
Analysis of Immune Associated Co-Expression Networks Reveals Immune-Related Long Non-Coding RNAs during MI in the Presence and Absence of HDC

Abstract
:1. Introduction
2. Results
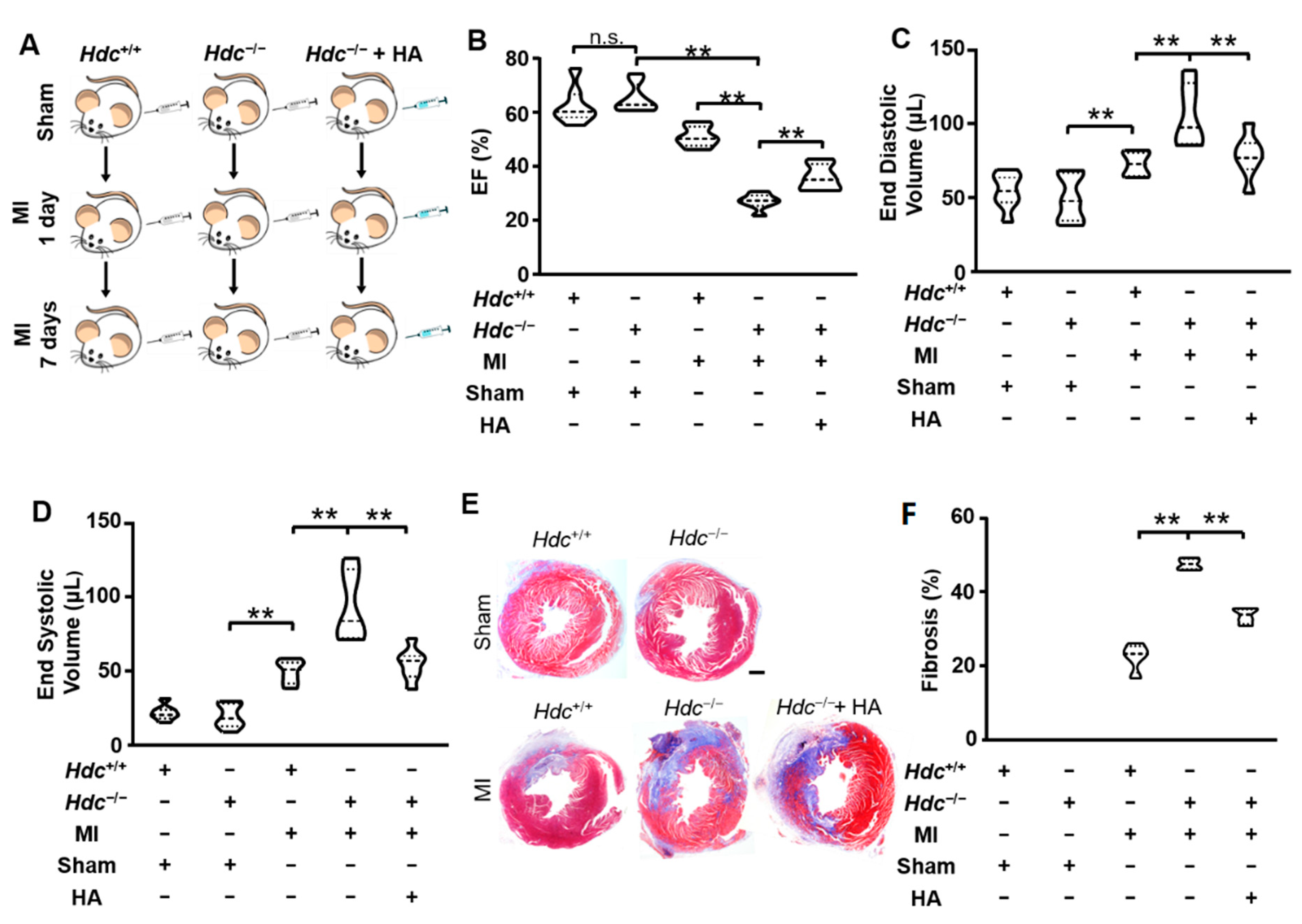
2.1. Hdc-Deletion Induced Aggravated Infarction Injury in Hearts
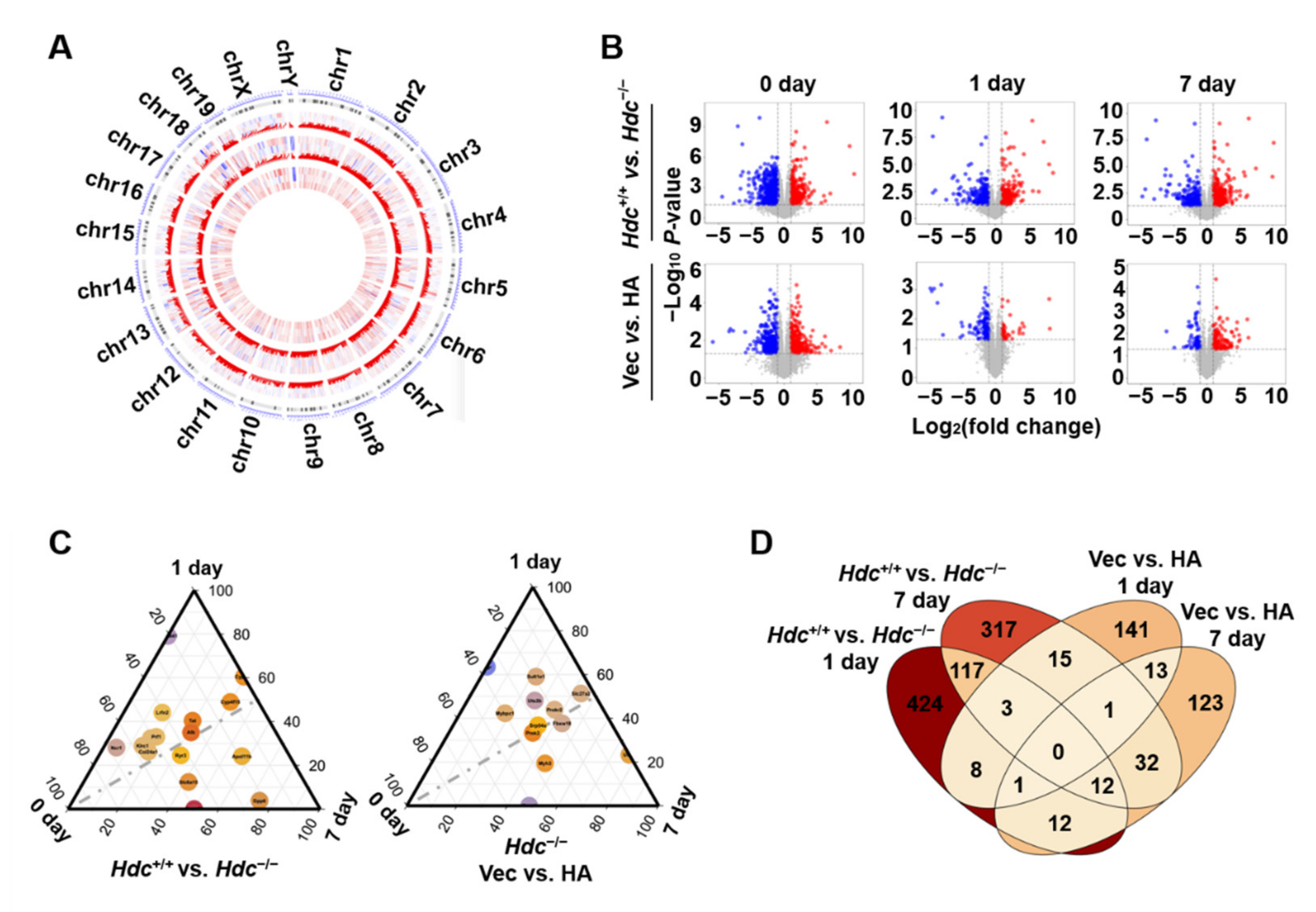
2.2. Identification of DEGs at Different Time Point Post Myocardial Infarction in Hdc Deficiency Mice
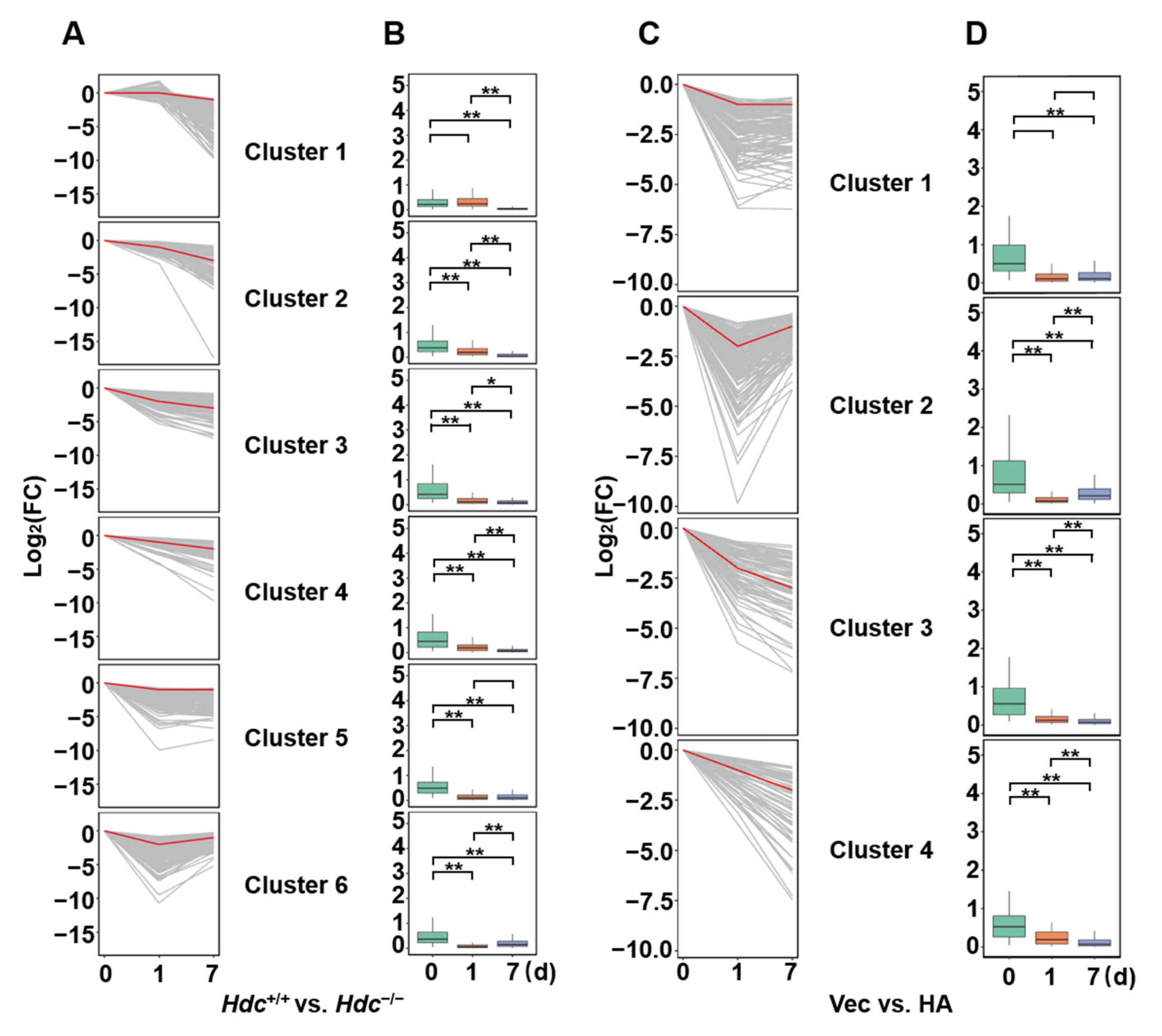
2.3. Time-Dependent Changes in DEGs
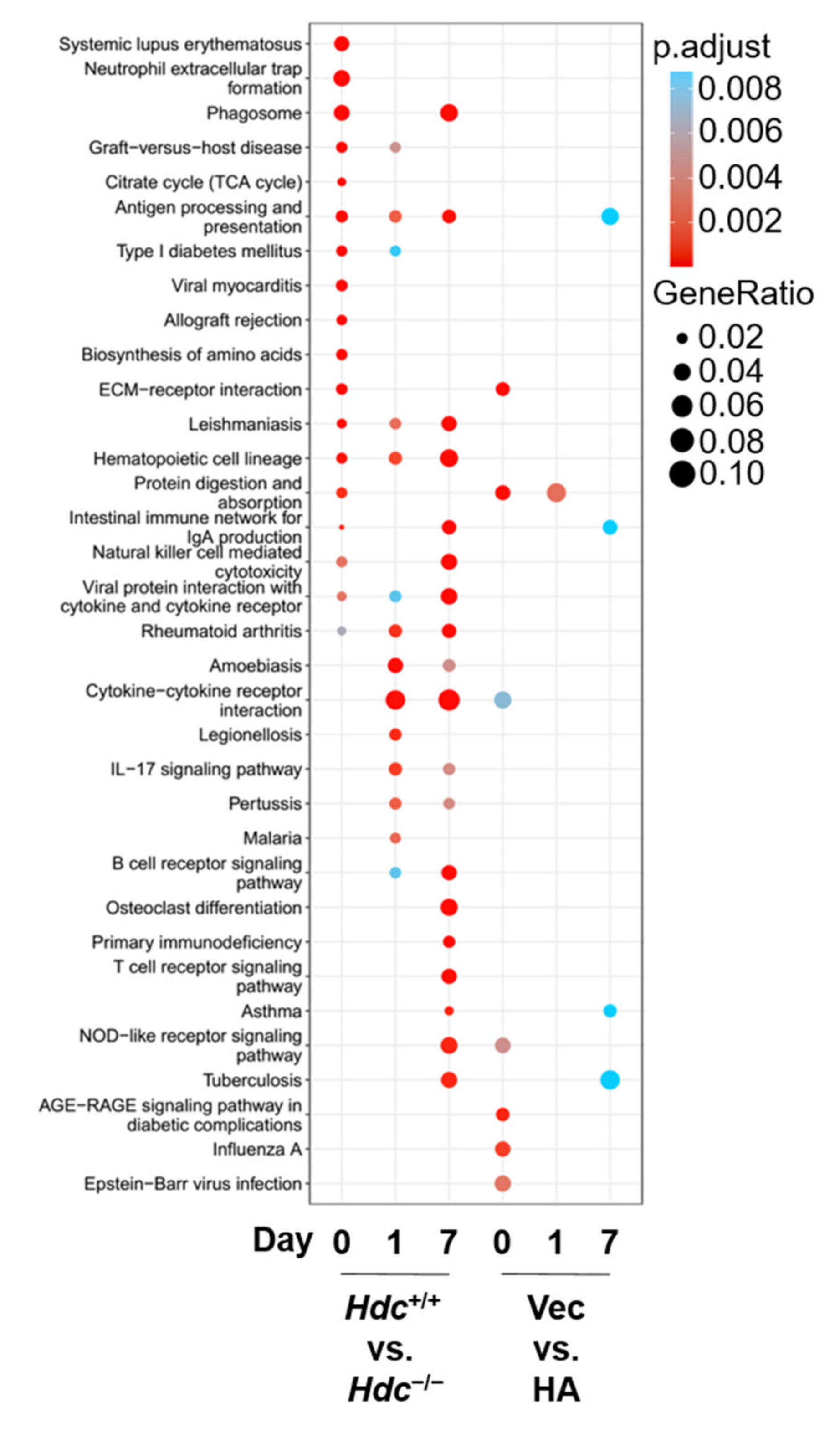
2.4. KEGG Functional Enrichment of DEGs
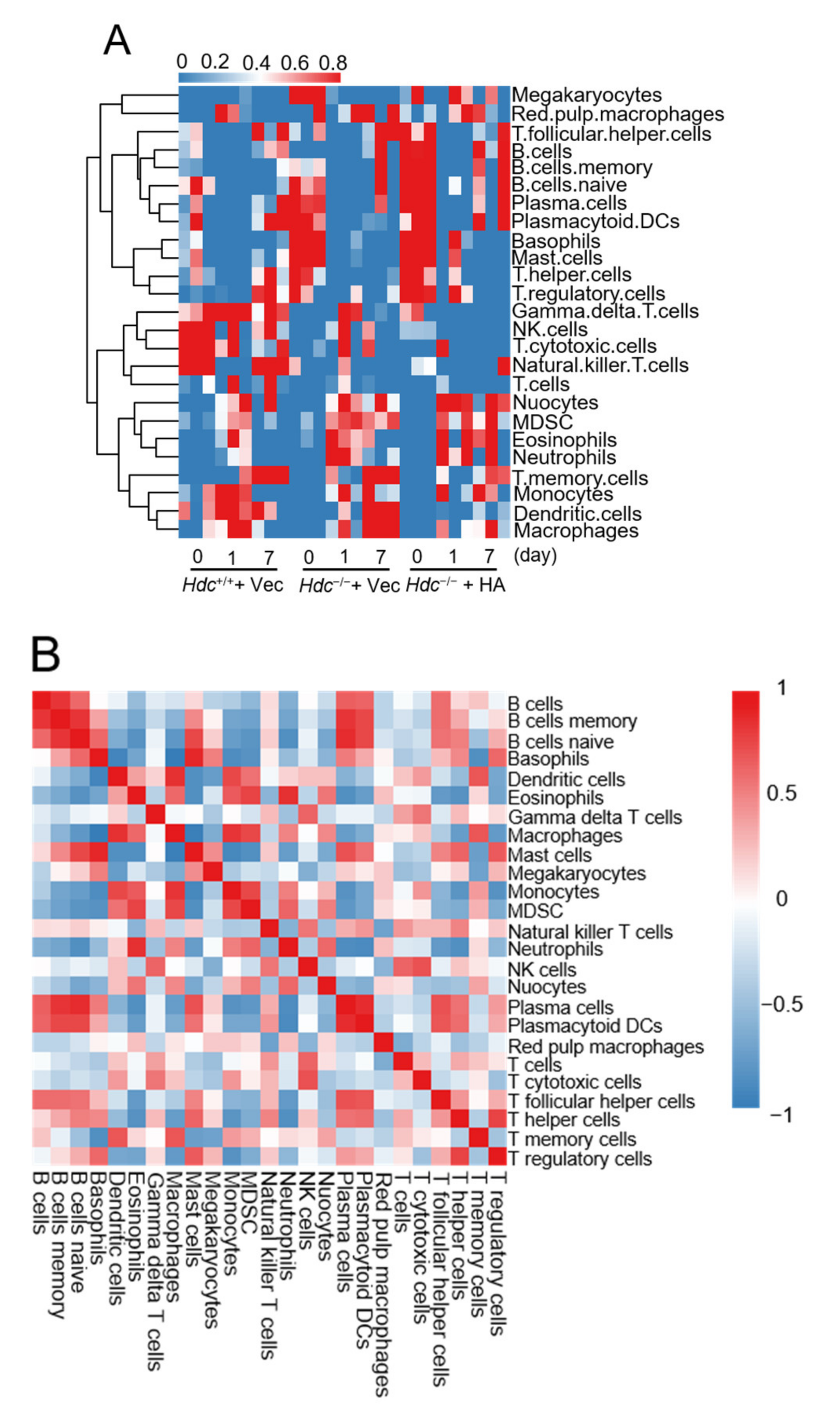
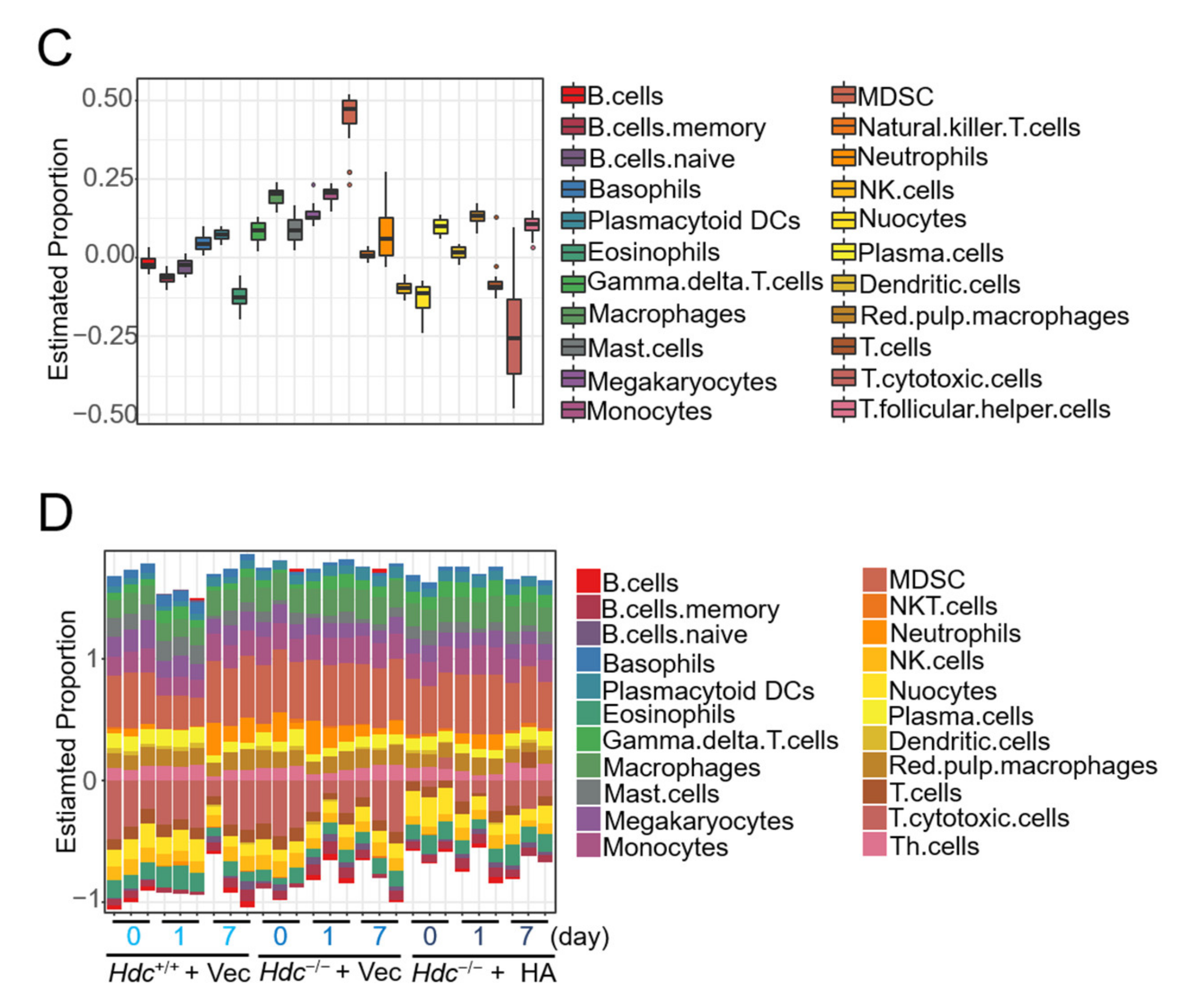
2.5. Immune Cell Profiles of Ischemic Heart in a Histamine-Treated/Hdc-Deficient Myocardial Infarction Model
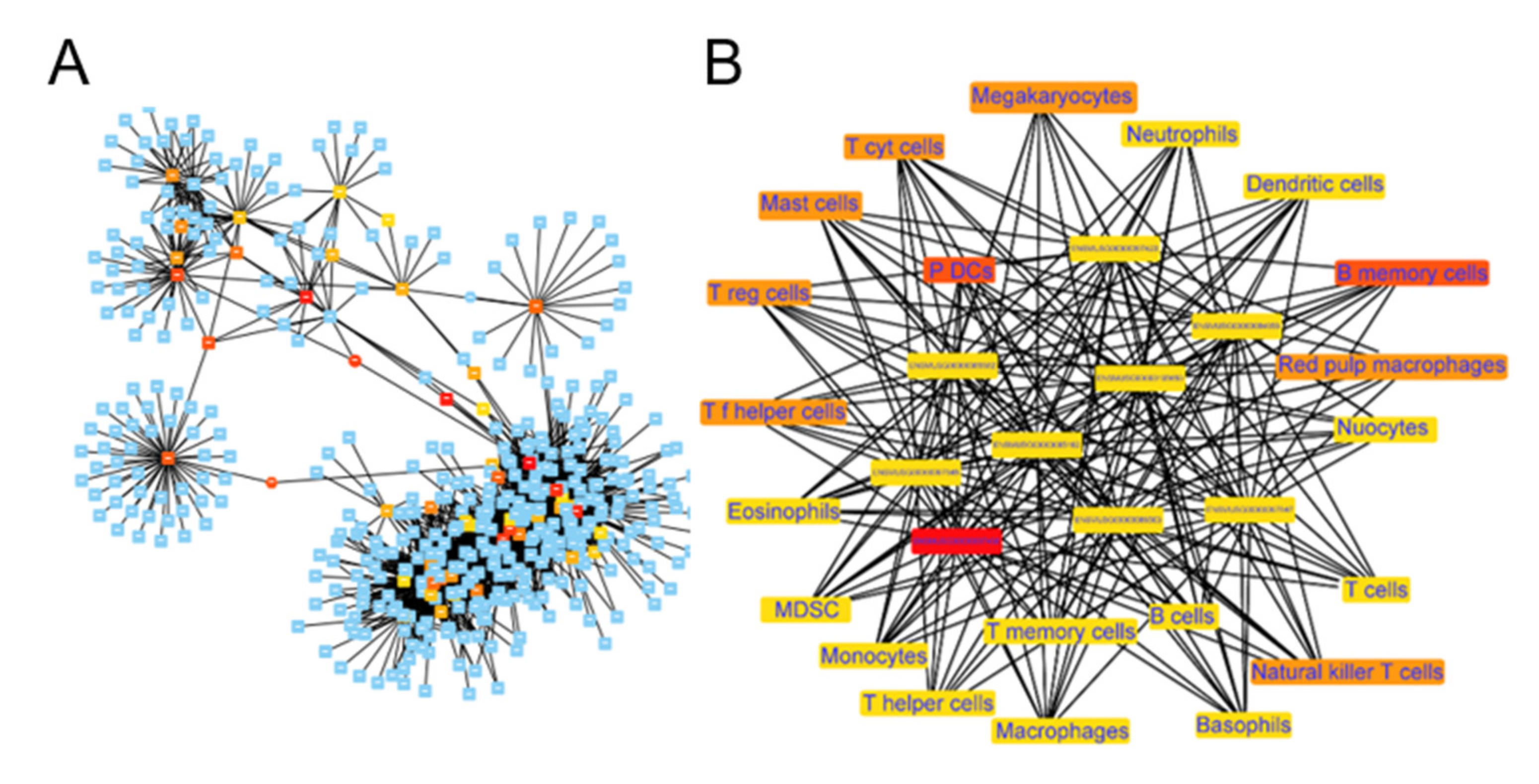
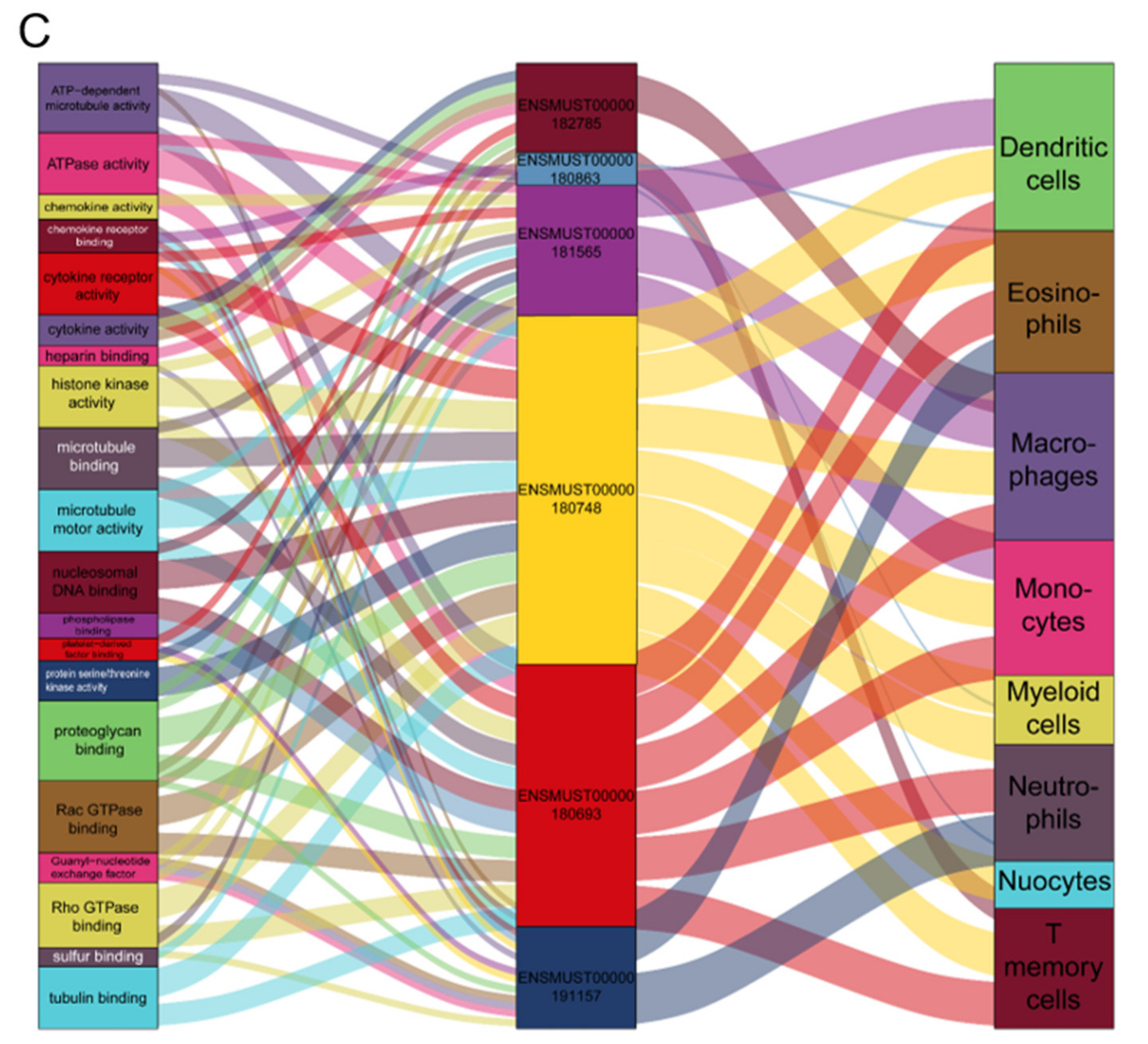
2.6. Functional Co-Expression Network of Imm-lncRNAs
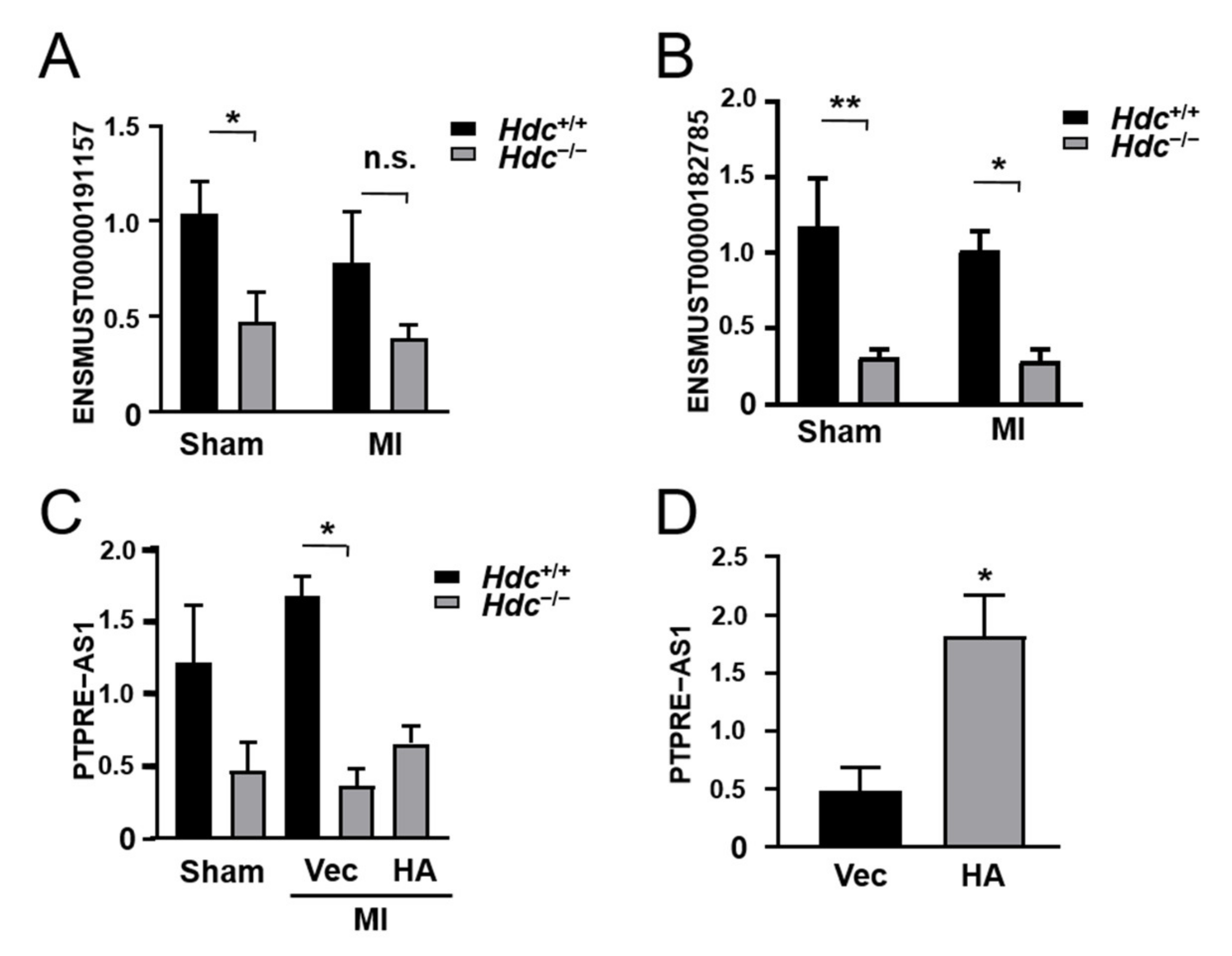
2.7. qPCR Verification of Immune-Related lncRNA Expression in Hdc−/− Mice Model of MI
3. Discussion
4. Materials and Methods
4.1. Animals and Establishment of the MI Mouse Model
4.2. Echocardiography
4.3. Histological Analysis
4.4. RNA-Seq and Data Processing
4.5. Transcriptome Analysis
4.6. Cell Culture
