

The Role of Small Molecules and Their Effect on the Molecular Mechanisms of Early Retinal Organoid Development
 , , and
, , and
Abstract
:1. Introduction
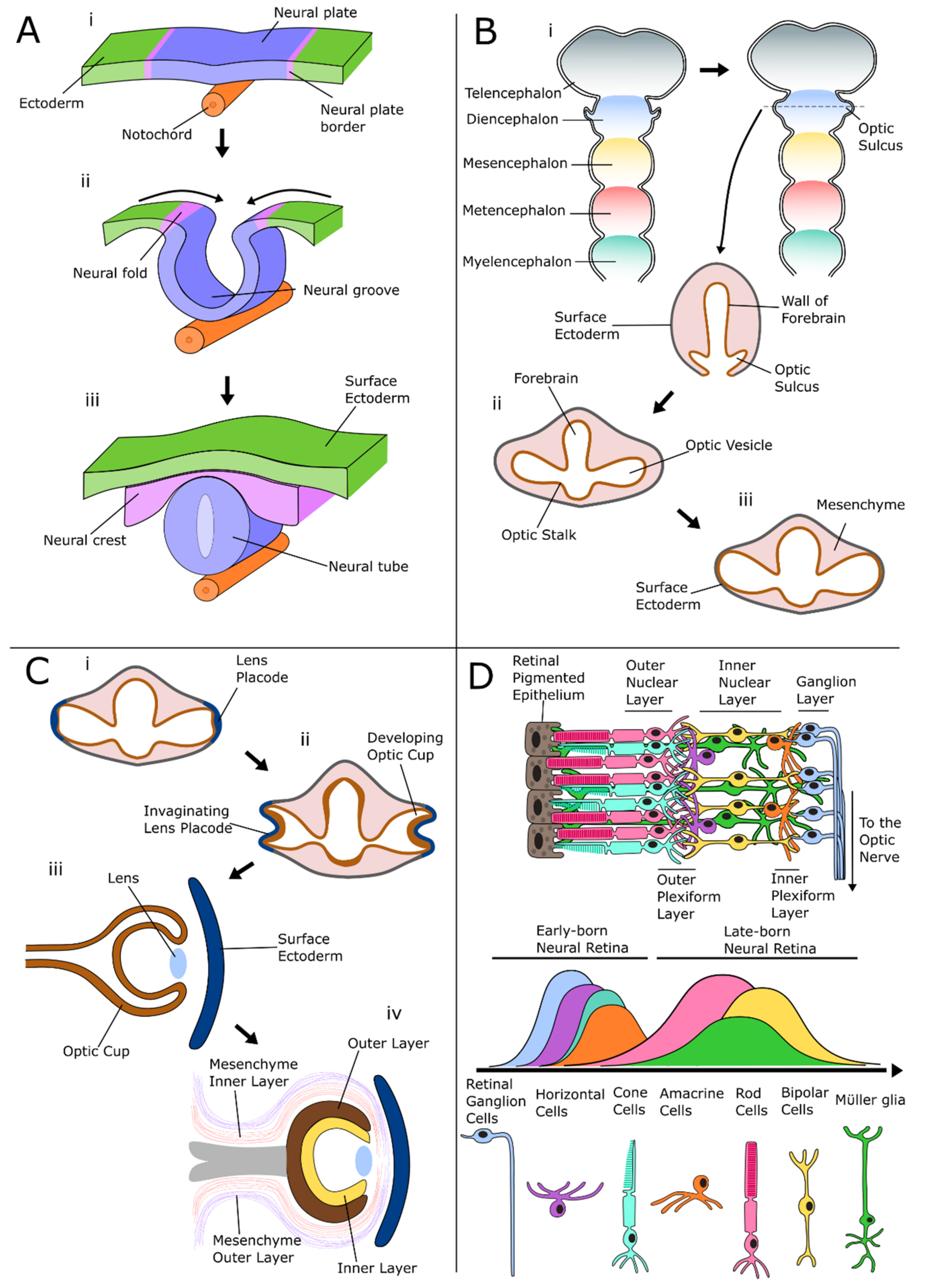
Development of the Retina
2. The Signaling Pathways of the Developing Retina
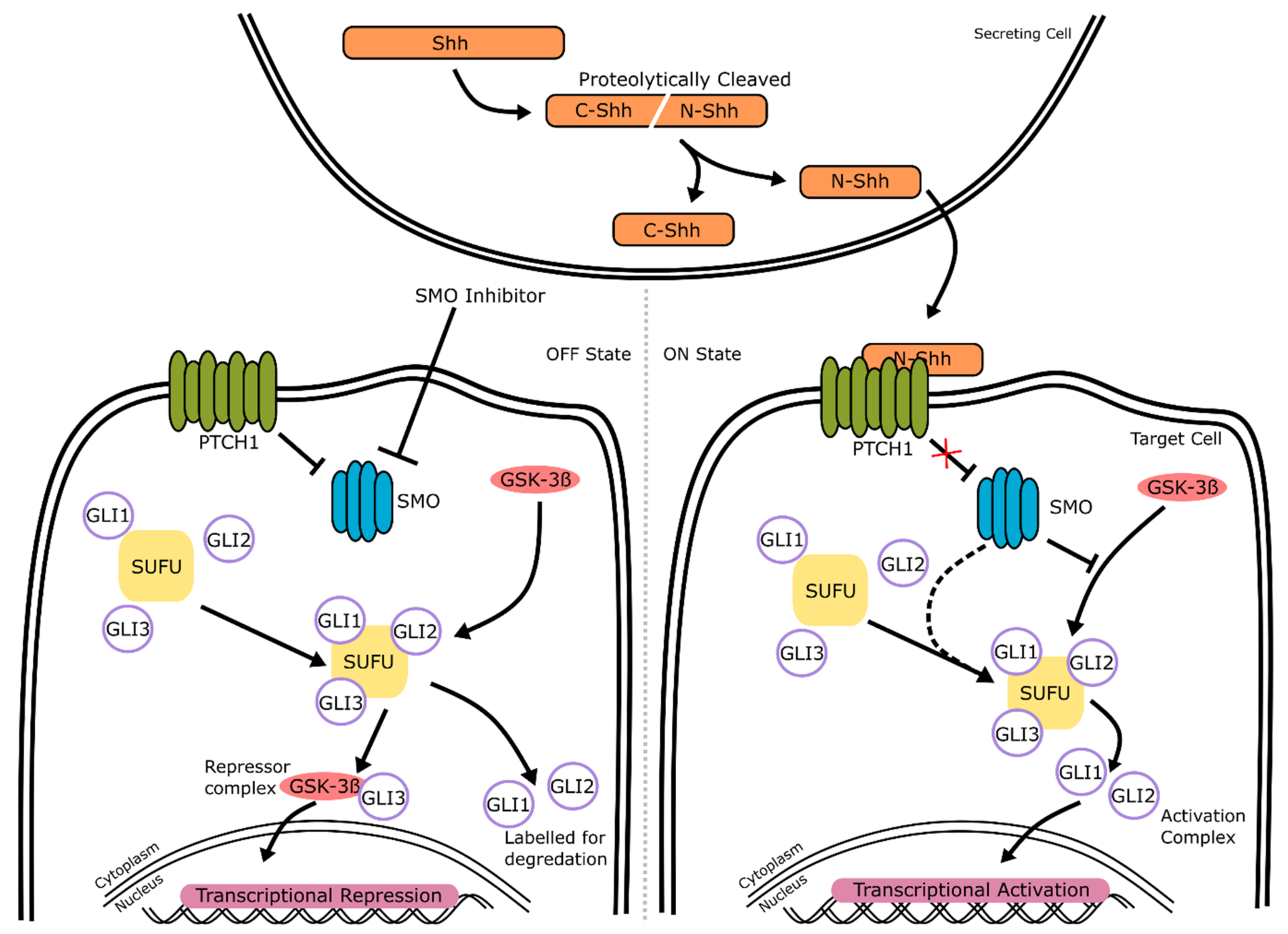
2.1. Hedgehog Signaling
2.1.1. Hedgehog Signaling: Mechanism of Action
2.1.2. Hedgehog Signaling: Neural and Retinal Development
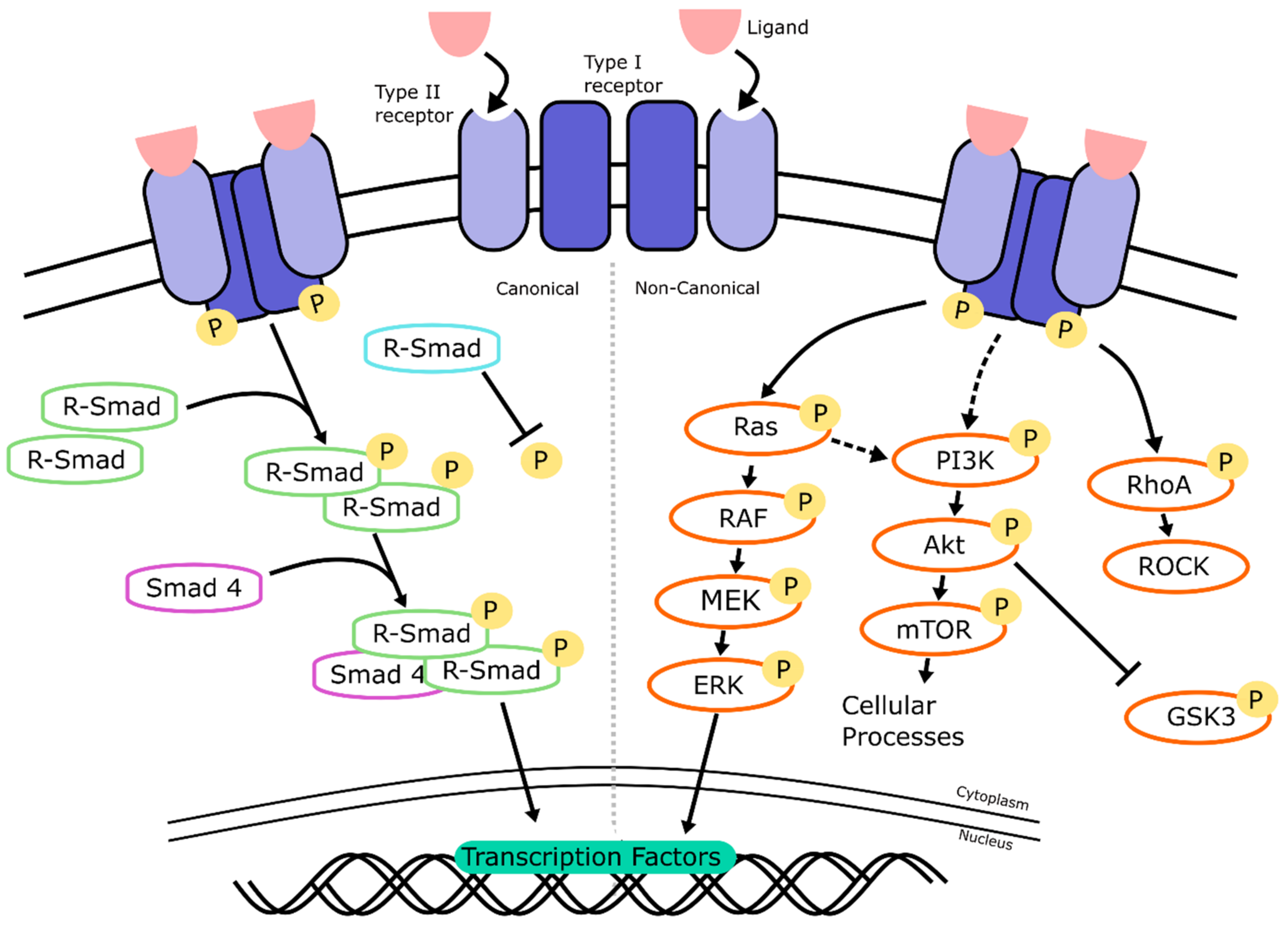
2.2. TGF-β/BMP Signaling
2.2.1. TGF-β/BMP Signaling: Mechanism of Action
2.2.2. TGF-β/BMP Signaling: Neural and Retinal Development
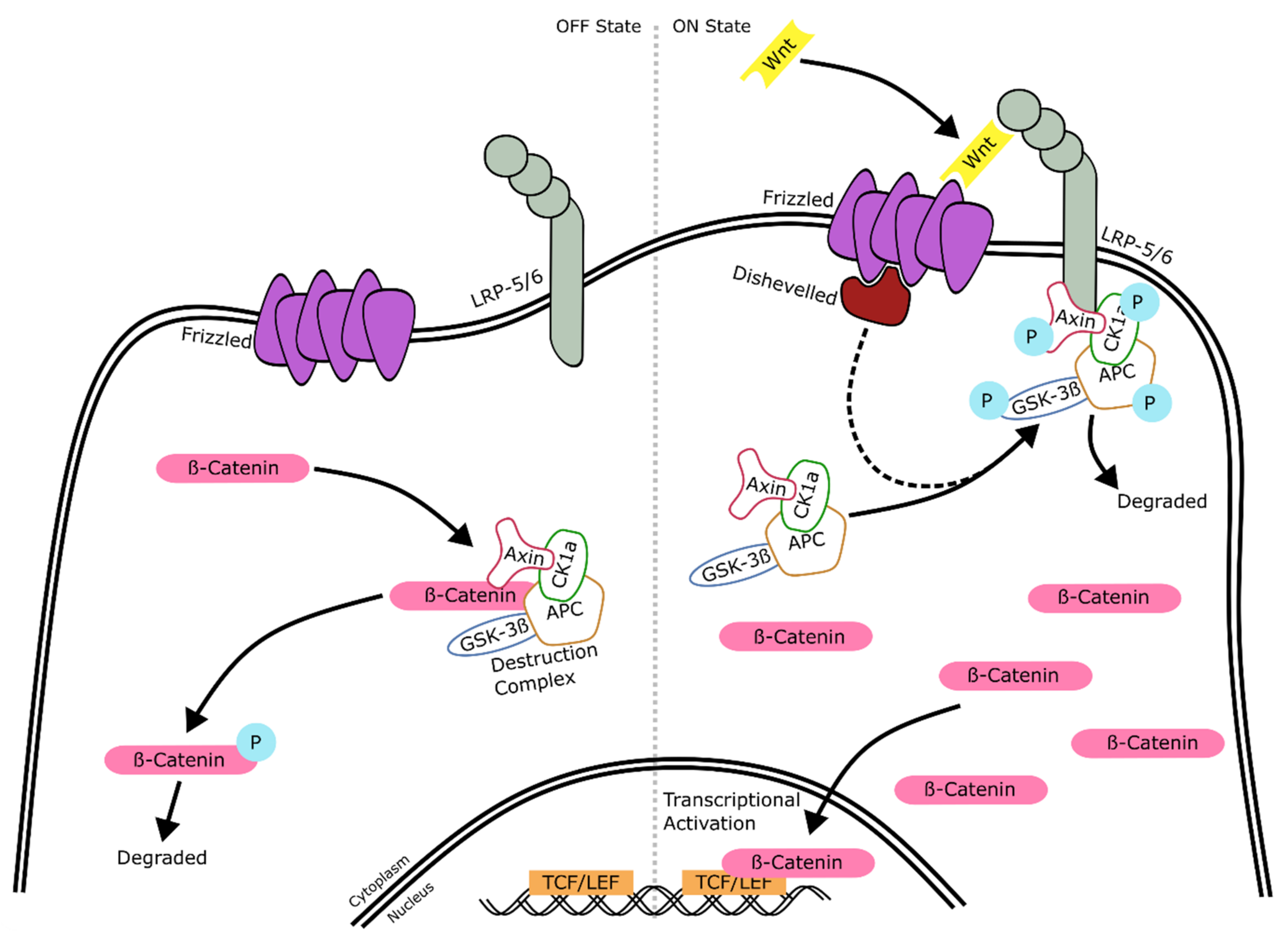
2.3. Wnt Signaling
2.3.1. Wnt Signaling: Mechanism of Action
2.3.2. Wnt Signaling: Neural and Retinal Development
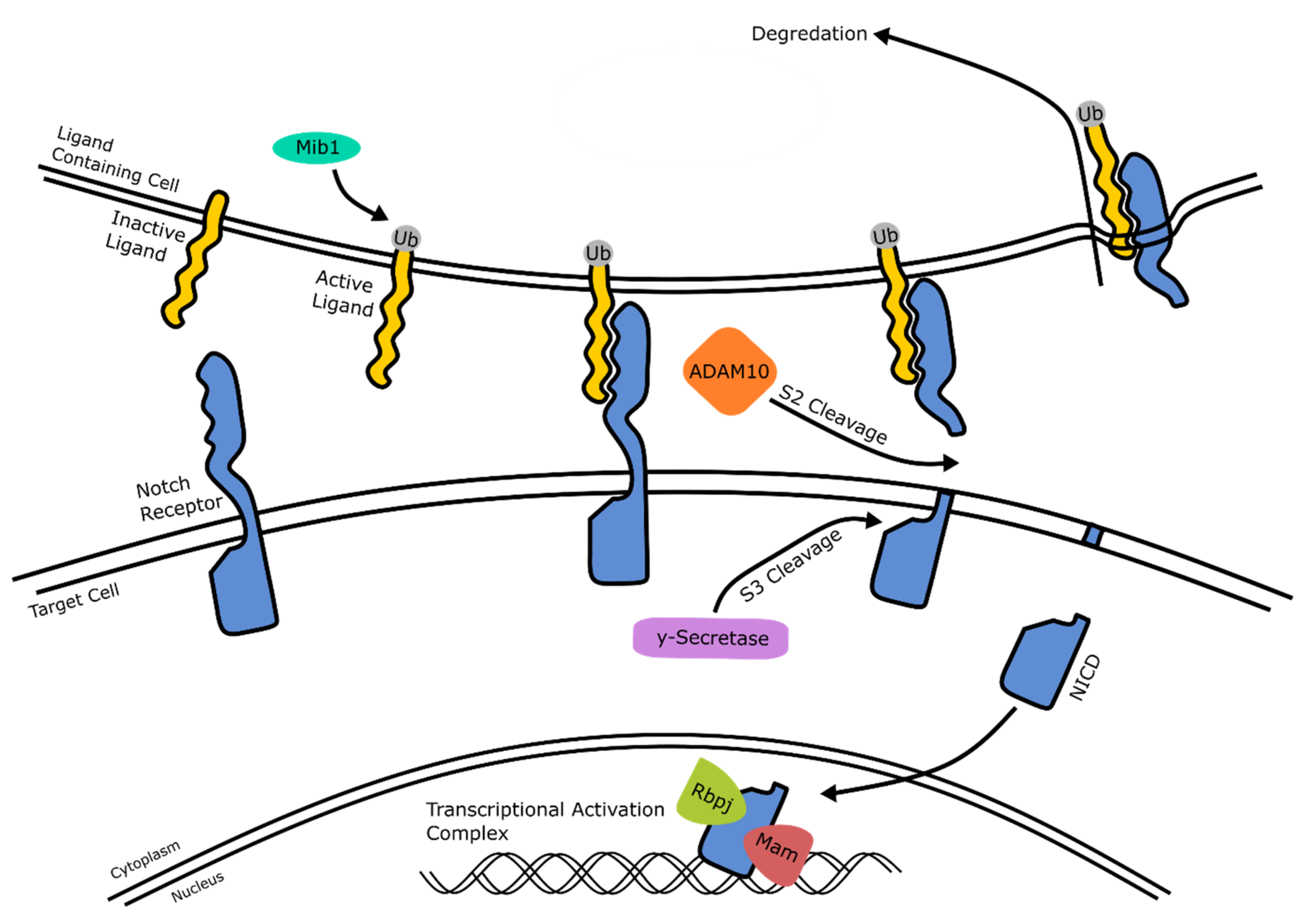
2.4. Notch Signaling
2.4.1. Notch Signaling: Mechanism of Action
2.4.2. Notch Signaling: Neural and Retinal Development
3. Agonists and Antagonists Involved in Regulating the Signaling Pathways of the Developing Retina
3.1. Agonists and Antagonists of the Hedgehog Signaling Pathway
3.2. Agonists and Antagonists of the TGF- β/BMP Signaling Pathway
3.3. Agonists and Antagonists of the Wnt Signaling Pathway
3.4. Agonists and Antagonists of the Notch Signaling Pathway
3.5. Agonists and Antagonists Used in Retinal Models That Regulate Additional Signaling Pathways
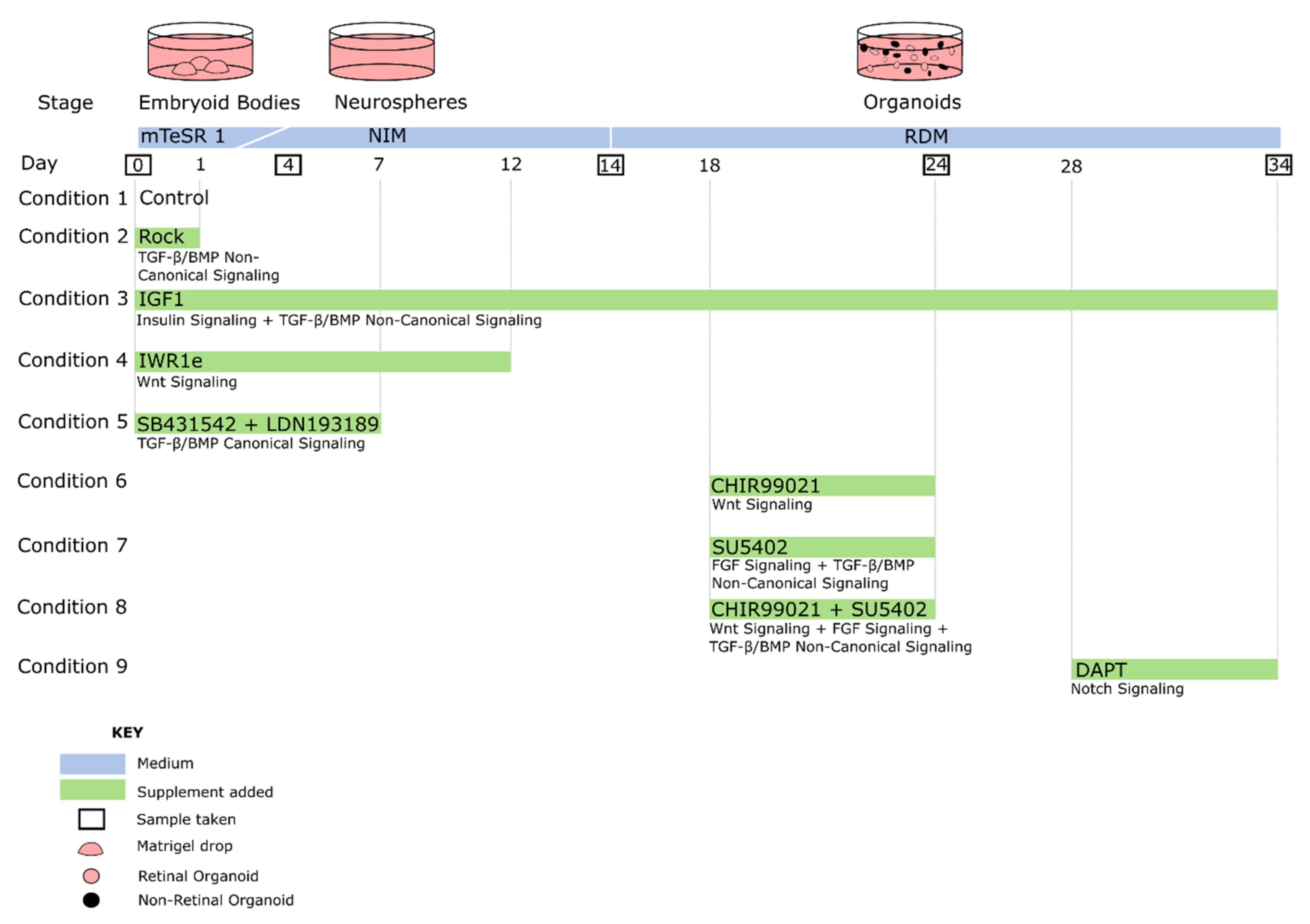
4. Systematic Comparisons of Protocols and Supplements
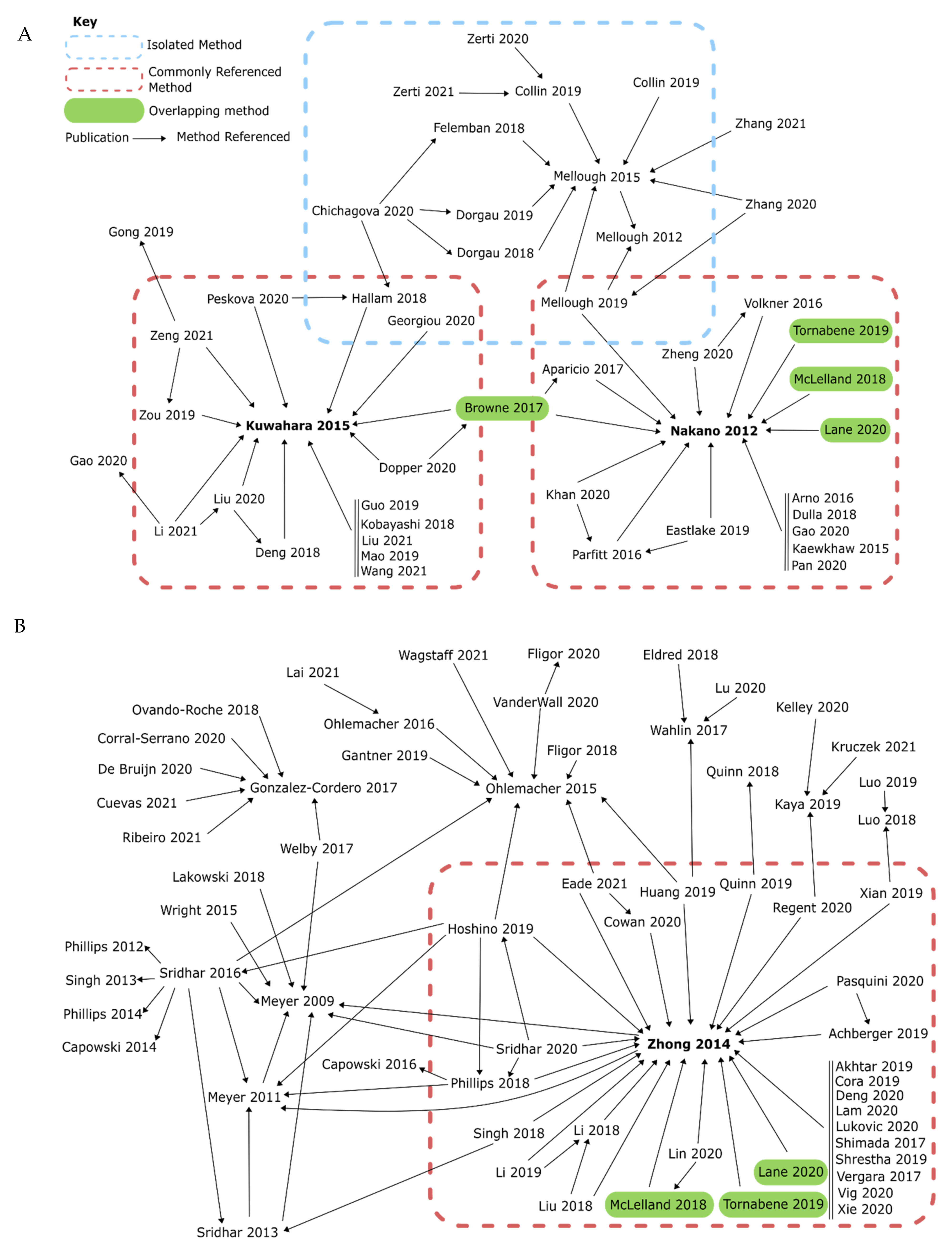
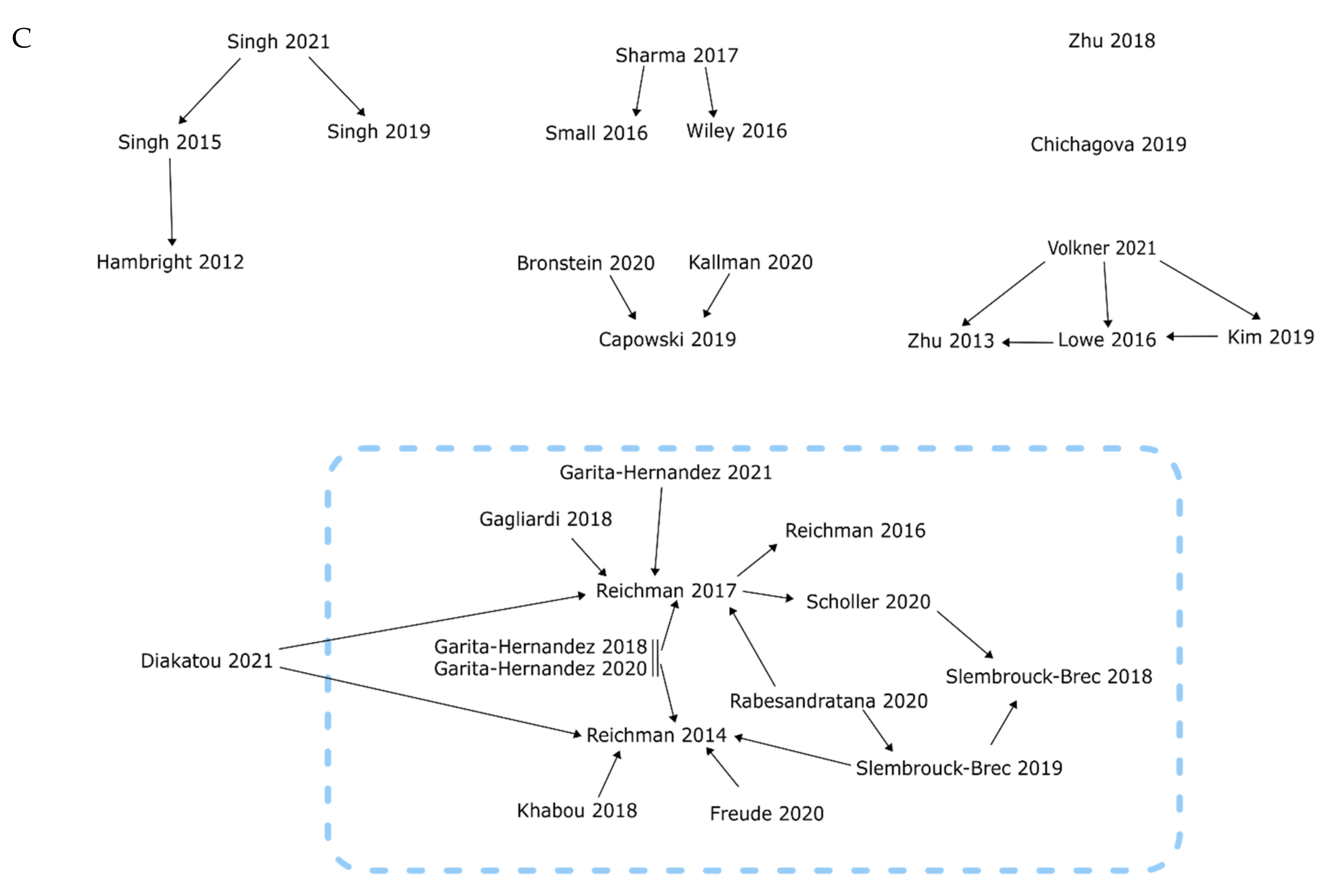
4.1. Directly Comparable Uses of Agonists and Antagonists in Retinal Organoid Models



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Concentration | Length of Addition | Reference | |
|---|---|---|---|---|
| BDNF | Activator of TGF-β and BMP signaling through PI3K and ERK signaling cascades | 20 ng/mL | Day 63 onwards | Singh ‘19 [235] |
| bFGF | FGF signaling ligand | 10 ng/mL | Day 14 onwards | Singh ‘15 [234], Singh ‘19 [235], Singh ‘21 [236] |
| Blebbistatin | Inhibitor of NMII-ATPase, downstream of Rock inhibition | 5 µM | Day 0–1 | Lu ‘20 [211] |
| 10 µM | Day 0–1 | Achberger ‘19 [159], Akhtar ‘19 [160], Cora ‘19 [165], Deng ‘20 [170], Lane ‘20 [202], Li ‘19 [203], Luo ‘18 [214], Luo ‘19 [213], Quinn ‘18 [226], Quinn ‘19 [225], Tornabene ‘19 [240], Vergara ‘17 [242], Vig ‘20 [243], Xian ‘19 [250], Xie ‘20 [251], Zhong ‘14 [155] | ||
| 10 mM | Day 0–1 | Li ‘18 [204], Lin ‘20 [206], Liu ‘18 [209] | ||
| BMP4 | BMP signaling ligand | 50 ng/mL | Day 6–15 | VanderWall ‘20 [241] |
| Day 6–18 | Mao ‘19 [215] | |||
| 55 ng/mL | Day 6–15 | Döpper ‘20 [108] | ||
| Day 6–20 | Khan ‘20 [196] | |||
| 100 ng/mL | Day 0–10 | Meyer ‘09 [144] | ||
| 0.05 µg/µL | Day 5–20 | Hoshino ‘19 [189] | ||
| 0.5 µg/µL | Day 6–12 | Sridhar ‘20 [237] | ||
| 1.5 nM | Day 6–15 | Bronstein ‘20 [130], Capowski ‘19 [49]. Kallman ‘20 [192] | ||
| Day 6–16 | Fligor ‘20 [131] | |||
| Day 6–18 | Browne ‘17 [107], Deng ‘18 [169], Guo ‘19 [188], Hallam ‘18 [145], Kobayashi ‘18 [129], Kuwahara ‘15 [127], Li ‘21 [205], Liu ‘20 [208], Liu ‘21 [207], Phillips ‘18 [224], Wang ‘21 [246], Zeng ‘21 [252], Zou ‘19 [259] | |||
| 2.2 nM | Day 6–18 | Peskova ‘20 [223] | ||
| 2.25 nM | Day 6–18 | Chichagova ‘20 [48], Georgiou ‘20 [146] | ||
| CHIR99021 | Wnt signaling activator through inhibition of GSK-3β | 3 µM | Day 15–18 | Aparicio ‘17 [74], Arno ‘16 [161], Browne ‘17 [107], Lane ‘20 [202], Nakano ‘12 [154], Pan ‘20 [113], Parfitt ‘16 [221], Zheng ‘20 [257] |
| Day 15–21 | Döpper ‘20 [108] | |||
| Day 18–24 * | Guo ‘19 [188], Hallam ‘18 [145] | |||
| 4 µM | Day 18–24 | Georgiou ‘20 [146] | ||
| 10 µM | Unknown | Luo ‘18 [214], Luo ‘19 [213] | ||
| 3 mM | Day 14–17 | Wiley ‘16 [248] | ||
| Day 15–18 | Sharma ‘17 [230] | |||
| Day 18–21 | Kobayashi ‘18 [129] | |||
| COCO | A BMP and Wnt inhibitor, and dual modulator of TGF-β signaling | 30 µM | Day 0–12 | Pan ‘20 [113] |
| Day 0–30 | Pan ‘20 [113] | |||
| DAPT | Inhibitor of notch signaling through inhibition of γ-secretase | 10 µM | Day 28–35 | Khabou ‘18 [195] |
| Day 28–42 * | Eldred ‘18 [177], Lu ‘20 [211], Zerti ‘20 [253] | |||
| Day 29–42 | Shrestha ‘19 [232] | |||
| Day 29–45 | Wahlin ‘17 [245] | |||
| Day 42–49 | Garita-Hernandez ‘20 [186], Garita-Hernandez ‘21 [184] | |||
| Day 44–50 | Garita-Hernandez ‘18 [185] | |||
| Day 60–72 | Zerti ‘20 [253] | |||
| Day 90–102 | Zerti ‘20 [253] | |||
| 10 mM | Day 30–40 | Sharma ‘17 [230], Wiley ‘16 [248] | ||
| DKK-1 | Inhibitor of Wnt signaling through binding of Fz | 10 ng/mL | Day 0–7 | Zhu ‘18 [109] |
| Day 18–21 | Singh ‘15 [234] | |||
| 20 ng/mL | Day 28–35 | Singh ‘19 [235], Singh ‘21 [236] | ||
| 100 ng/mL | Day 2–4 | Meyer ‘11 [135] | ||
| Unknown | Luo ‘18 [214], Luo ‘19 [213] | |||
| Dorsomorphin | BMP inhibitor targeting ALK-2, -3, and -6 inhibition | 100 ng/mL | Day 2–4 | Meyer ‘11 [135] |
| EC23 | Synthetic retinoid | 0.3 µM | Day 18–41 | Völkner ‘16 [101] |
| Day 25–120 | Völkner ‘21 [244] | |||
| FGF2 | FGF signaling ligand | 10 ng/mL | Day 14–21 | Khabou ‘18 [195], Reichman ‘14 [143] |
