
Zebrafish Motile Cilia as a Model for Primary Ciliary Dyskinesia

 , , and
, , and
Abstract
:1. Introduction
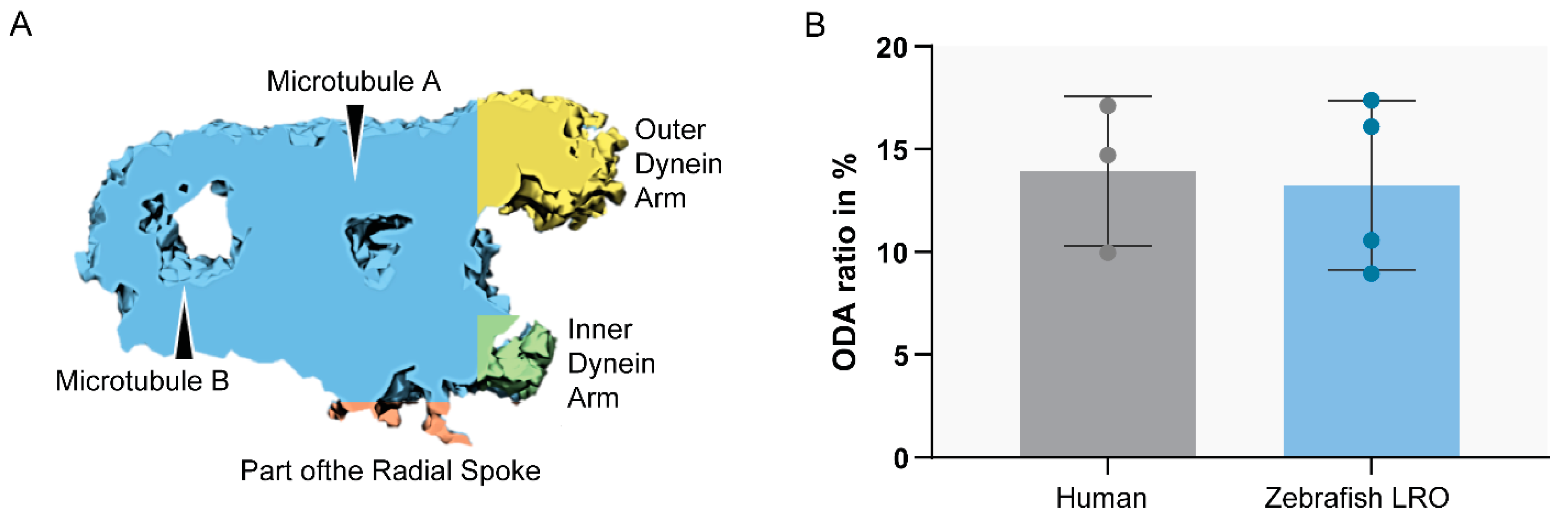
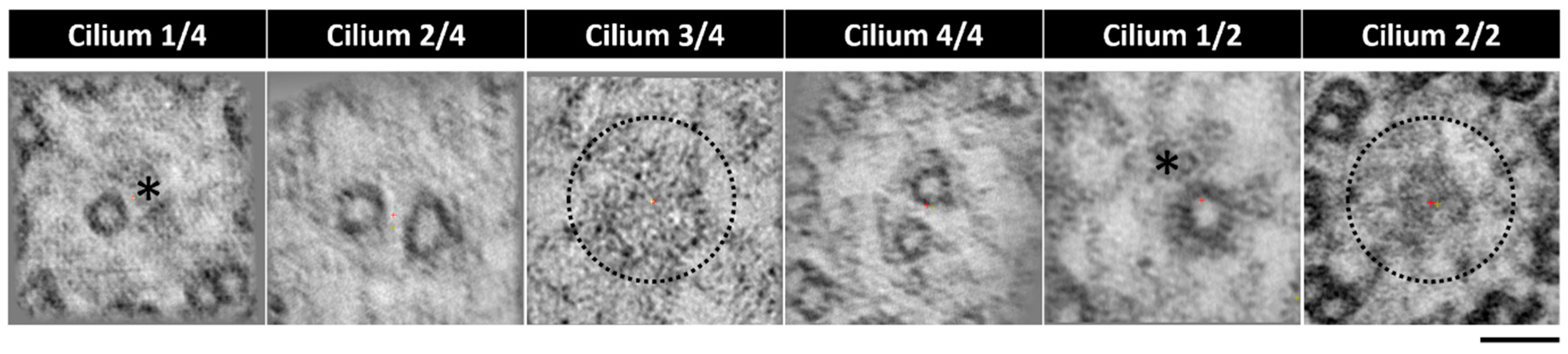
2. Results
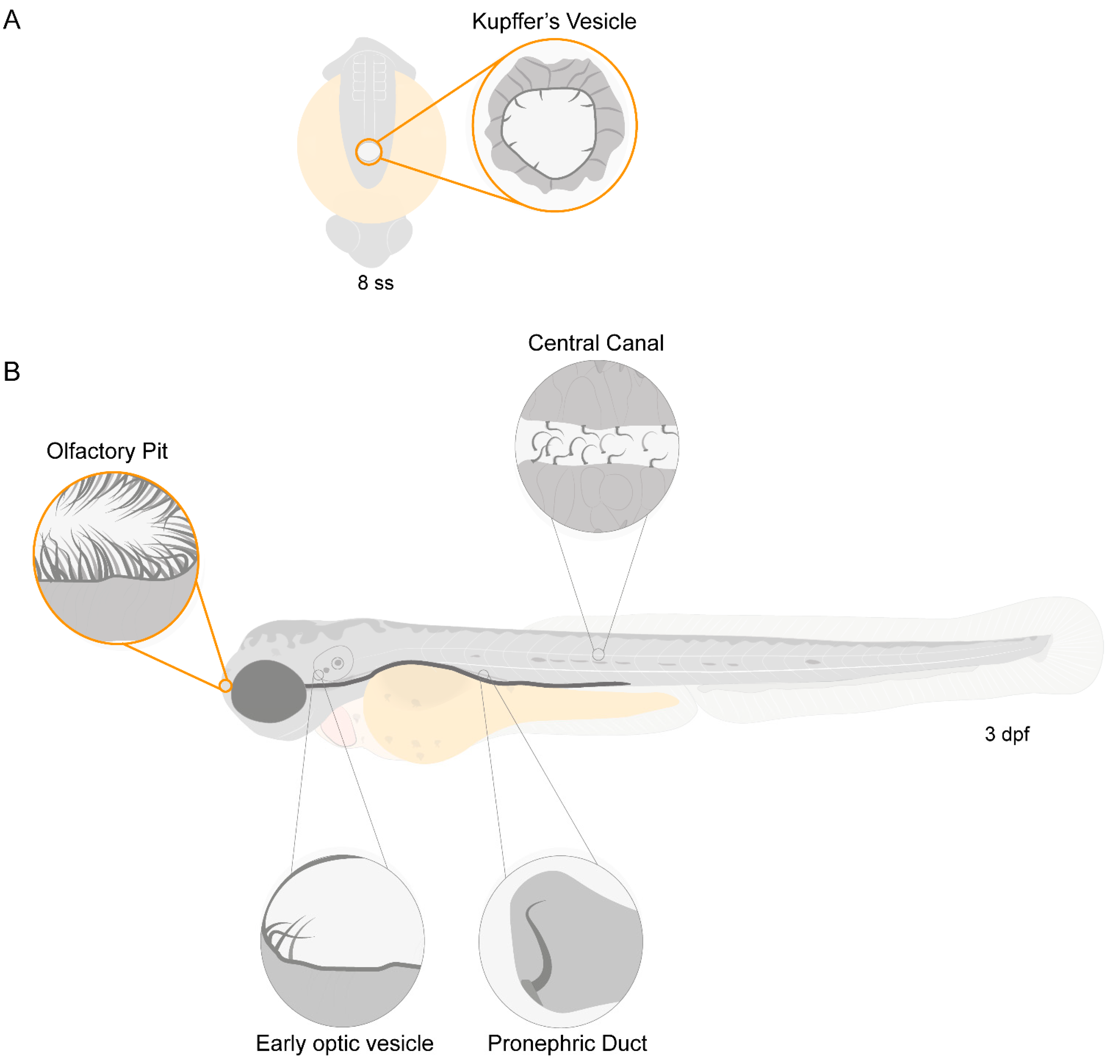
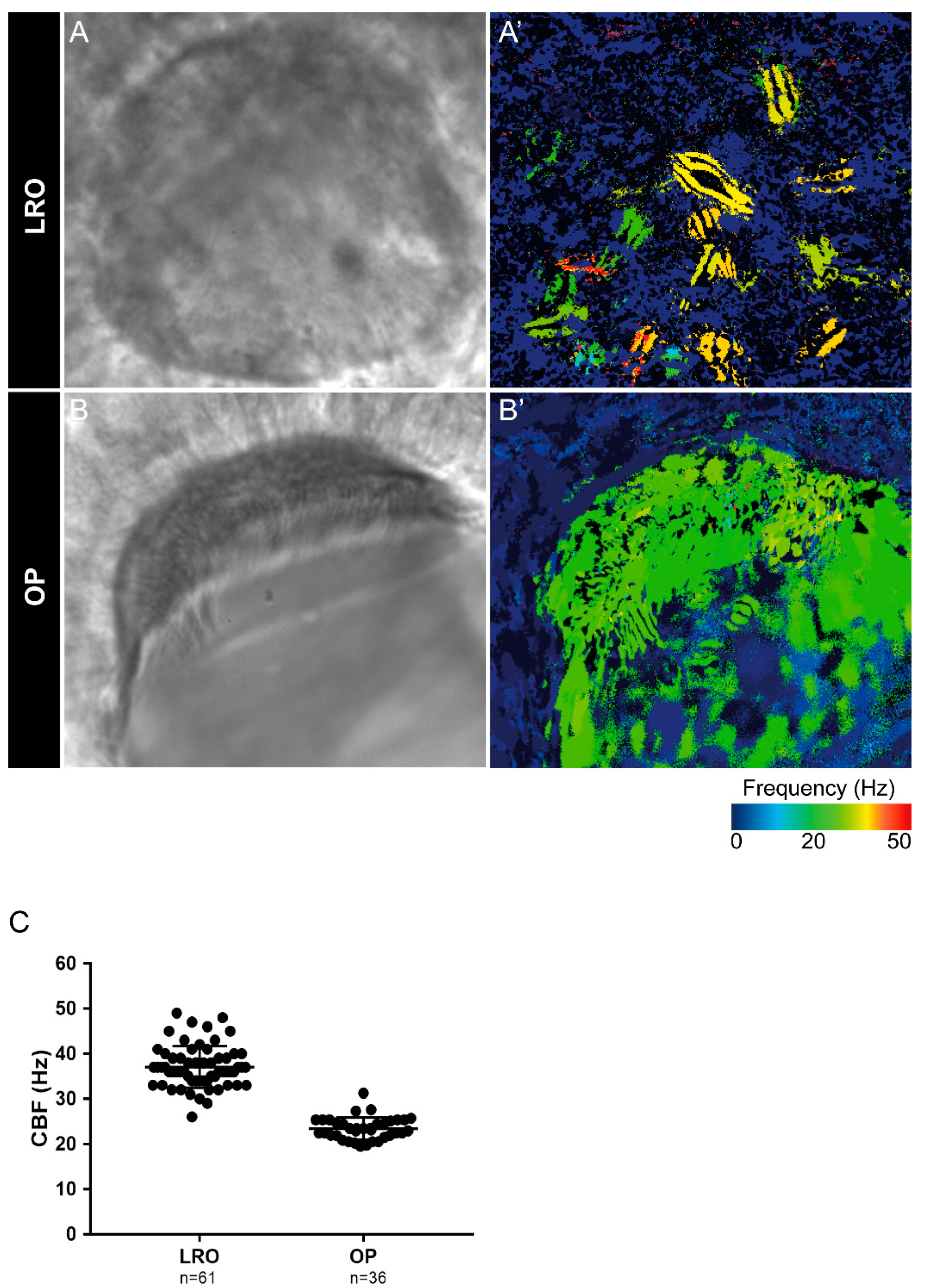
2.1. Zebrafish Motile Cilia
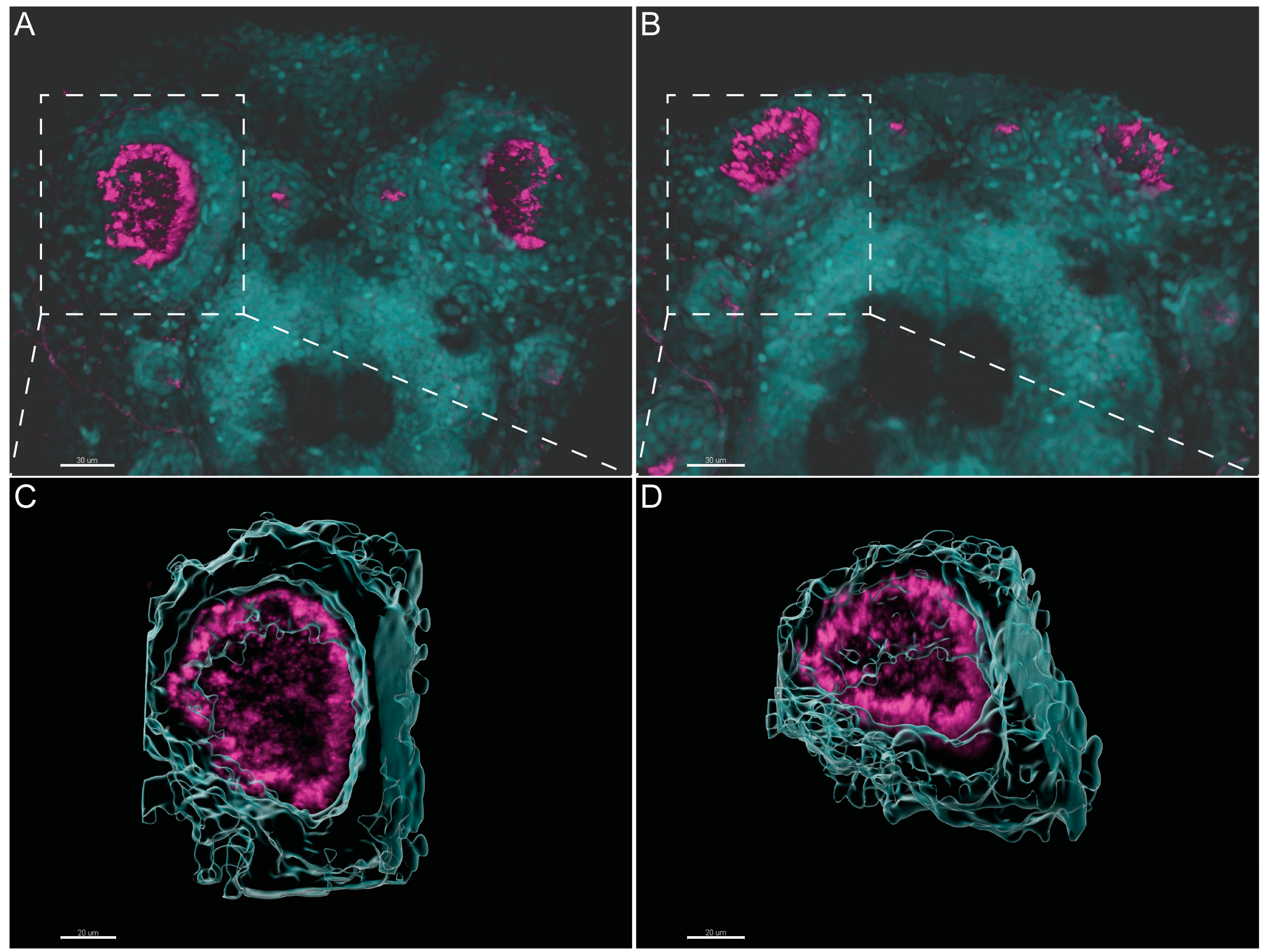
2.2. Different Cilia in the Zebrafish Olfactory Pit
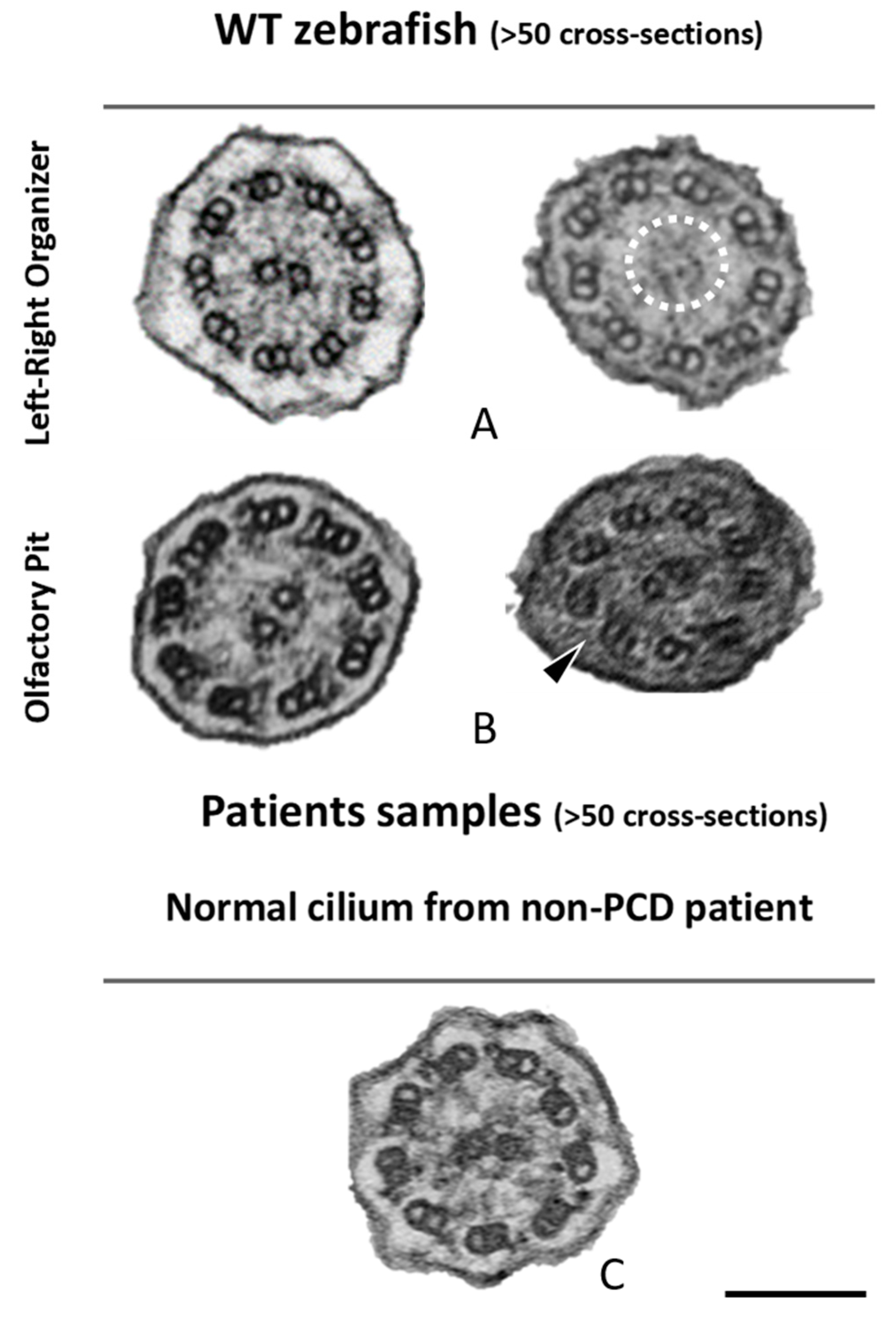
2.3. The Application of Transmission Electron Tomography for the Study of the Central Pair
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Animals
4.3. Immunofluorescence and Image Analysis
4.4. Transmission Electron Microscopy
4.5. Electron Tomography
4.6. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicastro, D.; Schwartz, C.; Pierson, J.; Gaudette, R.; Porter, M.E.; McIntosh, J.R. The molecular architecture of axonemes revealed by cryoelectron tomography. Science 2006, 313, 944–948. [Google Scholar] [CrossRef]
- Ishikawa, T. Axoneme structure from motile cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincensini, L.; Blisnick, T.; Bastin, P. 1001 Model Organisms to Study Cilia and Flagella. Biol. Cell 2011, 103, 109–130. [Google Scholar] [CrossRef]
- Legendre, M.; Zaragosi, L.E.; Mitchison, H.M. Motile cilia and airway disease. In Seminars in Cell & Developmental Biology; Elsevier: London, UK, 2020. [Google Scholar]
- Porter, K.R. The submicroscopic morphology of protoplasm. 1956. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 2005, 287, 1186–1204. [Google Scholar] [PubMed]
- Satir, P.; Christensen, S.T. Overview of Structure and Function of Mammalian Cilia. Annu. Rev. Physiol. 2007, 69, 377–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspary, T.; Larkins, C.E.; Anderson, K.V. The Graded Response to Sonic Hedgehog Depends on Cilia Architecture. Dev. Cell 2007, 12, 767–778. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, S.; Shiratori, H.; Saijoh, Y.; Hamada, H. Determination of left-right patterning of the mouse embryo by artificial nodal flow. Nature 2002, 418, 96–99. [Google Scholar] [CrossRef]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Gluenz, E.; Höög, J.L.; Smith, A.E.; Dawe, H.R.; Shaw, M.K.; Gull, K. Beyond 9 + 0: Noncanonical axoneme structures characterize sensory cilia from protists to humans. FASEB J. 2010, 24, 3117–3121. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, I.; Dynlacht, B.D. Cilium assembly and disassembly. Nat. Cell Biol. 2016, 18, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Lau, D.; Ng, C.P.; Roy, S. Cilia-driven fluid flow as an epigenetic cue for otolith biomineralization on sensory hair cells of the inner ear. Development 2011, 138, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, A.; Zeiske, E. The Peripheral Olfactory Organ of the Zebrafish, Danio Rerio: An Ultrastructural Study; Oxford University Press: Oxford, UK, 1998; Volume 23. [Google Scholar]
- Del Bigio, M.R. Ependymal cells: Biology and pathology. Acta Neuropathol. 2010, 119, 55–73. [Google Scholar] [CrossRef]
- Satir, P.; Sleigh, M.A. The physiology of cilia and mucociliary interactions. Annu. Rev. Physiol. 1990, 52, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, P.; Ferreira, R.R.; Guerrero, A.; Pintado, P.; Tavares, B.; Amaro, J.; Smith, A.A.; Montenegro-Johnson, T.; Smith, D.J.; Lopes, S.S. Article Left-Right Organizer Flow Dynamics: How Much Cilia Activity Reliably Yields Laterality? Dev. Cell 2014, 29, 716–728. [Google Scholar] [CrossRef] [Green Version]
- Roxo-Rosa, M.; Santos Lopes, S. The Zebrafish Kupffer’s Vesicle: A Special Organ in a Model Organism to Study Human Diseases. In Zebrafish in Biomedical Research; American College of Laboratory Animal Medicina series; USA IntechOpen: New York, NY, USA, 2020. [Google Scholar]
- Tavares, B.; Jacinto, R.; Sampaio, P.; Pestana, S.; Pinto, A.; Vaz, A.; Roxo-Rosa, M.; Gardner, R.; Lopes, T.; Schilling, B.; et al. Notch/Her12 signalling modulates, motile/immotile cilia ratio downstream of Foxj1a in zebrafish left-right organizer. Elife 2017, 6, e25165. [Google Scholar] [CrossRef] [PubMed]
- Feistel, K.; Blum, M. Three types of cilia including a novel 9 + 4 axoneme on the notochordal plate of the rabbit embryo. Dev. Dyn. 2006, 235, 3348–3358. [Google Scholar] [CrossRef] [PubMed]
