
Chorioamnionitis Precipitates Perinatal Alterations of Heme-Oxygenase-1 (HO-1) Homeostasis in the Developing Rat Brain

 , ,
, ,
Abstract
:1. Introduction
2. Results
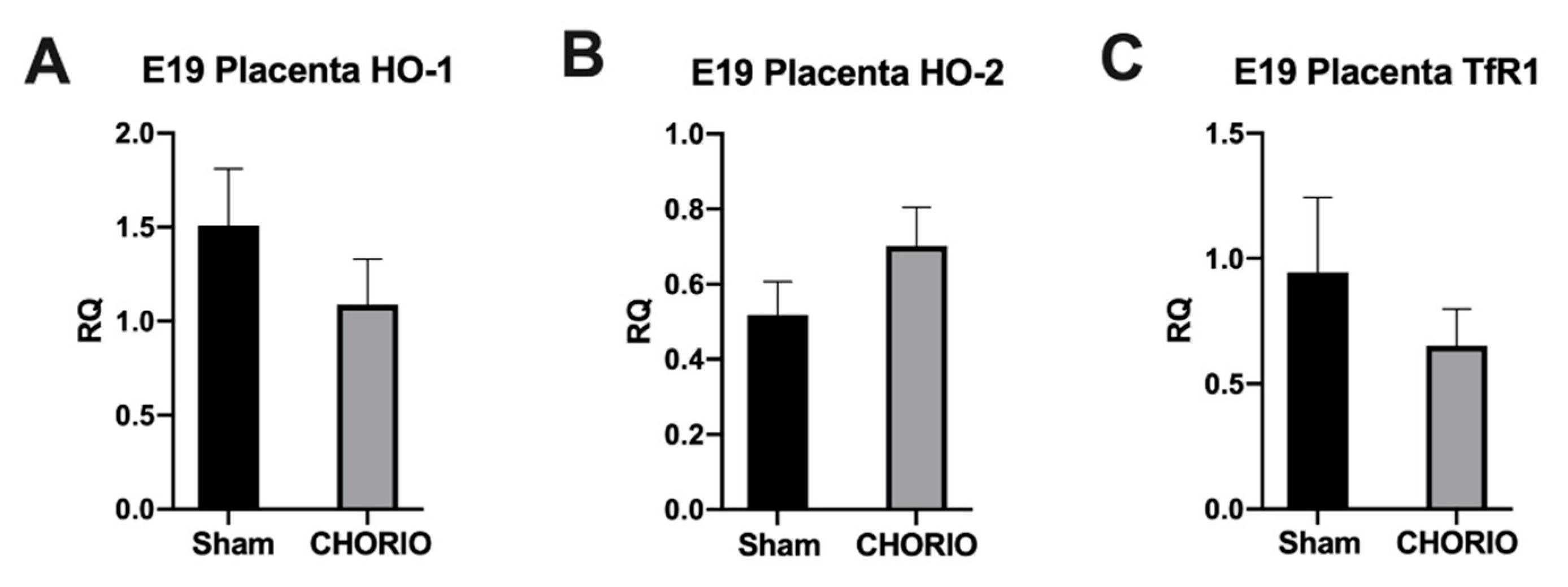
2.1. Placenta HO-1, HO-2, and TfR1 Expression
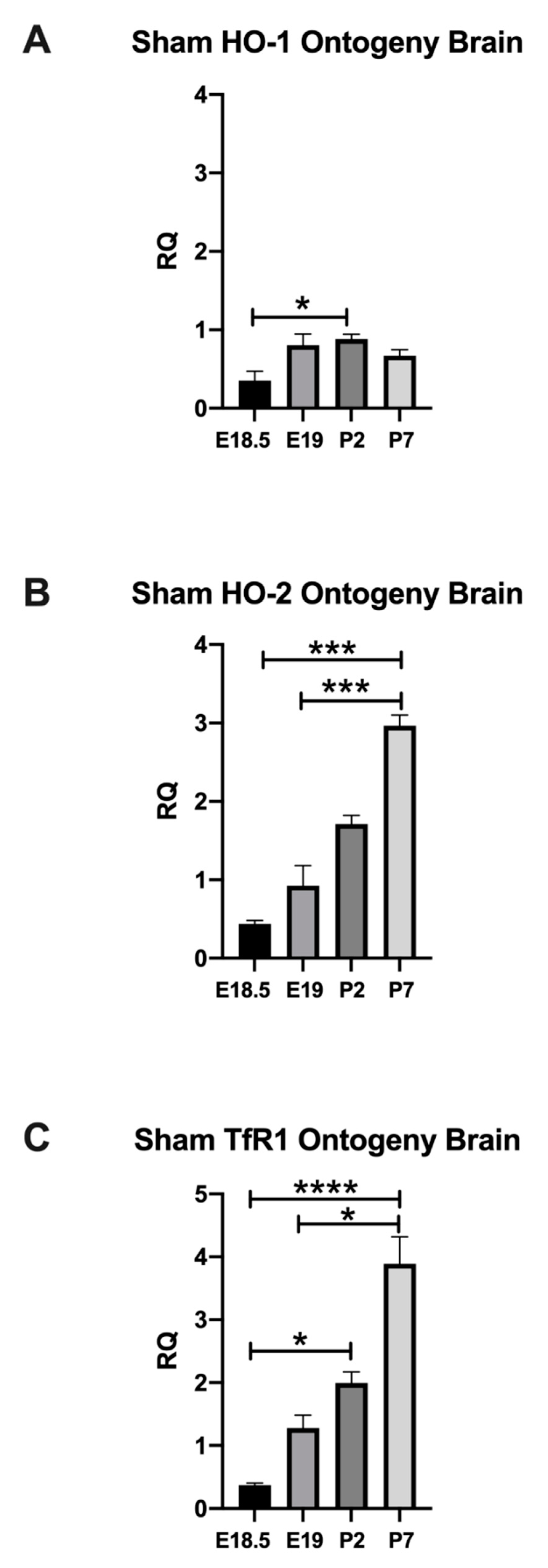
2.1.1. Normal Developmental Regulation of HO-1 in Rat Brain
