
Short-Term Effect of a Low-Protein High-Carbohydrate Diet on Mature Female and Male, and Neomale Rainbow Trout

 and
and
Abstract
:1. Introduction
2. Results
2.1. Zootechnical Parameters
2.2. Results in Females
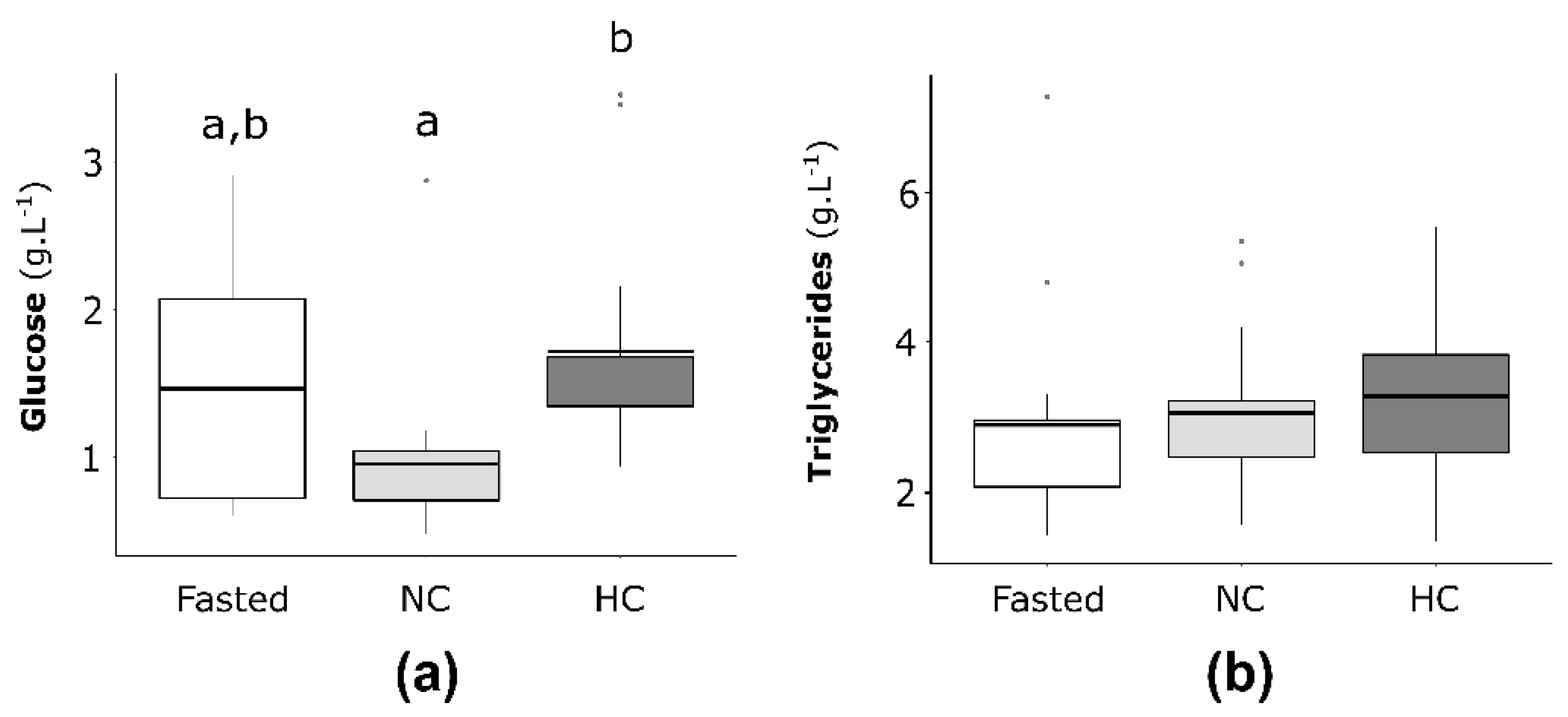
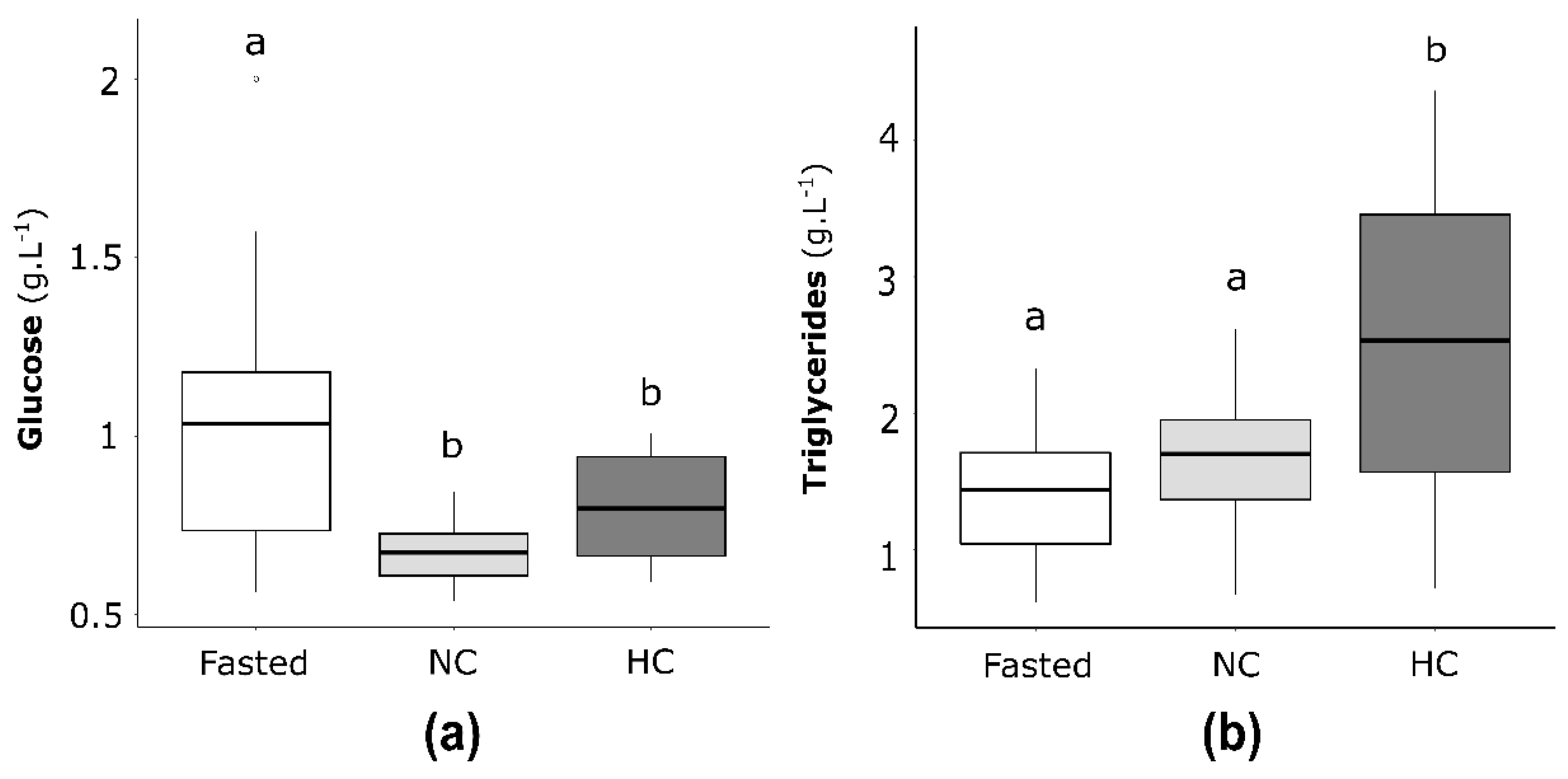
2.2.1. Plasma Glucose and Triglycerides Concentrations in Females
2.2.2. Relative mRNA Levels of Glucose and Lipid Metabolism-Related Genes in the Liver of Females
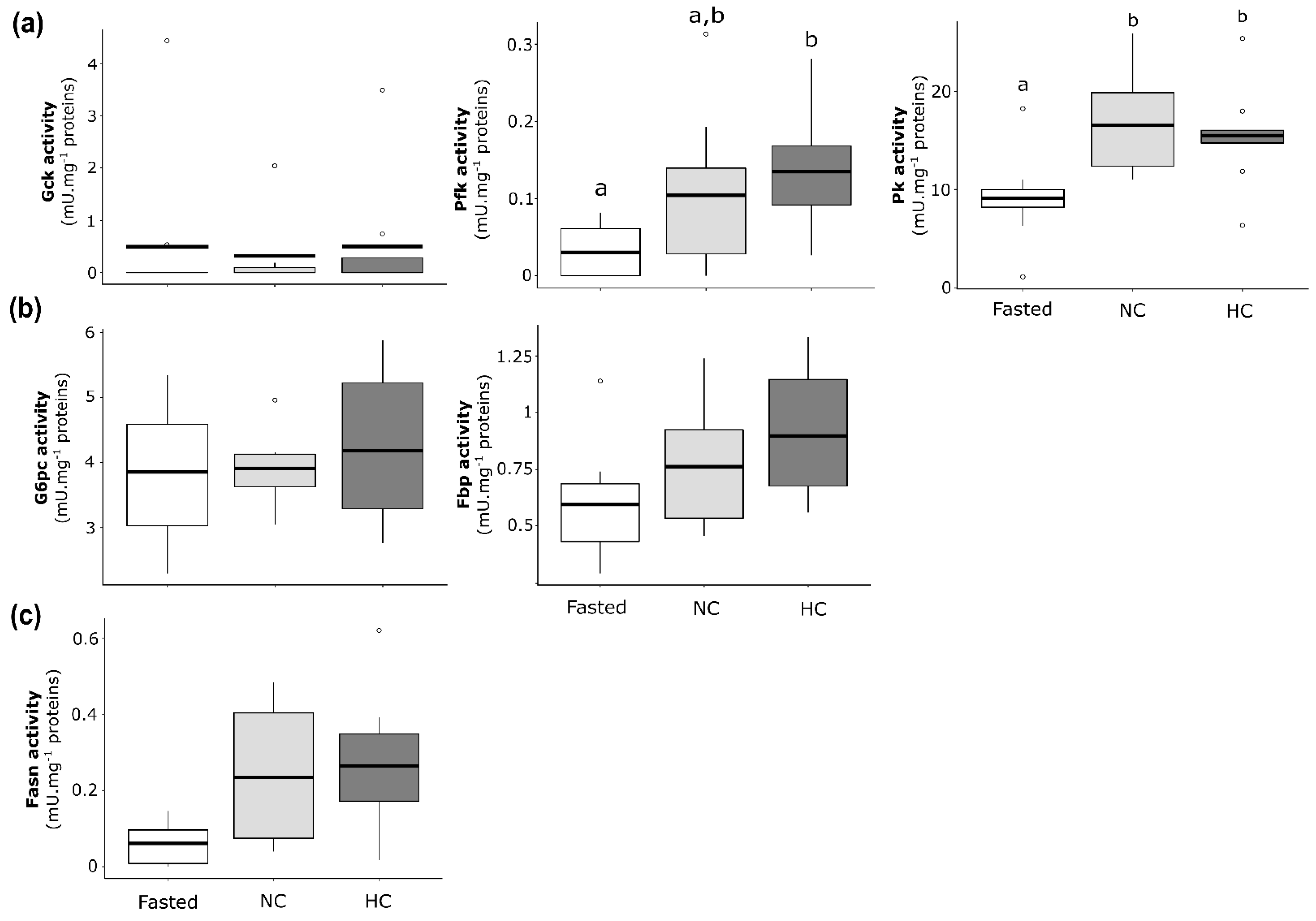
2.2.3. Enzymatic Activities of Glucose and Lipid Metabolisms in Liver of Females
2.3. Results in Males
2.3.1. Plasma Glucose and Triglycerides Concentrations in Males
2.3.2. Relative mRNA Levels of Glucose and Lipid Metabolism in Liver of Males
2.3.3. Enzymatic Activities of Glucose and Lipid Metabolism in Liver of Males
2.4. Results in Neomales
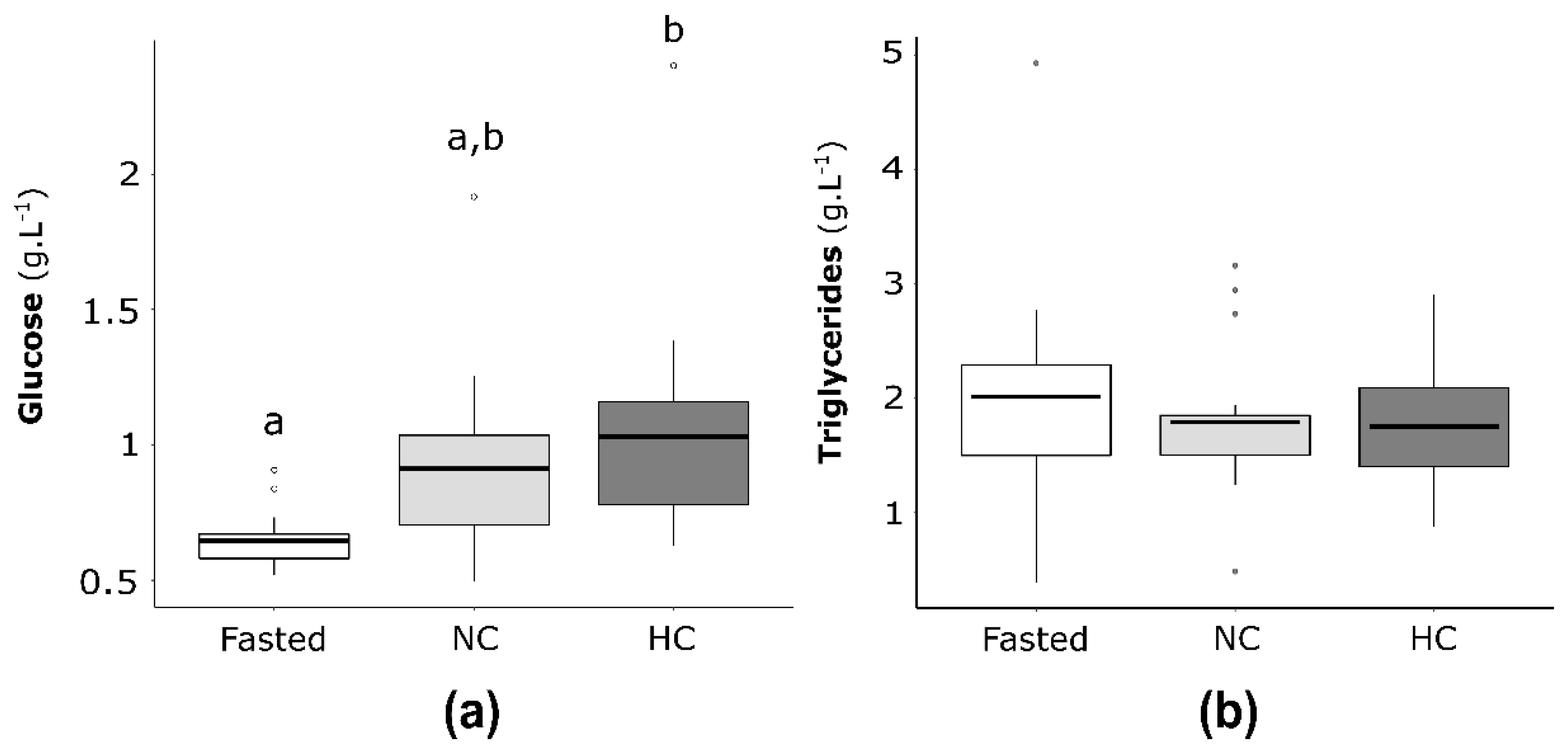
2.4.1. Plasma Glucose and Triglycerides Concentrations in Neomales
2.4.2. Relative mRNA Levels of Glucose and Lipid Metabolism in Liver of Neomales
2.4.3. Enzymatic Activities of Glucose and Lipid Metabolism in Liver of Neomales
3. Discussion
3.1. Glucose Metabolism Is Regulated at the Molecular Level by the Nutritional Status, Irrespective of the Carbohydrate Intake
3.2. At the Molecular Level, the De Novo Lipogenesis Is Upregulated and the β-Oxidation Is Downregulated by Feeding, Irrespective of the Carbohydrate Intake
3.3. The Two-Day Feeding of Mature Females and Males, and Neomales, Induced Different Regulation of the Glucose and Lipid Metabolism
4. Materials and Methods
4.1. Fish Diet
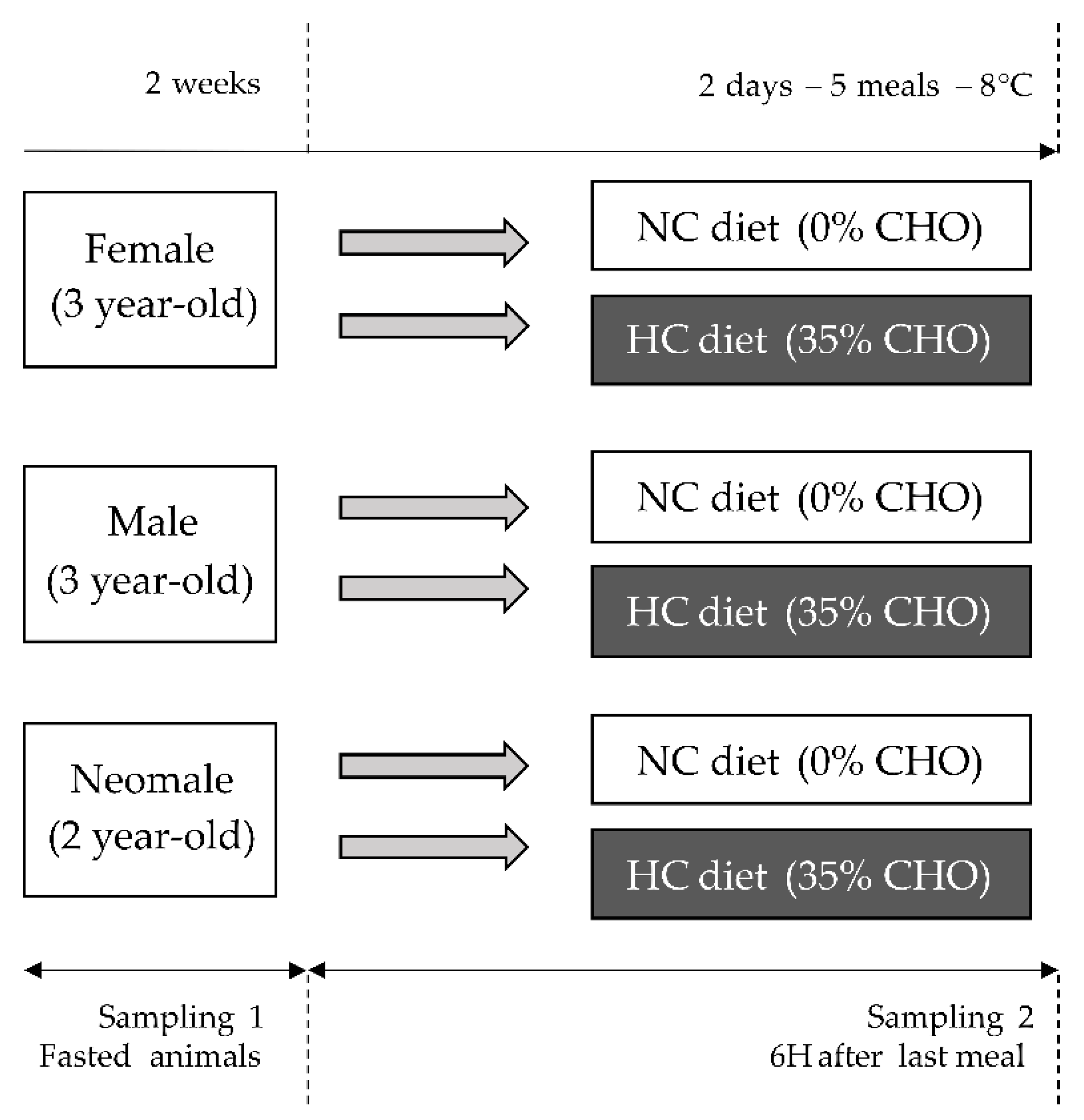
4.2. Fish Experimental Design
4.3. Plasma Metabolites
4.4. RNA Extraction and cDNA Synthesis
4.5. Quantitative Real-Time PCR
4.6. Enzymatic Activities
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Cardinaletti, G.; Belloni, A.; Giorgini, E.; Faccenda, F.; Cerri, R.; Tibaldi, E.; Olivotto, I. Physiological Response of Rainbow Trout (Oncorhynchus Mykiss) to Graded Levels of Hermetia Illucens or Poultry by-Product Meals as Single or Combined Substitute Ingredients to Dietary Plant Proteins. Aquaculture 2021, 538, 736550. [Google Scholar] [CrossRef]
