; Cunnion, K.M.; Troy, S.B. Effects of Well-Controlled HIV Infection on Complement Activation and Function. J. Acquir. Immune Defic. Syndr. 2016, 73, 20–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vujkovic-Cvijin, I.; Sortino, O.; Verheij, E.; Wit, F.W.; Kootstra, N.A.; Sellers, B.; Schim van der Loeff, M.; Belkaid, Y.; Reiss, P.; Sereti, I. The Complement Pathway Is Activated in People with Human Immunodeficiency Virus and Is Associated with Non-AIDS Comorbidities. J. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Khan, R.; Maduray, K.; Moodley, J.; Naicker, T. Activation of CD35 and CD55 in HIV associated normal and pre-eclamptic pregnant women. Eur J. Obs. Gynecol. Reprod. Biol. 2016, 204, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Perico, L.; Benigni, A.; Casiraghi, F.; Ng, L.F.P.; Renia, L.; Remuzzi, G. Immunity, endothelial injury and complement-induced coagulopathy in COVID-19. Nat. Rev. Nephrol. 2020, 17, 46–64. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Pillay, P.; Naicker, T.; Moodley, J.; Mackraj, I. Placental hypoxia inducible factor -1α & CHOP immuno-histochemical expression relative to maternal circulatory syncytiotrophoblast micro-vesicles in preeclamptic and normotensive pregnancies. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 220, 18–24. [Google Scholar]
- Masiá, M.; Padilla, S.; Fernández, M.; Rodríguez, C.; Moreno, A.; Oteo, J.A.; Antela, A.; Moreno, S.; Del Amo, J.; Gutiérrez, F. Oxidative Stress Predicts All-Cause Mortality in HIV-Infected Patients. PLoS ONE 2016, 11, e0153456. [Google Scholar] [CrossRef]
- Baliga, R.S.; Chaves, A.A.; Jing, L.; Ayers, L.W.; Bauer, J.A. AIDS-related vasculopathy: Evidence for oxidative and inflammatory pathways in murine and human AIDS. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H1373–H1380. [Google Scholar] [CrossRef]
- Mi, S.; Lee, X.; Li, X.P.; Veldman, G.M.; Finnerty, H.; Racie, L.; LaVallie, E.; Tang, X.Y.; Edouard, P.; Howes, S.; et al. Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis. Nature 2000, 403, 785–789. [Google Scholar] [CrossRef]
- Anand, A.R.; Rachel, G.; Parthasarathy, D. HIV Proteins and Endothelial Dysfunction: Implications in Cardiovascular Disease. Front. Cardiovasc. Med. 2018, 5, 185. [Google Scholar] [CrossRef] [PubMed]
- Blann, A.D.; Woywodt, A.; Bertolini, F.; Bull, T.M.; Buyon, J.P.; Clancy, R.M.; Haubitz, M.; Hebbel, R.P.; Lip, G.Y.; Mancuso, P.; et al. Circulating endothelial cells. Biomarker of vascular disease. Thromb. Haemost. 2005, 93, 228–235. [Google Scholar] [PubMed]
- Ganju, R.K.; Munshi, N.; Nair, B.C.; Liu, Z.Y.; Gill, P.; Groopman, J.E. Human immunodeficiency virus tat modulates the Flk-1/KDR receptor, mitogen-activated protein kinases, and components of focal adhesion in Kaposi’s sarcoma cells. J. Virol. 1998, 72, 6131–6137. [Google Scholar] [CrossRef] [Green Version]
- Albini, A.; Soldi, R.; Giunciuglio, D.; Giraudo, E.; Benelli, R.; Primo, L.; Noonan, D.; Salio, M.; Camussi, G.; Rockl, W.; et al. The angiogenesis induced by HIV-1 tat protein is mediated by the Flk-1/KDR receptor on vascular endothelial cells. Nat. Med. 1996, 2, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Mediouni, S.; Darque, A.; Baillat, G.; Ravaux, I.; Dhiver, C.; Tissot-Dupont, H.; Mokhtari, M.; Moreau, H.; Tamalet, C.; Brunet, C.; et al. Antiretroviral therapy does not block the secretion of the human immunodeficiency virus tat protein. Infect. Disord. Drug Targets 2012, 12, 81–86. [Google Scholar] [CrossRef]
- Shiu, C.; Barbier, E.; Di Cello, F.; Choi, H.J.; Stins, M. HIV-1 gp120 as well as alcohol affect blood-brain barrier permeability and stress fiber formation: Involvement of reactive oxygen species. Alcohol. Clin. Exp. Res. 2007, 31, 130–137. [Google Scholar] [CrossRef]
- Wang, T.; Green, L.A.; Gupta, S.K.; Kim, C.; Wang, L.; Almodovar, S.; Flores, S.C.; Prudovsky, I.A.; Jolicoeur, P.; Liu, Z.; et al. Transfer of intracellular HIV Nef to endothelium causes endothelial dysfunction. PLoS ONE 2014, 9, e91063. [Google Scholar] [CrossRef] [Green Version]