| Day 21–28 | Reichman ‘14 [143] | |||
| Day 28–35 | Diakatou ‘21 [171], Freude ‘20 [180], Gagliardi ‘18 [181], Garita-Hernandez ‘18 [185], Garita-Hernandez ‘20 [186], Garita-Hernandez ‘21 [184], Rabesandratana ‘20 [158], Reichman ‘14 [143], Reichman ‘17 [156], Scholler ‘20 [229], Slembrouck-Brec ‘18 [157], Slembrouck-Brec ‘19 [150] | |||
| FGF9 | FGF signaling ligand | 10 ng/mL | Day 21 onwards | Singh ‘15 [234] |
| Day 35 onwards | Singh ‘19 [235], Singh ‘21 [236] | |||
| IGF1 | Interacts with insulin receptors to regulate downstream signaling pathways such as Akt and ERK | 5 ng/mL | Day 0–35 | Chichagova ‘19 [162], Chichagova ‘20 [48] |
| Day 0–37 * | Collin ‘19 [163], Collin ‘19 [164], Dorgau ‘18 [173], Dorgau ‘19 [172], Felemban ‘18 [178], Mellough ‘15 [148], Mellough ‘19 [217], Zhang ‘20 [255], Zhang ‘21 [256] | |||
| Day 2–29 | Zerti ‘20 [253] | |||
| 10 ng/mL | Day 0–7 | Zhu ‘18 [109] | ||
| Day 18–21 | Singh ‘15 [234] | |||
| Day 30 onwards | Zerti ‘20 [253] | |||
| Day 35 onwards | Chichagova ‘19 [162], Chichagova ‘20 [48] | |||
| Day 37–90 | Collin ‘19 [163], Collin ‘19 [164], Dorgau ‘18 [173], Dorgau ‘19 [172], Felemban ‘18 [178], Mellough ‘15 [148], Mellough ‘19 [217], Zhang ‘20 [255], Zhang ‘21 [256] | |||
| Day 42–200 | Kaya ‘19 [193], Kelley ‘20 [194], Kruczek ‘21 [198] | |||
| 20 ng/mL | Day 20–30 | Regent ‘20 [227] | ||
| Day 21/28 onwards | Kaya ‘19 [193], Kelley ‘20 [194], Kruczek ‘21 [198] | |||
| Day 28–35 | Singh ‘19 [235], Singh ‘21 [236] | |||
| Day 35 onwards | Regent ‘20 [227] | |||
| IWR1e | Inhibitor of Wnt signaling through stabilization of AXIN | 3 nM | Day 0–12 | Sharma ‘17 [230], Wiley ‘16 [248] |
| 2 µM | Day 0–7 | Zhu ‘18 [109] | ||
| 3 µM | Day 0–6 | Döpper ‘20 [108] | ||
| Day 1–6 | Eldred ‘18 [177], Wahlin ‘17 [245] | |||
| Day 1–8 | Lu ‘20 [211] | |||
| Day 0–12 * | Arno ‘16 [161], Eastlake ‘19 [176], Gao ‘20 [183], Kaewkhaw ‘15 [191], Lane ‘20 [202], Nakano ‘12 [154], Pan ‘20 [113], Parfitt ‘16 [221], Völkner ‘16 [101], Zheng ‘20 [257] | |||
| Day 0–18 | Browne ‘17 [107] | |||
| Day 2–12 | Aparicio ‘17 [74], Browne ‘17 [107] | |||
| Day 2–14 | Dulla ‘18 [174] | |||
| 3 mM | Day 2–20 | Khan ‘20 [196] | ||
| LDN193189 | Inhibitor of BMP signaling through inhibition of ALK-2, -3, and -6 receptors | 100 nM | Day 0–6 | Döpper ‘20 [108] |
| Day 0–7* | Zhu ‘18 [109] | |||
| 3 µM | Day 0–6 | Browne ‘17 [107] | ||
| Noggin | TGF-β signaling ligand | 10 ng/mL | Day 0–7 | Zhu ‘18 [109] |
| 100 ng/mL | Day 0 onwards | Singh ‘15 [234], Singh ‘19 [235], Singh ‘21 [236] | ||
| Day 2–4 | Meyer ‘11 [135] | |||
| Rock Inhibitor (Y-27632) | Inhibitor of Rock | 10 µM | Day 0–1* | Achberger ‘19 [159], Aparicio ‘17 [74], Browne ‘17 [107], Cora ‘19 [165], Cowan ‘20 [152], Lai ‘21 [199], Lam ‘20 [201], Shrestha ‘19 [232], Zerti ‘20 [253], Zerti ‘21 [254] |
| Day 0–2 | Collin ‘19 [163], Collin ‘19 [164], Dorgau ‘18 [173], Dorgau ‘19 [172], Felemban ‘18 [178], Georgiou ‘20 [146], Hallam ‘18 [145] | |||
| Day 0–14 | Dulla ‘18 [174] | |||
| Day 24–25 | Achberger ‘19 [159], Cora ‘19 [165] | |||
| 20 µM | Day 0–1 | Döpper ‘20 [108], Kaya ‘19 [193], Kelley ‘20 [194], Kruczek ‘21 [198], Regent ‘20 [227], Zheng ‘20 [257] | ||
| Day 0–4 | Kaewkhaw ‘15 [191] | |||
| Day 0–6 | Deng ‘18 [169], Guo ‘19 [188], Kobayashi ‘18 [129], Kuwahara ‘15 [127], Wang ‘21 [246], Zeng ‘21 [252], Zou ‘19 [259] | |||
| Day 0–12 | Eastlake ‘19 [176], Gao ‘20 [183], Lane ‘20 [202], Nakano ‘12 [154], Pan ‘20 [113], Völkner ‘16 [101] | |||
| Day 0–18 | Li ‘21 [205], Liu ‘20 [208], Liu ‘21 [207], Parfitt ‘16 [221] | |||
| Day 0–20 | Khan ‘20 [196] | |||
| 10 mM | Day 0–1 | Shimada ‘17 [231] | ||
| 20 mM | Day 0–12 | Sharma ‘17 [230], Wiley ‘16 [248] | ||
| SAG | Hedgehog pathway activator through activation of Smo | 100 nM | Day 8–16 | Eldred ‘18 [177] |
| Day 10–18 | Lu ‘20 [211] | |||
| Day 12–18 | Eastlake ‘19 [176], Gao ‘20 [183], Kaewkhaw ‘15 [191], Völkner ‘16 [101], Wahlin ‘17 [245] | |||
| Day 12–20 | Khan ‘20 [196] | |||
| Day 14–17 | Wiley ‘16 [248] | |||
| Day 14–20 | Dulla ‘18 [174] | |||
| Day 15–18 | Aparicio ‘17 [74], Arno ‘16 [161], Browne ‘17 [107], Lane ‘20 [202], Nakano ‘12 [154], Pan ‘20 [113], Parfitt ‘16 [221], Sharma ‘17 [230], Zheng ‘20 [257] | |||
| SB431542 | Inhibitor of TGF-β signaling through inhibition of ALK-4, -5, and -7 receptors | 3 µM | Day 0–6 | Browne ‘17 [107] |
| 10 µM | Day 0–6 | Döpper ‘20 [108] | ||
| Day 0–7* | Zhu ‘18 [109] | |||
| SU5402 | Inhibitor of FGF signaling through inhibition of fibroblast growth fact receptor 1 (FGFR-1) | 2.5 µM | Day 18–24 | Chichagova ‘20 [48], Georgiou ‘20 [146] |
| 5 µM | Day 15–21 | Döpper ‘20 [108] | ||
| Day 18–24* | Guo ‘19 [188], Hallam ‘18 [145] | |||
| 10 µM | Day 0–10 | Meyer ‘09 [144] | ||
| Day 16–40 | Meyer ‘09 [144] | |||
| 3 mM | Day 18–21 | Kobayashi ‘18 [129] | ||
| Wnt3a | Wnt signaling activator | 100 ng/mL | Day 0–10 | Meyer ‘09 [144] |
| XAV939 | Wnt signaling inhibitor through stabilizing AXIN | 100 ng/mL | Day 2–4 | Meyer ‘11 [135] |