4.7. Quantitative Real-Time PCR
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kalman, J.M.; Lavandero, S.; Mahfoud, F.; Nahrendorf, M.; Yacoub, M.H.; Zhao, D. Looking back and thinking forwards-15 years of cardiology and cardiovascular research. Nat. Rev. Cardiol. 2019, 16, 651–660. [Google Scholar] [CrossRef]
- Severino, P.; D’Amato, A.; Pucci, M.; Infusino, F.; Adamo, F.; Birtolo, L.I.; Netti, L.; Montefusco, G.; Chimenti, C.; Lavalle, C.; et al. Ischemic Heart Disease Pathophysiology Paradigms Overview: From Plaque Activation to Microvascular Dysfunction. Int. J. Mol. Sci. 2020, 21, 8118. [Google Scholar] [CrossRef] [PubMed]
- Monda, E.; Palmiero, G.; Rubino, M.; Verrillo, F.; Amodio, F.; Di Fraia, F.; Pacileo, R.; Fimiani, F.; Esposito, A.; Cirillo, A.; et al. Molecular Basis of Inflammation in the Pathogenesis of Cardiomyopathies. Int. J. Mol. Sci. 2020, 21, 6462. [Google Scholar] [CrossRef]
- Baci, D.; Bosi, A.; Parisi, L.; Buono, G.; Mortara, L.; Ambrosio, G.; Bruno, A. Innate Immunity Effector Cells as Inflammatory Drivers of Cardiac Fibrosis. Int. J. Mol. Sci. 2020, 21, 7165. [Google Scholar] [CrossRef] [PubMed]
- Swirski, F.K.; Nahrendorf, M. Cardioimmunology: The immune system in cardiac homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 733–744. [Google Scholar] [CrossRef]