- Panserat, S.; Médale, F.; Brèque, J.; Plagnes-Juan, E.; Kaushik, S. Lack of Significant Long-Term Effect of Dietary Carbohydrates on Hepatic Glucose-6-Phosphatase Expression in Rainbow Trout (Oncorhynchus Mykiss)11The Genbank Accession Number for the Rainbow Trout G6Pase Sequence Is AF120150. J. Nutr. Biochem. 2000, 11, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Panserat, S.; Capilla, E.; Gutierrez, J.; Frappart, P.O.; Vachot, C.; Aguirre, P.; Breque, J.; Kaushik, S. Glucokinase Is Highly Induced and Glucose-6-Phosphatase Poorly Repressed in Liver of Rainbow Trout ž Oncorhynchus Mykiss/by a Single Meal with Glucose. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 128, 275–283. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose Metabolism in Fish: A Review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef]
- Marandel, L.; Seiliez, I.; Véron, V.; Skiba-Cassy, S.; Panserat, S. New Insights into the Nutritional Regulation of Gluconeogenesis in Carnivorous Rainbow Trout (Oncorhynchus Mykiss): A Gene Duplication Trail. Physiol. Genom. 2015, 47, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panserat, S.; Skiba-Cassy, S.; Seiliez, I.; Lansard, M.; Plagnes-Juan, E.; Vachot, C.; Aguirre, P.; Larroquet, L.; Chavernac, G.; Medale, F.; et al. Metformin Improves Postprandial Glucose Homeostasis in Rainbow Trout Fed Dietary Carbohydrates: A Link with the Induction of Hepatic Lipogenic Capacities? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R707–R715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polakof, S.; Médale, F.; Larroquet, L.; Vachot, C.; Corraze, G.; Panserat, S. Regulation of de Novo Hepatic Lipogenesis by Insulin Infusion in Rainbow Trout Fed a High-Carbohydrate Diet1. J. Anim. Sci. 2011, 89, 3079–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamalam, B.S.; Medale, F.; Panserat, S. Utilisation of Dietary Carbohydrates in Farmed Fishes: New Insights on Influencing Factors, Biological Limitations and Future Strategies. Aquaculture 2017, 467, 3–27. [Google Scholar] [CrossRef]
- Luquet, P.; Watanabe, T. Interaction “Nutrition-Reproduction” in Fish. Fish. Physiol. Biochem. 1986, 2, 121–129. [Google Scholar] [CrossRef]
- Washburn, B.S.; Frye, D.J.; Hung, S.S.O.; Doroshov, S.I.; Conte, F.S. Dietary Effects on Tissue Composition, Oogenesis and the Reproductive Performance of Female Rainbow Trout (Oncorhynchus Mykiss). Aquaculture 1990, 90, 179–195. [Google Scholar] [CrossRef]
- Barciela, P.; Soengas, J.L.; Rey, P.; Aldegunde, M.; Rozas, G. Carbohydrate Metabolism in Several Tissues of Rainbow Trout, Oncorhynchus Mykiss, Is Modified during Ovarian Recrudescence. Comp. Biochem. Physiol. Part B Comp. Biochem. 1993, 106, 943–948. [Google Scholar] [CrossRef]
