
Palmitoylation of Prolactin-Releasing Peptide Increased Affinity for and Activation of the GPR10, NPFF-R2 and NPFF-R1 Receptors: In Vitro Study

Abstract
:1. Introduction
2. Results
2.1. Binding Affinity for GPR10, NPFF-R2 and Potential Off-Target Receptors
2.1.1. Palmitoylated PrRP31 Analogs Have a High Binding Affinity for GPR10, NPFF-R2 and NPFF-R1
2.1.2. Palm-PrRP31 Shows a Higher Affinity for Other Potential Off-Target Receptors than Palm11-PrRP31
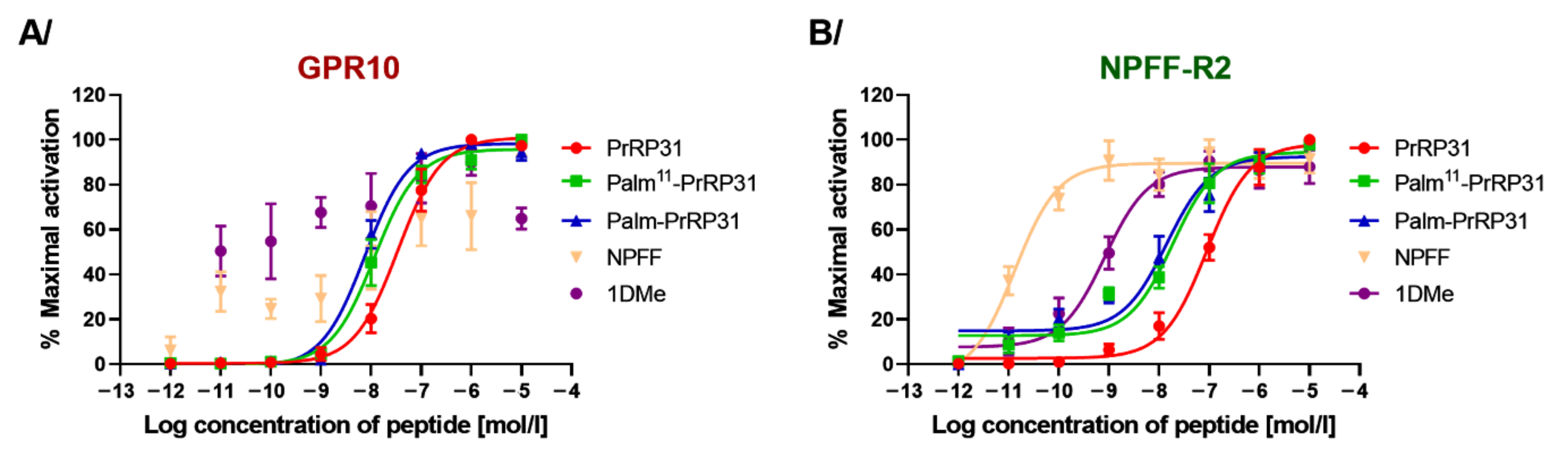
2.2. PrRP31 and Its Palmitoylated Analogs Stimulate Ca2+ Mobilization in CHO-K1 Cells Expressing GPR10 or NPFF-R2
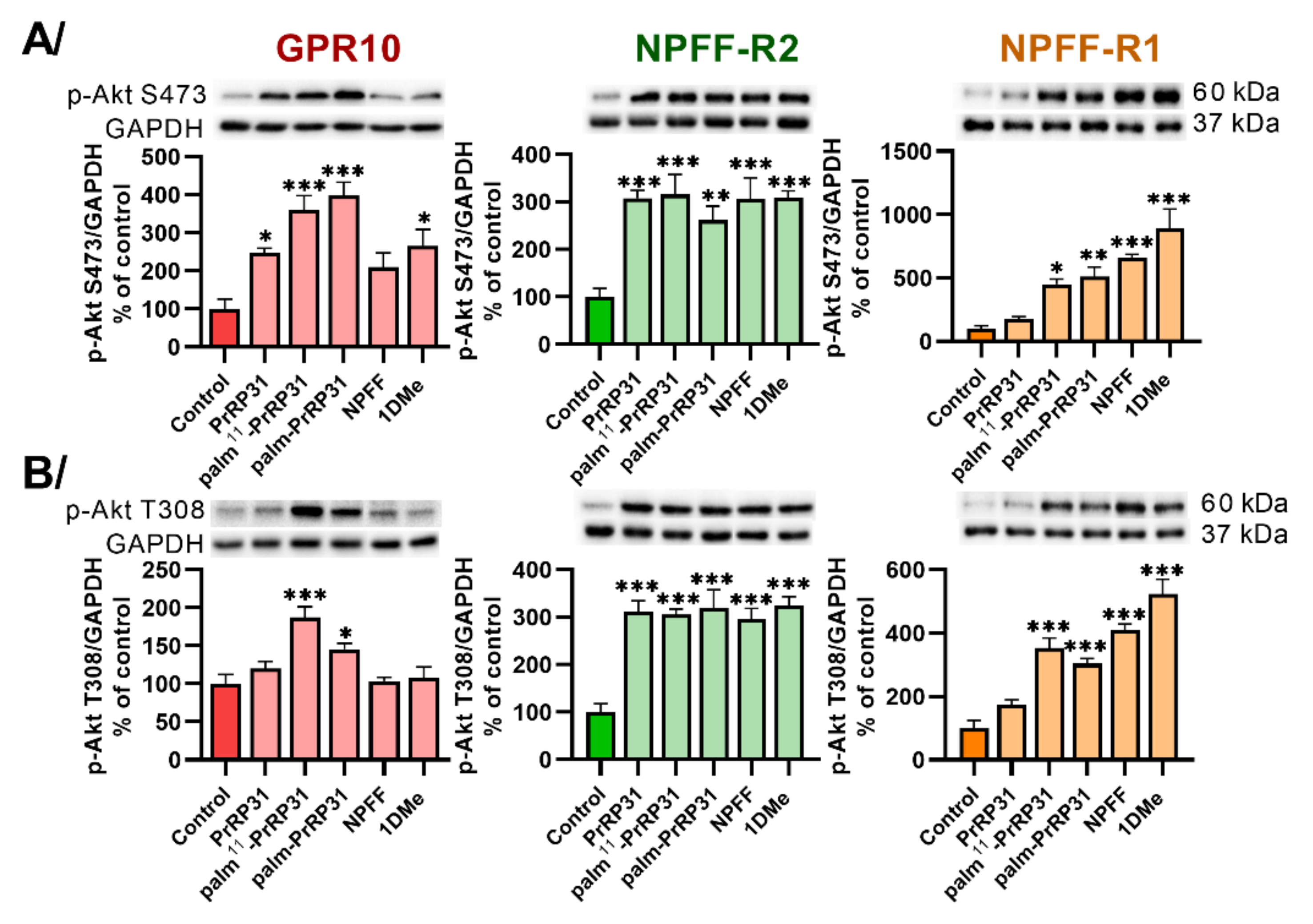
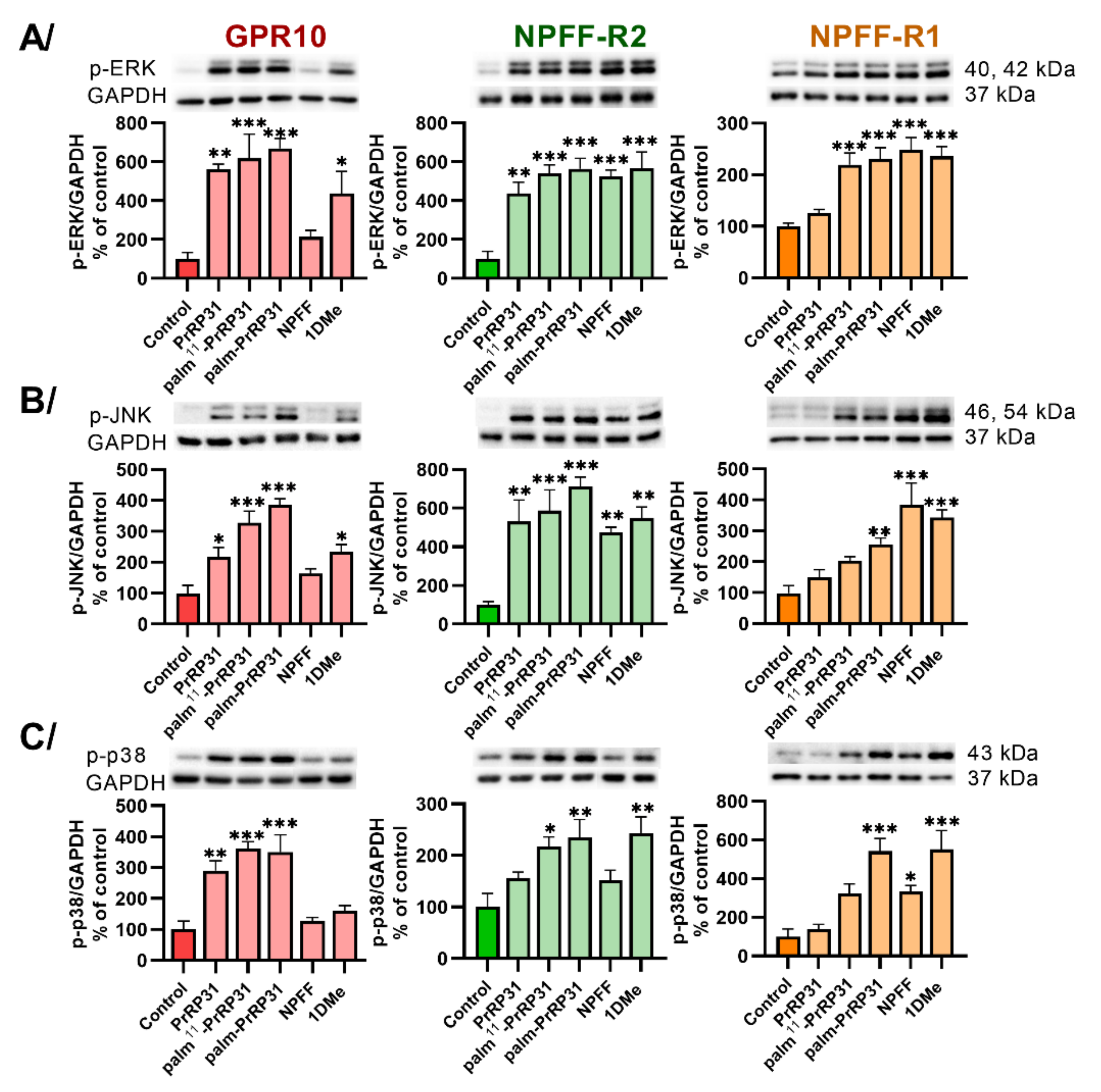
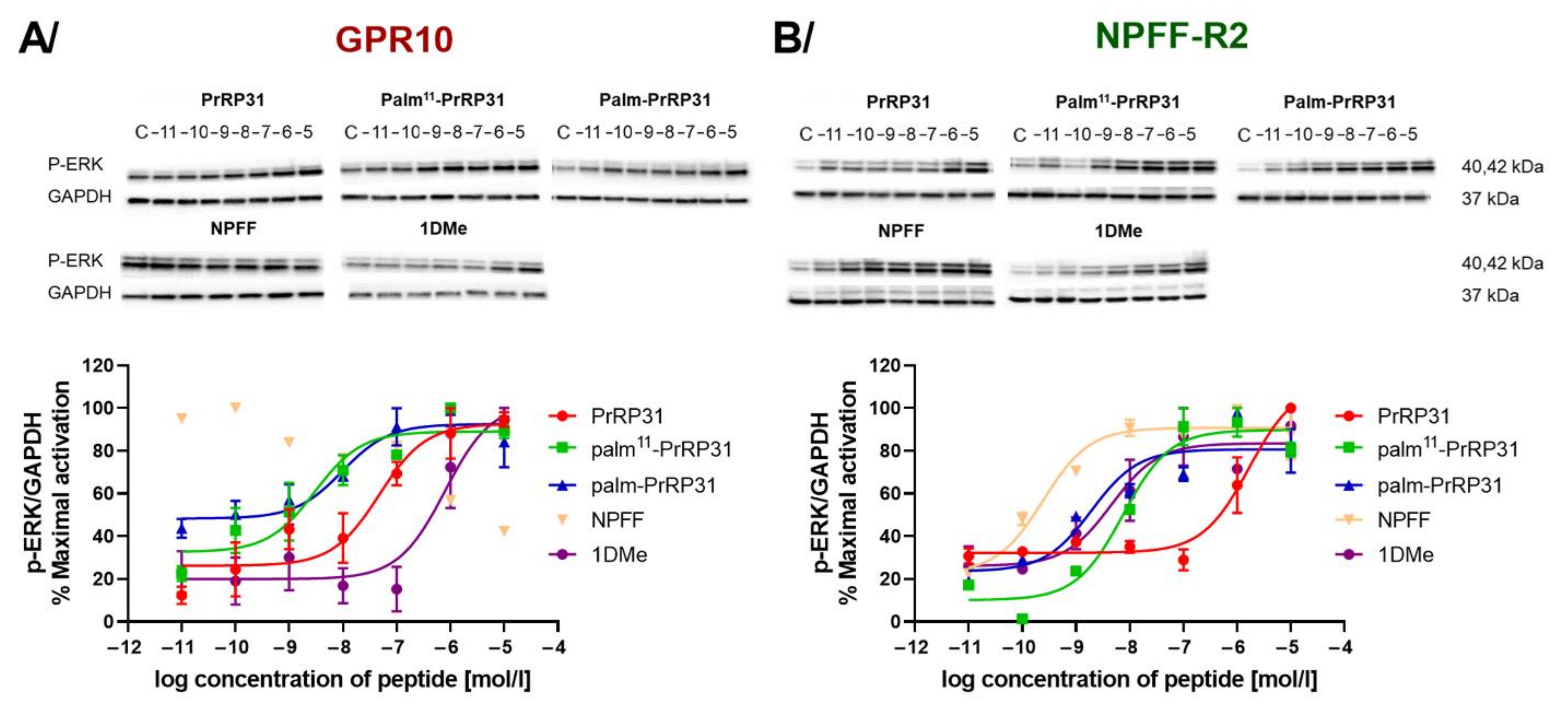
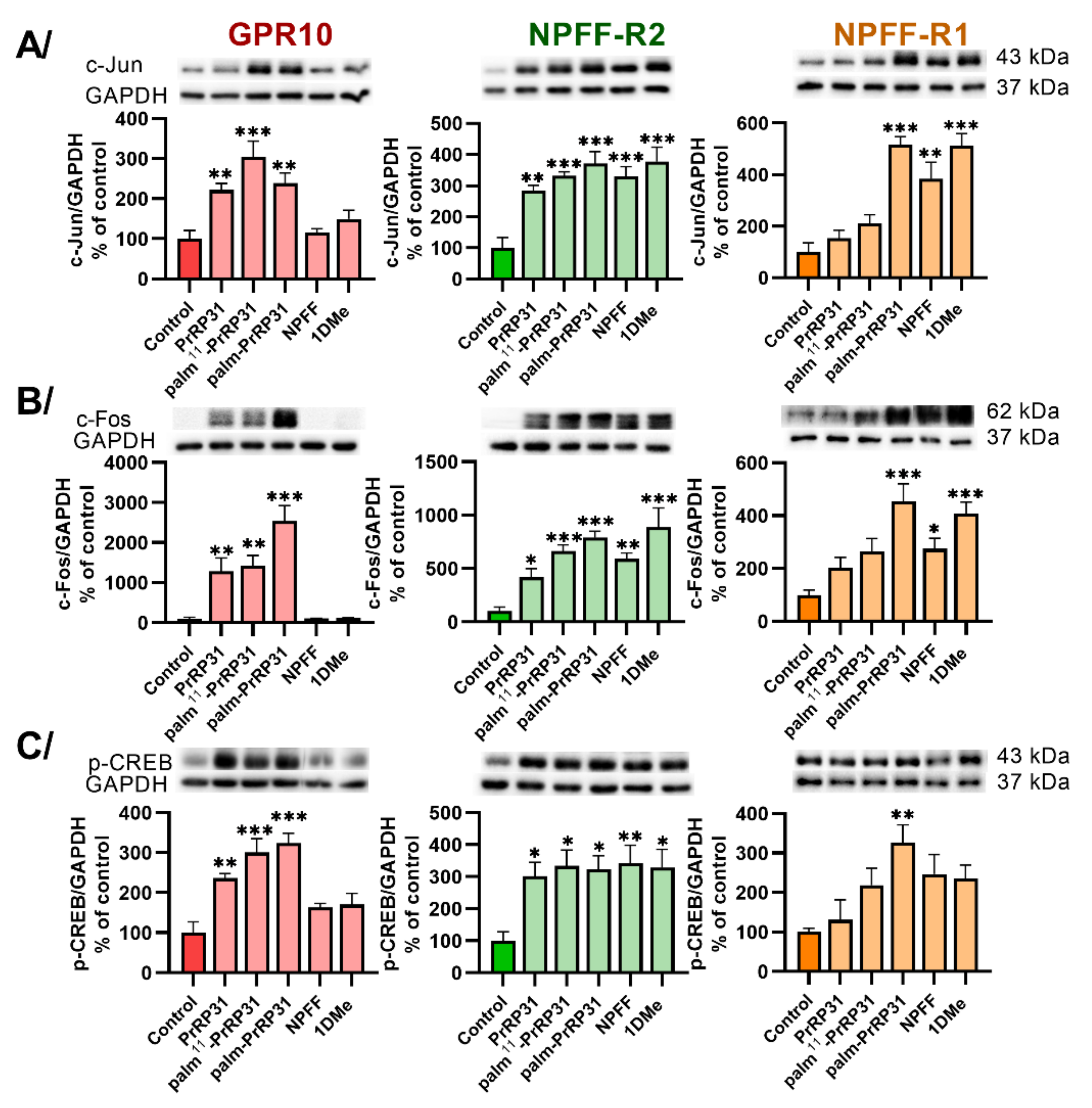
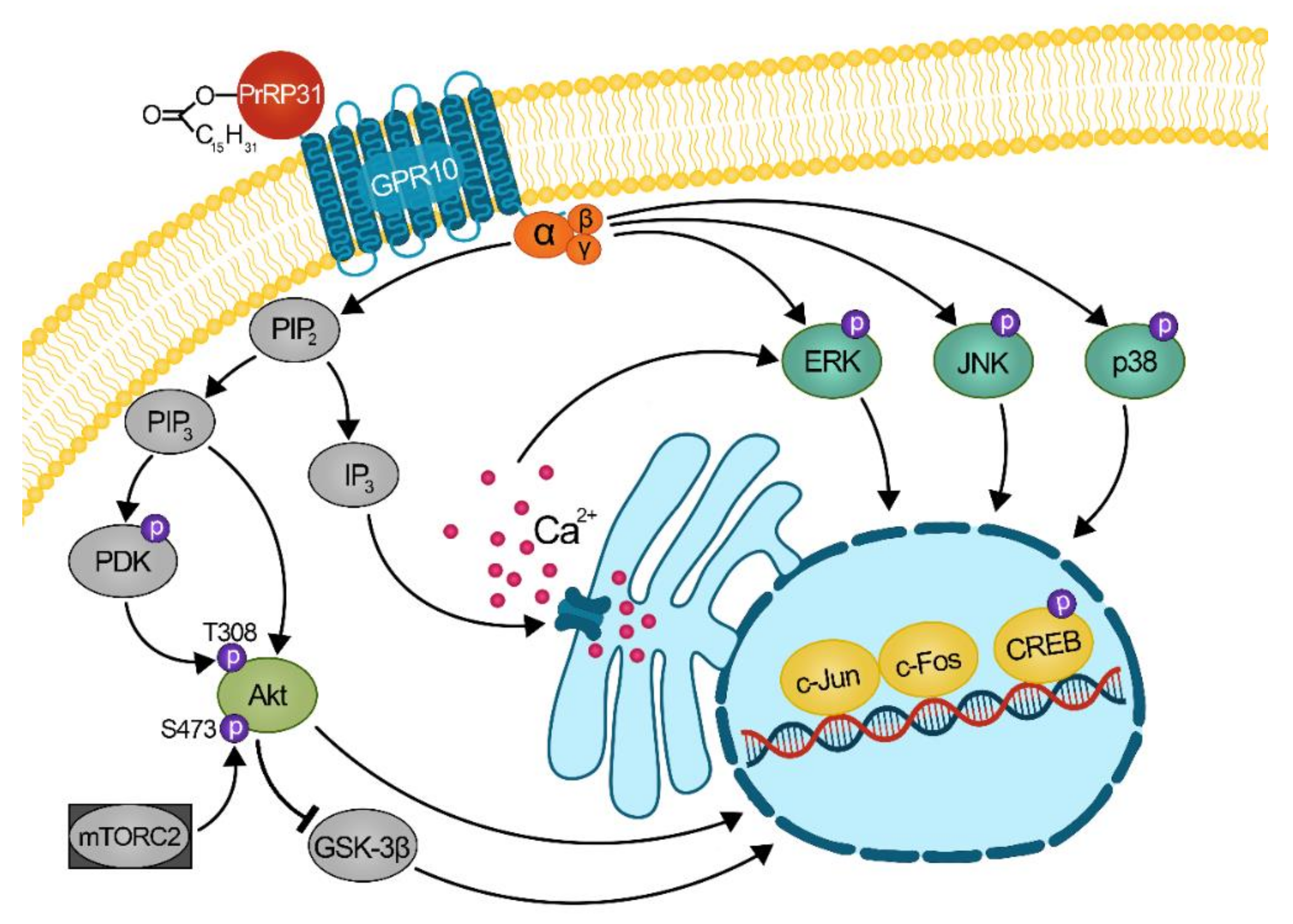
2.3. Palmitoylated PrRP31 Analogs Activate Different Intracellular Signaling Pathways in GPR10-, NPFF-R2- or NPFF-R1-Expressing Cells
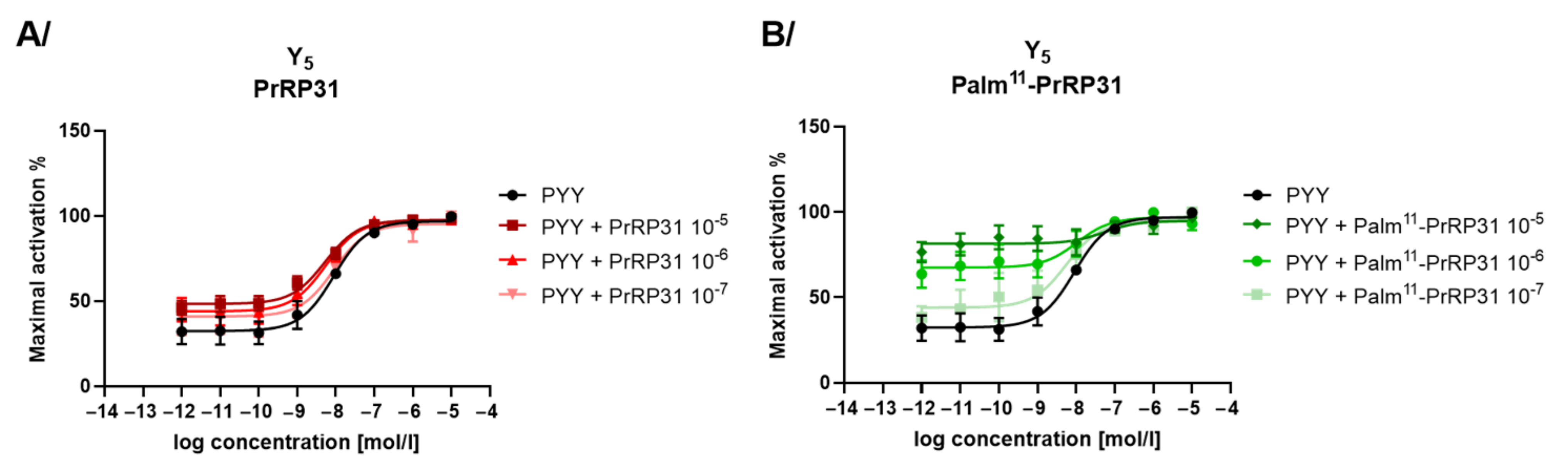
2.4. Agonist and Antagonist Properties of PrRP31 and Its Palmitoylated Analogs at Other Potential Off-Target Receptors
3. Discussion
4. Materials and Methods
4.1. Material
4.2. Peptide Iodination
4.3. Cell Cultures
4.4. Cell Membrane Isolation
4.5. Competitive Binding Experiments
4.6. Cell Signaling Detection by Immunoblotting
4.7. Calcium Mobilization Assays
4.8. Cell Signaling Determined Using Beta-Lactamase Reporter System
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hinuma, S.; Habata, Y.; Fujii, R.; Kawamata, Y.; Hosoya, M.; Fukusumi, S.; Kitada, C.; Masuo, Y.; Asano, T.; Matsumoto, H.; et al. A prolactin-releasing peptide in the brain. Nature 1998, 393, 272–276. [Google Scholar] [CrossRef]
- Matsumoto, H.; Noguchi, J.; Horikoshi, Y.; Kawamata, Y.; Kitada, C.; Hinuma, S.; Onda, H.; Nishimura, O.; Fujino, M. Stimulation of Prolactin Release by Prolactin-Releasing. Biochem. Biophys. Res. Commun. 1999, 259, 321–324. [Google Scholar] [CrossRef]