- Yang, X.D.; Ai, W.; Asfaha, S.; Bhagat, G.; Friedman, R.A.; Jin, G.; Park, H.; Shykind, B.; Diacovo, T.G.; Falus, A. Histamine deficiency promotes inflammation-associated carcinogenesis through reduced myeloid maturation and accumulation of CD11b+ Ly6G+ immature myeloid cells. Nat. Med. 2011, 17, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.; Abudupataer, M.; Zhou, Z.; Chen, J.; Li, H.; Xu, L.; Zhang, W.; Zhang, S.; Zou, Y.; Hong, T. Histamine deficiency aggravates cardiac injury through miR-206/216b-Atg13 axis-mediated autophagic-dependant apoptosis. Cell Death Dis. 2018, 9, 694. [Google Scholar] [CrossRef]
- Noguchi, K.; Ishida, J.; Kim, J.D.; Muromachi, N.; Kako, K.; Mizukami, H.; Lu, W.; Ishimaru, T.; Kawasaki, S.; Kaneko, S.; et al. Histamine receptor agonist alleviates severe cardiorenal damages by eliciting anti-inflammatory programming. Proc. Natl. Acad. Sci. USA 2020, 117, 3150–3156. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Hong, T.; Lin, J.; Ding, S.; Huang, Z.; Chen, J.; Jia, J.; Zou, Y.; Wang, T.C.; Yang, X. Histamine deficiency exacerbates myocardial injury in acute myocardial infarction through impaired macrophage infiltration and increased cardiomyocyte apoptosis. Sci. Rep. 2015, 5, 13131. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Hong, T.; Ding, S.; Deng, L.; Abudupataer, M.; Zhang, W.; Tong, M.; Jia, J.; Gong, H.; Zou, Y.; et al. Aggravated myocardial infarction-induced cardiac remodeling and heart failure in histamine-deficient mice. Sci. Rep. 2017, 7, 44007. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, J.; Li, G.; Xiao, J. Non-coding RNAs in Physiological Cardiac Hypertrophy. Adv. Exp. Med. Biol. 2020, 1229, 149–161. [Google Scholar] [CrossRef]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, discovery, and classification of lncRNAs. Long Non Coding RNA Biol. 2017, 1–46. [Google Scholar]
- Martens, C.R.; Bansal, S.S.; Accornero, F. Cardiovascular inflammation: RNA takes the lead. J. Mol. Cell. Cardiol. 2019, 129, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Ohtsu, H.; Tanaka, S.; Terui, T.; Hori, Y.; Makabe-Kobayashi, Y.; Pejler, G.; Tchougounova, E.; Hellman, L.; Gertsenstein, M.; Hirasawa, N. Mice lacking histidine decarboxylase exhibit abnormal mast cells. FEBS Lett. 2001, 502, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Gerlich, M.; Neumann, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar]
- Barbie, D.A.; Tamayo, P.; Boehm, J.S.; Kim, S.Y.; Moody, S.E.; Dunn, I.F.; Schinzel, A.C.; Sandy, P.; Meylan, E.; Scholl, C.; et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature 2009, 462, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Várkonyi, J.; Zalatnai, A.; Timár, J.; Matolcsi, A.; Falus, A.; Bencsáth, M.; László, V.; Pócsik, E.; Kotlán, B.; Császár, A. Secondary cutaneous infiltration in B cell chronic lymphocytic leukemia. Acta Haematol. 2000, 103, 116–121. [Google Scholar] [CrossRef]
- Tehrani, B.N.; Basir, M.B.; Kapur, N.K. Acute myocardial infarction and cardiogenic shock: Should we unload the ventricle before percutaneous coronary intervention? Prog. Cardiovasc. Dis. 2020, 63, 607–622. [Google Scholar] [CrossRef]
- Nikolsky, E.; Mehran, R.; Dangas, G.; Fahy, M.; Na, Y.; Pocock, S.J.; Lincoff, A.M.; Stone, G.W. Development and validation of a prognostic risk score for major bleeding in patients undergoing percutaneous coronary intervention via the femoral approach. Eur. Heart J. 2007, 28, 1936–1945. [Google Scholar] [CrossRef] [Green Version]
- Ridker, P.M.; Thuren, T.; Zalewski, A.; Libby, P. Interleukin-1β inhibition and the prevention of recurrent cardiovascular events: Rationale and design of the Canakinumab Anti-inflammatory Thrombosis Outcomes Study (CANTOS). Am. Heart J. 2011, 162, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Tardif, J.