
Differential Serotonin Uptake Mechanisms at the Human Maternal–Fetal Interface

 , , , , , and
, , , , , and 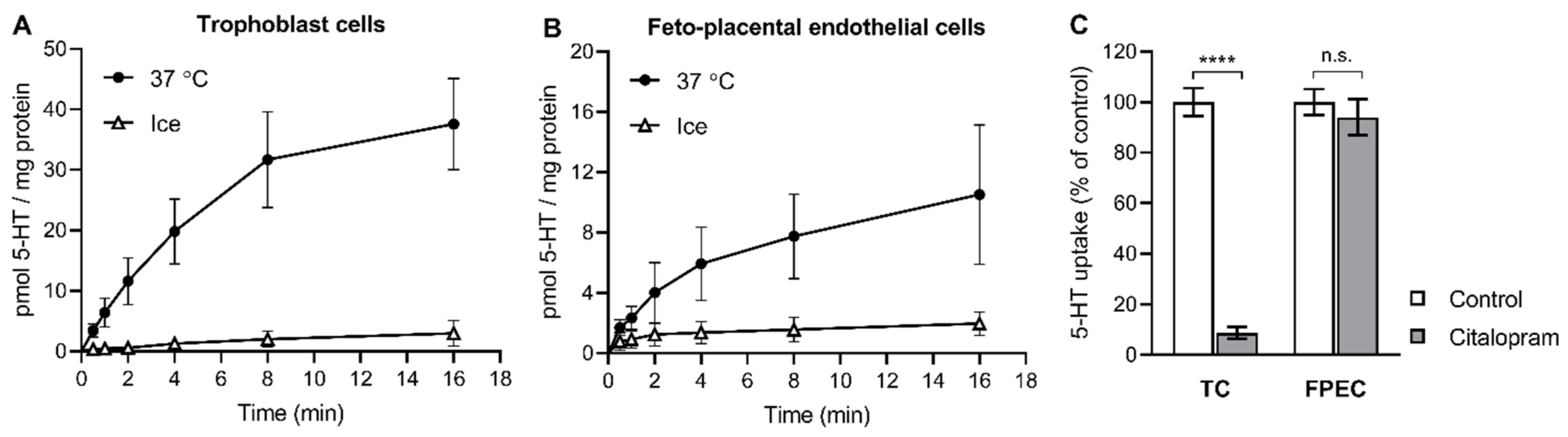{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
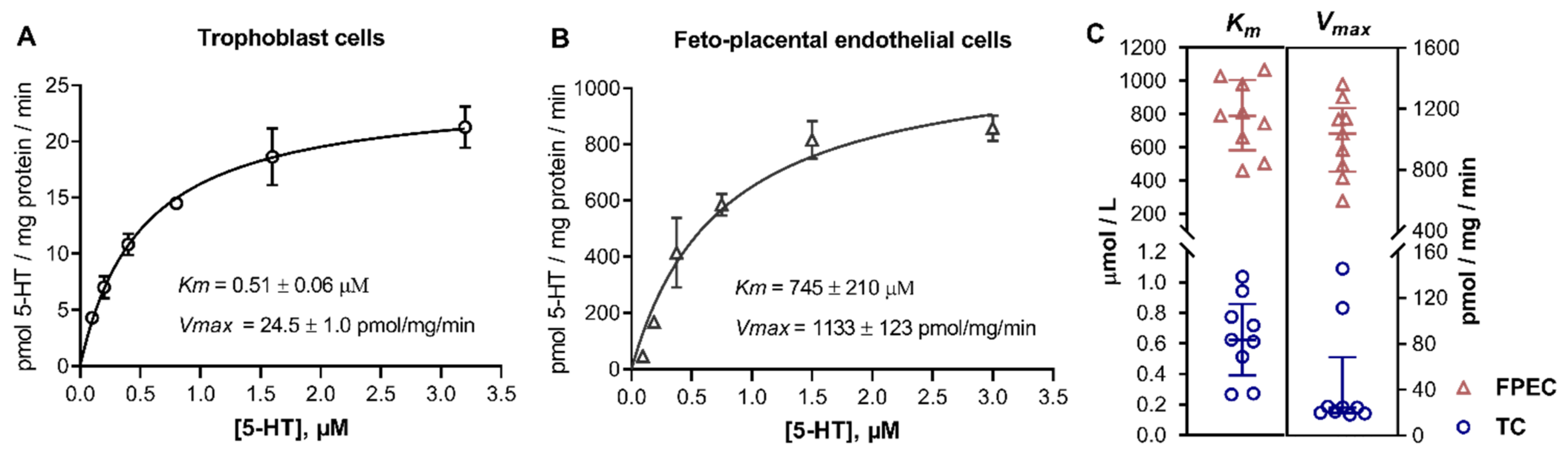
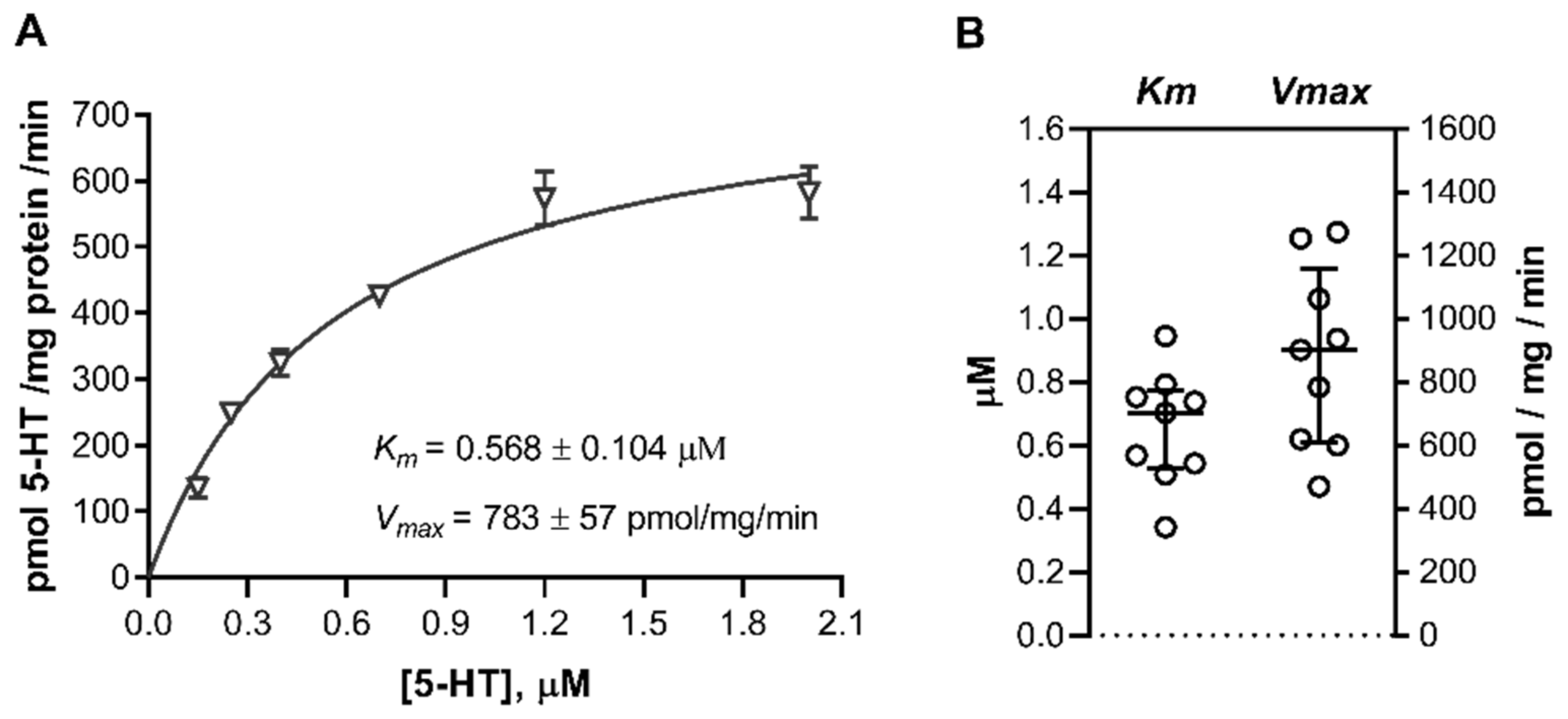
2.1. 5-HT Uptake into Primary Placental Cells
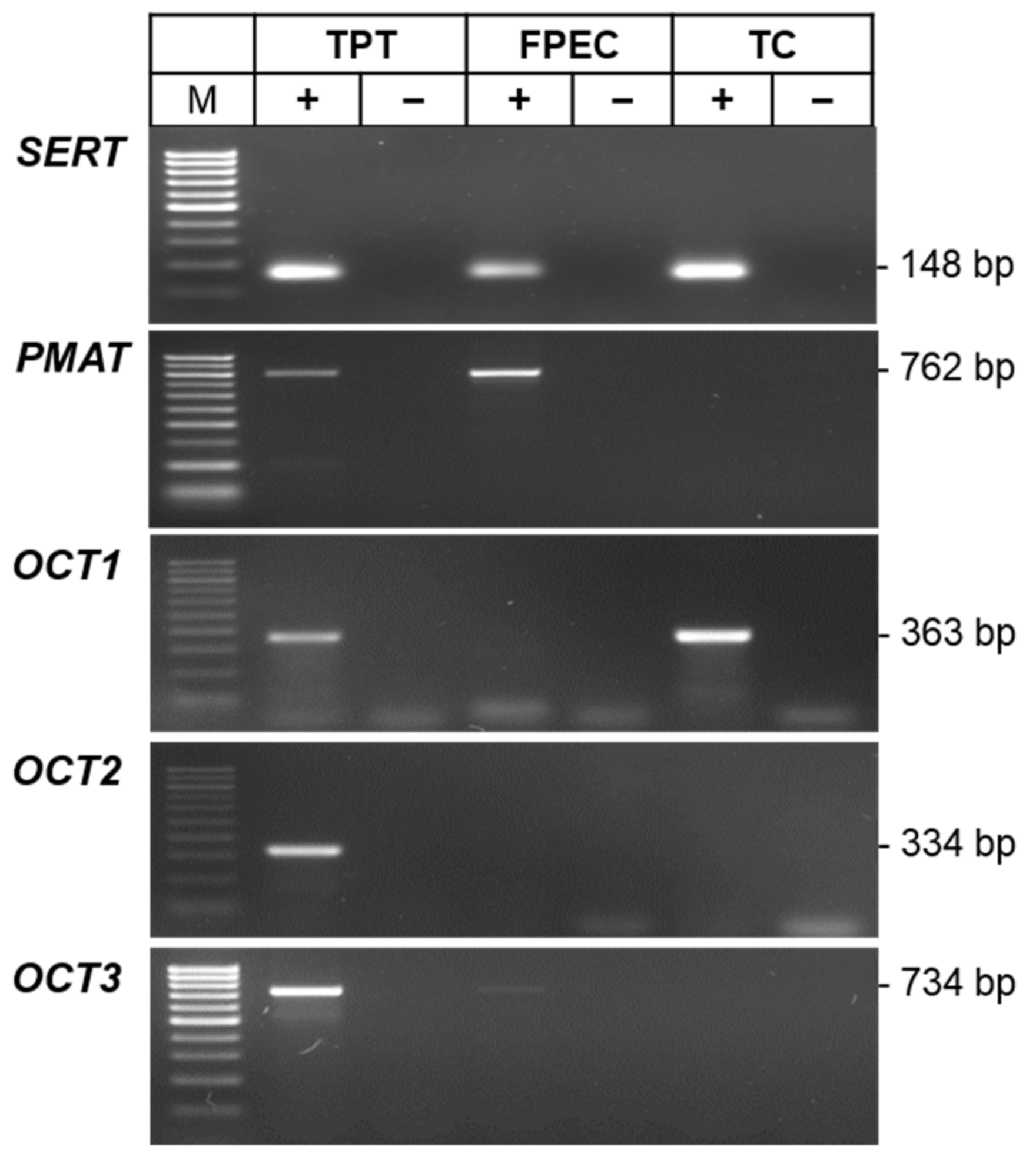
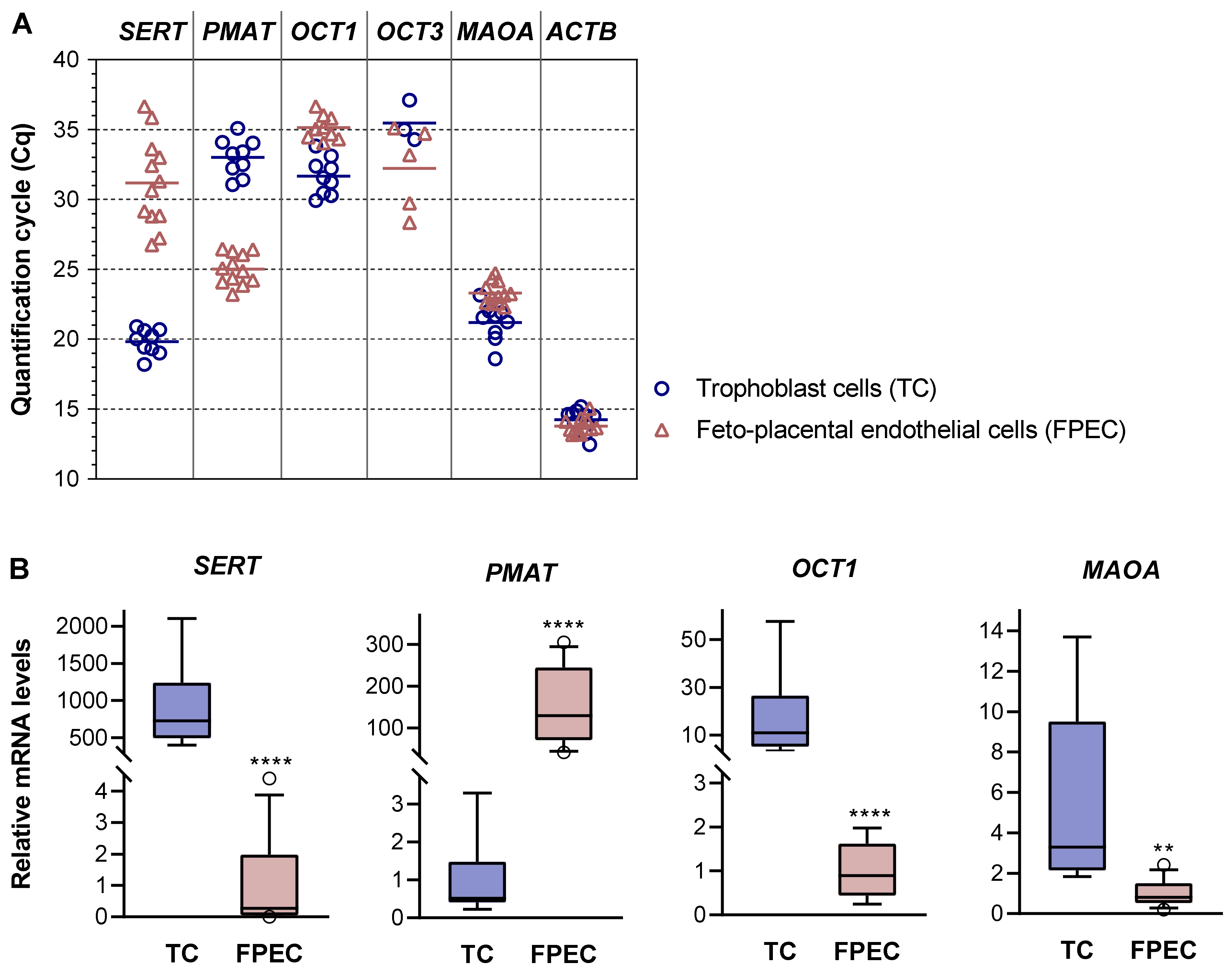
2.2. Expression of 5-HT-Regulating Genes in Primary Placental Cells
2.3. Uptake of 5HT in Cord Blood Platelets
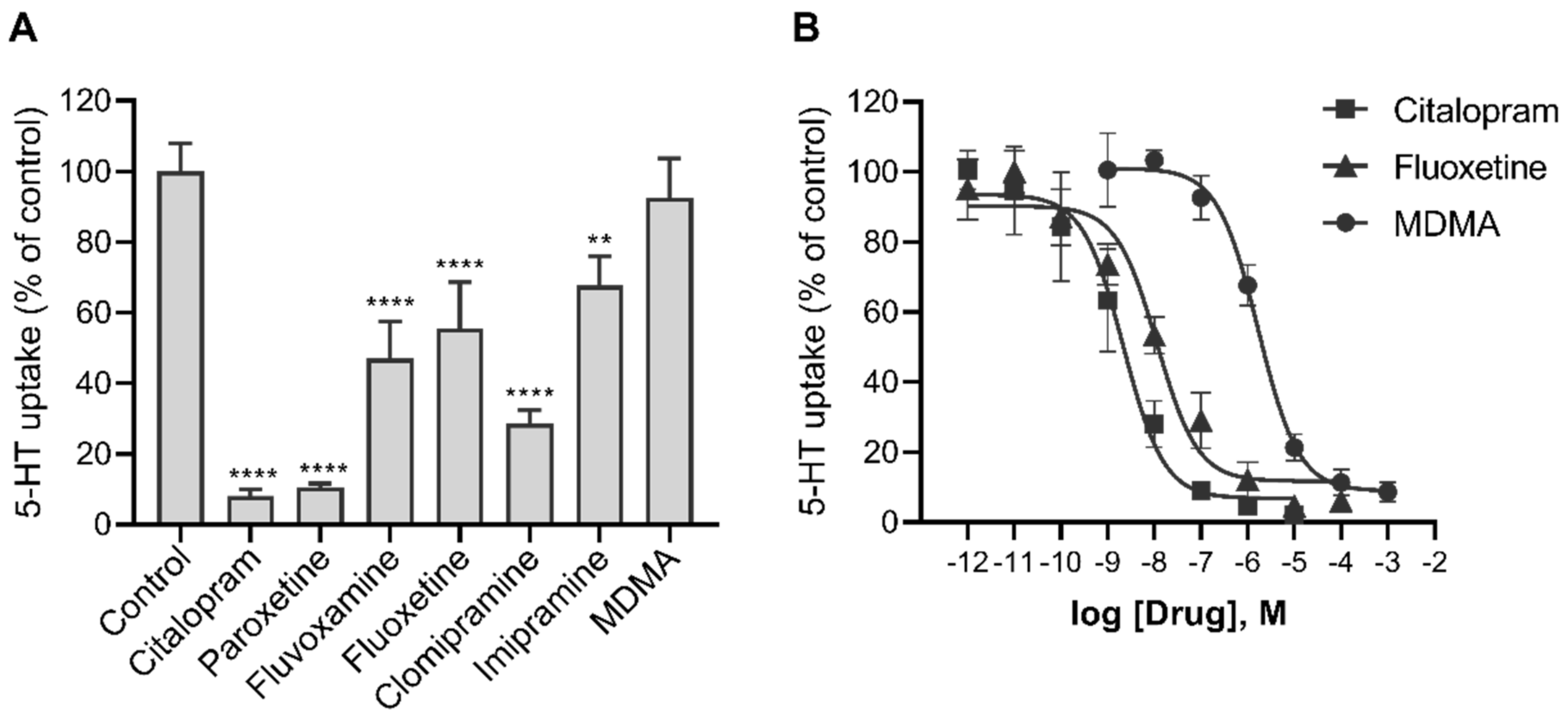
2.4. Effects of Various Psychotropic Drugs on the Uptake of 5-HT in Human Primary Trophoblasts
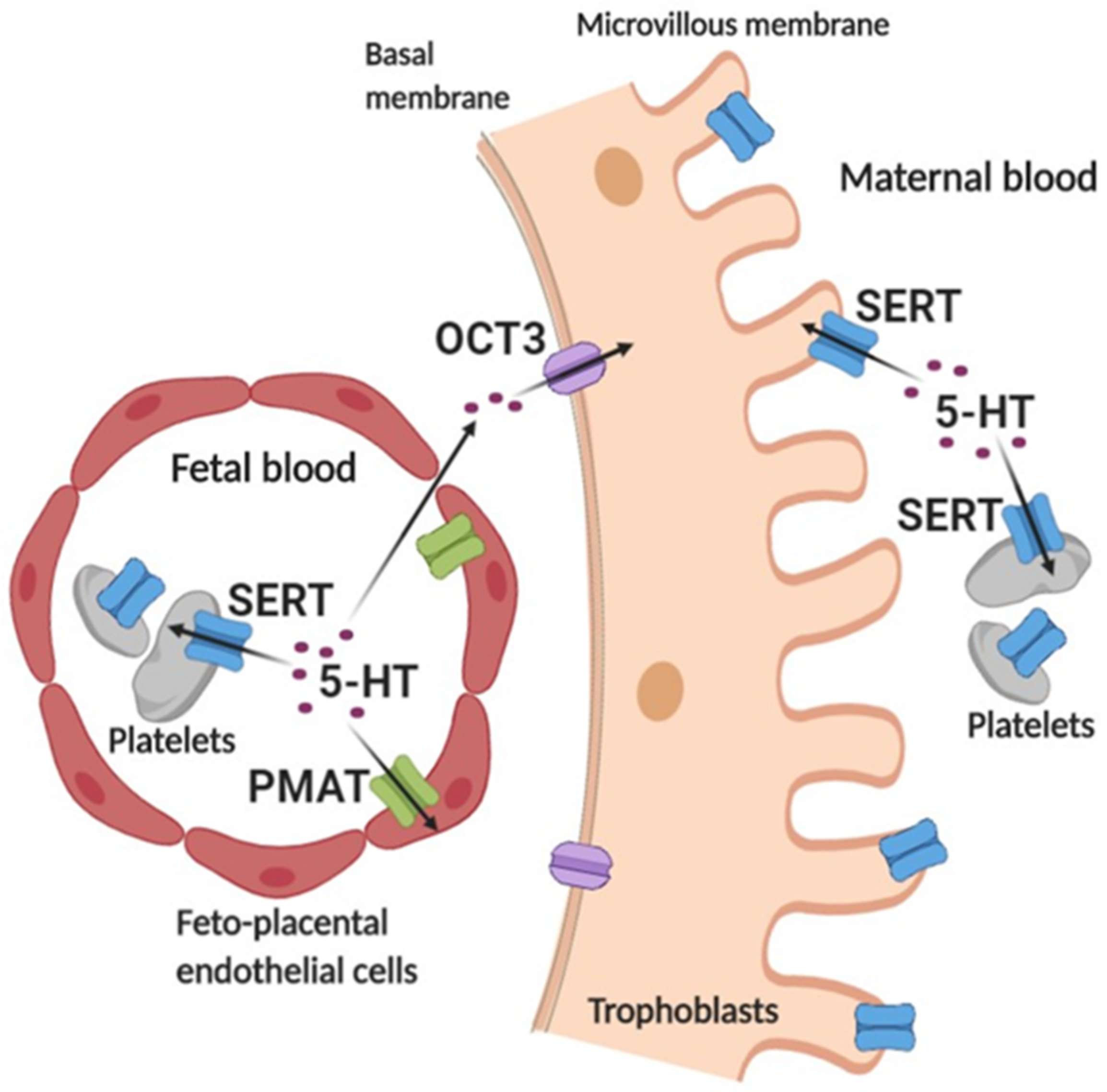
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation and Culture of Human Primary Placental Cells
4.3. 5-HT Uptake in Human Primary Placental Cells
4.4. Pharmacological Studies
4.5. Gene Expression Analyses
4.6. Isolation of Platelet Rich Plasma from Cord Blood
4.7. 5-HT Uptake in Cord Blood Platelets
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Spohn, S.N.; Mawe, G.M. Non-conventional features of peripheral serotonin signalling-the gut and beyond. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 412–420. [Google Scholar] [CrossRef]
- Gaspar, P.; Cases, O.; Maroteaux, L. The developmental role of serotonin: News from mouse molecular genetics. Nat. Rev. Neurosci. 2003, 4, 1002–1012. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Placental serotonin signaling, pregnancy outcomes, and regulation of fetal brain development. Biol. Reprod. 2020, 102, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Sonier, B.; Lavigne, C.; Arseneault, M.; Ouellette, R.; Vaillancourt, C. Expression of the 5-HT2A serotoninergic receptor in human placenta and choriocarcinoma cells: Mitogenic implications of serotonin. Placenta 2005, 26, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Oufkir, T.; Arseneault, M.; Sanderson, J.T.; Vaillancourt, C. The 5-HT2A serotonin receptor enhances cell viability, affects cell cycle progression and activates MEK-ERK1/2 and JAK2-STAT3 signalling pathways in human choriocarcinoma cell lines. Placenta 2010, 31, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Oufkir, T.; Vaillancourt, C. Phosphorylation of JAK2 by serotonin 5-HT(2A) receptor activates both STAT3 and ERK1/2 pathways and increases growth of JEG-3 human placental choriocarcinoma cell. Placenta 2011, 32, 1033–1040. [Google Scholar] [CrossRef]
- Klempan, T.; Hudon-Thibeault, A.A.; Oufkir, T.; Vaillancourt, C.; Sanderson, J.T. Stimulation of serotonergic 5-HT2A receptor signaling increases placental aromatase (CYP19) activity and expression in BeWo and JEG-3 human choriocarcinoma cells. Placenta 2011, 32, 651–656. [Google Scholar] [CrossRef]