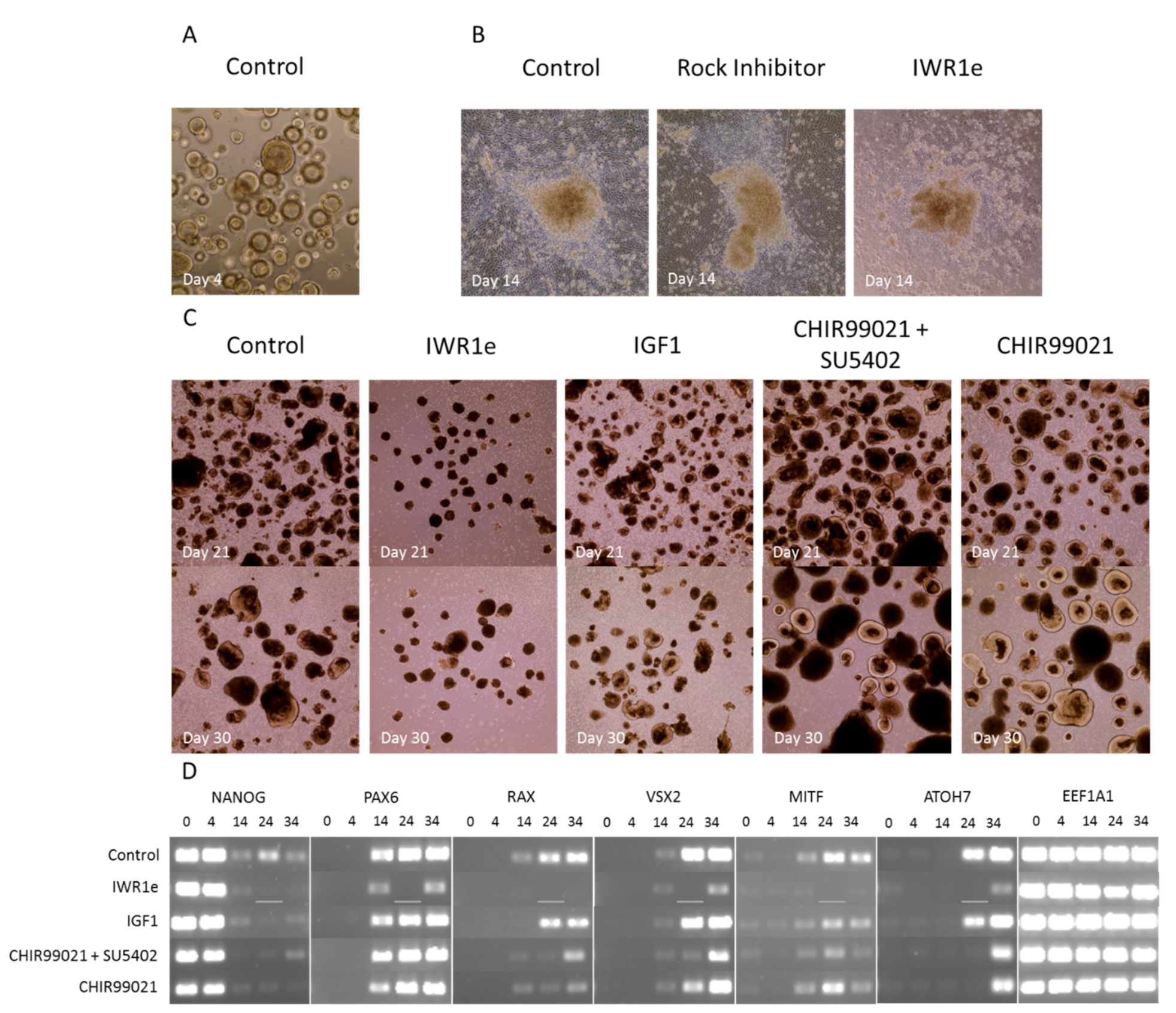
4.2. Comparison of Specific Signaling Pathway Modulators and Their Effect on Retinal Organoid Differentiation
4.2.1. Regulation of Signaling Pathways by External Factors Affecting Retinal Development
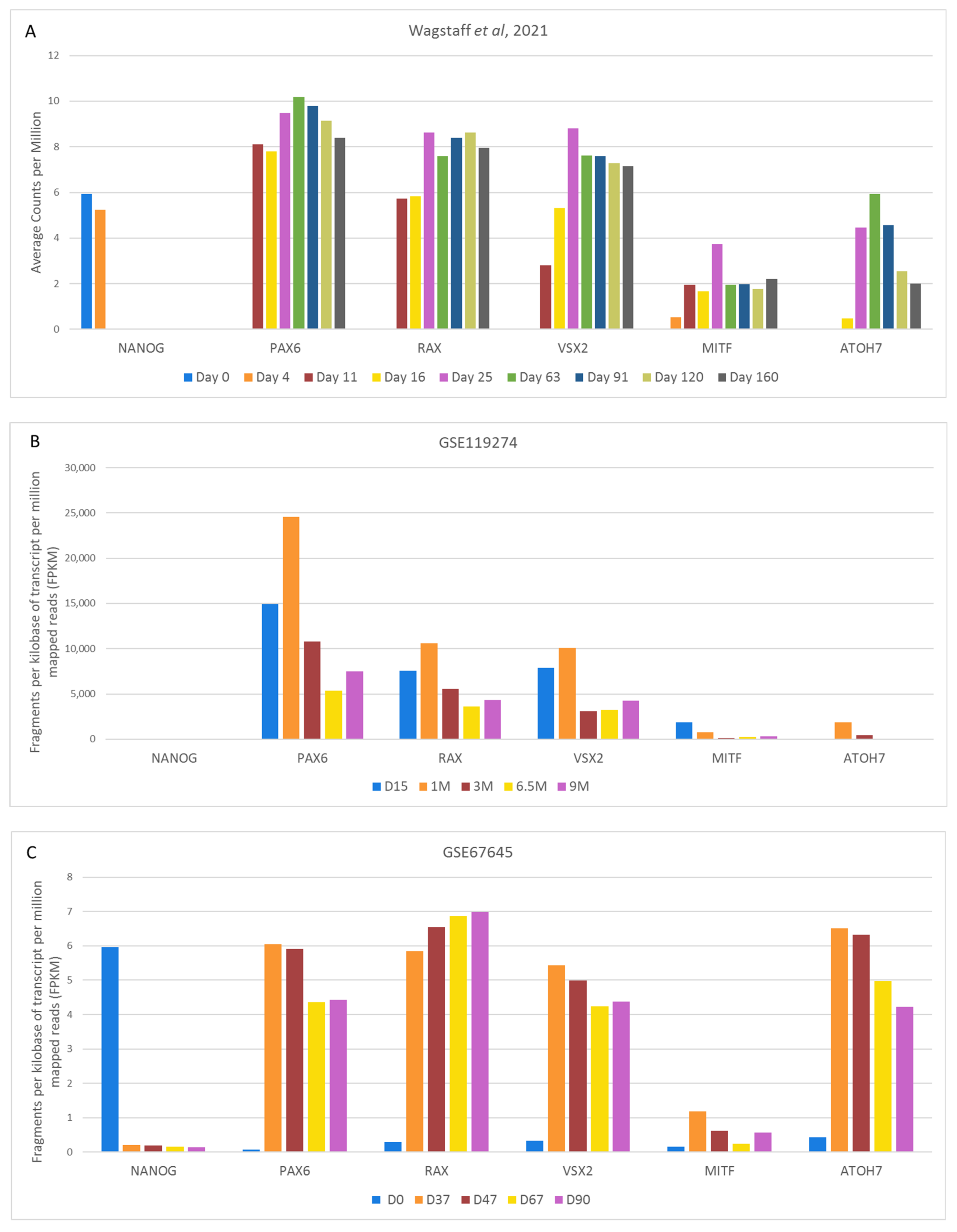
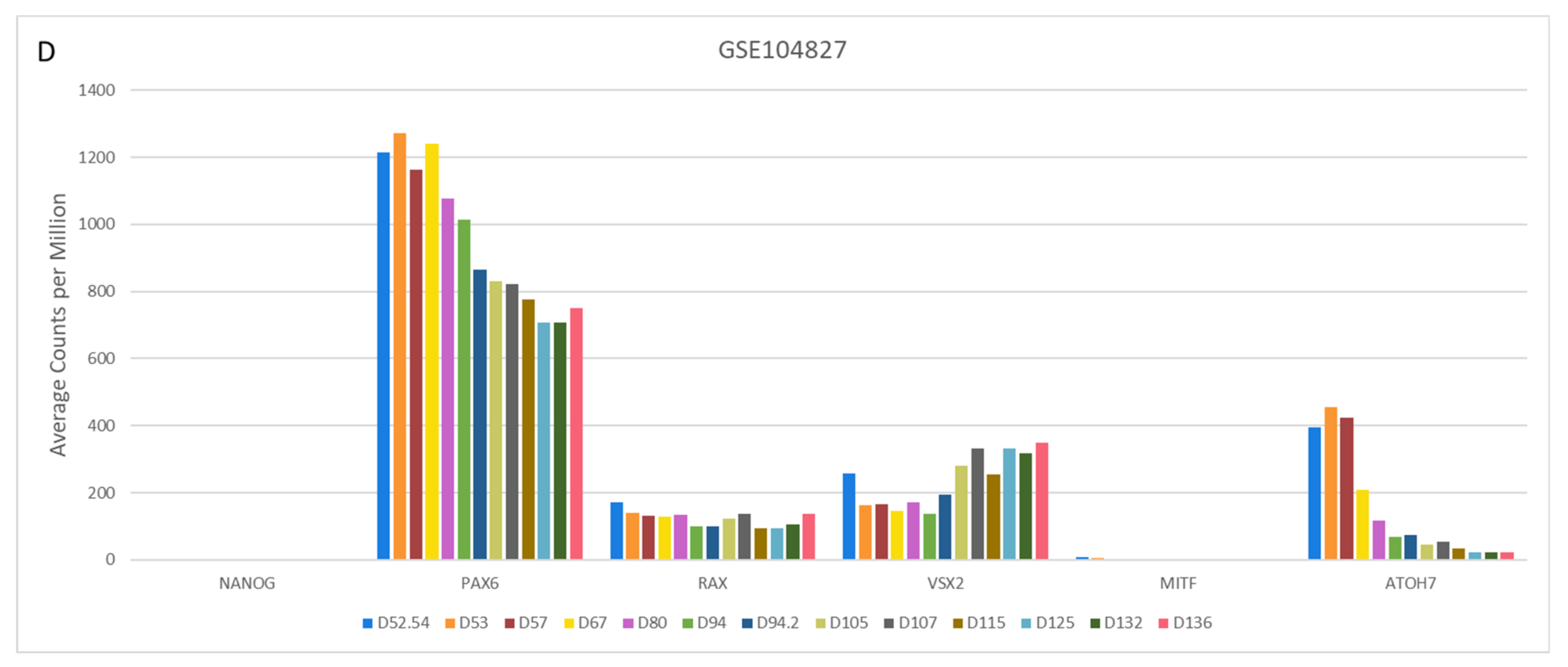
4.2.2. Variable Retinal Development Confirmed by Gene Expression Changes of Key Markers
4.3. Comparison of Cell Type-Specific Expression during Retinal Organoid Development
5. Discussion and Conclusions
5.1. The Use of Different Protocols Gives Rise to Variable and Difficult to Compare Cultures
5.2. Healthy Early Development of Embryoid Bodies Is Not a Guarantee for Healthy Organoids
5.3. One Pathway, One Agonist/Antagonist?
5.4. Does the Synergistic Addition of Multiple Factors Result in an Additive Effect, or Is It Not Necessary?
5.5. Research Implications and Future Perspectives
6. Materials and Methods
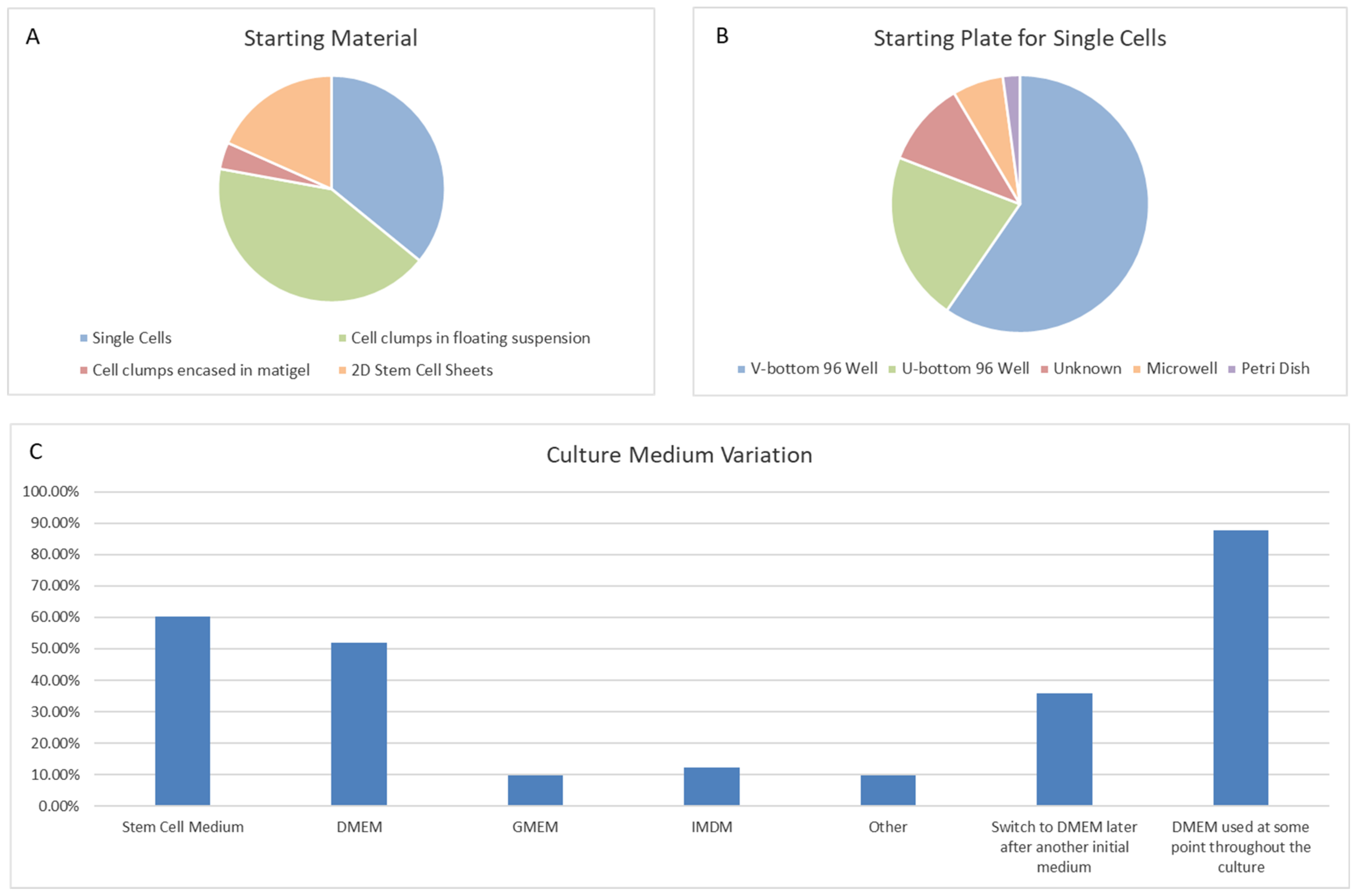
6.1. Protocol Search
6.2. Organoid Generation
6.3. RT-PCR and sqPCR
6.4. RNA-Seq Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Y.; Peng, S.; Finnell, R.H.; Zheng, Y. Organoids as a new model system to study neural tube defects. FASEB J. 2021, 35, e21545. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.M.; Wijnholds, J. Retinogenesis of the human fetal retina: An apical polarity perspective. Genes 2019, 10, 987. [Google Scholar] [CrossRef] [PubMed]
- O’Hara-Wright, M.; Gonzalez-Cordero, A. Retinal organoids: A window into human retinal development. Development 2020, 147, dev189746. [Google Scholar] [CrossRef] [PubMed]
- Lachke, S.; Maas, R.L. Building the developmental oculome: Systems biology in vertebrate eye development and disease. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 305–323. [Google Scholar] [CrossRef]
- Bryan, C.D.; Casey, M.A.; Pfeiffer, R.L.; Jones, B.W.; Kwan, K.M. Optic cup morphogenesis requires neural crest-mediated basement membrane assembly. Development 2020, 147, dev181420. [Google Scholar] [CrossRef] [PubMed]
- Gage, P.J.; Rhoades, W.; Prucka, S.K.; Hjalt, T. Fate maps of neural crest and mesoderm in the mammalian eye. Investig. Opthalmol. Vis. Sci. 2005, 46, 4200–4208. [Google Scholar] [CrossRef] [PubMed]
- Aavani, T.; Tachibana, N.; Wallace, V.; Biernaskie, J.; Schuurmans, C. Temporal profiling of photoreceptor lineage gene expression during murine retinal development. Gene Expr. Patterns 2017, 23–24, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Ratnapriya, R.; Brooks, M.J.; Chaitankar, V.; Wilken, M.S.; Zhang, C.; Starostik, M.R.; Gieser, L.; La Torre, A.; Nishio, M.; et al. Molecular anatomy of the developing human retina. Dev. Cell 2017, 43, 763–779.e4. [Google Scholar] [CrossRef]
- Centanin, L.; Wittbrodt, J. Retinal neurogenesis. Development 2014, 141, 241–244. [Google Scholar] [CrossRef]
- Mustafi, D.; Engel, A.H.; Palczewski, K. Structure of cone photoreceptors. Prog. Retin. Eye Res. 2009, 28, 289–302. [Google Scholar] [CrossRef]
- Stenkamp, D.L. Development of the vertebrate eye and retina. Prog. Mol. Biol. Transl. Sci. 2015, 134, 397–414. [Google Scholar] [CrossRef]
- Machold, R.; Fishell, G. Hedgehog patterns midbrain ARChitecture. Trends Neurosci. 2002, 25, 10–11. [Google Scholar] [CrossRef]
- Roelink, H.; Augsburger, A.; Heemskerk, J.; Korzh, V.; Norlin, S.; I Altaba, A.R.; Tanabe, Y.; Placzek, M.; Edlund, T.; Jessell, T.; et al. Floor plate and motor neuron induction by vhh-1, a vertebrate homolog of hedgehog expressed by the notochord. Cell 1994, 76, 761–775. [Google Scholar] [CrossRef]
- Ingham, P.W. Hedgehog signaling in animal development: Paradigms and principles. Genes Dev. 2001, 15, 3059–3087. [Google Scholar] [CrossRef]
- Choy, S.W.; Cheng, S.H. Hedgehog signaling. Vitam. Horm. 2012, 88, 1–23. [Google Scholar] [CrossRef]
- E Gallardo, V.; Bovolenta, P. Positive and negative regulation of Shh signalling in vertebrate retinal development. F1000Research 2018, 7, 1934. [Google Scholar] [CrossRef]
- Cavodeassi, F.; Creuzet, S.; Etchevers, H.C. The hedgehog pathway and ocular developmental anomalies. Qual. Life Res. 2019, 138, 917–936. [Google Scholar] [CrossRef]
- Rahnama, F.; Shimokawa, T.; Lauth, M.; Finta, C.; Kogerman, P.; Teglund, S.; Toftgård, R.; Zaphiropoulos, P.G. Inhibition of GLI1 gene activation by Patched1. Biochem. J. 2006, 394, 19–26. [Google Scholar] [CrossRef]
- Chiang, C.; Litingtung, Y.; Lee, E.; Young, K.E.; Corden, J.L.; Westphal, H.; Beachy, P.A. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nat. Cell Biol. 1996, 383, 407–413. [Google Scholar] [CrossRef]
- Neumann, C.J. Patterning of the Zebrafish retina by a wave of Sonic hedgehog activity. Science 2000, 289, 2137–2139. [Google Scholar] [CrossRef]
- Russell, C. The roles of hedgehogs and fibroblast growth factors in eye development and retinal cell rescue. Vis. Res. 2003, 43, 899–912. [Google Scholar] [CrossRef]
- Zhao, L.; Zevallos, S.E.; Rizzoti, K.; Jeong, Y.; Lovell-Badge, R.; Epstein, D.J. Disruption of SoxB1-dependent Sonic hedgehog expression in the hypothalamus causes septo-optic dysplasia. Dev. Cell 2012, 22, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Trousse, F.; Martí, E.; Gruss, P.; Torres, M.; Bovolenta, P. Control of retinal ganglion cell axon growth: A new role for Sonic hedgehog. Development 2001, 128, 3927–3936. [Google Scholar] [CrossRef] [PubMed]
- Dakubo, G.D.; Mazerolle, C.; Furimsky, M.; Yu, C.; St-Jacques, B.; McMahon, A.P.; Wallace, V.A. Indian hedgehog signaling from endothelial cells is required for sclera and retinal pigment epithelium development in the mouse eye. Dev. Biol. 2008, 320, 242–255. [Google Scholar] [CrossRef]
- Stenkamp, D.L.; Frey, R.A.; Prabhudesai, S.N.; Raymond, P. Function for hedgehog genes in Zebrafish retinal development. Dev. Biol. 2000, 220, 238–252. [Google Scholar] [CrossRef]
- Kerr, C.L.; Huang, J.; Williams, T.; West-Mays, J.A. Activation of the hedgehog signaling pathway in the developing lens stimulates EctopicFoxE3 expression and disruption in fiber cell differentiation. Investig. Opthalmol. Vis. Sci. 2012, 53, 3316–3330. [Google Scholar] [CrossRef]
- Hata, A.; Chen, Y.-G. TGF-β signaling from receptors to smads. Cold Spring Harb. Perspect. Biol. 2016, 8, a022061. [Google Scholar] [CrossRef]
- Braunger, B.M.; Pielmeier, S.; Demmer, C.; Landstorfer, V.; Kawall, D.; Abramov, N.; Leibinger, M.; Kleiter, I.; Fischer, D.; Jägle, H.; et al. TGF- signaling protects retinal neurons from programmed cell death during the development of the mammalian eye. J. Neurosci. 2013, 33, 14246–14258. [Google Scholar] [CrossRef]
- Nickel, J.; Mueller, T.D. Specification of BMP signaling. Cells 2019, 8, 1579. [Google Scholar] [CrossRef]
- Murali, D.; Kawaguchi-Niida, M.; Deng, C.-X.; Furuta, Y. Smad4 is required predominantly in the developmental processes dependent on the BMP branch of the TGF-β signaling system in the embryonic mouse retina. Investig. Opthalmol. Vis. Sci. 2011, 52, 2930–2937. [Google Scholar] [CrossRef]
- Heldin, C.-H.; Moustakas, A. Signaling receptors for TGF-β family members. Cold Spring Harb. Perspect. Biol. 2016, 8, a022053. [Google Scholar] [CrossRef]
- Byfield, S.D.; Major, C.; Laping, N.J.; Roberts, A.B. SB-505124 is a selective inhibitor of transforming growth factor-β Type I receptors ALK4, ALK5, and ALK7. Mol. Pharmacol. 2004, 65, 744–752. [Google Scholar] [CrossRef]
- Wharton, K.; Derynck, R. TGF family signaling: Novel insights in development and disease. Development 2009, 136, 3691–3697. [Google Scholar] [CrossRef]
- Zhang, Y.E. Non-Smad signaling pathways of the TGF-β family. Cold Spring Harb. Perspect. Biol. 2016, 9, a022129. [Google Scholar] [CrossRef]
- Nishina, S.; Kohsaka, S.; Yamaguchi, Y.; Handa, H.; Kawakami, A.; Fujisawa, H.; Azuma, N. PAX6 expression in the developing human eye. Br. J. Ophthalmol. 1999, 83, 723–727. [Google Scholar] [CrossRef]
- Castellano, E.; Downward, J. RAS interaction with PI3K: More than just another effector pathway. Genes Cancer 2011, 2, 261–274. [Google Scholar] [CrossRef]
- Duda, P.; Akula, S.M.; Abrams, S.L.; Steelman, L.S.; Martelli, A.M.; Cocco, L.; Ratti, S.; Candido, S.; Libra, M.; Montalto, G.; et al. Targeting GSK3 and associated signaling pathways involved in cancer. Cells 2020, 9, 1110. [Google Scholar] [CrossRef]
- Dünker, N.; Schuster, N.; Krieglstein, K. TGF-beta modulates programmed cell death in the retina of the developing chick embryo. Development 2001, 128, 1933–1942. [Google Scholar] [CrossRef]
- Chavarría, T.; Valenciano, A.I.; Mayordomo, R.; Egea, J.; Comella, J.X.; Hallböök, F.; De Pablo, F.; De La Rosa, E.J. Differential, age-dependent MEK-ERK and PI3K-Akt activation by insulin acting as a survival factor during embryonic retinal development. Dev. Neurobiol. 2007, 67, 1777–1788. [Google Scholar] [CrossRef]
- Kim, J.; Wu, H.-H.; Lander, A.D.; Lyons, K.M.; Matzuk, M.M.; Calof, A.L. GDF11 Controls the timing of progenitor cell competence in developing retina. Science 2005, 308, 1927–1930. [Google Scholar] [CrossRef]
- Chang, K.-C.; Sun, C.; Cameron, E.G.; Madaan, A.; Wu, S.; Xia, X.; Zhang, X.; Tenerelli, K.; Nahmou, M.; Knasel, C.M.; et al. Opposing effects of growth and differentiation factors in cell-fate specification. Curr. Biol. 2019, 29, 1963–1975.e5. [Google Scholar] [CrossRef]
- Schille, C.; Schambony, A. Signaling pathways and tissue interactions in neural plate border formation. Neurogenesis 2017, 4, e1292783. [Google Scholar] [CrossRef]
- Chesnutt, C.; Burrus, L.W.; Brown, A.M.; Niswander, L. Coordinate regulation of neural tube patterning and proliferation by TGFβ and WNT activity. Dev. Biol. 2004, 274, 334–347. [Google Scholar] [CrossRef]
- Chambers, S.M.; Fasano, C.A.; Papapetrou, E.P.; Tomishima, M.; Sadelain, M.; Studer, L. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nat. Biotechnol. 2009, 27, 275–280. [Google Scholar] [CrossRef]
- Ishikawa, M.; Aoyama, T.; Shibata, S.; Sone, T.; Miyoshi, H.; Watanabe, H.; Nakamura, M.; Morota, S.; Uchino, H.; Yoo, A.S.; et al. miRNA-Based rapid differentiation of purified neurons from hPSCs advancestowards quick screening for neuronal disease phenotypes In Vitro. Cells 2020, 9, 532. [Google Scholar] [CrossRef] [PubMed]
- Chavali, V.R.M.; Haider, N.; Rathi, S.; Vrathasha, V.; Alapati, T.; He, J.; Gill, K.; Nikonov, R.; Duong, T.T.; McDougald, D.S.; et al. Dual SMAD inhibition and Wnt inhibition enable efficient and reproducible differentiations of induced pluripotent stem cells into retinal ganglion cells. Sci. Rep. 2020, 10, 11828. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Xiao, Q.; Yip, H.K. Regulation of retinal progenitor cell differentiation by bone morphogenetic protein 4 is mediated by the Smad/Id cascade. Investig. Opthalmol. Vis. Sci. 2010, 51, 3764–3773. [Google Scholar] [CrossRef] [PubMed]
- Chichagova, V.; Hilgen, G.; Ghareeb, A.; Georgiou, M.; Carter, M.; Sernagor, E.; Lako, M.; Armstrong, L. Human iPSC differentiation to retinal organoids in response to IGF1 and BMP4 activation is line- and method-dependent. Stem Cells 2019, 38, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Capowski, E.E.; Samimi, K.; Mayerl, S.J.; Phillips, M.J.; Pinilla, I.; Howden, S.E.; Saha, J.; Jansen, A.D.; Edwards, K.L.; Jager, L.D.; et al. Reproducibility and staging of 3D human retinal organoids across multiple pluripotent stem cell lines. Development 2019, 146, dev171686. [Google Scholar] [CrossRef]
- Quillien, A.; Blanco-Sanchez, B.; Halluin, C.; Moore, J.C.; Lawson, N.; Blader, P.; Cau, E. BMP signaling orchestrates photoreceptor specification in the zebrafish pineal gland in collaboration with Notch. Development 2011, 138, 2293–2302. [Google Scholar] [CrossRef]
- Ueki, Y.; Wilken, M.S.; Cox, K.E.; Chipman, L.; Bermingham-McDonogh, O.; Reh, T.A. A transient wave of BMP signaling in the retina is necessary for Müller glial differentiation. Development 2015, 142, 533–543. [Google Scholar] [CrossRef]