- Lawrence, C.B.; Celsi, F.; Brennand, J.; Luckman, S.M. Alternative role for prolactin-releasing peptide in the regulation of food intake. Nat. Neurosc. 2000, 3, 645–646. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.B.; Ellacott, K.L.J.; Luckman, S.M. PRL-releasing peptide reduces food intake and may mediate satiety signaling. Endocrinology 2002, 143, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Bjursell, M.; Lenneras, M.; Goransson, M.; Elmgren, A.; Bohlooly, Y.M. GPR10 deficiency in mice results in altered energy expenditure and obesity. Biochem. Biophys. Res. Commun. 2007, 363, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Geddes, B.J.; Zhang, C.; Foley, K.P.; Stricker-Krongrad, A. The prolactin-releasing peptide receptor (GPR10) regulates body weight homeostasis in mice. J. Mol. Neurosc. 2004, 22, 93–103. [Google Scholar] [CrossRef]
- Prazienkova, V.; Funda, J.; Pirnik, Z.; Karnosova, A.; Hruba, L.; Korinkova, L.; Neprasova, B.; Janovska, P.; Benzce, M.; Kadlecova, M.; et al. GPR10 gene deletion in mice increases basal neuronal activity, disturbs insulin sensitivity and alters lipid homeostasis. Gene 2021, 774, 145427. [Google Scholar] [CrossRef]
- Maletinska, L.; Nagelova, V.; Ticha, A.; Zemenova, J.; Pirnik, Z.; Holubova, M.; Spolcova, A.; Mikulaskova, B.; Blechova, M.; Sykora, D.; et al. Novel lipidized analogs of prolactin-releasing peptide have prolonged half-lives and exert anti-obesity effects after peripheral administration. Int. J. Obesity 2015, 39, 986–993. [Google Scholar] [CrossRef]
- Prazienkova, V.; Holubova, M.; Pelantova, H.; Buganova, M.; Pirnik, Z.; Mikulaskova, B.; Popelova, A.; Blechova, M.; Haluzik, M.; Zelezna, B.; et al. Impact of novel palmitoylated prolactin-releasing peptide analogs on metabolic changes in mice with diet-induced obesity. PLoS ONE 2017, 12, e0183449. [Google Scholar] [CrossRef]
- Prazienkova, V.; Ticha, A.; Blechova, M.; Spolcova, A.; Zelezna, B.; Maletinska, L. Pharmacological characterization of lipidized analogs of prolactin-releasing peptide with a modified C-terminalaromatic ring. J. Physiol. Pharmacol. 2016, 67, 121–128. [Google Scholar] [PubMed]
- Pirnik, Z.; Kolesarova, M.; Zelezna, B.; Maletinska, L. Repeated peripheral administration of lipidized prolactin-releasing peptide analog induces c-fos and FosB expression in neurons of dorsomedial hypothalamic nucleus in male C57 mice. Neurochem. Int. 2018, 116, 77–84. [Google Scholar] [CrossRef]
- Pirnik, Z.; Zelezna, B.; Kiss, A.; Maletinska, L. Peripheral administration of palmitoylated prolactin-releasing peptide induces Fos expression in hypothalamic neurons involved in energy homeostasis in NMRI male mice. Brain Res. 2015, 1625, 151–158. [Google Scholar] [CrossRef]