- Hadden, C.; Fahmi, T.; Cooper, A.; Savenka, A.V.; Lupashin, V.V.; Roberts, D.J.; Maroteaux, L.; Hauguel-de Mouzon, S.; Kilic, F. Serotonin transporter protects the placental cells against apoptosis in caspase 3-independent pathway. J. Cell. Physiol. 2017, 232, 3520–3529. [Google Scholar] [CrossRef]
- Cruz, M.A.; Gallardo, V.; Miguel, P.; Carrasco, G.; González, C. Serotonin-induced vasoconstriction is mediated by thromboxane release and action in the human fetal-placental circulation. Placenta 1997, 18, 197–204. [Google Scholar] [CrossRef]
- Gonzalez, C.; Cruz, M.A.; Gallardo, V.; Albornoz, J.; Bravo, I. Serotonin-induced vasoconstriction in human placental chorionic veins: Interaction with prostaglandin F2 alpha. Gynecol. Obstet. Investig. 1993, 35, 86–90. [Google Scholar]
- Côté, F.; Fligny, C.; Bayard, E.; Launay, J.M.; Gershon, M.D.; Mallet, J.; Vodjdani, G. Maternal serotonin is crucial for murine embryonic development. Proc. Natl. Acad. Sci. USA 2007, 104, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Kliman, H.J.; Quaratella, S.B.; Setaro, A.C.; Siegman, E.C.; Subha, Z.T.; Tal, R.; Milano, K.M.; Steck, T.L. Pathway of maternal serotonin to the human embryo and fetus. Endocrinology 2018, 159, 1609–1629. [Google Scholar] [CrossRef]
- Carrasco, G.; Cruz, M.A.; Gallardo, V.; Miguel, P.; Dominguez, A.; González, C. Transport and metabolism of serotonin in the human placenta from normal and severely pre-eclamptic pregnancies. Gynecol. Obstet. Investig. 2000, 49, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Ranzil, S.; Ellery, S.; Walker, D.W.; Vaillancourt, C.; Alfaidy, N.; Bonnin, A.; Borg, A.; Wallace, E.M.; Ebeling, P.R.; Erwich, J.J.; et al. Disrupted placental serotonin synthetic pathway and increased placental serotonin: Potential implications in the pathogenesis of human fetal growth restriction. Placenta 2019, 84, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Murthi, P.; Vaillancourt, C. RETRACTED: Placental serotonin systems in pregnancy metabolic complications associated with maternal obesity and gestational diabetes mellitus. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165391. [Google Scholar] [CrossRef]