- Kanmogne, G.D.; Primeaux, C.; Grammas, P. HIV-1 gp120 proteins alter tight junction protein expression and brain endothelial cell permeability: Implications for the pathogenesis of HIV-associated dementia. J. Neuropathol. Exp. Neurol. 2005, 64, 498–505. [Google Scholar] [CrossRef] [Green Version]
- Acheampong, E.A.; Parveen, Z.; Muthoga, L.W.; Kalayeh, M.; Mukhtar, M.; Pomerantz, R.J. Human Immunodeficiency virus type 1 Nef potently induces apoptosis in primary human brain microvascular endothelial cells via the activation of caspases. J. Virol. 2005, 79, 4257–4269. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Fu, W.; Wang, X.; Lin, P.H.; Yao, Q.; Chen, C. HIV gp120 induces endothelial dysfunction in tumour necrosis factor-alpha-activated porcine and human endothelial cells. Cardiovasc. Res. 2010, 87, 366–374. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.B.; Khan, M.; Garcia-Barrio, M.; Powell, M.; Bond, V.C. Apoptotic effects in primary human umbilical vein endothelial cell cultures caused by exposure to virion-associated and cell membrane-associated HIV-1 gp120. J. Acquir. Immune Defic. Syndr. 2001, 27, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Didier, N.; Banks, W.A.; Créminon, C.; Dereuddre-Bosquet, N.; Mabondzo, A. HIV-1-induced production of endothelin-1 in an in vitro model of the human blood-brain barrier. Neuroreport 2002, 13, 1179–1183. [Google Scholar] [CrossRef] [PubMed]
- Caccuri, F.; Giagulli, C.; Bugatti, A.; Benetti, A.; Alessandri, G.; Ribatti, D.; Marsico, S.; Apostoli, P.; Slevin, M.A.; Rusnati, M.; et al. HIV-1 matrix protein p17 promotes angiogenesis via chemokine receptors CXCR1 and CXCR2. Proc. Natl. Acad. Sci. USA 2012, 109, 14580–14585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, D.I.R.; Zachary, I. The vascular endothelial growth factor (VEGF) family: Angiogenic factors in health and disease. Genome Biol. 2005, 6, 209–209.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaman, S.; Leppänen, V.-M.; Alitalo, K. Vascular endothelial growth factor signaling in development and disease. Development 2018, 145, 151019. [Google Scholar] [CrossRef] [Green Version]
- Roy, H.; Bhardwaj, S.; Ylä-Herttuala, S. Biology of vascular endothelial growth factors. FEBS Lett. 2006, 580, 2879–2887. [Google Scholar] [CrossRef] [Green Version]
- Rana, S.; Powe, C.E.; Salahuddin, S.; Verlohren, S.; Perschel, F.H.; Levine, R.J.; Lim, K.-H.; Wenger, J.B.; Thadhani, R.; Karumanchi, S.A. Angiogenic factors and the risk of adverse outcomes in women with suspected preeclampsia. Circulation 2012, 125, 911–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pajusola, K.; Aprelikova, O.; Korhonen, J.; Kaipainen, A.; Pertovaara, L.; Alitalo, R.; Alitalo, K. FLT4 receptor tyrosine kinase contains seven immunoglobulin-like loops and is expressed in multiple human tissues and cell lines. Cancer Res. 1992, 52, 5738–5743. [Google Scholar] [PubMed]
- Oplawski, M.; Dziobek, K.; Zmarzły, N.; Grabarek, B.; Halski, T.; Januszyk, P.; Kuś-Kierach, A.; Adwent, I.; Dąbruś, D.; Kiełbasiński, K.; et al. Expression Profile of VEGF-C, VEGF-D, and VEGFR-3 in Different Grades of Endometrial Cancer. Curr Pharm. Biotechnol. 2019, 20, 1004–1010. [Google Scholar] [CrossRef]
- Padayachee, S.; Moodley, J.; Naicker, T. A Review of Angiogenic Imbalance in HIV-Infected Hypertensive Disorders of Pregnancy. Curr. Hypertens. Rep. 2019, 21, 69. [Google Scholar] [CrossRef]
- Pandey, A.K.; Singhi, E.K.; Arroyo, J.P.; Ikizler, T.A.; Gould, E.R.; Brown, J.; Beckman, J.A.; Harrison, D.G.; Moslehi, J. Mechanisms of VEGF (Vascular Endothelial Growth Factor) Inhibitor-Associated Hypertension and Vascular Disease. Hypertension 2018, 71, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.J.; Lam, C.; Qian, C.; Yu, K.F.; Maynard, S.E.; Sachs, B.P.; Sibai, B.M.; Epstein, F.H.; Romero, R.; Thadhani, R. Soluble endoglin and other circulating antiangiogenic factors in preeclampsia. N. Engl. J. Med. 2006, 355, 992–1005. [Google Scholar] [CrossRef]