- Engstrom, M.; Brandt, A.; Wurster, S.; Savola, J.M.; Panula, P. Prolactin releasing peptide has high affinity and efficacy at neuropeptide FF2 receptors. J. Pharmacol. Exp. Therapeut. 2003, 305, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Elhabazi, K.; Humbert, J.P.; Bertin, I.; Schmitt, M.; Bihel, F.; Bourguignon, J.J.; Bucher, B.; Becker, J.A.; Sorg, T.; Meziane, H.; et al. Endogenous mammalian RF-amide peptides, including PrRP, kisspeptin and 26RFa, modulate nociception and morphine analgesia via NPFF receptors. Neuropharmacology 2013, 75, 164–171. [Google Scholar] [CrossRef]
- Murase, T.; Arima, H.; Kondo, K.; Oiso, Y. Neuropeptide FF reduces food intake in rats. Peptides 1996, 17, 353–354. [Google Scholar] [CrossRef]
- Sunter, D.; Hewson, A.K.; Lynam, S.; Dickson, S.L. Intracerebroventricular injection of neuropeptide FF, an opioid modulating neuropeptide, acutely reduces food intake and stimulates water intake in the rat. Neurosci. Lett. 2001, 313, 145–148. [Google Scholar] [CrossRef]
- Mouledous, L.; Mollereau, C.; Zajac, J.M. Opioid-modulating properties of the neuropeptide FF system. BioFactors 2010, 36, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Nicklous, D.M.; Simansky, K.J. Neuropeptide FF exerts pro- and anti-opioid actions in the parabrachial nucleus to modulate food intake. Am. J. Physiol. Regulat. Integr. Compar. Physiol. 2003, 285, 1046–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panula, P.; Aarnisalo, A.A.; Wasowicz, K. Neuropeptide FF, a mammalian neuropeptide with multiple functions. Progress Neurobiol. 1996, 48, 461–479. [Google Scholar] [CrossRef]
- Simonin, F.; Schmitt, M.; Laulin, J.-P.; Laboureyras, E.; Jhamandas, J.H.; MacTavish, D.; Matifas, A.; Mollereau, C.; Laurent, P. RF9, a potent and selective neuropeptide FF receptor antagonist, prevents opioid-induced tolerance associated with hyperalgesia. Proc. Natl. Acad. Sci. USA 2006, 103, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Maletinska, L.; Ticha, A.; Nagelova, V.; Spolcova, A.; Blechova, M.; Elbert, T.; Zelezna, B. Neuropeptide FF analog RF9 is not an antagonist of NPFF receptor and decreases food intake in mice after its central and peripheral