- Yokeid, H.; Ueno, H.; Narita, A.; Sakai, T.; Horiuchiid, K.; Shingyojiid, C.; Hamada, H.; Shinoharaid, K. Rsph4a is essential for the triplet radial spoke head assembly of the mouse motile cilia. PLoS Genet. 2020, 16, e1008664. [Google Scholar]
- Castleman, V.H.; Romio, L.; Chodhari, R.; Hirst, R.A.; de Castro, S.C.P.; Parker, K.A.; Ybot-Gonzalez, P.; Emes, R.D.; Wilson, S.W.; Wallis, C.; et al. Mutations in radial spoke head protein genes RSPH9 and RSPH4A cause primary ciliary dyskinesia with central-microtubular-pair abnormalities. Am. J. Hum. Genet. 2008, 84, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Driever, W.; Solnica-Krezel, L.; Schier, A.F.; Neuhauss, S.C.F.; Malicki, J.; Stemple, D.L.; Stainier, D.Y.R.; Zwartkruis, F.; Abdelilah, S.; Rangini, Z.; et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development 1996, 123, 37–46. [Google Scholar] [CrossRef]
- Liu, G.; Wang, L.; Pan, J.; Yao, X. Chlamydomonas WDR92 in association with R2TP-like complex and multiple DNAAFs to regulate ciliary dynein preassembly. J. Mol. Cell Biol. 2019, 11, 770–780. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Drummond, I.; Austin-Tse, C. Zebrafish cilia. In Methods in Enzymology; Elsevier Group: London, UK, 2013; Volume 525, pp. 219–244. ISBN 9780123979445. [Google Scholar]
- Hansen, A.; Zielinski, B.S. Diversity in the olfactory epithelium of bony fishes: Development, lamellar arrangement, sensory neuron cell types and transduction components. J. Neurocytol. 2006, 208, 183–208. [Google Scholar] [CrossRef] [PubMed]
- Olivares, J.; Schmachtenberg, O. An update on anatomy and function of the teleost olfactory system. PeerJ 2019, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Neuhauss, S.C.F. Olfaction: How Fish Catch a Whiff. Curr. Biol. 2017, 27, R57–R58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, A.; Rolen, S.H.; Anderson, K.; Morita, Y.; Caprio, J.; Finger, T.E. Correlation between Olfactory Receptor Cell Type and Function in the Channel Catfish. J. Neurosci. 2003, 23, 9328–9339. [Google Scholar] [CrossRef]
- Song, Z.; Zhang, X.; Jia, S.; Yelick, P.C.; Zhao, C. Zebrafish as a Model for Human Ciliopathies. J. Genet. Genom. 2016, 43, 107–120. [Google Scholar] [CrossRef]