- Maynard, S.E.; Min, J.Y.; Merchan, J.; Lim, K.H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating angiogenic factors and the risk of preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkuchi, A.; Hirashima, C.; Takahashi, K.; Suzuki, H.; Matsubara, S.; Suzuki, M. Onset threshold of the plasma levels of soluble fms-like tyrosine kinase 1/placental growth factor ratio for predicting the imminent onset of preeclampsia within 4 weeks after blood sampling at 19–31 weeks of gestation. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2013, 36, 1073–1080. [Google Scholar] [CrossRef]
- Chaiworapongsa, T.; Romero, R.; Kim, Y.M.; Kim, G.J.; Kim, M.R.; Espinoza, J.; Bujold, E.; Gonçalves, L.; Gomez, R.; Edwin, S.; et al. Plasma soluble vascular endothelial growth factor receptor-1 concentration is elevated prior to the clinical diagnosis of pre-eclampsia. J. Matern. Fetal Neonatal Med. 2005, 17, 3–18. [Google Scholar] [CrossRef]
- Eremina, V.; Jefferson, J.A.; Kowalewska, J.; Hochster, H.; Haas, M.; Weisstuch, J.; Richardson, C.; Kopp, J.B.; Kabir, M.G.; Backx, P.H.; et al. VEGF inhibition and renal thrombotic microangiopathy. N. Engl. J. Med. 2008, 358, 1129–1136. [Google Scholar] [CrossRef]
- Patel, T.V.; Morgan, J.A.; Demetri, G.D.; George, S.; Maki, R.G.; Quigley, M.; Humphreys, B.D. A preeclampsia-like syndrome characterized by reversible hypertension and proteinuria induced by the multitargeted kinase inhibitors sunitinib and sorafenib. J. Natl. Cancer Inst. 2008, 100, 282–284. [Google Scholar] [CrossRef] [Green Version]
- Verlohren, S.; Herraiz, I.; Lapaire, O.; Schlembach, D.; Moertl, M.; Zeisler, H.; Calda, P.; Holzgreve, W.; Galindo, A.; Engels, T.; et al. The sFlt-1/PlGF ratio in different types of hypertensive pregnancy disorders and its prognostic potential in preeclamptic patients. Am. J. Obs. Gynecol. 2012, 206, e1–e8. [Google Scholar] [CrossRef]
- Stepan, H.; Hund, M.; Andraczek, T. Combining Biomarkers to Predict Pregnancy Complications and Redefine Preeclampsia: The Angiogenic-Placental Syndrome. Hypertension 2020, 75, 918–926. [Google Scholar] [CrossRef]
- Schrey-Petersen, S.; Stepan, H. Anti-angiogenesis and Preeclampsia in 2016. Curr. Hypertens. Rep. 2017, 19, 6. [Google Scholar] [CrossRef]
- Zeisler, H.; Llurba, E.; Chantraine, F.; Vatish, M.; Staff, A.C.; Sennström, M.; Olovsson, M.; Brennecke, S.P.; Stepan, H.; Allegranza, D. Predictive value of the sFlt-1: PlGF ratio in women with suspected preeclampsia. N. Engl. J. Med. 2016, 374, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.H.; Yuan, H.T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Maharaj, A.S.; Walshe, T.E.; Saint-Geniez, M.; Venkatesha, S.; Maldonado, A.E.; Himes, N.C.; Matharu, K.S.; Karumanchi, S.A.; D’Amore, P.A. VEGF and TGF-beta are required for the maintenance of the choroid plexus and ependyma. J. Exp. Med. 2008, 205, 491–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govender, N.; Naicker, T.; Moodley, J. Maternal imbalance between pro-angiogenic and anti-angiogenic factors in HIV-infected women with pre-eclampsia. Cardiovasc. J. Afr. 2013, 24, 174–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govender, N.; Naicker, T.; Rajakumar, A.; Moodley, J. Soluble fms-like tyrosine kinase-1 and soluble endoglin in HIV-associated preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Sansone, M.; Sarno, L.; Saccone, G.; Berghella, V.; Maruotti, G.M.; Migliucci, A.; Capone, A.; Martinelli, P. Risk of Preeclampsia in Human Immunodeficiency Virus-Infected Pregnant Women. Obstet. Gynecol. 2016, 127, 1027–1032. [Google Scholar] [CrossRef]
- Mazibuko, M.; Moodley, J.; Naicker, T. Dysregulation of circulating sTie2 and sHER2 in HIV-infected women with preeclampsia. Hypertens. Pregnancy 2019, 38, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Mafuika, S.N.; Naicker, T. The role of Tenascin-C in HIV associated pre-eclampsia. Pregnancy Hypertens. 2021, 25, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Mbhele, N.; Moodley, J.; Naicker, T. Role of angiopoietin-2, endoglin, and placental growth factor in HIV-associated preeclampsia. Hypertens. Pregnancy 2017, 36, 240–246. [Google Scholar] [CrossRef]