administration. Brain Res. 2013, 1498, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Kalliomäki, M.L.; Pertovaara, A.; Brandt, A.; Wei, H.; Pietilä, P.; Kalmari, J.; Xu, M.; Kalso, E.; Panula, P. Prolactin-releasing peptide affects pain, allodynia and autonomicreflexes through medullary mechanisms. Neuropharmacology 2004, 46, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Laurent, P.; Becker, J.A.; Valverde, O.; Ledent, C.; de Kerchove d’Exaerde, A.; Schiffmann, S.N.; Maldonado, R.; Vassart, G.; Parmentier, M. The prolactin-releasing peptide antagonizes the opioid system through its receptor GPR10. Nat. Neurosci. 2005, 8, 1735–1741. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, J.C.; Felix, R.C.; Fonseca, V.G.; Power, D.M. Feeding and the rhodopsin family g-protein coupled receptors in nematodes and arthropods. Front. Endocrinol. 2012, 3, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, A.; Heiber, M.; Nguyen, T.; Heng, H.H.Q.; Saldivia, V.R.; Cheng, R.; Murphy, P.M.; Tsui, L.C.; Shi, X.; Gregor, P.; et al. Cloning and Chromosomal Mapping of Three Novel Genes, GPR9, GPR10, and GPR14, Encoding Receptors Related to Interleukin 8, Neuropeptide Y, and Somatostatin Receptors. Genomics 1995, 29, 335–344. [Google Scholar] [CrossRef]
- Bonini, J.A.; Jones, K.A.; Adham, N.; Forray, C.; Artymyshyn, R.; Durkin, M.M.; Smith, K.E.; Tamm, J.A.; Boteju, L.W.; Lakhlani, P.P.; et al. Identification and characterization of two G protein-coupled receptors for neuropeptide FF. J. Biol. Chem. 2000, 275, 39324–39331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, C.J.; Szekeres, P.G.; Chambers, J.K.; Ratcliffe, S.J.; Jones, D.N.; Hirst, W.D.; Price, G.W.; Herdon, H.J. Characterization of the binding of [(125)I]-human prolactin releasing peptide (PrRP) to GPR10, a novel G protein coupled receptor. British J. Pharmacol. 2000, 131, 683–688. [Google Scholar] [CrossRef] [Green Version]
- Roland, B.L.; Sutton, S.W.; Wilson, S.J.; Luo, L.; Pyati, J.; Huvar, R.; Erlander, M.G.; Lovenberg, T.W. Anatomical distribution of prolactin-releasing peptide and its receptor suggests additional functions in the central nervous system and periphery. Endocrinology 1999, 140, 5736–5745. [Google Scholar] [CrossRef]