- Bonnin, A.; Levitt, P. Fetal, maternal, and placental sources of serotonin and new implications for developmental programming of the brain. Neuroscience 2011, 197, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sato, K. Placenta-derived hypo-serotonin situations in the developing forebrain cause autism. Med. Hypotheses 2013, 80, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Räikkönen, K.; Pesonen, A.K.; O’Reilly, J.R.; Tuovinen, S.; Lahti, M.; Kajantie, E.; Villa, P.; Laivuori, H.; Hämäläinen, E.; Seckl, J.R.; et al. Maternal depressive symptoms during pregnancy, placental expression of genes regulating glucocorticoid and serotonin function and infant regulatory behaviors. Psychol. Med. 2015, 45, 3217–3226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goeden, N.; Velasquez, J.; Arnold, K.A.; Chan, Y.; Lund, B.T.; Anderson, G.M.; Bonnin, A. Maternal inflammation disrupts fetal neurodevelopment via increased placental output of serotonin to the fetal brain. J. Neurosci. 2016, 36, 6041–6049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanswijk, S.I.; Spoelder, M.; Shan, L.; Verheij, M.M.M.; Muilwijk, O.G.; Li, W.; Liu, C.; Kolk, S.M.; Homberg, J.R. Gestational factors throughout fetal neurodevelopment: The serotonin link. Int. J. Mol. Sci. 2020, 21, 5850. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, D.; Hannon, J.P.; Martin, G.R. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol. Biochem. Behav. 2002, 71, 533–554. [Google Scholar] [CrossRef]
- Muma, N.A.; Mi, Z. Serotonylation and transamidation of other monoamines. ACS Chem. Neurosci. 2015, 6, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Farrelly, L.A.; Thompson, R.E.; Zhao, S.; Lepack, A.E.; Lyu, Y.; Bhanu, N.V.; Zhang, B.; Loh, Y.H.E.; Ramakrishnan, A.; Vadodaria, K.C.; et al. Histone serotonylation is a permissive modification that enhances TFIID binding to H3K4me3. Nature 2019, 567, 535–539. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Bauman, A.L.; Moore, K.R.; Han, H.; Yang-Feng, T.; Chang, A.S.; Ganapathy, V.; Blakely, R.D. Antidepressant- and cocaine-sensitive human serotonin transporter: Molecular cloning, expression, and chromosomal localization. Proc. Natl. Acad. Sci. USA 1993, 90, 2542–2546. [Google Scholar] [CrossRef] [Green Version]
- Balkovetz, D.F.; Tiruppathi, C.; Leibach, F.H.; Mahesh, V.B.; Ganapathy, V. Evidence for an imipramine-sensitive serotonin transporter in human placental brush-border membranes. J. Biol. Chem. 1989, 264, 2195–2198. [Google Scholar] [CrossRef]
- Mercado, C.P.; Kilic, F. Molecular mechanisms of SERT in platelets: Regulation of plasma serotonin levels. Mol. Interv. 2010, 10, 231–241. [Google Scholar] [CrossRef]