- Soengas, J.L.; Sanmartin, B.; Barciela, P.; Aldegunde, M.; Rozas, G. Changes in Carbohydrate Metabolism Related to the Onset of Ovarian Recrudescence in Dmesticated Rainbow Trout (Oncorhynchus Mykiss). Comp. Biochem. Physiol. Part A Physiol. 1993, 105, 293–301. [Google Scholar] [CrossRef]
- Soengas, J.L.; Sanmartín, B.; Barciela, P.; Aldegunde, M.; Rozas, G. Changes in Carbohydrate Metabolism in Domesticated Rainbow Trout (Oncorhynchus Mykiss) Related to Spermatogenesis. Comp. Biochem. Physiol. Part B Comp. Biochem. 1993, 105, 665–671. [Google Scholar] [CrossRef]
- Callet, T.; Hu, H.; Larroquet, L.; Surget, A.; Liu, J.; Plagnes-Juan, E.; Maunas, P.; Turonnet, N.; Mennigen, J.A.; Bobe, J.; et al. Exploring the Impact of a Low-Protein High-Carbohydrate Diet in Mature Broodstock of a Glucose-Intolerant Teleost, the Rainbow Trout. Front. Physiol. 2020, 11, 303. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, R.K.; Sarosiek, B.; Demianowicz, W.; Judek, J.; Goryczko, K.; Dobosz, S.; Kuźmiński, H.; Demska-Zakęś, K.; Babiak, I.; Glogowski, J. Quantitative Characteristics of Rainbow Trout, Oncorhynchus Mykiss, Neo-Males (XX Genotype) and Super-Males (YY Genotype) Sperm. World Acad. Sci. Eng. Technol. 2011. [Google Scholar] [CrossRef]
- Walton, M.J.; Cowey, C.B. Gluconeogenesis by Isolated Hepatocytes from Rainbow Trout Salmo Gairdneri. Comp. Biochem. Physiol. Part B Comp. Biochem. 1979, 62, 75–79. [Google Scholar] [CrossRef]
- Cowey, C.B.; Cooke, D.J.; Matty, A.J.; Adron, J.W. Effects of Quantity and Quality of Dietary Protein on Certain Enzyme Activities in Rainbow Trout. J. Nutr. 1981, 111, 336–345. [Google Scholar] [CrossRef] [Green Version]
- French, C.J.; Mommsen, T.P.; Hochachka, P.W. Amino Acid Utilisation in Isolated Hepatocytes from Rainbow Trout. Eur. J. Biochem. 1981, 113, 311–317. [Google Scholar] [CrossRef]
- Groen, A.K.; Vervoorn, R.C.; Tager, J.M. Control of Gluconeogenesis in Rat Liver Cells. Biochem. Soc. Trans. 1983, 11, 88. [Google Scholar] [CrossRef]
- Washizu, T.; Tanaka, A.; Sako, T.; Washizu, M.; Arai, T. Comparison of the Activities of Enzymes Related to Glycolysis and Gluconeogenesis in the Liver of Dogs and Cats. Res. Vet. Sci. 1999, 67, 205–206. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Vijayan, M.M.; Moon, T.W. In Vitro Hepatocyte Metabolism of Alanine and Glucose and the Response to Insulin in Fed and Fasted Rainbow Trout. J. Exp. Zool. 1995, 271, 425–431. [Google Scholar] [CrossRef]