- Luo, G.; Hofmann, C.; Bronckers, A.; Sohocki, M.; Bradley, A.; Karsenty, G. BMP-7 is an inducer of nephrogenesis, and is also required for eye development and skeletal patterning. Genes Dev. 1995, 9, 2808–2820. [Google Scholar] [CrossRef]
- Ochi, H.; Ogino, H.; Kageyama, Y.; Yasuda, K. The stability of the lens-specific Maf protein is regulated by Fibroblast Growth Factor (FGF)/ERK signaling in lens fiber differentiation. J. Biol. Chem. 2003, 278, 537–544. [Google Scholar] [CrossRef]
- Pachenari, N.; Kiani, S.; Javan, M. Inhibition of glycogen synthase kinase 3 increased subventricular zone stem cells proliferation. Biomed. Pharmacother. 2017, 93, 1074–1082. [Google Scholar] [CrossRef]
- Kisseleff, E.; Vigouroux, R.J.; Hottin, C.; Lourdel, S.; Shah, P.; Chédotal, A.; Perron, M.; Swaroop, A.; Roger, J.E. Glycogen Synthase Kinase 3 regulates the genesis of the rare displaced ganglion cell retinal subtype. bioRxiv 2021. [Google Scholar] [CrossRef]
- Huang, J.; Guo, X.; Li, W.; Zhang, H. Activation of Wnt/β-catenin signalling via GSK3 inhibitors direct differentiation of human adipose stem cells into functional hepatocytes. Sci. Rep. 2017, 7, 40716. [Google Scholar] [CrossRef]
- Aikin, R.A.; Ayers, K.L.; Thérond, P.P. The role of kinases in the Hedgehog signalling pathway. EMBO Rep. 2008, 9, 330–336. [Google Scholar] [CrossRef]
- Kaidanovich-Beilin, O.; Woodgett, J.R. GSK-3: Functional insights from cell biology and animal models. Front. Mol. Neurosci. 2011, 4, 40. [Google Scholar] [CrossRef]
- Ekim, W.-Y.; Snider, W.D. Functions of GSK-3 signaling in development of the nervous system. Front. Mol. Neurosci. 2011, 4, 44. [Google Scholar] [CrossRef]
- Marchena, M.; Villarejo-Zori, B.; Zaldivar-Diez, J.; Palomo, V.; Gil, C.; Sánchez, C.H.; Martínez, A.; De La Rosa, E.J. Small molecules targeting glycogen synthase kinase 3 as potential drug candidates for the treatment of retinitis pigmentosa. J. Enzym. Inhib. Med. Chem. 2017, 32, 522–526. [Google Scholar] [CrossRef]
- Moon, R.T. The promise and perils of Wnt signaling through beta -Catenin. Science 2002, 296, 1644–1646. [Google Scholar] [CrossRef]
- Clark, C.E.; Nourse, C.C.; Cooper, H.M. The tangled web of non-canonical Wnt signalling in neural migration. Neurosignals 2012, 20, 202–220. [Google Scholar] [CrossRef]
- Cerpa, W.; Gambrill, A.; Inestrosa, N.C.; Barria, A. Regulation of NMDA-receptor synaptic transmission by Wnt signaling. J. Neurosci. 2011, 31, 9466–9471. [Google Scholar] [CrossRef]
- Ciani, L.; Salinas, P.C. WNTS in the vertebrate nervous system: From patterning to neuronal connectivity. Nat. Rev. Neurosci. 2005, 6, 351–362. [Google Scholar] [CrossRef]
- Carter, M.; Chen, X.; Slowinska, B.; Minnerath, S.; Glickstein, S.; Shi, L.; Campagne, F.; Weinstein, H.; Ross, M.E. Crooked tail (Cd) model of human folate-responsive neural tube defects is mutated in Wnt coreceptor lipoprotein receptor-related protein 6. Proc. Natl. Acad. Sci. USA 2005, 102, 12843–12848. [Google Scholar] [CrossRef]
- Shi, D.-L.; Goisset, C.; Boucaut, J.-C. Expression of Xfz3, a Xenopus frizzled family member, is restricted to the early nervous system. Mech. Dev. 1998, 70, 35–47. [Google Scholar] [CrossRef]
- Rasmussen, J.T.; Deardorff, M.A.; Tan, C.; Rao, M.S.; Klein, P.S.; Vetter, M.L. Regulation of eye development by frizzled signaling in Xenopus. Proc. Natl. Acad. Sci. USA 2001, 98, 3861–3866. [Google Scholar] [CrossRef] [PubMed]
- Cavodeassi, F.; Carreira-Barbosa, F.; Young, R.M.; Concha, M.L.; Allende, M.L.; Houart, C.; Tada, M.; Wilson, S.W. Early stages of Zebrafish eye formation require the coordinated activity of Wnt11, Fz5, and the Wnt/β-Catenin pathway. Neuron 2005, 47, 43–56. [Google Scholar] [CrossRef]
- Liu, W.; Lagutin, O.; Swindell, E.; Jamrich, M.; Oliver, G. Neuroretina specification in mouse embryos requires Six3-mediated suppression of Wnt8b in the anterior neural plate. J. Clin. Investig. 2010, 120, 3568–3577. [Google Scholar] [CrossRef]
- Tao, Y.; Cao, J.; Li, M.; Hoffmann, B.; Xu, K.; Chen, J.; Lu, X.; Guo, F.; Li, X.; Phillips, M.J.; et al. PAX 6D instructs neural retinal specification from human embryonic stem cell-derived neuroectoderm. EMBO Rep. 2020, 21, e50000. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, M.A.; Yi, H.; Nakamura, R.E.I.; Hackam, A.S. The Wnt signaling pathway protects retinal ganglion Cell 5 (RGC-5) cells from elevated pressure. Cell. Mol. Neurobiol. 2010, 31, 163–173. [Google Scholar] [CrossRef]
- Patel, A.K.; Surapaneni, K.; Yi, H.; Nakamura, R.E.I.; Karli, S.Z.; Syeda, S.; Lee, T.; Hackam, A.S. Activation of Wnt/β-catenin signaling in Muller glia protects photoreceptors in a mouse model of inherited retinal degeneration. Neuropharmacology 2015, 91, 1–12. [Google Scholar] [CrossRef]
- Liu, B.; Hunter, D.J.; Rooker, S.; Chan, A.; Paulus, Y.; Leucht, P.; Nusse, Y.; Nomoto, H.; Helms, J.A. Wnt signaling promotes Müller cell proliferation and survival after injury. Investig. Opthalmol. Vis. Sci. 2013, 54, 444–453. [Google Scholar] [CrossRef]
- Aparicio, J.; Hopp, H.; Choi, A.; Comar, J.M.; Liao, V.; Harutyunyan, N.; Lee, T. Temporal expression of CD184(CXCR4) and CD171(L1CAM) identifies distinct early developmental stages of human retinal ganglion cells in embryonic stem cell derived retina. Exp. Eye Res. 2017, 154, 177–189. [Google Scholar] [CrossRef]
- Menendez, L.; Yatskievych, T.A.; Antin, P.; Dalton, S. Wnt signaling and a Smad pathway blockade direct the differentiation of human pluripotent stem cells to multipotent neural crest cells. Proc. Natl. Acad. Sci. USA 2011, 108, 19240–19245. [Google Scholar] [CrossRef]
- Chambers, S.M.; Mica, Y.; Lee, G.; Studer, L.; Tomishima, M.J. Dual-SMAD inhibition/WNT activation-based methods to induce neural crest and derivatives from human pluripotent stem cells. Methods Mol. Biol. 2013, 1307, 329–343. [Google Scholar] [CrossRef]
- Zhou, S.; Flamier, A.; Abdouh, M.; Tétreault, N.; Barabino, A.; Wadhwa, S.; Bernier, G. Differentiation of human embryonic stem cells into cone photoreceptors through simultaneous inhibition of BMP, TGFβ and Wnt signaling. Development 2015, 142, 3294–3306. [Google Scholar] [CrossRef]
- Hägglund, A.-C.; Berghard, A.; Carlsson, L. Canonical Wnt/β-Catenin signalling is essential for optic cup formation. PLoS ONE 2013, 8, e81158. [Google Scholar] [CrossRef]
- Fujimura, N. WNT/β-Catenin signaling in vertebrate eye development. Front. Cell Dev. Biol. 2016, 4, 138. [Google Scholar] [CrossRef]
- Sugiyama, Y.; Lovicu, F.J.; McAvoy, J.W. Planar cell polarity in the mammalian eye lens. Organogenesis 2011, 7, 191–201. [Google Scholar] [CrossRef]
- Aguirre, A.; Rubio, M.E.; Gallo, V. Notch and EGFR pathway interaction regulates neural stem cell number and self-renewal. Nat. Cell Biol. 2010, 467, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Grego-Bessa, J.; Luna-Zurita, L.; del Monte, G.; Bolós, V.; Melgar, P.; Arandilla, A.; Garratt, A.; Zang, H.; Mukouyama, Y.-S.; Chen, H.; et al. Notch signaling is essential for ventricular chamber development. Dev. Cell 2007, 12, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, L.C.; Stanger, B.Z.; Kwan, K.M.; Melton, D.A. Notch signaling controls multiple steps of pancreatic differentiation. Proc. Natl. Acad. Sci. USA 2003, 100, 14920–14925. [Google Scholar] [CrossRef]
- Stupnikov, M.R.; Yang, Y.; Mori, M.; Lu, J.; Cardoso, W.V. Jagged and Delta-like ligands control distinct events during airway progenitor cell differentiation. eLife 2019, 8, e50487. [Google Scholar] [CrossRef]
- Van Tetering, G.; van Diest, P.; Verlaan, I.; van der Wall, E.; Kopan, R.; Vooijs, M. Metalloprotease ADAM10 is required for notch1 site 2 cleavage. J. Biol. Chem. 2009, 284, 31018–31027. [Google Scholar] [CrossRef]
- Mills, E.A.; Goldman, D. The regulation of notch signaling in retinal development and regeneration. Curr. Pathobiol. Rep. 2017, 5, 323–331. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Liao, W.; Lou, Y.-L.; Li, Q.; Hu, B.; Wang, Y.; Deng, Z.-F. Inhibition of Notch signaling facilitates the differentiation of human-induced pluripotent stem cells into neural stem cells. Mol. Cell. Biochem. 2014, 395, 291–298. [Google Scholar] [CrossRef]
- Banda, E.; McKinsey, A.; Germain, N.; Carter, J.; Anderson, N.C.; Grabel, L. Cell polarity and neurogenesis in embryonic stem cell-derived neural rosettes. Stem Cells Dev. 2015, 24, 1022–1033. [Google Scholar] [CrossRef]
- Toonen, J.A.; Ronchetti, A.; Sidjanin, D.J. A Disintegrin and metalloproteinase10 (ADAM10) regulates NOTCH signaling during early retinal development. PLoS ONE 2016, 11, e0156184. [Google Scholar] [CrossRef]
- Jadhav, A.P.; Cho, S.-H.; Cepko, C.L. Notch activity permits retinal cells to progress through multiple progenitor states and acquire a stem cell property. Proc. Natl. Acad. Sci. USA 2006, 103, 18998–19003. [Google Scholar] [CrossRef]
- Nelson, B.R.; Gumuscu, B.; Hartman, B.H.; Reh, T.A. Notch activity is downregulated just prior to retinal ganglion cell differentiation. Dev. Neurosci. 2006, 28, 128–141. [Google Scholar] [CrossRef]
- Riesenberg, A.N.; Conley, K.W.; Le, T.T.; Brown, N.L. Separate and coincident expression ofHes1andHes5in the developing mouse eye. Dev. Dyn. 2018, 247, 212–221. [Google Scholar] [CrossRef]
- Silva, A.O.; Ercole, C.E.; McLoon, S.C. Regulation of ganglion cell production by notch signaling during retinal development. J. Neurobiol. 2003, 54, 511–524. [Google Scholar] [CrossRef]
- Riazifar, H.; Jia, Y.; Chen, J.; Lynch, G.; Huang, T. Chemically induced specification of retinal ganglion cells from human embryonic and induced pluripotent stem cells. Stem Cells Transl. Med. 2014, 3, 424–432. [Google Scholar] [CrossRef]
- Jadhav, A.P.; Mason, H.A.; Cepko, C.L. Notch 1 inhibits photoreceptor production in the developing mammalian retina. Development 2006, 133, 913–923. [Google Scholar] [CrossRef]
- Mizeracka, K.; Trimarchi, J.; Stadler, M.B.; Cepko, C.L. Analysis of gene expression in wild-type and Notch1 mutant retinal cells by single cell profiling. Dev. Dyn. 2013, 242, 1147–1159. [Google Scholar] [CrossRef]
- Mochizuki, Y.; Iida, A.; Lyons, E.; Kageyama, R.; Nakauchi, H.; Murakami, A.; Watanabe, S. Use of cell type-specific transcriptome to identify genes specifically involved in Müller glia differentiation during retinal development. Dev. Neurobiol. 2013, 74, 426–437. [Google Scholar] [CrossRef]
- Teotia, P.; Chopra, D.A.; Dravid, S.M.; Van Hook, M.J.; Qiu, F.; Morrison, J.; Rizzino, A.; Ahmad, I. Generation of functional human retinal ganglion cells with target specificity from pluripotent stem cells by chemically defined recapitulation of developmental mechanism. Stem Cells 2017, 35, 572–585. [Google Scholar] [CrossRef]
- Stanton, B.Z.; Peng, L.F. Small-molecule modulators of the Sonic Hedgehog signaling pathway. Mol. BioSyst. 2010, 6, 44–54. [Google Scholar] [CrossRef]
- Gao, L.; Chen, X.; Zeng, Y.; Li, Q.; Zou, T.; Chen, S.; Wu, Q.; Fu, C.; Xu, H.; Yin, Z.Q. Intermittent high oxygen influences the formation of neural retinal tissue from human embryonic stem cells. Sci. Rep. 2016, 6, 29944. [Google Scholar] [CrossRef]
- Völkner, M.; Zschätzsch, M.; Rostovskaya, M.; Overall, R.; Busskamp, V.; Anastassiadis, K.; Karl, M.O. Retinal organoids from pluripotent stem cells efficiently recapitulate retinogenesis. Stem Cell Rep. 2016, 6, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Worthington, K.S.; Wiley, L.A.; Kaalberg, E.E.; Collins, M.M.; Mullins, R.F.; Stone, E.M.; Tucker, B.A. Two-photon polymerization for production of human iPSC-derived retinal cell grafts. Acta Biomater. 2017, 55, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.K.; Yung, J.S.Y.; Choy, K.W.; Cao, D.; Leung, C.K.S.; Cheung, H.S.; Pang, C.P. Transdifferentiation of periodontal ligament-derived stem cells into retinal ganglion-like cells and its microRNA signature. Sci. Rep. 2015, 5, 16429. [Google Scholar] [CrossRef] [PubMed]
- Inman, G.; Nicolás, F.J.; Callahan, J.F.; Harling, J.D.; Gaster, L.M.; Reith, A.D.; Laping, N.J.; Hill, C. SB-431542 is a potent and specific inhibitor of transforming growth Factor-β superfamily Type I activin receptor-like Kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol. Pharmacol. 2002, 62, 65–74. [Google Scholar] [CrossRef]