- Thakoordeen, S.; Moodley, J.; Naicker, T. Serum levels of platelet endothelial cell adhesion molecule-1 (PECAM-1) and soluble vascular endothelial growth factor receptor (sVEGFR)-1 and -2 in HIV associated preeclampsia. Hypertens. Pregnancy 2017, 36, 168–174. [Google Scholar] [CrossRef]
- Ngene, N.C.; Moodley, J.; Naicker, T. The performance of pre-delivery serum concentrations of angiogenic factors in predicting postpartum antihypertensive drug therapy following abdominal delivery in severe preeclampsia and normotensive pregnancy. PLoS ONE 2019, 14, e0215807. [Google Scholar]
- Zozo, B.; Govender, N.; Moodley, J.; Naicker, T. Expression of plasma nuclear factor-kappa B cells (NF-κB) and Inhibitory subunit kappa B alpha (IκB-α) in HIV-associated pre-eclampsia. Hypertens. Pregnancy 2021, 40, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Helske, S.; Vuorela, P.; Carpén, O.; Hornig, C.; Weich, H.; Halmesmäki, E. Expression of vascular endothelial growth factor receptors 1, 2 and 3 in placentas from normal and complicated pregnancies. Mol. Hum. Reprod. 2001, 7, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Munaut, C.; Lorquet, S.; Pequeux, C.; Coulon, C.; Le Goarant, J.; Chantraine, F.; Noël, A.; Goffin, F.; Tsatsaris, V.; Subtil, D. Differential expression of Vegfr-2 and its soluble form in preeclampsia. PLoS ONE 2012, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Yamaguchi, S.; Chida, K.; Shibuya, M. A single autophosphorylation site on KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-gamma and DNA synthesis in vascular endothelial cells. EMBO J. 2001, 20, 2768–2778. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, Y.; Zhang, J.; Rao, M.; Liang, H.; Liu, G. The defect of both angiogenesis and lymphangiogenesis is involved in preeclampsia. Placenta 2015, 36, 279–286. [Google Scholar] [CrossRef]
- Marini, M.; Vichi, D.; Toscano, A.; Zappoli Thyrion, G.D.; Parretti, E.; Mello, G.; Gheri, G.; Pacini, A.; Sgambati, E. Expression of vascular endothelial growth factor receptor types 1, 2 and 3 in placenta from pregnancies complicated by hypertensive disorders. Reprod. Fertil. Dev. 2007, 19, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Tsatsaris, V.; Goffin, F.; Munaut, C.; Brichant, J.F.; Pignon, M.R.; Noel, A.; Schaaps, J.P.; Cabrol, D.; Frankenne, F.; Foidart, J.M. Overexpression of the soluble vascular endothelial growth factor receptor in preeclamptic patients: Pathophysiological consequences. J. Clin. Endocrinol. Metab. 2003, 88, 5555–5563. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Liu, H.; Qiao, F.; Wu, Y.; Xu, J. VEGF deficit is involved in endothelium dysfunction in preeclampsia. J. Huazhong Univ. Sci. Technol. Med. Sci. 2010, 30, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Lecarpentier, E.; Tsatsaris, V. Angiogenic balance (sFlt-1/PlGF) and preeclampsia. Ann. Endocrinol. 2016, 77, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Lely, A.T.; Salahuddin, S.; Holwerda, K.M.; Karumanchi, S.A.; Rana, S. Circulating lymphangiogenic factors in preeclampsia. Hypertens. Pregnancy 2013, 32, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shange, G.P.; Moodley, J.; Naicker, T. Effect of vascular endothelial growth factors A, C, and D in HIV-associated pre-eclampsia. Hypertens. Pregnancy 2017, 36, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Risau, W. Development and differentiation of endothelium. Kidney Int. Suppl. 1998, 67, S3–S6. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.N.; Grimwood, J.; Taylor, R.S.; McMaster, M.T.; Fisher, S.J.; North, R.A. Longitudinal serum concentrations of placental growth factor: Evidence for abnormal placental angiogenesis in pathologic pregnancies. Am. J. Obs. Gynecol. 2003, 188, 177–182. [Google Scholar] [CrossRef]