- Viegas, I.; Rito, J.; González, J.D.; Jarak, I.; Carvalho, R.A.; Metón, I.; Pardal, M.A.; Baanante, I.V.; Jones, J.G. Effects of Food-Deprivation and Refeeding on the Regulation and Sources of Blood Glucose Appearance in European Seabass (Dicentrarchus Labrax L.). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 399–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panserat, S.; Médale, F.; Blin, C.; Brèque, J.; Vachot, C.; Plagnes-Juan, E.; Gomes, E.; Krishnamoorthy, R.; Kaushik, S. Hepatic Glucokinase Is Induced by Dietary Carbohydrates in Rainbow Trout, Gilthead Seabream, and Common Carp. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R1164–R1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Shima, T.; Furuita, H.; Sugita, T.; Suzuki, N. Effects of Feeding Time, Water Temperature, Feeding Frequency and Dietary Composition on Apparent Nutrient Digestibility in Rainbow Trout Oncorhynchus Mykiss and Common Carp Cyprinus Carpio. Fish Sci. 2007, 73, 161–170. [Google Scholar] [CrossRef]
- Figueiredo-Silva, A.C.; Panserat, S.; Kaushik, S.; Geurden, I.; Polakof, S. High Levels of Dietary Fat Impair Glucose Homeostasis in Rainbow Trout. J. Exp. Biol. 2012, 215, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Romsos, D.R.; Tack, P.I.; Leveille, G.A. Effects of Fasting and Feeding Various Diets on Hepatic Lipogenic Enzyme Activities in Coho Salmon (Oncorhynchus Kisutch (Walbaum)). J. Nutr. 1977, 107, 1477–1483. [Google Scholar] [CrossRef]
- Tucci, S.; Alatibi, K.I.; Wehbe, Z. Altered Metabolic Flexibility in Inherited Metabolic Diseases of Mitochondrial Fatty Acid Metabolism. Int. J. Mol. Sci. 2021, 22, 3799. [Google Scholar] [CrossRef]
- Kamalam, B.S.; Medale, F.; Kaushik, S.; Polakof, S.; Skiba-Cassy, S.; Panserat, S. Regulation of Metabolism by Dietary Carbohydrates in Two Lines of Rainbow Trout Divergently Selected for Muscle Fat Content. J. Exp. Biol. 2012, 215, 2567–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechanisms of Migration in Fishes; McCleave, J.D. (Ed.) IV Marine Sciences; Springer US: Berlin/Heidelberg, Germany, 1984; ISBN 978-1-4612-9708-6. [Google Scholar]
- Banks, J.W. A Review of the Literature on the Upstream Migration of Adult Salmonids. J. Fish. Biol. 1969, 1, 85–136. [Google Scholar] [CrossRef]