- Chuderland, D.; Seger, R. Calcium regulates ERK signaling by modulating its protein-protein interactions. Commun. Integr. Biol. 2008, 1, 4–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, A.; Ohmichi, M.; Tasaka, K.; Kanda, Y.; Ikegami, H.; Hayakawa, J.; Hisamoto, K.; Morishige, K.; Hinuma, S.; Kurachi, H.; et al. Prolactin-releasing peptide activation of the prolactin promoter is differentially mediated by extracellular signal-regulated protein kinase and c-Jun N-terminal protein kinase. J. Biol. Chem. 2000, 275, 3667–3674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maixnerova, J.; Spolcova, A.; Pychova, M.; Blechova, M.; Elbert, T.; Rezacova, M.; Zelezna, B.; Maletinska, L. Characterization of prolactin-releasing peptide: Binding, signaling and hormone secretion in rodent pituitary cell lines endogenously expressing its receptor. Peptides 2011, 32, 811–817. [Google Scholar] [CrossRef]
- Hayakawa, J.; Ohmichi, M.; Tasaka, K.; Kanda, Y.; Adachi, K.; Nishio, Y.; Hisamoto, K.; Mabuchi, S.; Hinuma, S.; Murata, Y. Regulation of the PRL promoter by Akt through cAMP response element binding protein. Endocrinology 2002, 143, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Prusoff, W.H. Relationship between the inhibition constant (K1) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem. Pharmacol. 1973, 22, 3099–3108. [Google Scholar] [CrossRef] [PubMed]
- Holubova, M.; Zemenova, J.; Mikulaskova, B.; Panajotova, V.; Stohr, J.; Haluzik, M.; Kunes, J.; Zelezna, B.; Maletinska, L. Palmitoylated PrRP analog decreases body weight in DIO rats but not in ZDF rats. J. Endocrinol. 2016, 229, 85–96. [Google Scholar] [CrossRef]
- Maletinska, L.; Popelova, A.; Zelezna, B.; Bencze, M.; Kunes, J. The impact of anorexigenic peptides in experimental models of Alzheimer's disease pathology. J. Endocr. 2019, 240, R47–R72. [Google Scholar] [CrossRef] [Green Version]
- Nanmoku, T.; Takekoshi, K.; Isobe, K.; Kawakami, Y.; Nakai, T.; Okuda, Y. Prolactin-releasing peptide stimulates catecholamine release but not proliferation in rat pheochromocytoma PC12 cells. Neurosci. Lett. 2003, 350, 33–36. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.Y.; Wu, Y.; Huang, G.; Li, J.; Leung, F.C. Identification of the receptors for prolactin-releasing peptide (PrRP) and Carassius RFamide peptide (C-RFa) in chickens. Endocrinology 2012, 153, 1861–1874. [Google Scholar] [CrossRef]