- Koepsell, H. Organic cation transporters in health and disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Lee, N.; Hebert, M.F.; Prasad, B.; Easterling, T.R.; Kelly, E.J.; Unadkat, J.D.; Wang, J. Effect of gestational age on mRNA and protein expression of polyspecific organic cation transporters during pregnancy. Drug Metab. Dispos. 2013, 41, 2225–2232. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, N.; Furugen, A.; Ono, K.; Koishikawa, M.; Miyazawa, Y.; Nishimura, A.; Umazume, T.; Narumi, K.; Kobayashi, M.; Iseki, K. Cellular uptake properties of lamotrigine in human placental cell lines: Investigation of involvement of organic cation transporters (SLC22A1–5). Drug Metab. Pharmacokinet. 2020, 35, 266–273. [Google Scholar] [CrossRef]
- Cool, D.R.; Leibach, F.H.; Bhalla, V.K.; Mahesh, V.B.; Ganapathy, V. Expression and cyclic AMP-dependent regulation of a high affinity serotonin transporter in the human placental choriocarcinoma cell line (JAR). J. Biol. Chem. 1991, 266, 15750–15757. [Google Scholar] [CrossRef]
- Karahoda, R.; Horackova, H.; Kastner, P.; Matthios, A.; Cerveny, L.; Kucera, R.; Kacerovsky, M.; Duintjer Tebbens, J.; Bonnin, A.; Abad, C.; et al. Serotonin homeostasis in the materno-foetal interface at term: Role of transporters (SERT/SLC6A4 and OCT3/SLC22A3) and monoamine oxidase A (MAO-A) in uptake and degradation of serotonin by human and rat term placenta. Acta Physiol. 2020, 229, e13478. [Google Scholar] [CrossRef] [PubMed]
- Hyttel, J. Pharmacological characterization of selective serotonin reuptake inhibitors (SSRIs). Int. Clin. Psychopharmacol. 1994, 9, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Wang, J. Selective transport of monoamine neurotransmitters by human plasma membrane monoamine transporter and organic cation transporter 3. J. Pharmacol. Exp. Ther. 2010, 335, 743–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.W.S.; Yang, C.; Kwan, Y.W.; Chan, S.W.; Lee, S.M.Y.; Leung, G.P.H. Involvement of organic cation transporter-3 and plasma membrane monoamine transporter in serotonin uptake in human brain vascular smooth muscle cells. Front. Pharmacol. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naganuma, F.; Yoshikawa, T.; Nakamura, T.; Iida, T.; Harada, R.; Mohsen, A.S.; Miura, Y.; Yanai, K. Predominant role of plasma membrane monoamine transporters in monoamine transport in 1321N1, a human astrocytoma-derived cell line. J. Neurochem. 2014, 129, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J. The plasma membrane monoamine transporter (PMAT): Structure, function, and role in organic cation disposition. Clin. Pharmacol. Ther. 2016, 100, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Boxberger, K.H.; Hagenbuch, B.; Lampe, J.N. Common drugs inhibit human organic cation transporter 1 (OCT1)-mediated neurotransmitter uptake. Drug Metab. Dispos. 