- Liu, D.; Wang, Z.; Xiao, A.; Zhang, Y.; Li, W.; Zu, Y.; Yao, S.; Lin, S.; Zhang, B. Efficient Gene Targeting in Zebrafish Mediated by a Zebrafish-Codon-Optimized Cas9 and Evaluation of Off-Targeting Effect. J. Genet. Genom. 2014, 41, 43–46. [Google Scholar] [CrossRef]
- Chang, N.; Sun, C.; Gao, L.; Zhu, D.; Xu, X.; Zhu, X.; Xiong, J.W.; Xi, J.J. Genome editing with RNA-guided Cas9 nuclease in Zebrafish embryos. Cell Res. 2013, 23, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Kouis, P.; Yiallouros, P.K.; Middleton, N.; Evans, J.S.; Kyriacou, K.; Papatheodorou, S.I. Prevalence of primary ciliary dyskinesia in consecutive referrals of suspect cases and the transmission electron microscopy detection rate: A systematic review and meta-analysis. Pediatr. Res. 2016, 81, 398–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, J.S.; Burgess, A.; Mitchison, H.M.; Moya, E.; Williamson, M.; Hogg, C. Diagnosis and management of primary ciliary dyskinesia. Arch. Dis. Child. 2014, 99, 850–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchison, H.M.; Valente, E.M. Motile and non-motile cilia in human pathology: From function to phenotypes. J. Pathol. 2017, 241, 294–309. [Google Scholar] [CrossRef]
- Wallmeier, J.; Nielsen, K.G.; Kuehni, C.E.; Lucas, J.S.; Leigh, M.W.; Zariwala, M.A.; Omran, H. Motile ciliopathies. Nat. Rev. Dis. Prim. 2020, 6, 1–29. [Google Scholar] [CrossRef]
- Reiten, I.; Uslu, F.E.; Fore, S.; Pelgrims, R.; Ringers, C.; Diaz Verdugo, C.; Hoffman, M.; Lal, P.; Kawakami, K.; Pekkan, K.; et al. Motile-Cilia-Mediated Flow Improves Sensitivity and Temporal Resolution of Olfactory Computations. Curr. Biol. 2017, 27, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef] [PubMed]
- Olstad, E.W.; Ringers, C.; Hansen, J.N.; Wens, A.; Brandt, C.; Wachten, D.; Yaksi, E.; Jurisch-Yaksi, N. Ciliary Beating Compartmentalizes Cerebrospinal Fluid Flow in the Brain and Regulates Ventricular Development. Curr. Biol. 2019, 29, 229–241.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, A.; Amack, J.D. Cilia in vertebrate left—Right patterning. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, P.; da Silva, M.F.; Vale, I.; Roxo-Rosa, M.; Pinto, A.; Constant, C.; Pereira, L.; Quintão, C.M.; Lopes, S.S. CiliarMove: New software for evaluating ciliary beat frequency helps find novel mutations by a Portuguese multidisciplinary team on primary ciliary dyskinesia. ERJ Open Res. 2021, 7, 00792–02020. [Google Scholar] [CrossRef]