- Ridelman, J.M.; Hardy, R.W.; Brannon, E.L. The Effect of Short-Term Starvation on Ovarian Development and Egg Viability in Rainbow Trout (Salmo Gairdneri). Aquaculture 1984, 37, 133–140. [Google Scholar] [CrossRef]
- Nynca, J.; Kuźmiński, H.; Dietrich, G.J.; Hliwa, P.; Dobosz, S.; Liszewska, E.; Karol, H.; Ciereszko, A. Biochemical and Physiological Characteristics of Semen of Sex-Reversed Female Rainbow Trout (Oncorhynchus Mykiss, Walbaum). Theriogenology 2012, 77, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Marandel, L.; Labbe, C.; Bobe, J.; Le Bail, P.-Y. Evolutionary History of c-Myc in Teleosts and Characterization of the Duplicated c-Myca Genes in Goldfish Embryos: TELEOST c-Myc HISTORY AND EMBRYONIC EXPRESSION. Mol. Reprod. Dev. 2012, 79, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Alegre, M.; Ciudad, C.J.; Fillat, C.; Guinovart, J.J. Determination of Glucose-6-Phosphatase Activity Using the Glucose Dehydrogenase-Coupled Reaction. Anal. Biochem. 1988, 173, 185–189. [Google Scholar] [CrossRef]
- Kirchner, S.; Kaushik, S.; Panserat, S. Low Protein Intake Is Associated with Reduced Hepatic Gluconeogenic Enzyme Expression in Rainbow Trout (Oncorhynchus Mykiss). J. Nutr. 2003, 133, 2561–2564. [Google Scholar] [CrossRef] [PubMed]
- Tranulis, M.A.; Christophersen, B. A Glucokinase-Like Enzyme in the Liver of Atlantic Salmon (Salmo Salar). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 114, 35–39. [Google Scholar] [CrossRef]
- Metón, I.; Caseras, A.; Fernández, F.; Baanante, I.V. Molecular Cloning of Hepatic Glucose-6-Phosphatase Catalytic Subunit from Gilthead Sea Bream (Sparus Aurata): Response of Its MRNA Levels and Glucokinase Expression to Refeeding and Diet Composition. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2004, 138, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, K.; Leveille, G.A. Acetyl CoA Carboxylase and Fatty Acid Synthetase Activities in Liver and Adipose Tissue of Meal-Fed Rats. Proc. Soc. Exp. Biol. Med. 1969, 131, 1051–1054. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway/Genes | Fasted | NC | HC | p-Value |
|---|---|---|---|---|
| Glucose transport | ||||
| glut2a | 1.92 ± 0.60 | 2.36 ± 1.33 | 1.82 ± 1.17 | 0.573 |
| glut2b | 1.91 ± 0.64 | 2.37 ± 0.82 | 1.99 ± 1.09 | 0.567 |
| Glycolysis | ||||
| gcka | 3.01 ± 7.0 | 0 | 3.36 ± 5.37 | 0.119 |
| gckb | 1.83 ± 4.12 a,b | 0.15 ± 0.36 a | 4.16 ± 6.18 b | 0.049 |