2014, 42, 990–995. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.E.; Karbach, U.; Miska, D.; Gorboulev, V.; Akhoundova, A.; Volk, C.; Arndt, P.; Ulzheimer, J.C.; Sonders, M.S.; Baumann, C.; et al. Human neurons express the polyspecific cation transporter hOCT2, which translocates monoamine neurotransmitters, amantadine, and memantine. Mol. Pharmacol. 1998, 54, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Viau, M.; Lafond, J.; Vaillancourt, C. Expression of placental serotonin transporter and 5-HT2A receptor in normal and gestational diabetes mellitus pregnancies. Reprod. Biomed. Online 2009, 19, 207–215. [Google Scholar] [CrossRef]
- Sata, R.; Ohtani, H.; Tsujimoto, M.; Murakami, H.; Koyabu, N.; Nakamura, T.; Uchiumi, T.; Kuwano, M.; Nagata, H.; Tsukimori, K.; et al. Functional analysis of organic cation transporter 3 expressed in human placenta. J. Pharmacol. Exp. Ther. 2005, 315, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayer-Zillgen, M.; Brüss, M.; Bönisch, H. Expression and pharmacological profile of the human organic cation transporters hOCT1, hOCT2 and hOCT3. Br. J. Pharmacol. 2002, 136, 829–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.; Hebert, M.F.; Wagner, D.J.; Esterling, T.R.; Liang, C.J.; Rice, K.; Wang, J. Organic cation transporter 3 facilitates fetal exposure to metformin during pregnancy. Mol. Pharmacol. 2018, 94, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Dallmann, A.; Liu, X.I.; Burckart, G.J.; van den Anker, J. Drug transporters expressed in the human placenta and models for studying maternal-fetal drug transfer. J. Clin. Pharmacol. 2019, 59, S70–S81. [Google Scholar] [CrossRef] [Green Version]
- Shih, J.C. Monoamine oxidase isoenzymes: Genes, functions and targets for behavior and cancer therapy. J. Neural Transm. 2018, 125, 1553–1566. [Google Scholar] [CrossRef] [PubMed]
- Banović, M.; Bordukalo-Nikšić, T.; Balija, M.; Čičin-Šain, L.; Jernej, B. Platelet serotonin transporter (5HTt): Physiological influences on kinetic characteristics in a large human population. Platelets 2010, 21, 429–438. [Google Scholar] [CrossRef]
- Balija, M.; Bordukalo- Nikšić, T.; Mokrović, G.; Banović, M.; Čičin-Šain, L.; Jernej, B. Serotonin level and serotonin uptake in human platelets: A variable interrelation under marked physiological influences. Clin. Chim. Acta 2011, 412, 299–304. [Google Scholar] [CrossRef]
- Beikmann, B.S.; Tomlinson, I.D.; Rosenthal, S.J.; Andrews, A.M. Serotonin uptake is largely mediated by platelets versus lymphocytes in peripheral blood cells. ACS Chem. Neurosci. 2013, 4, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Cool, D.R.; Leibach, F.H.; Ganapathy, V. High-affinity paroxetine binding to the human placental serotonin transporter. Am. J. Physiol. 1990, 259, C196–C204. [Google Scholar] [CrossRef]