-C.; Kouz, S.; Waters, D.D.; Bertrand, O.F.; Diaz, R.; Maggioni, A.P.; Pinto, F.J.; Ibrahim, R.; Gamra, H.; Kiwan, G.S. Efficacy and safety of low-dose colchicine after myocardial infarction. N. Engl. J. Med. 2019, 381, 2497–2505. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Takemoto, Y.; Deng, H.; Middelhoff, M.; Friedman, R.A.; Chu, T.H.; Churchill, M.J.; Ma, Y.; Nagar, K.K.; Tailor, Y.H.; et al. Histidine decarboxylase (HDC)-expressing granulocytic myeloid cells induce and recruit Foxp3(+) regulatory T cells in murine colon cancer. Oncoimmunology 2017, 6, e1290034. [Google Scholar] [CrossRef] [Green Version]
- Hercor, M.; Anciaux, M.; Denanglaire, S.; Debuisson, D.; Leo, O.; Andris, F. Antigen-presenting cell-derived IL-6 restricts the expression of GATA3 and IL-4 by follicular helper T cells. J. Leukoc. Biol. 2017, 101, 5–14. [Google Scholar] [CrossRef]
- Han, X.; Huang, S.; Xue, P.; Fu, J.; Liu, L.; Zhang, C.; Yang, L.; Xia, L.; Sun, L.; Huang, S.-K. LncRNA PTPRE-AS1 modulates M2 macrophage activation and inflammatory diseases by epigenetic promotion of PTPRE. Sci. Adv. 2019, 5, eaax9230. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Nurakhayev, S.; Nurkesh, A.; Zharkinbekov, Z.; Saparov, A. Macrophage Polarization in Cardiac Tissue Repair Following Myocardial Infarction. Int. J. Mol. Sci. 2021, 22, 2715. [Google Scholar] [CrossRef]