| pfkla | 0.92 ± 0.28 a | 1.45 ± 0.35 b | 1.27 ± 0.51 a,b | 0.045 |
| pfklb | 0.94 ± 0.32 | 1.5 ± 0.57 | 1.3 ± 0.33 | 0.051 |
| pkl | 0.8 ± 0.35 | 1.37 ± 0.48 | 1.25 ± 0.63 | 0.1 |
| Gluconeogenesis | ||||
| g6pca | 0.93 ± 0.37 a | 0.75 ± 0.64 a,b | 0.42 ± 0.20 b | 0.004 |
| g6pcb1a | 3.08 ± 3.17 a | 0.94 ± 1.71 a,b | 0.12 ± 0.37 b | 0.005 |
| g6pcb1b | 0.35 ± 0.18 a | 0.7 ± 0.76 a | 0.09 ± 0.06 b | 0.001 |
| g6pcb2a | 0.49 ± 0.57 a | 1.49 ± 0.87 a,b | 1.8 ± 1.46 b | 0.020 |
| g6pcb2b | nd | nd | nd | - |
| fbp1a | nd | nd | nd | - |
| fbp1b1 | 0.9 ± 0.43 | 1.02 ± 0.90 | 0.73 ± 0.23 | 0.848 |
| fbp1b2 | 2.01 ± 0.67 | 2.11 ± 2.0 | 1.6 ± 0.76 | 0.340 |
| pck1 | 0 | 0.11 ± 0.23 | 0 | 0.157 |
| pck2a | 1.79 ± 1.33 | 1.28 ± 0.58 | 1.45 ± 0.77 | 0.557 |
| pck2b | 1.05 ± 0.53 | 1.56 ± 0.72 | 1.47 ± 0.65 | 0.266 |
| Lipid metabolism | ||||
| srebf1 | 0.2 ± 0.11 a | 0.99 ± 0.60 b | 1.34 ± 0.88 b | <0.001 |
| g6pd | 1.63 ± 1.12 | 3.19 ± 2.45 | 2.95 ± 2.14 | 0.289 |
| acly | 0.92 ± 0.46 a | 2.64 ± 1.14 b | 2.04 ± 2.14 b | 0.007 |
| aca-αa | 0.15 ± 0.11 a | 2.08 ± 2.06 b | 2.72 ± 1.26 b | <0.001 |
| aca-αb | 0.17 ± 0.09 a | 1.66 ± 1.49 b | 2.53 ± 2.44 b | <0.001 |
| aca-βa | 2.53 ± 0.67 | 1.87 ± 2.37 | 2.06 ± 1.26 | 0.160 |
| aca-βb | 1.29 ± 0.59 | 1.3 ± 0.59 | 1.28 ± 0.87 | 0.701 |
| fasn | 0.12 ± 0.22 a | 3.28 ± 2.20 b | 1.67 ± 1.36 b | 0.001 |
| Pathway/Genes | Fasted | NC | HC | p-Value |
|---|---|---|---|---|
| Glucose transport | ||||
| glut2a | 2.51 ± 1.48 | 2.88 ± 1.66 | 2.15 ± 1.21 | 0.790 |
| glut2b | 2.12 ± 1.23 | 2.59 ± 1.35 | 2.07 ± 0.96 | 0.656 |
| Glycolysis | ||||
| gcka | 1.61 ± 1.64 a | 0 b | 0 b | 0.001 |
| gckb | 0 | 0 | 0.29 ± 0.87 | 0.434 |
| pfkla | 0.6 ± 0.30 | 1.21 ± 0.68 | 0.9 ± 0.37 | 0.076 |
| pfklb | 0.59 ± 0.31 | 1.2 ± 0.64 | 0.93 ± 0.38 | 0.062 |
| pkl | 0.91 ± 0.61 | 1.48 ± 0.73 | 1.43 ± 0.39 | 0.100 |
| Gluconeogenesis | ||||
| g6pca | 1.85 ± 1.1 | 1.82 ± 1.2 | 1.23 ± 0.75 | 0.393 |
| g6pcb1a | 0.38 ± 0.61 | 0.53 ± 1.59 | 0 | 0.127 |
| g6pcb1b | 1.1 ± 0.7 a,b | 2.76 ± 3.14 a | 0.62 ± 0.43 b | 0.046 |
| g6pcb2a | 0.36 ± 0.17a | 1.71 ± 1.1 b | 1.87 ± 1.5 b | 0.018 |
| g6pcb2b | nd | nd | nd | - |
| fbp1a | nd | nd | nd | - |
| fbp1b1 | 1.66 ± 0.66 | 1.86 ± 0.92 | 1.62 ± 0.86 | 0.773 |
| fbp1b2 | 0.84 ± 0.68 | 0.63 ± 0.43 | 0.74 ± 0.29 | 0.592 |
| pck1 | 0.19 ± 0.23 a | 4.88 ± 6.35 b | 0.3 ± 0.3 a | 0.005 |
| pck2a | 2.99 ± 2.26 | 1.87 ± 1.09 | 2.50 ± 1.77 | 0.317 |
| pck2b | 1.21 ± 0.55 | 1.80 ± 0.83 | 1.33 ± 0.69 | 0.191 |