- Madhu, V.; Dighe, A.S.; Cui, Q.; Deal, D.N. Dual inhibition of activin/Nodal/TGF-βand BMP signaling pathways by SB431542 and dorsomorphin induces neuronal differentiation of human adipose derived stem cells. Stem Cells Int. 2016, 2016, 1035374. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, X.-J.; Renier, N.; Wu, Z.; Atkin, T.; Sun, Z.; Ozair, M.Z.; Tchieu, J.; Zimmer, B.; Fattahi, F.; et al. Combined small-molecule inhibition accelerates the derivation of functional cortical neurons from human pluripotent stem cells. Nat. Biotechnol. 2017, 35, 154–163. [Google Scholar] [CrossRef]
- Browne, A.W.; Arnesano, C.; Harutyunyan, N.; Khuu, T.; Martinez, J.C.; Pollack, H.A.; Koos, D.S.; Lee, T.C.; Fraser, S.E.; Moats, R.A.; et al. Structural and functional characterization of human stem-cell-derived retinal organoids by live imaging. Investig. Opthalmol. Vis. Sci. 2017, 58, 3311–3318. [Google Scholar] [CrossRef]
- Döpper, H.; Menges, J.; Bozet, M.; Brenzel, A.; Lohmann, D.; Steenpass, L.; Kanber, D. Differentiation protocol for 3D retinal organoids, immunostaining and signal quantitation. Curr. Protoc. Stem Cell Biol. 2020, 55, e120. [Google Scholar] [CrossRef]
- Zhu, J.; Reynolds, J.; Garcia, T.; Cifuentes, H.; Chew, S.; Zeng, X.; Lamba, D.A. Generation of transplantable retinal photoreceptors from a current good manufacturing practice-manufactured human induced pluripotent stem cell line. Stem Cells Transl. Med. 2017, 7, 210–219. [Google Scholar] [CrossRef]
- Popovic, N.; Hooker, E.; Barabino, A.; Flamier, A.; Provost, F.; Buscarlet, M.; Bernier, G.; Larrivée, B. COCO/DAND5 inhibits developmental and pathological ocular angiogenesis. EMBO Mol. Med. 2021, 13, e12005. [Google Scholar] [CrossRef]
- Bates, T.J.D.; Vonica, A.; Heasman, J.; Brivanlou, A.H.; Bell, E. Coco regulates dorsoventral specification of germ layers via inhibition of TGFβ signalling. Development 2013, 140, 4177–4181. [Google Scholar] [CrossRef]
- Schweickert, A.; Vick, P.; Getwan, M.; Weber, T.; Schneider, I.; Eberhardt, M.; Beyer, T.; Pachur, A.; Blum, M. The nodal inhibitor coco is a critical target of leftward flow in Xenopus. Curr. Biol. 2010, 20, 738–743. [Google Scholar] [CrossRef]
- Pan, D.; Xia, X.-X.; Zhou, H.; Jin, S.-Q.; Lu, Y.-Y.; Liu, H.; Gao, M.-L.; Jin, Z.-B. COCO enhances the efficiency of photoreceptor precursor differentiation in early human embryonic stem cell-derived retinal organoids. Stem Cell Res. Ther. 2020, 11, 366. [Google Scholar] [CrossRef]
- Deglincerti, A.; Haremaki, T.; Warmflash, A.; Sorre, B.; Brivanlou, A.H. Coco is a dual-activity modulator of TGF-β signaling. Development 2015, 142, 2678–2685. [Google Scholar] [CrossRef]
- Blázquez-Medela, A.M.; Jumabay, M.; Boström, K.I. Beyond the bone: Bone morphogenetic protein signaling in adipose tissue. Obes. Rev. 2019, 20, 648–658. [Google Scholar] [CrossRef]
- Merino, R.; Macias, D.; Gañan, Y.; Economides, A.; Wang, X.; Wu, Q.; Stahl, N.; Sampath, K.; Varona, P.; Hurle, J.M. Expression and function ofGdf-5 during digit skeletogenesis in the embryonic chick leg bud. Dev. Biol. 1999, 206, 33–45. [Google Scholar] [CrossRef]
- Chang, C.; Hemmati-Brivanlou, A. Xenopus GDF6, a new antagonist of noggin and a partner of BMPs. Development 1999, 126, 3347–3357. [Google Scholar] [CrossRef]
- Messina, A.; Lan, L.; Incitti, T.; Bozza, A.; Andreazzoli, M.; Vignali, R.; Cremisi, F.; Bozzi, Y.; Casarosa, S. Noggin-Mediated retinal induction reveals a novel interplay between bone morphogenetic protein inhibition, transforming growth Factor β, and Sonic hedgehog signaling. Stem Cells 2015, 33, 2496–2508. [Google Scholar] [CrossRef]
- Lamba, D.A.; McUsic, A.; Hirata, R.K.; Wang, P.-R.; Russell, D.; Reh, T.A. Generation, purification and transplantation of photoreceptors derived from human induced pluripotent stem cells. PLoS ONE 2010, 5, e8763. [Google Scholar] [CrossRef]
- Tucker, B.A.; Mullins, R.F.; Streb, L.M.; Anfinson, K.; Eyestone, M.E.; Kaalberg, E.; Riker, M.J.; Drack, A.V.; Braun, T.A.; Stone, E.M. Patient-specific iPSC-derived photoreceptor precursor cells as a means to investigate retinitis pigmentosa. eLife 2013, 2, e00824. [Google Scholar] [CrossRef]
- Kim, D.-S.; Lee, J.S.; Leem, J.W.; Huh, Y.J.; Kim, J.Y.; Kim, H.-S.; Park, I.-H.; Daley, G.Q.; Hwang, D.-Y.; Kim, N.-W. Robust enhancement of neural differentiation from human ES and iPS cells regardless of their innate difference in differentiation propensity. Stem Cell Rev. Rep. 2010, 6, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Morizane, A.; Doi, D.; Kikuchi, T.; Nishimura, K.; Takahashi, J. Small-molecule inhibitors of bone morphogenic protein and activin/nodal signals promote highly efficient neural induction from human pluripotent stem cells. J. Neurosci. Res. 2011, 89, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.B.; Hong, C.C.; Sachidanandan, C.; Babitt, J.L.; Deng, D.Y.; A Hoyng, S.; Lin, H.Y.; Bloch, K.D.; Peterson, R.T. Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism. Nat. Chem. Biol. 2008, 4, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Boergermann, J.; Kopf, J.; Yu, P.; Knaus, P. Dorsomorphin and LDN-193189 inhibit BMP-mediated Smad, p38 and Akt signalling in C2C12 cells. Int. J. Biochem. Cell Biol. 2010, 42, 1802–1807. [Google Scholar] [CrossRef] [PubMed]
- Vogt, J.; Traynor, R.; Sapkota, G.P. The specificities of small molecule inhibitors of the TGFß and BMP pathways. Cell. Signal. 2011, 23, 1831–1842. [Google Scholar] [CrossRef]
- Cuny, G.D.; Yu, P.B.; Laha, J.K.; Xing, X.; Liu, J.-F.; Lai, C.S.; Deng, D.Y.; Sachidanandan, C.; Bloch, K.D.; Peterson, R.T. Structure–activity relationship study of bone morphogenetic protein (BMP) signaling inhibitors. Bioorganic Med. Chem. Lett. 2008, 18, 4388–4392. [Google Scholar] [CrossRef]
- Kuwahara, A.; Ozone, C.; Nakano, T.; Saito, K.; Eiraku, M.; Sasai, Y. Generation of a ciliary margin-like stem cell niche from self-organizing human retinal tissue. Nat. Commun. 2015, 6, 6286. [Google Scholar] [CrossRef]
- Sluch, V.M.; Chamling, X.; Liu, M.M.; Berlinicke, C.A.; Cheng, J.; Mitchell, K.L.; Welsbie, D.S.; Zack, D.J. Enhanced stem cell differentiation and immunopurification of genome engineered human retinal ganglion cells. Stem Cells Transl. Med. 2017, 6, 1972–1986. [Google Scholar] [CrossRef]
- Kobayashi, W.; Onishi, A.; Tu, H.-Y.; Takihara, Y.; Matsumura, M.; Tsujimoto, K.; Inatani, M.; Nakazawa, T.; Takahashi, M. Culture systems of dissociated mouse and human pluripotent stem cell–derived retinal ganglion cells purified by two-step immunopanning. Investig. Opthalmol. Vis. Sci. 2018, 59, 776–787. [Google Scholar] [CrossRef]
- Bronstein, R.; Capowski, E.E.; Mehrotra, S.; Jansen, A.D.; Navarro-Gomez, D.; Maher, M.; Place, E.; Sangermano, R.; Bujakowska, K.M.; Gamm, D.M.; et al. A combined RNA-seq and whole genome sequencing approach for identification of non-coding pathogenic variants in single families. Hum. Mol. Genet. 2020, 29, 967–979. [Google Scholar] [CrossRef]
- Fligor, C.M.; Huang, K.-C.; Lavekar, S.S.; VanderWall, K.B.; Meyer, J.S. Differentiation of retinal organoids from human pluripotent stem cells. Methods Cell Biol. 2020, 159, 279–302. [Google Scholar] [CrossRef]
- Chen, B.; Dodge, M.E.; Tang, W.; Lu, J.; Ma, Z.; Fan, C.-W.; Wei, S.; Hao, W.; Kilgore, J.; Williams, N.S.; et al. Small molecule–mediated disruption of Wnt-dependent signaling in tissue regeneration and cancer. Nat. Chem. Biol. 2009, 5, 100–107. [Google Scholar] [CrossRef]
- Kegeles, E.; Perepelkina, T.; Baranov, P. Semi-Automated approach for retinal tissue differentiation. Transl. Vis. Sci. Technol. 2020, 9, 24. [Google Scholar] [CrossRef]
- Sakata, T.; Chen, J.K. Chemical ‘Jekyll and Hyde’s: Small-molecule inhibitors of developmental signaling pathways. Chem. Soc. Rev. 2011, 40, 4318–4331. [Google Scholar] [CrossRef]
- Meyer, J.S.; Howden, S.E.; Wallace, K.A.; Verhoeven, A.D.; Wright, L.S.; Capowski, E.E.; Pinilla, I.; Martin, J.M.; Tian, S.; Stewart, R.; et al. Optic vesicle-like structures derived from human pluripotent stem cells facilitate a customized approach to retinal disease treatment. STEM CELLS 2011, 29, 1206–1218. [Google Scholar] [CrossRef]
- Semënov, M.V.; Tamai, K.; Brott, B.K.; Kühl, M.; Sokol, S.; He, X. Head inducer Dickkopf-1 is a ligand for Wnt coreceptor LRP6. Curr. Biol. 2001, 11, 951–961. [Google Scholar] [CrossRef]
- Deng, F.; Chen, M.; Liu, Y.; Hu, H.; Xiong, Y.; Xu, C.; Liu, Y.; Li, K.; Zhuang, J.; Ge, J. Stage-specific differentiation of iPSCs toward retinal ganglion cell lineage. Mol. Vis. 2016, 22, 536–547. [Google Scholar]
- Gill, K.P.; Hung, S.; Sharov, A.; Lo, C.Y.; Needham, K.; Lidgerwood, G.E.; Jackson, S.; Crombie, D.E.; Nayagam, B.; Cook, A.L.; et al. Enriched retinal ganglion cells derived from human embryonic stem cells. Sci. Rep. 2016, 6, 30552. [Google Scholar] [CrossRef]
- Shao, J.; Zhou, P.-Y.; Peng, G.-H. Experimental study of the biological properties of human embryonic stem cell–derived retinal progenitor cells. Sci. Rep. 2017, 7, 42363. [Google Scholar] [CrossRef]
- Leach, L.L.; Buchholz, D.E.; Nadar, V.P.; Lowenstein, S.E.; Clegg, D.O. Canonical/β-Catenin Wnt pathway activation improves retinal pigmented epithelium derivation from human embryonic stem cells. Investig. Opthalmol. Vis. Sci. 2015, 56, 1002–1013. [Google Scholar] [CrossRef]
- Nelson, B.R.; Hartman, B.H.; Georgi, S.A.; Lan, M.S.; Reh, T.A. Transient inactivation of Notch signaling synchronizes differentiation of neural progenitor cells. Dev. Biol. 2007, 304, 479–498. [Google Scholar] [CrossRef]
- Kruczek, K.; Cordero, A.G.; Goh, D.; Naeem, A.; Jonikas, M.; Blackford, S.; Kloc, M.; Duran, Y.; Georgiadis, A.; Sampson, R.; et al. Differentiation and transplantation of embryonic stem cell-derived cone photoreceptors into a mouse model of end-stage retinal degeneration. Stem Cell Rep. 2017, 8, 1659–1674. [Google Scholar] [CrossRef]
- Reichman, S.; Terray, A.; Slembrouck, A.; Nanteau, C.; Orieux, G.; Habeler, W.; Nandrot, E.F.; Sahel, J.; Monville, C.; Goureau, O. From confluent human iPS cells to self-forming neural retina and retinal pigmented epithelium. Proc. Natl. Acad. Sci. USA 2014, 111, 8518–8523. [Google Scholar] [CrossRef]
- Meyer, J.; Shearer, R.L.; Capowski, E.E.; Wright, L.S.; Wallace, K.A.; McMillan, E.L.; Zhang, S.-C.; Gamm, D.M. Modeling early retinal development with human embryonic and induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2009, 106, 16698–16703. [Google Scholar] [CrossRef] [PubMed]
- Hallam, D.; Hilgen, G.; Dorgau, B.; Zhu, L.; Yu, M.; Bojic, S.; Hewitt, P.; Schmitt, M.; Uteng, M.; Kustermann, S.; et al. Human-Induced pluripotent stem cells generate light responsive retinal organoids with variable and nutrient-dependent efficiency. Stem Cells 2018, 36, 1535–1551. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, M.; Chichagova, V.; Hilgen, G.; Dorgau, B.; Sernagor, E.; Armstrong, L.; Lako, M. Room temperature shipment does not affect the biological activity of pluripotent stem cell-derived retinal organoids. PLoS ONE 2020, 15, e0233860. [Google Scholar] [CrossRef]
- Lamba, D.A.; Karl, M.; Ware, C.B.; Reh, T.A. Efficient generation of retinal progenitor cells from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2006, 103, 12769–12774. [Google Scholar] [CrossRef]
- Mellough, C.; Collin, J.; Khazim, M.; White, K.; Sernagor, E.; Steel, D.; Lako, M. IGF-1 signaling plays an important role in the formation of three-dimensional laminated neural retina and other ocular structures from human embryonic stem cells. Stem Cells 2015, 33, 2416–2430. [Google Scholar] [CrossRef]
- Wagstaff, E.L.; Asbroek, A.L.M.A.T.; Brink, J.B.T.; Jansonius, N.M.; Bergen, A.A.B. An alternative approach to produce versatile retinal organoids with accelerated ganglion cell development. Sci. Rep. 2021, 11, 1101. [Google Scholar] [CrossRef]