- Keijer, J.; Li, M.; Speakman, J.R. What is the best housing temperature to translate mouse experiments to humans? Mol. Metab. 2019, 25, 168–176. [Google Scholar] [CrossRef]
- Gao, E.; Lei, Y.H.; Shang, X.; Huang, Z.M.; Zuo, L.; Boucher, M.; Fan, Q.; Chuprun, J.K.; Ma, X.L.; Koch, W.J. A novel and efficient model of coronary artery ligation and myocardial infarction in the mouse. Circ. Res. 2010, 107, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Hamilton, N. An Extension to ‘ggplot2’, for the Creation of Ternary Diagrams [R package ggtern version 2.2.1]. Anesthesiology 2015, 64, 72–86. [Google Scholar]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef] [Green Version]
- Shaw, D.; Ashburner, M.; Blake, J.; Baldarelli, R.; Botstein, D.; Davis, A.; Cherry, J.; Lewis, S.; Lutz, C.; Richardson, J. Gene Ontology: A controlled vocabulary to describe the function, biological process and cellular location of gene products in genome databases. Am. J. Hum. Genet. 1999, 65, A419. [Google Scholar]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Franzén, O.; Gan, L.M.; Björkegren, J.L.M. PanglaoDB: A web server for exploration of mouse and human single-cell RNA sequencing data. Database 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| ENSMUST00000180693 (PTPRE-AS1) | CAGTGAATGAGTGTGGCTCCTG | ACATGTAGAGTGTCCCTCGTTG |
| ENSMUST00000191157 | CTGCCGCTAAGAAGGCGATT | CCCCTGCCAACCCATTCTT |
| ENSMUST00000182785 | TGGTGGTTCCTGACTGTGGAC | AGCACCCACCCAAACAAGTCT |
| Actb | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Ding, S.; Yang, X.; Ge, J. Analysis of Immune Associated Co-Expression Networks Reveals Immune-Related Long Non-Coding RNAs during MI in the Presence and Absence of HDC. Int. J. Mol. Sci. 2021, 22, 7401. https://doi.org/10.3390/ijms22147401
Zhang Z, Ding S, Yang X, Ge J. Analysis of Immune Associated Co-Expression Networks Reveals Immune-Related Long Non-Coding RNAs during MI in the Presence and Absence of HDC. International Journal of Molecular Sciences. 2021; 22(14):7401. https://doi.org/10.3390/ijms22147401
Chicago/Turabian StyleZhang, Zhiwei, Suling Ding, Xiangdong Yang, and Junbo Ge. 2021. "Analysis of Immune Associated Co-Expression Networks Reveals Immune-Related Long Non-Coding RNAs during MI in the Presence and Absence of HDC" International Journal of Molecular Sciences 22, no. 14: 7401. https://doi.org/10.3390/ijms22147401