| Lipid metabolism | ||||
| srebf1 | 0.17 ± 0.06 a | 1.19 ± 0.86 b | 0.92 ± 0.59 b | <0.001 |
| g6pd | 1.64 ± 0.95 | 1.63 ± 1.02 | 1.69 ± 0.75 | 0.989 |
| acly | 0.96 ± 0.63 | 2.01 ± 1.27 | 1.57 ± 0.72 | 0.132 |
| aca-αa | 0.13 ± 0.09 a | 1.37 ± 1.59 b | 1.67 ± 1.43 b | <0.001 |
| aca-αb | 0.15 ± 0.09 a | 1.27 ± 1.24 b | 1.67 ± 1.31 b | <0.001 |
| aca-βa | 1.95 ± 0.84 a | 0.53 ± 0.39 b | 0.53 ± 0.36 b | <0.001 |
| aca-βb | 1.18 ± 0.62 | 1.94 ± 1.30 | 1.41 ± 0.67 | 0.177 |
| fasn | 0.21 ± 0.12 a | 1.97 ± 1.24 b | 2.13 ± 1.34 b | 0.004 |
| Pathway/Genes | Fasted | NC | HC | p-Value |
|---|---|---|---|---|
| Glucose transport | ||||
| glut2a | 1.96 ± 1.46 | 2.20 ± 1.10 | 1.92 ± 0.83 | 0.782 |
| glut2b | 1.64 ± 1.08 | 1.89 ± 1.05 | 1.75 ± 0.84 | 0.917 |
| Glycolysis | ||||
| gcka | 0 | 0 | 0.89 ± 1.51 | 0.125 |
| gckb | 0.02 ± 0.06 a | 0.02 ± 0.06 a | 2.51 ± 2.92 b | 0.006 |
| pfkla | 0.88 ± 0.24 a | 1.48 ± 0.77 b | 1.55 ± 0.99 b | 0.042 |
| pfklb | 0.83 ± 0.21 a | 1.43 ± 0.7 b | 1.46 ± 0.88 b | 0.037 |
| pkl | 0.75 ± 0.35 | 1.04 ± 0.46 | 1.13 ± 0.69 | 0.156 |
| Gluconeogenesis | ||||
| g6pca | 1.22 ± 0.37 | 1.04 ± 0.51 | 0.84 ± 0.93 | 0.081 |
| g6pcb1a | 1.92 ± 1.06a | 0 b | 0 b | <0.001 |
| g6pcb1b | 0.84 ± 0.74 a | 3.7 ± 3.25 b | 1.04 ± 2.10 a,b | 0.015 |
| g6pcb2a | 0.32 ± 0.37 a | 1.73 ± 1.60 b | 1.35 ± 1.25 b | 0.001 |
| g6pcb2b | nd | nd | nd | - |
| fbp1a | nd | nd | nd | - |
| fbp1b1 | 0.96 ± 0.64 | 1.34 ± 0.67 | 0.76 ± 0.40 | 0.067 |
| fbp1b2 | 1.3 ± 0.63 a | 0.97 ± 0.52 a,b | 0.59 ± 0.24 b | 0.006 |
| pck1 | 0.03 ± 0.08 a | 5.32 ± 4.21 b | 2.17 ± 5.76 a | <0.001 |
| pck2a | 2.99 ± 2.26 | 1.87 ± 1.09 | 2.50 ± 1.77 | 0.317 |
| pck2b | 1.21 ± 0.55 | 1.80 ± 0.83 | 1.33 ± 0.69 | 0.191 |
| Lipid metabolism | ||||
| srebf1 | 0.4 ± 0.23 a | 1.2 ± 0.65 b | 1.9 ± 1.32 b | <0.001 |
| g6pd | 0.6 ± 0.87 a | 2.51 ± 1.37 b | 1.99 ± 1.24 b | 0.001 |
| acly | 0.68 ± 0.9 a | 2.4 ± 1.08 b | 2.15 ± 1.53 b | 0.001 |
| aca-αa | 0.18 ± 0.11 a | 1.87 ± 1.27 b | 3.63 ± 3.24 b | <0.001 |
| aca-αb | 0.22 ± 0.13 a | 1.78 ± 1.15 b | 3.49 ± 3.09 b | <0.001 |
| aca-βa | 2.31 ± 1.23 a | 1.12 ± 0.74 a,b | 0.79 ± 0.52 b | 0.005 |
| aca-βb | 2.35 ± 0.60 a | 1.19 ± 0.52 b | 1.38 ± 0.78 a,b | 0.005 |
| fasn | 0.05 ± 0.11 a | 2.06 ± 1.25 b | 2.5 ± 1.85 b | <0.001 |
| Ingredients (%) | NC | HC | ||
|---|---|---|---|---|
| 5 mm | 9 mm | 5 mm | 9 mm | |
| Fish meal 1 | 77.77 | 77.77 | 45 | 45 |