- Nanmoku, T.; Takekoshi, K.; Fukuda, T.; Ishii, K.; Isobe, K.; Kawakami, Y. Stimulation of catecholamine biosynthesis via the PKC pathway by prolactin-releasing peptide in PC12 rat pheochromocytoma cells. J. Endocrinol. 2005, 186, 233–239. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [Green Version]
- Canovas, B.; Nebreda, A.R. Diversity and versatility of p38 kinase signalling in health and disease. Nat. Rev. Mol. Cell Biol. 2021. [Google Scholar] [CrossRef]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The role of the transcription factor CREB in immune function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, K.; Herzog, H.; Shi, Y.C. Regulation of energy homeostasis by the NPY system. Trends Endocrinol. Metab. 2015, 26, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.D.; Mitchell, N.F.; Lin, S.; Macia, L.; Yulyaningsih, E.; Baldock, P.A.; Enriquez, R.F.; Zhang, L.; Shi, Y.C.; Zolotukhin, S.; et al. Y1 and Y5 receptors are both required for the regulation of food intake and energy homeostasis in mice. PLoS ONE 2012, 7, e40191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maletinska, L.; Pychova, M.; Holubova, M.; Blechova, M.; Demianova, Z.; Elbert, T.; Zelezna, B. Characterization of new stable ghrelin analogs with prolonged orexigenic potency. J. Pharmacol. Exp. Ther. 2012, 340, 781–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spolcova, A.; Mikulaskova, B.; Holubova, M.; Nagelova, V.; Pirnik, Z.; Zemenova, J.; Haluzik, M.; Zelezna, B.; Galas, M.C.; Maletinska, L. Anorexigenic lipopeptides ameliorate central insulin signaling and attenuate tau phosphorylation in hippocampi of mice with monosodium glutamate-induced obesity. J. Alzheimers Dis. 2015, 45, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Motulsky, H.; Neubig, R. Analyzing radioligand binding data. Curr. Protoc. Neurosci. 2002, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analog | Sequence |
|---|---|
| PrRP31 | SRAHQHSMETRTPDINPAWYTGRGIRPVGRF-NH2 |
| Palm11-PrRP31 | SRTHRHSMEIK(γ-E (N-palm))TPDINPAWYASRGIRPVGRF-NH2 |
| Palm-PrRP31 | (N-palm)SRTHRHSMEIRTPDINPAWYASRRGIRPVGRF-NH2 |
| NPFF | FLFQPQRF-NH2 |
| 1DMe | yL(N-Me)FQPQRF-NH2 |
| Receptor | GPR10 | NPFF-R2 | NPFF-R1 | KOR |
|---|---|---|---|---|