- Slembrouck-Brec, A.; Rodrigues, A.; Rabesandratana, O.; Gagliardi, G.; Nanteau, C.; Fouquet, S.; Thuret, G.; Reichman, S.; Orieux, G.; Goureau, O. Reprogramming of adult retinal Müller glial cells into human-induced pluripotent stem cells as an efficient source of retinal cells. Stem Cells Int. 2019, 2019, 7858796. [Google Scholar] [CrossRef] [PubMed]
- Gamm, D.M.; Clark, E.; Capowski, E.E.; Singh, R. The role of FGF9 in the production of neural retina and RPE in a pluripotent stem cell model of early human retinal development. Am. J. Ophthalmol. 2019, 206, 113–131. [Google Scholar] [CrossRef]
- Cameron Cowan, A.S.; Renner, M.; De Gennaro, M.; Roma, G.; Nigsch, F.; Roska Correspondence, B.; Cowan, C.S.; Gross-Scherf, B.; Goldblum, D.; Hou, Y.; et al. Cell types of the human retina and its organoids at single-cell resolution. Cell 2020, 182, 1623–1640.e34. [Google Scholar] [CrossRef]
- Tso, A.; Ragi, S.D.; Costa, B.L.D.; Fehnel, A.; Li, Y.; Quinn, P.M.J. Molecular and therapeutic strategies for Retinitis Pigmentosa: Generation of Human iPSC-Derived Retinal Organoids for Assessment of AAV-Mediated Gene Delivery. In: Walker, J.M., ed. Methods Mol. Biol. 2021, in press. [Google Scholar]
- Nakano, T.; Ando, S.; Takata, N.; Kawada, M.; Muguruma, K.; Sekiguchi, K.; Saito, K.; Yonemura, S.; Eiraku, M.; Sasai, Y. Self-formation of optic cups and storable stratified neural retina from human ESCs. Cell Stem Cell 2012, 10, 771–785. [Google Scholar] [CrossRef]
- Zhong, X.; Gutierrez, C.; Xue, T.; Hampton, C.; Vergara, M.N.; Cao, L.-H.; Peters, A.; Park, T.S.; Zambidis, E.T.; Meyer, J.; et al. Generation of three-dimensional retinal tissue with functional photoreceptors from human iPSCs. Nat. Commun. 2014, 5, 4047. [Google Scholar] [CrossRef]
- Reichman, S.; Slembrouck, A.; Gagliardi, G.; Chaffiol, A.; Terray, A.; Nanteau, C.; Potey, A.; Belle, M.; Rabesandratana, O.; Duebel, J.; et al. Generation of storable retinal organoids and retinal pigmented epithelium from adherent human iPS cells in xeno-free and feeder-free conditions. Stem Cells 2017, 35, 1176–1188. [Google Scholar] [CrossRef]
- Slembrouck-Brec, A.; Nanteau, C.; Sahel, J.-A.; Goureau, O.; Reichman, S. Defined xeno-free and feeder-free culture conditions for the generation of human iPSC-derived retinal cell models. J. Vis. Exp. 2018, 139, e57795. [Google Scholar] [CrossRef]
- Rabesandratana, O.; Chaffiol, A.; Mialot, A.; Slembrouck-Brec, A.; Joffrois, C.; Nanteau, C.; Rodrigues, A.; Gagliardi, G.; Reichman, S.; Sahel, J.-A.; et al. Generation of a transplantable population of human iPSC-derived retinal ganglion cells. Front. Cell Dev. Biol. 2020, 8, 585675. [Google Scholar] [CrossRef]
- Achberger, K.; Probst, C.; Haderspeck, J.; Bolz, S.; Rogal, J.; Chuchuy, J.; Nikolova, M.; Cora, V.; Antkowiak, L.; Haq, W.; et al. Merging organoid and organ-on-a-chip technology to generate complex multi-layer tissue models in a human retina-on-a-chip platform. eLife 2019, 8, e46188. [Google Scholar] [CrossRef]
- Akhtar, T.; Xie, H.; Khan, M.I.; Zhao, H.; Bao, J.; Zhang, M.; Xue, T. Accelerated photoreceptor differentiation of hiPSC-derived retinal organoids by contact co-culture with retinal pigment epithelium. Stem Cell Res. 2019, 39, 101491. [Google Scholar] [CrossRef]
- Arno, G.; Agrawal, S.A.; Eblimit, A.; Bellingham, J.; Xu, M.; Wang, F.; Chakarova, C.; Parfitt, D.A.; Lane, A.; Burgoyne, T.; et al. Mutations in REEP6 cause autosomal-recessive retinitis pigmentosa. Am. J. Hum. Genet. 2016, 99, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Chichagova, V.; Dorgau, B.; Felemban, M.; Georgiou, M.; Armstrong, L.; Lako, M. Differentiation of retinal organoids from human pluripotent stem cells. Curr. Protoc. Stem Cell Biol. 2019, 50, e95. [Google Scholar] [CrossRef] [PubMed]
- Collin, J.; Queen, R.; Zerti, D.; Dorgau, B.; Hussain, R.; Coxhead, J.; Cockell, S.; Lako, M. Deconstructing retinal organoids: Single cell RNA-Seq reveals the cellular components of human pluripotent stem cell-derived retina. Stem Cells 2019, 37, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Collin, J.; Zerti, D.; Queen, R.; Santos-Ferreira, T.; Bauer, R.; Coxhead, J.; Hussain, R.; Steel, D.; Mellough, C.; Ader, M.; et al. CRX expression in pluripotent stem cell-derived photoreceptors marks a transplantable subpopulation of early cones. Stem Cells 2019, 37, 609–622. [Google Scholar] [CrossRef]
- Cora, V.; Haderspeck, J.; Antkowiak, L.; Mattheus, U.; Neckel, P.H.; Mack, A.F.; Bolz, S.; Ueffing, M.; Pashkovskaia, N.; Achberger, K.; et al. A cleared view on retinal organoids. Cells 2019, 8, 391. [Google Scholar] [CrossRef]
- Corral-Serrano, J.C.; Lamers, I.J.C.; Van Reeuwijk, J.; Duijkers, L.; Hoogendoorn, A.D.M.; Yildirim, A.; Argyrou, N.; Ruigrok, R.A.A.; Letteboer, S.J.F.; Butcher, R.; et al. PCARE and WASF3 regulate ciliary F-actin assembly that is required for the initiation of photoreceptor outer segment disk formation. Proc. Natl. Acad. Sci. USA 2020, 117, 9922–9931. [Google Scholar] [CrossRef]
- Cuevas, E.; Holder, D.L.; AlShehri, A.H.; Tréguier, J.; Lakowski, J.; Sowden, J.C. NRL−/− gene edited human embryonic stem cells generate rod-deficient retinal organoids enriched in S-cone-like photoreceptors. Stem Cells 2021, 39, 414–428. [Google Scholar] [CrossRef]
- De Bruijn, S.E.; Fiorentino, A.; Ottaviani, D.; Fanucchi, S.; Melo, U.S.; Corral-Serrano, J.C.; Mulders, T.; Georgiou, M.; Rivolta, C.; Pontikos, N.; et al. Structural variants create new topological-associated domains and ectopic retinal enhancer-gene contact in dominant retinitis pigmentosa. Am. J. Hum. Genet. 2020, 107, 802–814. [Google Scholar] [CrossRef]
- Deng, W.-L.; Gao, M.-L.; Lei, X.-L.; Lv, J.-N.; Zhao, H.; He, K.-W.; Xia, X.-X.; Li, L.-Y.; Chen, Y.-C.; Li, Y.-P.; et al. Gene correction reverses ciliopathy and photoreceptor loss in iPSC-derived retinal organoids from retinitis pigmentosa patients. Stem Cell Rep. 2018, 10, 1267–1281. [Google Scholar] [CrossRef]
- Deng, X.; Iwagawa, T.; Fukushima, M.; Watanabe, S. Characterization of human-induced pluripotent stem cells carrying homozygous RB1 gene deletion. Genes Cells 2020, 25, 510–517. [Google Scholar] [CrossRef]
- Diakatou, M.; Dubois, G.; Erkilic, N.; Sanjurjo-Soriano, C.; Meunier, I.; Kalatzis, V. Allele-Specific knockout by CRISPR/Cas to treat autosomal dominant retinitis pigmentosa caused by the G56R mutation in NR2E3. Int. J. Mol. Sci. 2021, 22, 2607. [Google Scholar] [CrossRef]
- Dorgau, B.; Felemban, M.; Hilgen, G.; Kiening, M.; Zerti, D.; Hunt, N.C.; Doherty, M.; Whitfield, P.; Hallam, D.; White, K.; et al. Decellularised extracellular matrix-derived peptides from neural retina and retinal pigment epithelium enhance the expression of synaptic markers and light responsiveness of human pluripotent stem cell derived retinal organoids. Biomaterials 2019, 199, 63–75. [Google Scholar] [CrossRef]
- Dorgau, B.; Felemban, M.; Sharpe, A.; Bauer, R.; Hallam, D.; Steel, D.H.; Lindsay, S.; Mellough, C.; Lako, M. Laminin γ3 plays an important role in retinal lamination, photoreceptor organisation and ganglion cell differentiation. Cell Death Dis. 2018, 9, 615. [Google Scholar] [CrossRef]
- Dulla, K.; Aguila, M.; Lane, A.; Jovanovic, K.; Parfitt, D.A.; Schulkens, I.; Chan, H.L.; Schmidt, I.; Beumer, W.; Vorthoren, L.; et al. Splice-Modulating oligonucleotide QR-110 restores CEP290 mRNA and function in human c.2991+1655A>G LCA10 models. Mol. Ther. Nucleic Acids 2018, 12, 730–740. [Google Scholar] [CrossRef]
- Eade, K.; Giles, S.; Harkins-Perry, S.; Friedlander, M. Toxicity screens in human retinal organoids for pharmaceutical discovery. J. Vis. Exp. 2021, 169, e62269. [Google Scholar] [CrossRef]
- Eastlake, K.; Wang, W.; Jayaram, H.; Murray-Dunning, C.; Carr, A.-J.; Ramsden, C.M.; Vugler, A.; Gore, K.; Clemo, N.; Stewart, M.; et al. Phenotypic and functional characterization of Müller glia isolated from induced pluripotent stem cell-derived retinal organoids: Improvement of retinal ganglion cell function upon transplantation. Stem Cells Transl. Med. 2019, 8, 775–784. [Google Scholar] [CrossRef]
- Eldred, K.C.; Hadyniak, S.E.; Hussey, K.A.; Brenerman, B.; Zhang, P.-W.; Chamling, X.; Sluch, V.M.; Welsbie, D.S.; Hattar, S.; Taylor, J.; et al. Thyroid hormone signaling specifies cone subtypes in human retinal organoids. Science 2018, 362, eaau6348. [Google Scholar] [CrossRef]
- Felemban, M.; Dorgau, B.; Hunt, N.C.; Hallam, D.; Zerti, D.; Bauer, R.; Ding, Y.; Collin, J.; Steel, D.; Krasnogor, N.; et al. Extracellular matrix component expression in human pluripotent stem cell-derived retinal organoids recapitulates retinogenesis in vivo and reveals an important role for IMPG1 and CD44 in the development of photoreceptors and interphotoreceptor matrix. Acta Biomater. 2018, 74, 207–221. [Google Scholar] [CrossRef]
- Fligor, C.M.; Langer, K.B.; Sridhar, A.; Ren, Y.; Shields, P.K.; Edler, M.C.; Ohlemacher, S.K.; Sluch, V.M.; Zack, D.J.; Zhang, C.; et al. Three-Dimensional retinal organoids facilitate the investigation of retinal ganglion cell development, organization and neurite outgrowth from human pluripotent stem cells. Sci. Rep. 2018, 8, 14520. [Google Scholar] [CrossRef]
- Freude, K.K.; Saruhanian, S.; McCauley, A.; Paterson, C.; Odette, M.; Oostenink, A.; Hyttel, P.; Gillies, M.; Haukedal, H.; Kolko, M. Enrichment of retinal ganglion and Müller glia progenitors from retinal organoids derived from human induced pluripotent stem cells—Possibilities and current limitations. World J. Stem Cells 2020, 12, 1171–1183. [Google Scholar] [CrossRef]
- Gagliardi, G.; BEN M’Barek, K.; Chaffiol, A.; Slembrouck-Brec, A.; Conart, J.-B.; Nanteau, C.; Rabesandratana, O.; Sahel, J.-A.; Duebel, J.; Orieux, G.; et al. Characterization and transplantation of CD73-positive photoreceptors isolated from human iPSC-derived retinal organoids. Stem Cell Rep. 2018, 11, 665–680. [Google Scholar] [CrossRef]
- Gantner, M.L.; Eade, K.; Wallace, M.; Handzlik, M.K.; Fallon, R.; Trombley, J.; Bonelli, R.; Giles, S.; Harkins-Perry, S.; Heeren, T.; et al. Serine and lipid metabolism in macular disease and peripheral neuropathy. N. Engl. J. Med. 2019, 381, 1422–1433. [Google Scholar] [CrossRef]
- Gao, M.-L.; Lei, X.-L.; Han, F.; He, K.-W.; Jin, S.-Q.; Zhang, Y.-Y.; Jin, Z.-B. Patient-Specific retinal organoids recapitulate disease features of late-onset retinitis pigmentosa. Front. Cell Dev. Biol. 2020, 8, 128. [Google Scholar] [CrossRef]
- Garita-Hernandez, M.; Chaffiol, A.; Guibbal, L.; Routet, F.; Khabou, H.; Riancho, L.; Toualbi, L.; Picaud, S.; Sahel, J.-A.; Goureau, O.; et al. Control of microbial opsin expression in stem cell derived cones for improved outcomes in cell therapy. Front. Cell. Neurosci. 2021, 15, 648210. [Google Scholar] [CrossRef] [PubMed]
- Garita-Hernandez, M.; Guibbal, L.; Toualbi, L.; Routet, F.; Chaffiol, A.; Winckler, C.; Harinquet, M.; Robert, C.; Fouquet, S.; Bellow, S.; et al. Optogenetic light sensors in human retinal organoids. Front. Neurosci. 2018, 12, 789. [Google Scholar] [CrossRef]
- Garita-Hernandez, M.; Routet, F.; Guibbal, L.; Khabou, H.; Toualbi, L.; Riancho, L.; Reichman, S.; Duebel, J.; Sahel, J.-A.; Goureau, O.; et al. AAV-Mediated gene delivery to 3D retinal organoids derived from human induced pluripotent stem cells. Int. J. Mol. Sci. 2020, 21, 994. [Google Scholar] [CrossRef] [PubMed]
- Cordero, A.G.; Kruczek, K.; Naeem, A.; Fernando, M.; Kloc, M.; Ribeiro, J.F.C.; Goh, D.; Duran, Y.; Blackford, S.; Abelleira-Hervas, L.; et al. Recapitulation of human retinal development from human pluripotent stem cells generates transplantable populations of cone photoreceptors. Stem Cell Rep. 2017, 9, 820–837. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, P.; Ma, J.H.; Cui, Z.; Yu, Q.; Liu, S.; Xue, Y.; Zhu, D.; Cao, J.; Li, Z.; et al. Modeling retinitis pigmentosa: Retinal organoids generated from the iPSCs of a patient with the USH2A mutation show early developmental abnormalities. Front. Cell. Neurosci. 2019, 13, 361. [Google Scholar] [CrossRef]