- Pinto, A.L.; Rai, R.K.; Hogg, C.; Burgoyne, T. Ciliary feature counter: A program for the quantitative assessment of cilia to diagnose primary ciliary dyskinesia. Diagnostics 2020, 10, 524. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Montenegro-Johnson, T.D.; Baker, D.I.; Smith, D.J.; Lopes, S.S. Three-dimensional flow in Kupffer’s Vesicle. J. Math. Biol. 2016, 73, 705–725. [Google Scholar] [CrossRef] [Green Version]
- Waters, A.M.; Beales, P.L. Ciliopathies: An expanding disease spectrum. Pediatr. Nephrol. 2011, 26, 1039–1056. [Google Scholar] [CrossRef] [Green Version]
- Oda, T.; Yanagisawa, H.; Kamiya, R.; Kikkawa, M. A molecular ruler determines the repeat length in eukaryotic cilia and flagella. Science 2014, 346, 857–860. [Google Scholar] [CrossRef]
- Nicastro, D.; Fu, X.; Heuser, T.; Tso, A.; Porter, M.E.; Linck, R.W. Cryo-electron tomography reveals conserved features of doublet microtubules in flagella. Proc. Natl. Acad. Sci. USA 2011, 108, E845–E853. [Google Scholar] [CrossRef] [Green Version]
- Austin-Tse, C.; Halbritter, J.; Zariwala, M.A.; Gilberti, R.M.; Gee, H.Y.; Hellman, N.; Pathak, N.; Liu, Y.; Panizzi, J.R.; Patel-King, R.S.; et al. Zebrafish ciliopathy screen plus human mutational analysis identifies C21orf59 and CCDC65 defects as causing primary ciliary dyskinesia. Am. J. Hum. Genet. 2013, 93, 672–686. [Google Scholar] [CrossRef] [Green Version]
- Oura, S.; Kazi, S.; Savolainen, A.; Nozawa, K.; Castañeda, J.; Yu, Z.; Miyata, H.; Matzuk, R.M.; Hansen, J.N.; Wachten, D.; et al. Cfap97d1 is important for flagellar axoneme maintenance and male mouse fertility. PLoS Genet. 2020, 16, e1008954. [Google Scholar] [CrossRef] [PubMed]
- Mangos, S.; Lam, P.Y.; Zhao, A.; Liu, Y.; Mudumana, S.; Vasilyev, A.; Liu, A.; Drummond, I.A. The ADPKD genes pkd1a/b and pkd2 regulate extracellular matrix formation. DMM Dis. Model. Mech. 2010, 3, 354–365. [Google Scholar] [CrossRef] [Green Version]