- Cool, D.R.; Liebach, F.H.; Ganapathy, V. Interaction of fluoxetine with the human placental serotonin transporter. Biochem. Pharmacol. 1990, 40, 2161–2167. [Google Scholar] [CrossRef]
- Ramamoorthy, S.; Cool, D.R.; Leibach, F.H.; Mahesh, V.B.; Ganapathy, V. Reconstitution of the human placental 5-hydroxytryptamine transporter in a catalytically active form after detergent solubilization. Biochem. J. 1992, 286, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Prasad, P.D.; Leibach, F.H.; Mahesh, V.B.; Ganapathy, V. Human placenta as a target organ for cocaine action: Interaction of cocaine with the placental serotonin transporter. Placenta 1994, 15, 267–278. [Google Scholar] [CrossRef]
- Jayanthi, L.D.; Ramamoorthy, S.; Mahesh, V.B.; Leibach, F.H.; Ganapathys, V. Calmodulin-dependent regulation of the catalytic function of the human serotonin transporter in placental choriocarcinoma cells. J. Biol. Chem. 1994, 269, 14424–14429. [Google Scholar] [CrossRef]
- Decker, A.M.; Blough, B.E. Development of serotonin transporter reuptake inhibition assays using JAR cells. J. Pharmacol. Toxicol. Methods 2018, 92, 52–56. [Google Scholar] [CrossRef]
- Andrade, S.E.; Raebel, M.A.; Brown, J.; Lane, K.; Livingston, J.; Boudreau, D.; Rolnick, S.J.; Roblin, D.; Smith, D.H.; Willy, M.E.; et al. Use of antidepressant medications during pregnancy: A multisite study. Am. J. Obstet. Gynecol. 2008, 198, 194.e1–194.e5. [Google Scholar] [CrossRef]
- Bakker, M.K.; Kölling, P.; van den Berg, P.B.; de Walle, H.E.K.; de Jong van den Berg, L.T.W. Increase in use of selective serotonin reuptake inhibitors in pregnancy during the last decade, a population-based cohort study from the Netherlands. Br. J. Clin. Pharmacol. 2008, 65, 600–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smid, M.C.; Metz, T.D.; Gordon, A.J. Stimulant use in pregnancy: An under-recognized epidemic among pregnant women. Clin. Obstet. Gynecol. 2019, 62, 168–184. [Google Scholar] [CrossRef]
- Hendrick, V.; Stowe, Z.N.; Altshuler, L.L.; Hwang, S.; Lee, E.; Haynes, D. Placental passage of antidepressant medications. Am. J. Psychiatry 2003, 160, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Rampono, J.; Simmer, K.; Ilett, K.F.; Hackett, L.P.; Doherty, D.A.; Elliot, R.; Kok, C.H.; Coenen, A.; Forman, T. Placental transfer of SSRI and SNRI antidepressants and effects on the neonate. Pharmacopsychiatry 2009, 42, 95–100. [Google Scholar] [CrossRef]