- Sela, S.; Itin, A.; Natanson-Yaron, S.; Greenfield, C.; Goldman-Wohl, D.; Yagel, S.; Keshet, E. A novel human-specific soluble vascular endothelial growth factor receptor 1: Cell-type-specific splicing and implications to vascular endothelial growth factor homeostasis and preeclampsia. Circ. Res. 2008, 102, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.P.; Andrews, J.I.; Liu, K.Z. Intronic polyadenylation signal sequences and alternate splicing generate human soluble Flt1 variants and regulate the abundance of soluble Flt1 in the placenta. FASEB J. 2007, 21, 3885–3895. [Google Scholar] [CrossRef]
- Chaiworapongsa, T.; Romero, R.; Espinoza, J.; Bujold, E.; Mee Kim, Y.; Gonçalves, L.F.; Gomez, R.; Edwin, S. Evidence supporting a role for blockade of the vascular endothelial growth factor system in the pathophysiology of preeclampsia. Young Investigator Award. Am. J. Obs. Gynecol. 2004, 190, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.J.; Thadhani, R.; Qian, C.; Lam, C.; Lim, K.H.; Yu, K.F.; Blink, A.L.; Sachs, B.P.; Epstein, F.H.; Sibai, B.M.; et al. Urinary placental growth factor and risk of preeclampsia. JAMA 2005, 293, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, K.; Morris, R.; Kyle, P.B.; Cornelius, D.; Darby, M.; Scott, J.; Moseley, J.; Chatman, K.; Lamarca, B. Hypertension, inflammation and T lymphocytes are increased in a rat model of HELLP syndrome. Hypertens. Pregnancy 2014, 33, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Noori, M.; Donald, A.E.; Angelakopoulou, A.; Hingorani, A.D.; Williams, D.J. Prospective study of placental angiogenic factors and maternal vascular function before and after preeclampsia and gestational hypertension. Circulation 2010, 122, 478–487. [Google Scholar] [CrossRef] [Green Version]
- Schmella, M.J.; Assibey-Mensah, V.; Parks, W.T.; Roberts, J.M.; Jeyabalan, A.; Hubel, C.A.; Catov, J.M. Plasma concentrations of soluble endoglin in the maternal circulation are associated with maternal vascular malperfusion lesions in the placenta of women with preeclampsia. Placenta 2019, 78, 29–35. [Google Scholar] [CrossRef]
- Zachary, I.; Gliki, G. Signaling transduction mechanisms mediating biological actions of the vascular endothelial growth factor family. Cardiovasc Res 2001, 49, 568–581. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-L.; Hong, T.-M.; Yang, P.-C. Neuropilin 1 and Lung Cancer Progression: Potential Role of Targeting Neuropilin 1 as an Antitumor Strategy. Clin. Cancer Res. 2007, 13, 4759–4768. [Google Scholar]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [Green Version]
- Soker, S.; Takashima, S.; Miao, H.Q.; Neufeld, G.; Klagsbrun, M. Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 1998, 92, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Arad, A.; Nammouz, S.; Nov, Y.; Ohel, G.; Bejar, J.; Vadasz, Z. The Expression of Neuropilin-1 in Human Placentas from Normal and Preeclamptic Pregnancies. Int. J. Gynecol. Pathol. 2017, 36, 42–49. [Google Scholar] [CrossRef]
- Xu, X.; Yang, X.Y.; He, B.W.; Yang, W.J.; Cheng, W.W. Placental NRP1 and VEGF expression in pre-eclamptic women and in a homocysteine-treated mouse model of pre-eclampsia. Eur J. Obs. Gynecol. Reprod. Biol. 2016, 196, 69–75. [Google Scholar] [CrossRef]
- Maulik, D.; De, A.; Ragolia, L.; Evans, J.; Grigoryev, D.; Lankachandra, K.; Mundy, D.; Muscat, J.; Gerkovich, M.M.; Ye, S.Q. Down-regulation of placental neuropilin-1 in fetal growth restriction. Am. J. Obs. Gynecol. 2016, 214, e1–e279. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.Y.; Song, Y.; Wang, Y.; Mazagness, R.R.; Zheng, J. Differential expression of vascular endothelial growth factor (VEGF), endocrine gland derived-VEGF, and VEGF receptors in human placentas from normal and preeclamptic pregnancies. J. Clin. Endocrinol. Metab. 2004, 89, 2484–2490. [Google Scholar] [CrossRef] [Green Version]