- Obara, T.; Mangos, S.; Liu, Y.; Zhao, J.; Wiessner, S.; Kramer-Zucker, A.G.; Olale, F.; Schier, A.F.; Drummond, I.A. Polycystin-2 immunolocalization and function in zebrafish. J. Am. Soc. Nephrol. 2006, 17, 2706–2718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bataille, S.; Demoulin, N.; Devuyst, O.; Audrézet, M.P.; Dahan, K.; Godin, M.; Fonts, M.; Pirson, Y.; Burtey, S. Association of PKD2 (Polycystin 2) mutations with left-right laterality defects. Am. J. Kidney Dis. 2011, 58, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Vick, P.; Kreis, J.; Schneider, I.; Tingler, M.; Getwan, M.; Thumberger, T.; Beyer, T.; Schweickert, A.; Blum, M. An Early Function of Polycystin-2 for Left-Right Organizer Induction in Xenopus. iScience 2018, 2, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Rachev, E.; Schuster-Gossler, K.; Fuhl, F.; Ott, T.; Tveriakhina, L.; Beckers, A.; Hegermann, J.; Boldt, K.; Mai, M.; Kremmer, E.; et al. CFAP43 modulates ciliary beating in mouse and Xenopus. Dev. Biol. 2020, 459, 109–125. [Google Scholar] [CrossRef] [Green Version]
- Nicastro, D. Cryo-electron microscope tomography to study axonemal organization. Methods Cell Biol. 2009, 91, 1–39. [Google Scholar] [PubMed]
- Essner, J.J.; Amack, J.D.; Nyholm, M.K.; Harris, E.B.; Yost, H.J. Kupffer’s vesicle is a ciliated organ of asymmetry in the zebrafish embryo that initiates left-right development of the brain, heart and gut. Development 2005, 132, 1247–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoemark, A.; Burgoyne, T.; Kwan, R.; Dixon, M.; Patel, M.P.; Rogers, A.V.; Onoufriadis, A.; Scully, J.; Daudvohra, F.; Cullup, T.; et al. Primary ciliary dyskinesia with normal ultrastructure: Three-dimensional tomography detects absence of DNAH11. Eur. Respir. J. 2018, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Panizzi, J.R.; Becker-Heck, A.; Castleman, V.H.; Al-Mutairi, D.A.; Liu, Y.; Loges, N.T.; Pathak, N.; Austin-Tse, C.; Sheridan, E.; Schmidts, M.; et al. CCDC103 mutations cause primary ciliary dyskinesia by disrupting assembly of ciliary dynein arms. Nat. Genet. 2012, 44, 714–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Computer Visualization of Three-Dimensional Image Data Using IMOD. Available online: https://bio3d.colorado.edu/imod/paper/ (accessed on 13 March 2021).