| Pre-gelatinised starch 2 | - | - | 37 | 37 |
| CPSP90 3 | 2 | 2 | 5 | 5 |
| Soybean meal 4 | 12 | 12 | - | - |
| Pea protein concentrated 5 | - | - | 5 | 5 |
| Fish oil 6 | 1.66 | 1.66 | 3.96 | 3.96 |
| Cellulosis 7 | 2.53 | 2.53 | - | - |
| Alginate 8 | 2 | 2 | 2 | 2 |
| Minerals premix 9 | 1 | 1 | 1 | 1 |
| Vitamins premix 9 | 1 | 1 | 1 | 1 |
| Astaxanthine 10 | 0.04 | 0.04 | 0.04 | 0.04 |
| Proximate composition | ||||
| Dry matter | 93.97 | 94.42 | 95.84 | 96.05 |
| Crude proteins (%DM) | 66.63 | 65.7 | 41.97 | 42.94 |
| Gross energy (kJ/gDM) | 20.21 | 20.48 | 18.73 | 20.44 |
| Ash (%DM) | 16.81 | 16.75 | 10.37 | 10.4 |
| Carbohydrates (%DM) | 0.55 | 0.55 | 32.55 | 32.55 |
| Crude lipids (%DM) | 8.71 | 9.69 | 7.57 | 6.47 |
| Gene | Forward Primer (5’-3’) | Reverse Primer (5’-3’) | GenBank Accession Number |
|---|---|---|---|
| srebf1 | CAGTTGCTGCTGTGTGACCT | TGATGTGTTCGTGTGGGACT | XM_021624594.1 |
| acly | CCTCTGTAAGGCCAAGTGGG | TTGGCATCCAGGTCTGCAAT | XM_021624819.1/XM_021557697.1/XM_021575392.1 |
| aca-αa | ACAGGACCCTAAAGCACAGG | GGTGAAAGAGGTGTCCAGGA | XM_021623125.1 |
| aca-αb | TCCAGTTCATGCTGCCTACC | GCTTAATGTCCCGAGTGCGA | XM_021618451.1 |
| aca-βa | TCGCTCAGAATTCCGGGTAC | CGCGTGGTGATGGTTACAAT | XM_021605386.1 |
| aca-βb | TGAACAGCTTGGTAAACAGCC | TCTCGTGCATTCTACCAGGG | XM_021620987.1 |
| fasn | TGATCTGAAAGCCCGTGTCAA | ATTTGGTTGCCGGGACGTAA | XM_021576228.1/XM_021581290.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Favalier, N.; Véron, V.; Marchand, M.; Surget, A.; Maunas, P.; Turonnet, N.; Panserat, S.; Marandel, L. Short-Term Effect of a Low-Protein High-Carbohydrate Diet on Mature Female and Male, and Neomale Rainbow Trout. Int. J. Mol. Sci. 2021, 22, 6149. https://doi.org/10.3390/ijms22116149
Favalier N, Véron V, Marchand M, Surget A, Maunas P, Turonnet N, Panserat S, Marandel L. Short-Term Effect of a Low-Protein High-Carbohydrate Diet on Mature Female and Male, and Neomale Rainbow Trout. International Journal of Molecular Sciences. 2021; 22(11):6149. https://doi.org/10.3390/ijms22116149
Chicago/Turabian StyleFavalier, Nathan, Vincent Véron, Michael Marchand, Anne Surget, Patrick Maunas, Nicolas Turonnet, Stéphane Panserat, and Lucie Marandel. 2021. "Short-Term Effect of a Low-Protein High-Carbohydrate Diet on Mature Female and Male, and Neomale Rainbow Trout" International Journal of Molecular Sciences 22, no. 11: 6149. https://doi.org/10.3390/ijms22116149