- Hoshino, A.; Horvath, S.; Sridhar, A.; Chitsazan, A.; Reh, T.A. Synchrony and asynchrony between an epigenetic clock and developmental timing. Sci. Rep. 2019, 9, 3770. [Google Scholar] [CrossRef]
- Huang, K.-C.; Wang, M.-L.; Chen, S.-J.; Kuo, J.-C.; Wang, W.-J.; Nguyen, P.N.N.; Wahlin, K.J.; Lu, J.-F.; Tran, A.A.; Shi, M.; et al. Morphological and molecular defects in human three-dimensional retinal organoid model of X-linked juvenile retinoschisis. Stem Cell Rep. 2019, 13, 906–923. [Google Scholar] [CrossRef]
- Kaewkhaw, R.; Kaya, K.D.; Brooks, M.; Homma, K.; Zou, J.; Chaitankar, V.; Rao, M.; Swaroop, A. Transcriptome dynamics of developing photoreceptors in three-dimensional retina cultures recapitulates temporal sequence of human cone and rod differentiation revealing cell surface markers and gene networks. Stem Cells 2015, 33, 3504–3518. [Google Scholar] [CrossRef]
- Kallman, A.; Capowski, E.E.; Wang, J.; Kaushik, A.M.; Jansen, A.D.; Edwards, K.L.; Chen, L.; Berlinicke, C.A.; Phillips, M.J.; Pierce, E.A.; et al. Investigating cone photoreceptor development using patient-derived NRL null retinal organoids. Commun. Biol. 2020, 3, 82. [Google Scholar] [CrossRef]
- Kaya, K.D.; Chen, H.Y.; Brooks, M.J.; Kelley, R.A.; Shimada, H.; Nagashima, K.; De Val, N.; Drinnan, C.T.; Gieser, L.; Kruczek, K.; et al. Transcriptome-based molecular staging of human stem cell-derived retinal organoids uncovers accelerated photoreceptor differentiation by 9-cis retinal. Mol. Vis. 2019, 25, 663–678. [Google Scholar]
- Kelley, R.A.; Chen, H.Y.; Swaroop, A.; Li, T. Accelerated development of rod photoreceptors in retinal organoids derived from human pluripotent stem cells by supplementation with 9-cis retinal. STAR Protoc. 2020, 1, 100033. [Google Scholar] [CrossRef]
- Khabou, H.; Garita-Hernandez, M.; Chaffiol, A.; Reichman, S.; Jaillard, C.; Brazhnikova, E.; Bertin, S.; Forster, V.; Desrosiers, M.; Winckler, C.; et al. Noninvasive gene delivery to foveal cones for vision restoration. JCI Insight 2018, 3, e96029. [Google Scholar] [CrossRef]
- Khan, M.; Arno, G.; Fakin, A.; Parfitt, D.A.; Dhooge, P.P.; Albert, S.; Bax, N.M.; Duijkers, L.; Niblock, M.; Hau, K.L.; et al. Detailed phenotyping and therapeutic strategies for intronic ABCA4 variants in stargardt disease. Mol. Ther. Nucleic Acids 2020, 21, 412–427. [Google Scholar] [CrossRef]
- Kim, S.; Lowe, A.; Dharmat, R.; Lee, S.; Owen, L.A.; Wang, J.; Shakoor, A.; Li, Y.; Morgan, D.J.; Hejazi, A.A.; et al. Generation, transcriptome profiling, and functional validation of cone-rich human retinal organoids. Proc. Natl. Acad. Sci. USA 2019, 116, 10824–10833. [Google Scholar] [CrossRef]
- Kruczek, K.; Qu, Z.; Gentry, J.; Fadl, B.R.; Gieser, L.; Hiriyanna, S.; Batz, Z.; Samant, M.; Samanta, A.; Chu, C.J.; et al. Gene therapy of dominant CRX-leber congenital amaurosis using patient stem cell-derived retinal organoids. Stem Cell Rep. 2021, 16, 252–263. [Google Scholar] [CrossRef]
- Lai, H.I.A.M.; Chou, S.-J.; Chien, Y.; Tsai, P.-H.; Chien, C.-S.; Hsu, C.-C.; Jheng, Y.-C.; Wang, M.-L.; Chiou, S.-H.; Chou, Y.-B.; et al. Expression of endogenous angiotensin-converting enzyme 2 in human induced pluripotent stem cell-derived retinal organoids. Int. J. Mol. Sci. 2021, 22, 1320. [Google Scholar] [CrossRef]
- Lakowski, J.; Welby, E.; Budinger, D.; Di Marco, F.; Di Foggia, V.; Bainbridge, J.W.; Wallace, K.; Gamm, D.M.; Ali, R.R.; Sowden, J.C. Isolation of human photoreceptor precursors via a cell surface marker panel from stem cell-derived retinal organoids and fetal retinae. Stem Cells 2018, 36, 709–722. [Google Scholar] [CrossRef]
- Lam, P.T.; Gutierrez, C.; Del Rio-Tsonis, K.; Robinson, M.L. Generation of a retina reporter hiPSC line to label progenitor, ganglion, and photoreceptor cell types. Transl. Vis. Sci. Technol. 2020, 9, 21. [Google Scholar] [CrossRef]
- Lane, A.; Jovanovic, K.; Shortall, C.; Ottaviani, D.; Panes, A.B.; Schwarz, N.; Guarascio, R.; Hayes, M.J.; Palfi, A.; Chadderton, N.; et al. Modeling and rescue of RP2 retinitis pigmentosa using iPSC-derived retinal organoids. Stem Cell Rep. 2020, 15, 67–79. [Google Scholar] [CrossRef]
- Li, G.; Gao, G.; Wang, P.; Song, X.; Xu, P.; Xie, B.; Zhou, T.; Pan, G.; Peng, F.; Zhang, Q.; et al. Generation and characterization of induced pluripotent stem cells and retinal organoids from a Leber’s congenital amaurosis patient with novel RPE65 mutations. Front. Mol. Neurosci. 2019, 12, 212. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xie, B.; He, L.; Zhou, T.; Gao, G.; Liu, S.; Pan, G.; Ge, J.; Peng, F.; Zhong, X. Generation of retinal organoids with mature rods and cones from urine-derived human induced pluripotent stem cells. Stem Cells Int. 2018, 2018, 4968658. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-P.; Deng, W.-L.; Jin, Z.-B. Modeling retinitis pigmentosa through patient-derived retinal organoids. STAR Protoc. 2021, 2, 100438. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; McLelland, B.T.; Aramant, R.B.; Thomas, B.B.; Nistor, G.; Keirstead, H.S.; Seiler, M.J. Retina organoid transplants develop photoreceptors and improve visual function in RCS rats With RPE dysfunction. Investig. Opthalmol. Vis. Sci. 2020, 61, 34. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hua, Z.-Q.; Jin, Z.-B. Modeling human retinoblastoma using embryonic stem cell-derived retinal organoids. STAR Protoc. 2021, 2, 100444. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Zhang, Y.-Y.; Li, Y.-P.; Hua, Z.-Q.; Zhang, C.-J.; Wu, K.-C.; Yu, F.; Zhang, Y.; Su, J.; et al. Human embryonic stem cell-derived organoid retinoblastoma reveals a cancerous origin. Proc. Natl. Acad. Sci. USA 2020, 117, 33628–33638. [Google Scholar] [CrossRef]
- Liu, S.; Xie, B.; Song, X.; Zheng, D.; He, L.; Li, G.; Gao, G.; Peng, F.; Yu, M.; Ge, J.; et al. Self-Formation of RPE spheroids facilitates enrichment and expansion of hiPSC-derived RPE generated on retinal organoid induction platform. Investig. Opthalmol. Vis. Sci. 2018, 59, 5659–5669. [Google Scholar] [CrossRef]
- Lowe, A.; Harris, R.; Bhansali, P.; Cvekl, A.; Liu, W. Intercellular adhesion-dependent cell survival and rock-regulated actomyosin-driven forces mediate self-formation of a retinal organoid. Stem Cell Rep. 2016, 6, 743–756. [Google Scholar] [CrossRef]
- Lu, Y.; Shiau, F.; Yi, W.; Lu, S.; Wu, Q.; Pearson, J.D.; Kallman, A.; Zhong, S.; Hoang, T.; Zuo, Z.; et al. Single-Cell analysis of human retina identifies evolutionarily conserved and species-specific mechanisms controlling development. Dev. Cell 2020, 53, 473–491.e9. [Google Scholar] [CrossRef]
- Lukovic, D.; Castro, A.A.; Kaya, K.D.; Munezero, D.; Gieser, L.; Davó-Martínez, C.; Corton, M.; Cuenca, N.; Swaroop, A.; Ramamurthy, V.; et al. Retinal organoids derived from hiPSCs of an AIPL1-LCA patient maintain cytoarchitecture despite reduced levels of mutant AIPL1. Sci. Rep. 2020, 10, 5426. [Google Scholar] [CrossRef]
- Luo, Z.; Xu, C.; Li, K.; Xian, B.; Liu, Y.; Li, K.; Liu, Y.; Rong, H.; Tang, M.; Hu, D.; et al. Islet1 and Brn3 expression pattern study in human retina and hiPSC-derived retinal organoid. Stem Cells Int. 2019, 2019, 8786396. [Google Scholar] [CrossRef]
- Luo, Z.; Zhong, X.; Li, K.; Xie, B.; Liu, Y.; Ye, M.; Li, K.; Xu, C.; Ge, J. An optimized system for effective derivation of three-dimensional retinal tissue via Wnt signaling regulation. Stem Cells 2018, 36, 1709–1722. [Google Scholar] [CrossRef]
- Mao, X.; An, Q.; Xi, H.; Yang, X.-J.; Zhang, X.; Yuan, S.; Wang, J.; Hu, Y.; Liu, Q.; Fan, G. Single-Cell RNA sequencing of hESC-derived 3D retinal organoids reveals novel genes regulating RPC commitment in early human retinogenesis. Stem Cell Rep. 2019, 13, 747–760. [Google Scholar] [CrossRef]
- McLelland, B.T.; Lin, B.; Mathur, A.; Aramant, R.B.; Thomas, B.B.; Nistor, G.; Keirstead, H.S.; Seiler, M.J. Transplanted hESC-derived retina organoid sheets differentiate, integrate, and improve visual function in retinal degenerate rats. Investig. Opthalmol. Vis. Sci. 2018, 59, 2586–2603. [Google Scholar] [CrossRef]
- Mellough, C.B.; Collin, J.; Queen, R.; Hilgen, G.; Dorgau, B.; Zerti, D.; Felemban, M.; White, K.; Sernagor, E.; Lako, M. Systematic comparison of retinal organoid differentiation from human pluripotent stem cells reveals stage specific, cell line, and methodological differences. Stem Cells Transl. Med. 2019, 8, 694–706. [Google Scholar] [CrossRef]
- Ohlemacher, S.K.; Iglesias, C.L.; Sridhar, A.; Gamm, D.M.; Meyer, J.S. Generation of highly enriched populations of optic vesicle−like retinal cells from human pluripotent stem cells. Curr. Protoc. Stem Cell Biol. 2015, 32, 1H.8.1–1H.8.20. [Google Scholar] [CrossRef]
- Ohlemacher, S.K.; Sridhar, A.; Xiao, Y.; Hochstetler, A.E.; Sarfarazi, M.; Cummins, T.R.; Meyer, J.S. Stepwise differentiation of retinal ganglion cells from human pluripotent stem cells enables analysis of glaucomatous neurodegeneration. Stem Cells 2016, 34, 1553–1562. [Google Scholar] [CrossRef]
- Ovando-Roche, P.; West, E.L.; Branch, M.J.; Sampson, R.D.; Fernando, M.; Munro, P.; Georgiadis, A.; Rizzi, M.; Kloc, M.; Naeem, A.; et al. Use of bioreactors for culturing human retinal organoids improves photoreceptor yields. Stem Cell Res. Ther. 2018, 9, 156. [Google Scholar] [CrossRef]
- Parfitt, D.A.; Lane, A.; Ramsden, C.M.; Carr, A.-J.; Munro, P.M.; Jovanovic, K.; Schwarz, N.; Kanuga, N.; Muthiah, M.N.; Hull, S.; et al. Identification and correction of mechanisms underlying inherited blindness in human iPSC-derived optic cups. Cell Stem Cell 2016, 18, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, G.; Cora, V.; Swiersy, A.; Achberger, K.; Antkowiak, L.; Müller, B.; Wimmer, T.; Fraschka, S.A.-K.; Casadei, N.; Ueffing, M.; et al. Using transcriptomic analysis to assess double-strand break repair activity: Towards precise In Vivo genome editing. Int. J. Mol. Sci. 2020, 21, 1380. [Google Scholar] [CrossRef] [PubMed]
- Peskova, L.; Jurcikova, D.; Vanova, T.; Krivanek, J.; Capandova, M.; Sramkova, Z.; Sebestikova, J.; Kolouskova, M.; Kotasova, H.; Streit, L.; et al. miR-183/96/182 cluster is an important morphogenetic factor targeting PAX6 expression in differentiating human retinal organoids. Stem Cells 2020, 38, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.J.; Capowski, E.E.; Petersen, A.; Jansen, A.D.; Barlow, K.; Edwards, K.L.; Gamm, D.M. Generation of a rod-specific NRL reporter line in human pluripotent stem cells. Sci. Rep. 2018, 8, 2370. [Google Scholar] [CrossRef]
- Quinn, P.M.; Buck, T.M.; Mulder, A.A.; Ohonin, C.; Alves, C.H.; Vos, R.M.; Bialecka, M.; van Herwaarden, T.; van Dijk, E.H.; Talib, M.; et al. Human iPSC-derived retinas recapitulate the fetal CRB1 CRB2 complex formation and demonstrate that photoreceptors and Müller Glia are targets of AAV5. Stem Cell Rep. 2019, 12, 906–919. [Google Scholar] [CrossRef]
- Quinn, P.M.; Buck, T.M.; Ohonin, C.; Mikkers, H.M.M.; Wijnholds, J. Production of iPS-derived human retinal organoids for use in transgene expression assays. Adv. Struct. Saf. Stud. 2017, 1715, 261–273. [Google Scholar] [CrossRef]
- Regent, F.; Chen, H.Y.; A Kelley, R.; Qu, Z.; Swaroop, A.; Li, T. A simple and efficient method for generating human retinal organoids. Mol. Vis. 2020, 26, 97–105. [Google Scholar]
- Ribeiro, J.; Procyk, C.A.; West, E.L.; O’Hara-Wright, M.; Martins, M.F.; Khorasani, M.M.; Hare, A.; Basche, M.; Fernando, M.; Goh, D.; et al. Restoration of visual function in advanced disease after transplantation of purified human pluripotent stem cell-derived cone photoreceptors. Cell Rep. 2021, 35, 109022. [Google Scholar] [CrossRef]
- Scholler, J.; Groux, K.; Goureau, O.; Sahel, J.-A.; Fink, M.; Reichman, S.; Boccara, C.; Grieve, K. Dynamic full-field optical coherence tomography: 3D live-imaging of retinal organoids. Light. Sci. Appl. 2020, 9, 140. [Google Scholar] [CrossRef]
- Sharma, T.; Wiley, L.A.; Whitmore, S.S.; Anfinson, K.R.; Cranston, C.M.; Oppedal, D.J.; Daggett, H.T.; Mullins, R.F.; Tucker, B.A.; Stone, E.M. Patient-specific induced pluripotent stem cells to evaluate the pathophysiology of TRNT1 -associated Retinitis pigmentosa. Stem Cell Res. 2017, 21, 58–70. [Google Scholar] [CrossRef]