| [125I]-PrRP31 | [125I]-1DMe | [125I]-1DMe | [125I]-Dynorphin | |
| Ki [nM] | ||||
| PrRP31 | 4.58 ± 0.66 | 26.73 ± 9.01 | 40.39 ± 4.20 | >10,000 |
| Palm11-PrRP31 | 3.44 ± 0.36 | 7.66 ± 1.33 | 13.52 ± 1.57 | 4278 ± 866 |
| Palm-PrRP31 | 4.04 ± 0.01 | 0.77 ± 0.19 | 0.78 ± 0.11 | 106 ± 15 |
| NPFF | >10,000 | 0.28 ± 0.06 | 1.08 ± 0.09 | - |
| 1DMe | >10,000 | 1.03 ± 0.23 | 0.79 ± 0.06 | - |
| Dynorphin | - | - | - | 0.36 ± 0.03 |
| Receptor | Y1 | Y2 | Y5 | GHSR |
| [125I]-PYY | [125I]-PYY | [125I]-PYY | [125I]-Ghrelin | |
| Ki [nM] | ||||
| PYY | 2.92 ± 0.28 | 6.51 ± 0.71 | 3.06 ± 0.49 | - |
| PrRP31 | >10,000 | >10,000 | 2863 ± 43 | >10,000 |
| Palm11-PrRP31 | >10,000 | >10,000 | 362 ± 96 | 2800 ± 466 |
| Palm-PrRP31 | 3147 ± 31 | >10,000 | 32.62 ± 6.16 | 160 ± 16 |
| Ghrelin | - | - | - | 4.59 ± 0.41 |
| PrRP31 | Palm11-PrRP31 | Palm-PrRP31 | NPFF | 1DMe | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Receptor | GPR10 | NPFF-R2 | NPFF-R1 | GPR10 | NPFF-R2 | NPFF-R1 | GPR10 | NPFF-R2 | NPFF-R1 | GPR10 | NPFF-R2 | NPFF-R1 | GPR10 | NPFF-R2 | NPFF-R1 |
| p-ERK | ↑ ** | ↑ ** | - | ↑ *** | ↑ *** | ↑ *** | ↑ *** | ↑ *** | ↑ *** | - | ↑ *** | ↑ *** | ↑ * | ↑ *** | ↑ *** |
| p-JNK | ↑ * | ↑ ** | - | ↑ *** | ↑ *** | - | ↑ *** | ↑ *** | ↑ ** | - | ↑ ** | ↑ *** | ↑ * | ↑ ** | ↑ *** |
| p-p38 | ↑ ** | - | - | ↑ *** | ↑ * | - | ↑ *** | ↑ ** | ↑ *** | - | - | ↑ * | - | ↑ ** | ↑ *** |
| p-Akt S473 | ↑ * | ↑ *** | - | ↑ *** | ↑ *** | ↑ * | ↑ *** | ↑ ** | ↑ ** | - | ↑ *** | ↑ *** | ↑ * | ↑ *** | ↑ *** |
| p-Akt T308 | - | ↑ *** | - | ↑ *** | ↑ *** | ↑ *** | ↑ * | ↑ *** | ↑ *** | - | ↑ *** | ↑ *** | - | ↑ *** | ↑ *** |
| p-PKA | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| c-Jun | ↑ ** | ↑ ** | - | ↑ *** | ↑ *** | - | ↑ ** | ↑ *** | ↑ *** | - | ↑ *** | ↑ ** | - | ↑ *** | ↑ *** |
| c-Fos | ↑ ** | ↑ * | - | ↑ ** | ↑ *** | - | ↑ *** | ↑ *** | ↑ *** | - | ↑ ** | ↑ * | - | ↑ *** | ↑ *** |
| p-CREB | ↑ ** | ↑ * | - | ↑ *** | ↑ * | - | ↑ *** | ↑ * | ↑ ** | - | ↑ ** | - | - | ↑ * | - |
| Receptor | GPR10 | Y5 | GHSR | KOR | DOR | MOR | ORL-1 |
|---|---|---|---|---|---|---|---|
| EC50 [pM] | EC50 [nM] | ||||||
| PrRP31 | 530.3 ± 70.5 | N | N | N | N | N | N |
| PYY | 19.4 ± 2.5 | ||||||
| Ghrelin | 2.8 ± 2.5 | ||||||
| U-50488 | 1.4 ± 1.0 | ||||||
| Deltorphin II | 5.6 ± 9.9 | ||||||