- UCSF Chimera Home Page. Available online: https://www.rbvi.ucsf.edu/chimera/ (accessed on 13 March 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCD Gene | Zebrafish Transcript Name | Zebrafish Transcript ID |
|---|---|---|
| DNAH5 | dnah5-201 | ENSDART00000123150.4 |
| dnah5-202 | ENSDART00000191818.1 | |
| CCDC114 (ODAD1) | ccdc114-201 | ENSDART00000023745.8 |
| ARMC4 (ODAD2) | cr847789.1-201 | ENSDART00000186851.1 |
| armc4-201 | ENSDART00000077453.5 | |
| armc4-204 | ENSDART00000170018.2 | |
| armc4-203 | ENSDART00000153115.2 | |
| armc4-202 | ENSDART00000152887.2 | |
| TTC25 (ODAD4) | ttc25-201 | ENSDART00000080946.5 |
| DNAH9 | dnah9-201 | ENSDART00000160926.2 |
| DNAH11 | dnah11-201 | ENSDART00000148294.4 |
| dnah11-202 | ENSDART00000020821.10 | |
| dnah11-203 | ENSDART00000138744.4 | |
| DNAI1 | dnai1.2-201 | ENSDART00000080431.5 |
| dnai1.2-202 | ENSDART00000142468.3 | |
| dnai1.1-205 | ENSDART00000170205.2 | |
| dnai1.1-201 | ENSDART00000160163.2 | |
| dnai1.1-204 | ENSDART00000169676.2 | |
| dnai1.1-202 | ENSDART00000163063.2 | |
| dnai1.1-203 | ENSDART00000165798.2 | |
| DNAI2 | dnai2a-201 | ENSDART00000162579.2 |
| dnai2a-202 | ENSDART00000164199.2 | |
| dnai2b-203 | ENSDART00000188726.1 | |
| dnai2b-201 | ENSDART00000003339.9 | |
| dnai2b-202 | ENSDART00000188648.1 | |
| DNAL1 | dnal1-203 | ENSDART00000188500.1 |
| dnal1-202 | ENSDART00000156182.2 | |
| dnal1-201 | ENSDART00000043651.7 | |
| TXNDC3 (NME8) | nme8-201 | ENSDART00000163684.2 |
| CCDC103 | ccdc103-201 | ENSDART00000075493.4 |
| ccdc103-202 | ENSDART00000132293.2 | |
| CFAP298 (C21orf59) | cfap298-201 | ENSDART00000051197.6 |
| cfap298-202 | ENSDART00000130093.3 | |
| cfap298-203 | ENSDART00000181950.1 | |
| CFAP300 (c11orf70) | cfap300-201 | ENSDART00000151109.2 |
| cfap300-202 | ENSDART00000192737.1 | |
| DNAAF1 (LRRC50) | dnaaf1-201 | ENSDART00000145762.4 |
| dnaaf1-203 | ENSDART00000173909.2 | |
| dnaaf1-202 | ENSDART00000173853.2 | |
| DNAAF2 (KTU) | dnaaf2-201 | ENSDART00000167840.2 |
| DNAAF3 | dnaaf3l-201 | ENSDART00000079233.5 |
| DNAAF4 (DYX1C1) | dnaaf4-201 | ENSDART00000165855.2 |
| DNAAF5 (HEATR2) | lo018183.1-201 | ENSDART00000194031.1 |
| DNAAF6 (PIH1D3) | pih1d3-201 | ENSDART00000056375.5 |
| pih1d3-203 | ENSDART00000145388.3 | |
| pih1d3-202 | ENSDART00000136858.2 | |
| pih1d3-204 | ENSDART00000183524.1 | |
| pih1d3-205 | ENSDART00000191761.1 | |
| LRRC6 | lrrc6-203 | ENSDART00000188883.1 |
| lrrc6-202 | ENSDART00000132346.3 | |
| lrrc6-201 | ENSDART00000075347.5 | |
| RPGR | rpgrb-201 | ENSDART00000088624.5 |
| rpgrb-202 | ENSDART00000124471.3 | |
| rpgrip-201 | ENSDART00000138541.3 | |
| rpgrip-203 | ENSDART00000190953.1 | |
| rpgrip-202 | ENSDART00000179003.2 | |
| rpgrip1l-202 | ENSDART00000185324.1 | |
| rpgrip1l-201 | ENSDART00000126326.5 | |
| SPAG1 | spag1b-201 | ENSDART00000101207.5 |
| spag1a-202 | ENSDART00000185960.1 | |
| spag1a-201 | ENSDART00000130537.3 | |
| ZMYND10 | zmynd10-201 | ENSDART00000017413.10 |
| zmynd10-202 | ENSDART00000189261.1 | |
| zmynd10-203 | ENSDART00000183251.1 | |
| CCDC39 | ccdc39-202 | ENSDART00000190769.1 |
| ccdc39-201 | ENSDART00000169709.2 | |