- Coutard, B.; Valle, C.; de Lamballerie, X.; Canard, B.; Seidah, N.G.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antivir. Res. 2020, 176, 104742. [Google Scholar] [CrossRef]
- Obimbo, M.M.; Zhou, Y.; McMaster, M.T.; Cohen, C.R.; Qureshi, Z.; Ong’ech, J.; Ogeng’o, J.A.; Fisher, S.J. Placental structure in preterm birth among HIV-positive versus HIV-negative women in Kenya. J. Acquir. Immune Defic. Syndr. 2019, 80, 94. [Google Scholar] [CrossRef] [PubMed]
- Machado, E.S.; Krauss, M.R.; Megazzini, K.; Coutinho, C.M.; Kreitchmann, R.; Melo, V.H.; Pilotto, J.H.; Ceriotto, M.; Hofer, C.B.; Siberry, G.K. Hypertension, preeclampsia and eclampsia among HIV-infected pregnant women from Latin America and Caribbean countries. J. Infect. 2014, 68, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Hernández, S.; Catalán-García, M.; Morén, C.; García-Otero, L.; López, M.; Guitart-Mampel, M.; Milisenda, J.; Coll, O.; Cardellach, F.; Gratacós, E. Placental mitochondrial toxicity, oxidative stress, apoptosis, and adverse perinatal outcomes in HIV pregnancies under antiretroviral treatment containing zidovudine. JAIDS J. Acquir. Immune Defic. Syndr. 2017, 75, e113–e119. [Google Scholar] [CrossRef]
- Song, L.; Ding, S.; Ge, Z.; Zhu, X.; Qiu, C.; Wang, Y.; Lai, E.; Yang, W.; Sun, Y.; Chow, S.A.; et al. Nucleoside/nucleotide reverse transcriptase inhibitors attenuate angiogenesis and lymphangiogenesis by impairing receptor tyrosine kinases signalling in endothelial cells. Br. J. Pharmacol. 2018, 175, 1241–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autran, B.; Carcelaint, G.; Li, T.S.; Gorochov, G.; Blanc, C.; Renaud, M.; Durali, M.; Mathez, D.; Calvez, V.; Leibowitch, J.; et al. Restoration of the immune system with anti-retroviral therapy. Immunol. Lett. 1999, 66, 207–211. [Google Scholar] [CrossRef]
- Powis, K.M.; Shapiro, R.L. Protease Inhibitors and Adverse Birth Outcomes: Is Progesterone the Missing Piece to the Puzzle? Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Kala, S.; Dunk, C.; Acosta, S.; Serghides, L. Periconceptional exposure to lopinavir, but not darunavir, impairs decidualization: A potential mechanism leading to poor birth outcomes in HIV-positive pregnancies. Hum. Reprod. 2020, 35, 1781–1796. [Google Scholar] [CrossRef] [PubMed]
- Francisci, D.; Giannini, S.; Baldelli, F.; Leone, M.; Belfiori, B.; Guglielmini, G.; Malincarne, L.; Gresele, P. HIV type 1 infection, and not short-term HAART, induces endothelial dysfunction. Aids 2009, 23, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Powis, K.M.; McElrath, T.F.; Hughes, M.D.; Ogwu, A.; Souda, S.; Datwyler, S.A.; von Widenfelt, E.; Moyo, S.; Nádas, M.; Makhema, J.; et al. High viral load and elevated angiogenic markers associated with increased risk of preeclampsia among women initiating highly active antiretroviral therapy in pregnancy in the Mma Bana study, Botswana. J. Acquir. Immune Defic. Syndr. 2013, 62, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattar, R.; Amed, A.M.; Lindsey, P.C.; Sass, N.; Daher, S. Preeclampsia and HIV infection. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 117, 240–241. [Google Scholar] [CrossRef]
- Sgadari, C.; Barillari, G.; Toschi, E.; Carlei, D.; Bacigalupo, I.; Baccarini, S.; Palladino, C.; Leone, P.; Bugarini, R.; Malavasi, L.; et al. HIV protease inhibitors are potent anti-angiogenic molecules and promote regression of Kaposi sarcoma. Nat. Med. 2002, 8, 225–232. [Google Scholar] [CrossRef]
- Govender, N.; Moodley, J.; Gathiram, P.; Naicker, T. Soluble fms-like tyrosine kinase-1 in HIV infected pre-eclamptic South African Black women. Placenta 2014, 35, 618–624. [Google Scholar] [CrossRef]
- Srinivasa, S.; Fitch, K.V.; Wong, K.; Torriani, M.; Mayhew, C.; Stanley, T.; Lo, J.; Adler, G.K.; Grinspoon, S.K. RAAS activation is associated with visceral adiposity and insulin resistance among HIV-infected patients. J. Clin. Endocrinol. Metab. 2015, 100, 2873–2882. [Google Scholar] [CrossRef]