- Campbell, N.G.; Koprich, J.B.; Kanaan, N.M.; Lipton, J.W. MDMA Administration to pregnant Sprague-Dawley rats results in its passage to the fetal compartment. Neurotoxicol. Teratol. 2006, 28, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Engel, K.; Zhou, M.; Wang, J. Identification and characterization of a novel monoamine transporter in the human brain. J. Biol. Chem. 2004, 279, 50042–50049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pletscher, A. The 5-hydroxytryptamine system of blood platelets: Physiology and pathophysiology. Int. J. Cardiol. 1987, 14, 177–188. [Google Scholar] [CrossRef]
- Brand, T.; Anderson, G.M. The measurement of platelet-poor plasma serotonin: A systematic review of prior reports and recommendations for improved analysis. Clin. Chem. 2011, 57, 1376–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.M.; Czarkowski, K.; Ravski, N.; Epperson, C.N. Platelet serotonin in newborns and infants: Ontogeny, heritability, and effect of in utero exposure to selective serotonin reuptake inhibitors. Pediatr. Res. 2004, 56, 418–422. [Google Scholar] [CrossRef] [Green Version]
- Daws, L.C. Unfaithful neurotransmitter transporters: Focus on serotonin uptake and implications for antidepressant efficacy. Pharmacol. Ther. 2009, 121, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Eddahibi, S.; Fabre, V.; Boni, C.; Martres, M.P.; Raffestin, B.; Hamon, M.; Adnot, S. Induction of serotonin transporter by hypoxia in pulmonary vascular smooth muscle cells relationship with the mitogenic action of serotonin. Circ. Res. 1999, 84, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Smith, G.N.; Liu, X.; Holden, J.J.A. Association of MAOA, 5-HTT, and NET promoter polymorphisms with gene expression and protein activity in human placentas. Physiol. Genom. 2010, 42, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hadden, C.; Singh, P.; Mercado, C.P.; Murphy, P.; Dajani, N.K.; Lowery, C.L.; Roberts, D.J.; Maroteaux, L.; Kilic, F. GDM-associated insulin deficiency hinders the dissociation of SERT from ERp44 and down-regulates placental 5-HT uptake. Proc. Natl. Acad. Sci. USA 2014, 111, E5697–E5705. [Google Scholar] [CrossRef] [Green Version]
- Blazević, S.; Horvatiček, M.; Kesić, M.; Zill, P.; Hranilović, D.; Ivanišević, M.; Desoye, G.; Štefulj, J. Epigenetic adaptation of the placental serotonin transporter gene (SLC6A4) to gestational diabetes mellitus. PLoS ONE 2017, 12, e0179934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmon, B.; Hartmann, M.; Jones, C.J.; Desoye, G. Insulin and glucose do not affect the glycogen content in isolated and cultured trophoblast cells of human term placenta. J. Clin. Endocrinol. Metab. 1991, 73, 888–893. [Google Scholar] [CrossRef]