| CCDC40 | ccdc40-202 | ENSDART00000169752.2 |
| Ccdc40-201 | ENSDART00000164275.2 | |
| Ccdc40-203 | ENSDART00000182267.1 | |
| TTC12 | ttc12-201 | ENSDART00000156234.2 |
| ttc12-202 | ENSDART00000157380.2 | |
| CCDC65 (DRC2) | ccdc65-201 | ENSDART00000043946.8 |
| ccdc65-202 | ENSDART00000177219.2 | |
| CCDC164 (DRC1) | drc1-201 | ENSDART00000061829.5 |
| GAS8 | gas8-202 | ENSDART00000170982.2 |
| gas8-201 | ENSDART00000165126.2 | |
| CFAP221 | not found in ZF | |
| DNAJB13 | dnajb13-204 | ENSDART00000148093.3 |
| dnajb13-201 | ENSDART00000063365.6 | |
| dnajb13-203 | ENSDART00000139097.2 | |
| dnajb13-202 | ENSDART00000133505.2 | |
| HYDIN | hydin-201 | ENSDART00000143265.4 |
| hydin-202 | ENSDART00000145701.2 | |
| hydin-203 | ENSDART00000169861.2 | |
| bx571975.1-201 | ENSDART00000185269.1 | |
| NME5 | nme5-201 | ENSDART00000060998.6 |
| RSPH1 | rsph1-201 | ENSDART00000160273.3 |
| ct573248.3-201 | ENSDART00000181186.1 | |
| RSPH3 | rsph3-202 | ENSDART00000128823.5 |
| rsph3-201 | ENSDART00000103394.3 | |
| RSPH4a | rsph4a-201 | ENSDART00000097340.5 |
| RSPH9 | rsph9-201 | ENSDART00000010903.8 |
| STK36 | stk36-201 | ENSDART00000086765.5 |
| stk36-202 | ENSDART00000139065.2 | |
| SPEF2 | spef2-201 | ENSDART00000159718.2 |
| spef2-202 | ENSDART00000168984.2 | |
| CFAP57 | cfap57-201 | ENSDART00000080900.6 |
| cfap57-202 | ENSDART00000149309.3 | |
| LRRC56 | lrrc56-202 | ENSDART00000161369.2 |
| lrrc56-201 | ENSDART00000150364.2 | |
| GAS2L2 | gas2l2-201 | ENSDART00000112744.4 |
| NEK10 | nek10-201 | ENSDART00000155162.2 |
| OFD1 | ofd1-201 | ENSDART00000000552.12 |
| CCNO | fq311924.1-201 | ENSDART00000158096.2 |
| FOXJ1 | foxj1b-201 | ENSDART00000126676.2 |
| foxj1b-203 | ENSDART00000181942.1 | |
| foxj1b-202 | ENSDART00000153327.2 | |
| foxj1a-201 | ENSDART00000157772.2 | |
| foxj1a-202 | ENSDART00000168280.2 | |
| MCIDAS | cu633857.1-201 | ENSDART00000192716.1 |
| Disarranged Cilia (%) | Dynein Arms Assessment (%) | ||||
|---|---|---|---|---|---|
| Counted > 50 Cilia | Both Arms Present | ODA Missing | IDA Missing | Both Arms Missing | |
| WT zebrafish (n = 3) | 14 (±8) | 62 (±1) | 23 (±2) | 14 (±2) | 14 (±2) |
| Human control (n = 3) | 3 (±1) | 93 (±12) | 1 (±1) | 3 (±6) | 2 (±4) |
| LROs | OP and Nasal Cilia | |
|---|---|---|
| Zebrafish |
|
|
| Human |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto, A.L.; Rasteiro, M.; Bota, C.; Pestana, S.; Sampaio, P.; Hogg, C.; Burgoyne, T.; Lopes, S.S. Zebrafish Motile Cilia as a Model for Primary Ciliary Dyskinesia. Int. J. Mol. Sci. 2021, 22, 8361. https://doi.org/10.3390/ijms22168361
Pinto AL, Rasteiro M, Bota C, Pestana S, Sampaio P, Hogg C, Burgoyne T, Lopes SS. Zebrafish Motile Cilia as a Model for Primary Ciliary Dyskinesia. International Journal of Molecular Sciences. 2021; 22(16):8361. https://doi.org/10.3390/ijms22168361
Chicago/Turabian StylePinto, Andreia L., Margarida Rasteiro, Catarina Bota, Sara Pestana, Pedro Sampaio, Claire Hogg, Thomas Burgoyne, and Susana S. Lopes. 2021. "Zebrafish Motile Cilia as a Model for Primary Ciliary Dyskinesia" International Journal of Molecular Sciences 22, no. 16: 8361. https://doi.org/10.3390/ijms22168361