| DAMGO | 14.7 ± 1.9 | ||||||
| Nociceptin | 3.8 ± 0.6 | ||||||
| Palm11-PrRP31 | 39.1 ± 5.1 | 583.3 ± 121.1 | 1068.1 ± 272.2 | >10,000 | N | N | N |
| Palm-PrRP31 | 71.8 ± 6.4 | 56.5 ± 18.4 | 1273.5 ± 167.9 | >10,000 | N | N | N |
| Antibody Against | Source | Dilution |
|---|---|---|
| Phospho-Akt (Thr308) (#2965) | Rabbit | 1:1000, 5% BSA, TBS/T-20 |
| Phospho-Akt (Ser473) (#4060) Akt (#4691S) | Rabbit Rabbit | 1:1000, 5% BSA, TBS/T-20 1:1000, 5% BSA, TBS/T-20 |
| Phospho-CREB (Ser133) (#9196) CREB (#9104S) | Mouse Mouse | 1:1000, 5% milk, TBS/T-20 1:1000, 5% milk, TBS/T-20 |
| Phospho-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) (#4370S) p44/42 MAPK (Erk1/2) (#9107S) | Rabbit Mouse | 1:2000, 5% BSA, TBS/T-20 1:2000, 5% milk, TBS/T-20 |
| Phospho-SAPK/JNK (Thr183/Tyr185) (#4668) SAPK/JNK (#9252) | Rabbit Rabbit | 1:1000, 5% BSA, TBS/T-20 1:1000, 5% BSA, TBS/T-20 |
| Phospho-p38 MAPK (Thr180/Tyr182) (#4511) p38 MAPK (#9212) | Rabbit Rabbit | 1:1000, 5% BSA, TBS/T-20 1:1000, 5% BSA, TBS/T-20 |
| Phospho-PKA C (Thr197) (#5661) | Rabbit | 1:1000, 5% BSA, TBS/T-20 |
| c-Fos (#2250) | Rabbit | 1:1000, 5% BSA, TBS/T-20 |
| c-Jun (#9165) | Rabbit | 1:1000, 5% BSA, TBS/T-20 |
| GAPDH (#97166) | Mouse | 1:1000, 5% milk, TBS/T-20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karnošová, A.; Strnadová, V.; Holá, L.; Železná, B.; Kuneš, J.; Maletínská, L. Palmitoylation of Prolactin-Releasing Peptide Increased Affinity for and Activation of the GPR10, NPFF-R2 and NPFF-R1 Receptors: In Vitro Study. Int. J. Mol. Sci. 2021, 22, 8904. https://doi.org/10.3390/ijms22168904
Karnošová A, Strnadová V, Holá L, Železná B, Kuneš J, Maletínská L. Palmitoylation of Prolactin-Releasing Peptide Increased Affinity for and Activation of the GPR10, NPFF-R2 and NPFF-R1 Receptors: In Vitro Study. International Journal of Molecular Sciences. 2021; 22(16):8904. https://doi.org/10.3390/ijms22168904
Chicago/Turabian StyleKarnošová, Alena, Veronika Strnadová, Lucie Holá, Blanka Železná, Jaroslav Kuneš, and Lenka Maletínská. 2021. "Palmitoylation of Prolactin-Releasing Peptide Increased Affinity for and Activation of the GPR10, NPFF-R2 and NPFF-R1 Receptors: In Vitro Study" International Journal of Molecular Sciences 22, no. 16: 8904. https://doi.org/10.3390/ijms22168904