- Shimada, H.; Lü, Q.; Insinna-Kettenhofen, C.; Nagashima, K.; English, M.A.; Semler, E.M.; Mahgerefteh, J.; Cideciyan, A.V.; Li, T.; Brooks, B.P.; et al. In Vitro modeling using ciliopathy-patient-derived cells reveals distinct cilia dysfunctions caused by CEP290 mutations. Cell Rep. 2017, 20, 384–396. [Google Scholar] [CrossRef]
- Shrestha, R.; Wen, Y.-T.; Ding, D.-C.; Tsai, R.-K. Aberrant hiPSCs-derived from human keratinocytes differentiates into 3D retinal organoids that acquire mature photoreceptors. Cells 2019, 8, 36. [Google Scholar] [CrossRef]
- Singh, D.; Wang, S.; Xia, T.; Tainsh, L.; Ghiassi-Nejad, M.; Xu, T.; Peng, S.; Adelman, R.A.; Rizzolo, L.J. A biodegradable scaffold enhances differentiation of embryonic stem cells into a thick sheet of retinal cells. Biomaterials 2018, 154, 158–168. [Google Scholar] [CrossRef]
- Singh, R.K.; Mallela, R.K.; Cornuet, P.K.; Reifler, A.N.; Chervenak, A.P.; West, M.D.; Wong, K.Y.; Nasonkin, I.O. Characterization of three-dimensional retinal tissue derived from human embryonic stem cells in adherent monolayer cultures. Stem Cells Dev. 2015, 24, 2778–2795. [Google Scholar] [CrossRef]
- Singh, R.K.; Occelli, L.M.; Binette, F.; Petersen-Jones, S.; Nasonkin, I.O. Transplantation of human embryonic stem cell-derived retinal tissue in the subretinal space of the cat eye. Stem Cells Dev. 2019, 28, 1151–1166. [Google Scholar] [CrossRef]
- Singh, R.K.; Winkler, P.A.; Binette, F.; Petersen-Jones, S.M.; Nasonkin, I.O. Comparison of developmental dynamics in human fetal retina and human pluripotent stem cell-derived retinal tissue. Stem Cells Dev. 2021, 30, 399–417. [Google Scholar] [CrossRef]
- Sridhar, A.; Hoshino, A.; Finkbeiner, C.R.; Chitsazan, A.; Dai, L.; Haugan, A.K.; Eschenbacher, K.M.; Jackson, D.L.; Trapnell, C.; Bermingham-McDonogh, O.; et al. Single-Cell transcriptomic comparison of human fetal retina, hPSC-derived retinal organoids, and long-term retinal cultures. Cell Rep. 2020, 30, 1644–1659.e4. [Google Scholar] [CrossRef]
- Sridhar, A.; Ohlemacher, S.K.; Langer, K.B.; Meyer, J.S. Robust differentiation of mRNA-reprogrammed human induced pluripotent stem cells toward a retinal lineage. Stem Cells Transl. Med. 2016, 5, 417–426. [Google Scholar] [CrossRef]
- Sridhar, A.; Steward, M.M.; Meyer, J.S. Nonxenogeneic growth and retinal differentiation of human induced pluripotent stem cells. Stem Cells Transl. Med. 2013, 2, 255–264. [Google Scholar] [CrossRef]
- Tornabene, P.; Trapani, I.; Minopoli, R.; Centrulo, M.; Lupo, M.; De Simone, S.; Tiberi, P.; Dell’Aquila, F.; Marrocco, E.; Iodice, C.; et al. Intein-mediated protein trans-splicing expands adeno-associated virus transfer capacity in the retina. Sci. Transl. Med. 2019, 11, eaav4523. [Google Scholar] [CrossRef]
- VanderWall, K.B.; Huang, K.-C.; Pan, Y.; Lavekar, S.S.; Fligor, C.M.; Allsop, A.R.; Lentsch, K.A.; Dang, P.; Zhang, C.; Tseng, H.C.; et al. Retinal ganglion cells with a glaucoma OPTN(E50K) mutation exhibit neurodegenerative phenotypes when derived from three-dimensional retinal organoids. Stem Cell Rep. 2020, 15, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Vergara, M.N.; Flores-Bellver, M.; Aparicio-Domingo, S.; McNally, M.; Wahlin, K.J.; Saxena, M.T.; Mumm, J.S.; Canto-Soler, M.V. Enabling quantitative screening in retinal organoids: 3D automated reporter quantification technology (3D-ARQ). Development 2017, 144, 3698–3705. [Google Scholar] [CrossRef] [PubMed]
- Vig, A.; Poulter, J.A.; Poulter, J.A.; Ottaviani, D.; Tavares, E.; Toropova, K.; Tracewska, A.M.; Mollica, A.; Kang, J.; Kehelwathugoda, O.; et al. DYNC2H1 hypomorphic or retina-predominant variants cause nonsyndromic retinal degeneration. Genet. Med. 2020, 22, 2041–2051. [Google Scholar] [CrossRef] [PubMed]
- Völkner, M.; Pavlou, M.M.; Büning, H.; Michalakis, S.; Karl, M. Optimized adeno-associated virus vectors for efficient transduction of human retinal organoids. Hum. Gene Ther. 2021, in press. [Google Scholar] [CrossRef]
- Wahlin, K.J.; Maruotti, J.A.; Sripathi, S.R.; Ball, J.; Angueyra, J.; Kim, C.; Grebe, R.; Li, W.; Jones, B.W.; Zack, D.J. Photoreceptor outer segment-like structures in long-term 3D retinas from human pluripotent stem cells. Sci. Rep. 2017, 7, 766. [Google Scholar] [CrossRef]
- Wang, S.; Poli, S.; Liang, X.; Peng, G.-H. Longitudinal single-cell RNA-seq of hESCs-derived retinal organoids. Sci. China Life Sci. 2021. [Google Scholar] [CrossRef]
- Welby, E.; Lakowski, J.; Di Foggia, V.; Budinger, D.; Cordero, A.G.; Lun, A.T.; Epstein, M.; Patel, A.; Cuevas, E.; Kruczek, K.; et al. Isolation and comparative transcriptome analysis of human Fetal AND iPSC-derived cone photoreceptor cells. Stem Cell Rep. 2017, 9, 1898–1915. [Google Scholar] [CrossRef]
- Wiley, L.; Burnight, E.R.; DeLuca, A.P.; Anfinson, K.R.; Cranston, C.M.; Kaalberg, E.E.; Penticoff, J.A.; Affatigato, L.M.; Mullins, R.F.; Stone, E.M.; et al. cGMP production of patient-specific iPSCs and photoreceptor precursor cells to treat retinal degenerative blindness. Sci. Rep. 2016, 6, 30742. [Google Scholar] [CrossRef]
- Wright, L.S.; Pinilla, I.; Saha, J.; Clermont, J.M.; Lien, J.S.; Borys, K.D.; Capowski, E.E.; Phillips, M.J.; Gamm, D.M. VSX2 and ASCL1 are indicators of neurogenic competence in human retinal progenitor cultures. PLoS ONE 2015, 10, e0135830. [Google Scholar] [CrossRef]
- Xian, B.; Luo, Z.; Li, K.; Li, K.; Tang, M.; Yang, R.; Lu, S.; Zhang, H.; Ge, J. Dexamethasone provides effective immunosuppression for improved survival of retinal organoids after epiretinal transplantation. Stem Cells Int. 2019, 2019, 7148032. [Google Scholar] [CrossRef]
- Xie, H.; Zhang, W.; Zhang, M.; Akhtar, T.; Li, Y.; Yi, W.; Sun, X.; Zuo, Z.; Wei, M.; Fang, X.; et al. Chromatin accessibility analysis reveals regulatory dynamics of developing human retina and hiPSC-derived retinal organoids. Sci. Adv. 2020, 6, eaay5247. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, M.; Zou, T.; Chen, X.; Li, Q.; Li, Y.; Ge, L.; Chen, S.; Xu, H. The impact of Particulate Matter (PM2.5) on human retinal development in hESC-derived retinal organoids. Front. Cell Dev. Biol. 2021, 9, 607341. [Google Scholar] [CrossRef]
- Zerti, D.; Dorgau, B.; Felemban, M.; Ghareeb, A.; Yu, M.; Ding, Y.; Krasnogor, N.; Lako, M. Developing a simple method to enhance the generation of cone and rod photoreceptors in pluripotent stem cell-derived retinal organoids. Stem Cells 2019, 38, 45–51. [Google Scholar] [CrossRef]
- Zerti, D.; Molina, M.M.; Dorgau, B.; Mearns, S.; Bauer, R.; Al-Aama, J.; Lako, M. IGFBPs mediate IGF-1’s functions in retinal lamination and photoreceptor development during pluripotent stem cell differentiation to retinal organoids. Stem Cells 2021, 39, 458–466. [Google Scholar] [CrossRef]
- Zhang, X.; Thompson, J.A.; Zhang, D.; Charng, J.; Arunachalam, S.; McLaren, T.L.; Lamey, T.M.; De Roach, J.N.; Jennings, L.; McLenachan, S.; et al. Characterization ofCRB1splicing in retinal organoids derived from a patient with adult-onset rod-cone dystrophy caused by the c.1892A>G and c.2548G>A variants. Mol. Genet. Genom. Med. 2020, 8, e1489. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Thompson, J.A.; Chen, S.; Huang, Z.; Jennings, L.; McLaren, T.L.; Lamey, T.M.; De Roach, J.N.; Chen, F.K.; et al. Gene correction of the CLN3 c.175G>A variant in patient-derived induced pluripotent stem cells prevents pathological changes in retinal organoids. Mol. Genet. Genom. Med. 2021, 9, e1601. [Google Scholar] [CrossRef]
- Zheng, C.; Schneider, J.W.; Hsieh, J. Role of RB1 in human embryonic stem cell-derived retinal organoids. Dev. Biol. 2020, 462, 197–207. [Google Scholar] [CrossRef]
- Zhu, Y.; Carido, M.; Meinhardt, A.; Kurth, T.; Karl, M.; Ader, M.; Tanaka, E.M. Three-Dimensional neuroepithelial culture from human embryonic stem cells and its use for quantitative conversion to retinal pigment epithelium. PLoS ONE 2013, 8, e54552. [Google Scholar] [CrossRef]
- Zou, T.; Gao, L.; Zeng, Y.; Li, Q.; Li, Y.; Chen, S.; Hu, X.; Chen, X.; Fu, C.; Xu, H.; et al. Organoid-Derived C-Kit+/SSEA4− human retinal progenitor cells promote a protective retinal microenvironment during transplantation in rodents. Nat. Commun. 2019, 10, 1205. [Google Scholar] [CrossRef]
- Eiraku, M.; Takata, N.; Ishibashi, H.; Kawada, M.; Sakakura, E.; Okuda, S.; Sekiguchi, K.; Adachi, T.; Sasai, Y. Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature 2011, 472, 51–56. [Google Scholar] [CrossRef]
- Furue, M.K.; Na, J.; Jackson, J.P.; Okamoto, T.; Jones, M.; Baker, D.; Hata, R.-I.; Moore, H.D.; Sato, J.D.; Andrews, P.W. Heparin promotes the growth of human embryonic stem cells in a defined serum-free medium. Proc. Natl. Acad. Sci. USA 2008, 105, 13409–13414. [Google Scholar] [CrossRef]
- Yamazoe, H.; Kobori, M.; Murakami, Y.; Yano, K.; Satoh, M.; Mizuseki, K.; Sasai, Y.; Iwata, H. One-Step induction of neurons from mouse embryonic stem cells in serum-free media containing vitamin B12 and heparin. Cell Transplant. 2006, 15, 135–145. [Google Scholar] [CrossRef]
- Colombres, M.; Henriquez, J.P.; Reig, G.F.; Scheu, J.; Calderón, R.; Alvarez, A.; Brandan, E.; Inestrosa, N.C. Heparin activates Wnt signaling for neuronal morphogenesis. J. Cell. Physiol. 2008, 216, 805–815. [Google Scholar] [CrossRef]
- Bejoy, J.; Wang, Z.; Bijonowski, B.; Yang, M.; Ma, T.; Sang, Q.-X.; Li, Y. Differential effects of heparin and hyaluronic acid on neural patterning of human induced pluripotent stem cells. ACS Biomater. Sci. Eng. 2018, 4, 4354–4366. [Google Scholar] [CrossRef]
- McCabe, K.; Gunther, E.; Reh, T. The development of the pattern of retinal ganglion cells in the chick retina: Mechanisms that control differentiation. Development 1999, 126, 5713–5724. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, J.; Carthew, R. Frizzled regulates mirror-symmetric pattern formation in the Drosophila eye. Development 1995, 121, 3045–3055. [Google Scholar] [CrossRef]
- Liu, C.; Nathans, J. An essential role for frizzled 5 in mammalian ocular development. Development 2008, 135, 3567–3576. [Google Scholar] [CrossRef]
- Wong, K.A.; Trembley, M.; Wahab, S.A.; Viczian, A.S. Efficient retina formation requires suppression of both Activin and BMP signaling pathways in pluripotent cells. Biol. Open 2015, 4, 573–583. [Google Scholar] [CrossRef]
- Todd, L.; Palazzo, I.; Squires, N.; Mendonca, N.; Fischer, A.J. BMP- and TGFβ-signaling regulate the formation of Müller glia-derived progenitor cells in the avian retina. Glia 2017, 65, 1640–1655. [Google Scholar] [CrossRef]











| Condition | Number of Retinal Organoids | Number of Non-Retinal Organoids | Yield of Retinal Organoids 1 |
|---|---|---|---|
| Control | 9 | 80 | 10.1% |
| Rock Inhibitor | 3 | 114 | 2.6% |
| IGF1 | 17 | 78 | 17.9% |
| IWR1e | 0 | 42 | 0.0% |
| SB431542 + LDN193189 | 1 | 106 | 0.9% |
| CHIR99021 | 105 | 99 | 51.5% |
| SU5402 | 10 | 118 | 7.8% |
| CHIR99021 + SU5402 | 56 | 175 | 24.2% |
| DAPT | 10 | 112 | 8.2% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagstaff, E.L.; Heredero Berzal, A.; Boon, C.J.F.; Quinn, P.M.J.; ten Asbroek, A.L.M.A.; Bergen, A.A. The Role of Small Molecules and Their Effect on the Molecular Mechanisms of Early Retinal Organoid Development. Int. J. Mol. Sci. 2021, 22, 7081. https://doi.org/10.3390/ijms22137081
Wagstaff EL, Heredero Berzal A, Boon CJF, Quinn PMJ, ten Asbroek ALMA, Bergen AA. The Role of Small Molecules and Their Effect on the Molecular Mechanisms of Early Retinal Organoid Development. International Journal of Molecular Sciences. 2021; 22(13):7081. https://doi.org/10.3390/ijms22137081
Chicago/Turabian StyleWagstaff, Ellie L., Andrea Heredero Berzal, Camiel J. F. Boon, Peter M. J. Quinn, Anneloor L. M. A. ten Asbroek, and Arthur A. Bergen. 2021. "The Role of Small Molecules and Their Effect on the Molecular Mechanisms of Early Retinal Organoid Development" International Journal of Molecular Sciences 22, no. 13: 7081. https://doi.org/10.3390/ijms22137081