- Chandel, N.; Ayasolla, K.; Lan, X.; Rai, P.; Mikulak, J.; Husain, M.; Malhotra, A.; McGowan, J.; Singhal, P.C. Renin modulates HIV replication in T cells. J. Leukoc. Biol. 2014, 96, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Tzoupis, H.; Leonis, G.; Megariotis, G.; Supuran, C.T.; Mavromoustakos, T.; Papadopoulos, M.G. Dual inhibitors for aspartic proteases HIV-1 PR and renin: Advancements in AIDS–hypertension–diabetes linkage via molecular dynamics, inhibition assays, and binding free energy calculations. J. Med. Chem. 2012, 55, 5784–5796. [Google Scholar] [CrossRef] [PubMed]
- Bouba, I.; Makrydimas, G.; Kalaitzidis, R.; Lolis, D.E.; Siamopoulos, K.C.; Georgiou, I. Interaction between the polymorphisms of the renin–angiotensin system in preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 110, 8–11. [Google Scholar] [CrossRef] [Green Version]
- Aung, M.; Konoshita, T.; Moodley, J.; Gathiram, P. Association of gene polymorphisms of four components of renin-angiotensin-aldosterone system and preeclampsia in South African black women. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 215, 180–187. [Google Scholar] [CrossRef]
- Williams, P.; Mistry, H.; Innes, B.; Bulmer, J.; Pipkin, F.B. Expression of AT1R, AT2R and AT4R and their roles in extravillous trophoblast invasion in the human. Placenta 2010, 31, 448–455. [Google Scholar] [CrossRef]
- Zhou, A. Renin Angiotensin System Polymorphisms and Pregnancy Complications. Ph.D. Thesis, University of Adelaide, Adelaide, Australia, 2012. [Google Scholar]
- Khaliq, O.P.; Konoshita, T.; Moodley, J.; Naicker, T. Soluble angiotensin IV receptor levels in preeclampsia: Is there a variation? J. Matern. Fetal. Neonatal. Med. 2020, 1–6. [Google Scholar] [CrossRef]
- Khaliq, O.P.; Konoshita, T.; Moodley, J.; Naicker, T. The role of LNPEP and ANPEP gene polymorphisms in the pathogenesis of pre-eclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 252, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Pangmekeh, P.J.; Awolu, M.M.; Gustave, S.; Gladys, T.; Cumber, S.N. Association between highly active antiretroviral therapy (HAART) and hypertension in persons living with HIV/AIDS at the Bamenda regional hospital, Cameroon. Pan Afr. Med. J. 2019, 33, 87. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.A.S.; Sampaio, W.O.; Alzamora, A.C.; Motta-Santos, D.; Alenina, N.; Bader, M.; Campagnole-Santos, M.J. The ACE2/angiotensin-(1–7)/MAS axis of the renin-angiotensin system: Focus on angiotensin-(1–7). Physiol. Rev. 2017. [Google Scholar] [CrossRef] [Green Version]
- Woldesenbet, S.A.; Kufa, T.; Barron, P.; Ayalew, K.; Cheyip, M.; Chirombo, B.C.; Lombard, C.; Manda, S.; Pillay, Y.; Puren, A.J. Assessment of readiness to transition from antenatal HIV surveillance surveys to PMTCT programme data-based HIV surveillance in South Africa: The 2017 Antenatal Sentinel HIV Survey. Int. J. Infect. Dis. 2020, 91, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, T.; Moodley, J.; Naicker, T. The Involvement of MicroRNAs in SARS-CoV-2 Infection Comorbid with HIV-Associated Preeclampsia. Curr. Hypertens. Rep. 2021, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Naicker, T.; Dorsamy, E.; Ramsuran, D.; Burton, G.J.; Moodley, J. The role of apoptosis on trophoblast cell invasion in the placental bed of normotensive and preeclamptic pregnancies. Hypertens. Pregnancy 2013, 32, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Raguema, N.; Moustadraf, S.; Bertagnolli, M. Immune and Apoptosis Mechanisms Regulating Placental Development and Vascularization in Preeclampsia. Front. Physiol. 2020, 11, 98. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps: Is immunity the second function of chromatin? J. Cell Biol. 2012, 198, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V. Neutrophil Extracellular Traps in the Second Decade. J. Innate Immun. 2018, 10, 414–421. [Google Scholar] [CrossRef]
- Moodley, M.; Moodley, J.; Naicker, T. The Role of Neutrophils and Their Extracellular Traps in the Synergy of Pre-eclampsia and HIV Infection. Curr. Hypertens. Rep. 2020, 22, 1–9. [Google Scholar] [CrossRef]