- Lang, I.; Schweizer, A.; Hiden, U.; Ghaffari-Tabrizi, N.; Hagendorfer, G.; Bilban, M.; Pabst, M.A.; Korgun, E.T.; Dohr, G.; Desoye, G. Human fetal placental endothelial cells have a mature arterial and a juvenile venous phenotype with adipogenic and osteogenic differentiation potential. Differentiation 2008, 76, 1031–1043. [Google Scholar] [CrossRef]
- Spandidos, A.; Wang, X.; Wang, H.; Seed, B. PrimerBank: A resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucleic Acids Res. 2009, 38, D792–D799. [Google Scholar] [CrossRef] [Green Version]
- Real Time PCR Primer Design—Real Time PCR Probe Design—GenScript. Available online: https://www.genscript.com/tools/real-time-pcr-taqman-primer-design-tool (accessed on 11 March 2021).
- Van Lelyveld, N.; Ter Linde, J.; Schipper, M.E.I.; Samsom, M. Regional differences in expression of TPH-1, SERT, 5-HT(3) and 5-HT(4) receptors in the human stomach and duodenum. Neurogastroenterol. Motil. 2007, 19, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, J.; Yuan, Y.; Yu, X.; Shen, Y.; Xu, Q. Study of a possible role of the monoamine oxidase A (MAOA) gene in paranoid schizophrenia among a chinese population. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2012, 159B, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Métayé, T.; Menet, E.; Guilhot, J.; Kraimps, J.L. Expression and activity of g protein-coupled receptor kinases in differentiated thyroid carcinoma. J. Clin. Endocrinol. Metab. 2002, 87, 3279–3286. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baković, P.; Kesić, M.; Perić, M.; Bečeheli, I.; Horvatiček, M.; George, M.; Čičin-Šain, L.; Desoye, G.; Wadsack, C.; Panzenboeck, U.; et al. Differential Serotonin Uptake Mechanisms at the Human Maternal–Fetal Interface. Int. J. Mol. Sci. 2021, 22, 7807. https://doi.org/10.3390/ijms22157807
Baković P, Kesić M, Perić M, Bečeheli I, Horvatiček M, George M, Čičin-Šain L, Desoye G, Wadsack C, Panzenboeck U, et al. Differential Serotonin Uptake Mechanisms at the Human Maternal–Fetal Interface. International Journal of Molecular Sciences. 2021; 22(15):7807. https://doi.org/10.3390/ijms22157807
Chicago/Turabian StyleBaković, Petra, Maja Kesić, Maja Perić, Ivona Bečeheli, Marina Horvatiček, Meekha George, Lipa Čičin-Šain, Gernot Desoye, Christian Wadsack, Ute Panzenboeck, and et al. 2021. "Differential Serotonin Uptake Mechanisms at the Human Maternal–Fetal Interface" International Journal of Molecular Sciences 22, no. 15: 7807. https://doi.org/10.3390/ijms22157807