- Mutua, V.; Gershwin, L.J. A review of neutrophil extracellular traps (NETs) in disease: Potential anti-NETs therapeutics. Clin. Rev. Allergy Immunol. 2020, 1–18. [Google Scholar] [CrossRef]
- Thierry, A.; Benoit, R. NETs by-products and extracellular DNA may play a key role in COVID-19 pathogenesis: Incidence on patient monitoring and therapy. Preprints 2020. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Ye, J.; Pan, N.; Tan, C.; Herrmann, M. Neutrophil Extracellular Traps Tied to Rheumatoid Arthritis: Points to Ponder. Front. Immunol. 2021, 11, 3668. [Google Scholar] [CrossRef]
- Gupta, A.K.; Hasler, P.; Holzgreve, W.; Gebhardt, S.; Hahn, S. Induction of neutrophil extracellular DNA lattices by placental microparticles and IL-8 and their presence in preeclampsia. Hum. Immunol. 2005, 66, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Moodley, M.; Moodley, J.; Naicker, T. Neutrophil extracellular traps: The synergy source in the placentae of HIV infected women with pre-eclampsia. Pregnancy Hypertens. 2020, 20, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Joshi, M.B.; Philippova, M.; Erne, P.; Hasler, P.; Hahn, S.; Resink, T.J. Activated endothelial cells induce neutrophil extracellular traps and are susceptible to NETosis--mediated cell death. FEBS Lett. 2010, 584, 3193–3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.Y.; Laivuori, H.; Livingston, J.C.; Ylikorkala, O.; Sibai, B.M.; Holzgreve, W.; Hahn, S. Elevation of both maternal and fetal extracellular circulating deoxyribonucleic acid concentrations in the plasma of pregnant women with preeclampsia. Am. J. Obstet. Gynecol. 2001, 184, 414–419. [Google Scholar] [CrossRef]
- Konečná, B.; Lauková, L.; Vlková, B. Immune activation by nucleic acids: A role in pregnancy complications. Scand. J. Immunol. 2018, 87, e12651. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Komano, J.; Saitoh, Y.; Misawa, T.; Takahama, M.; Kozaki, T.; Uehata, T.; Iwasaki, H.; Omori, H.; Yamaoka, S. Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Cell Host Microbe 2012, 12, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlowski, H.N.; Lai, E.T.; Havugimana, P.C.; White, C.; Emili, A.; Sakac, D.; Binnington, B.; Neschadim, A.; McCarthy, S.D.; Branch, D.R. Extracellular histones identified in crocodile blood inhibit in-vitro HIV-1 infection. Aids 2016, 30, 2043–2052. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, C.P.; Schatz, F.; Rahman, M.; Abrahams, V.; Lockwood, C. Pre--eclampsia is associated with dendritic cell recruitment into the uterine decidua. J. Pathol. 2008, 214, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Brander, C. Considerations for successful therapeutic immunization in HIV cure. Curr. Opin. HIV AIDS 2021, 16, 257–261. [Google Scholar] [CrossRef]


| Antiretroviral Type | Main Findings | References |
|---|---|---|
| Nucleoside/nucleotide reverse transcriptase inhibitors |
| [121,122] |
| Protease inhibitors |
| [123,124,125] |
| HAART |
| [83,88,126,127,128,129,130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naicker, T.; Govender, N.; Abel, T.; Naidoo, N.; Moodley, M.; Pillay, Y.; Singh, S.; Khaliq, O.P.; Moodley, J. HIV Associated Preeclampsia: A Multifactorial Appraisal. Int. J. Mol. Sci. 2021, 22, 9157. https://doi.org/10.3390/ijms22179157
Naicker T, Govender N, Abel T, Naidoo N, Moodley M, Pillay Y, Singh S, Khaliq OP, Moodley J. HIV Associated Preeclampsia: A Multifactorial Appraisal. International Journal of Molecular Sciences. 2021; 22(17):9157. https://doi.org/10.3390/ijms22179157
Chicago/Turabian StyleNaicker, Thajasvarie, Nalini Govender, Tashlen Abel, Nitalia Naidoo, Merantha Moodley, Yazira Pillay, Shoohana Singh, Olive Pearl Khaliq, and Jagidesa Moodley. 2021. "HIV Associated Preeclampsia: A Multifactorial Appraisal" International Journal of Molecular Sciences 22, no. 17: 9157. https://doi.org/10.3390/ijms22179157