2.1.2. Normal Developmental Regulation of HO-2 in Rat Brain
2.1.3. Normal Developmental Regulation of TfR1 in Rat Brain
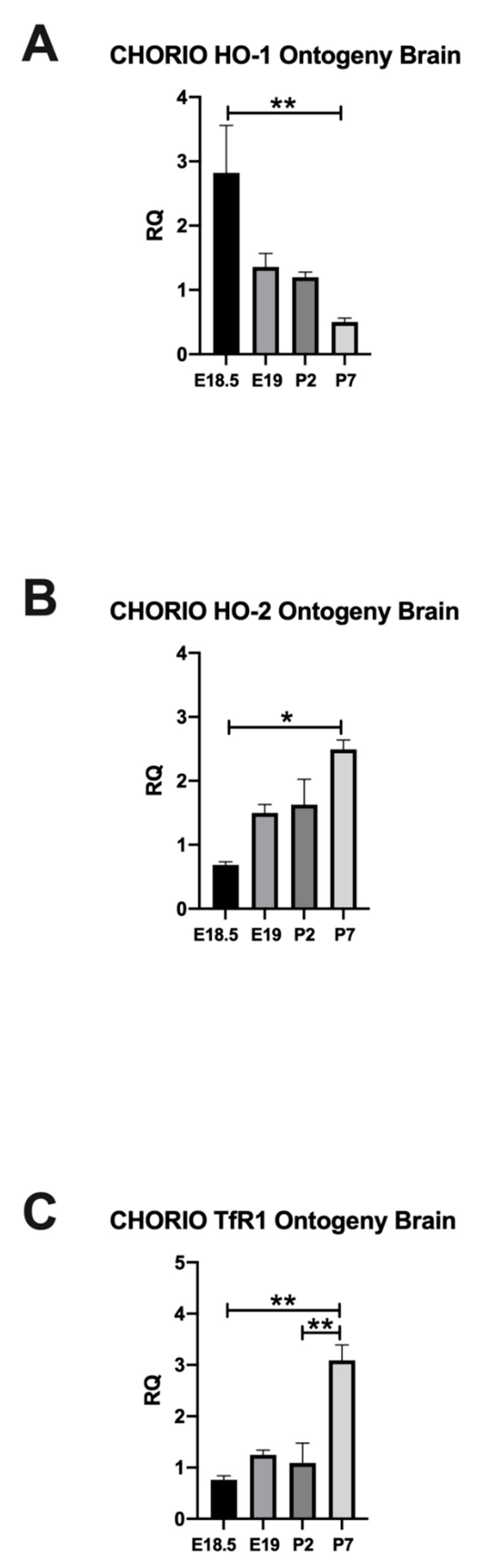
2.2. CHORIO Altered the Ontogeny of HO-1, HO-2, and TfR1 in Rat Brain
2.2.1. HO-1 Expression in Rat CHORIO Brain
2.2.2. HO-2 Expression in Rat CHORIO Brain
2.3. TfR1 Expression in Rat CHORIO Brain
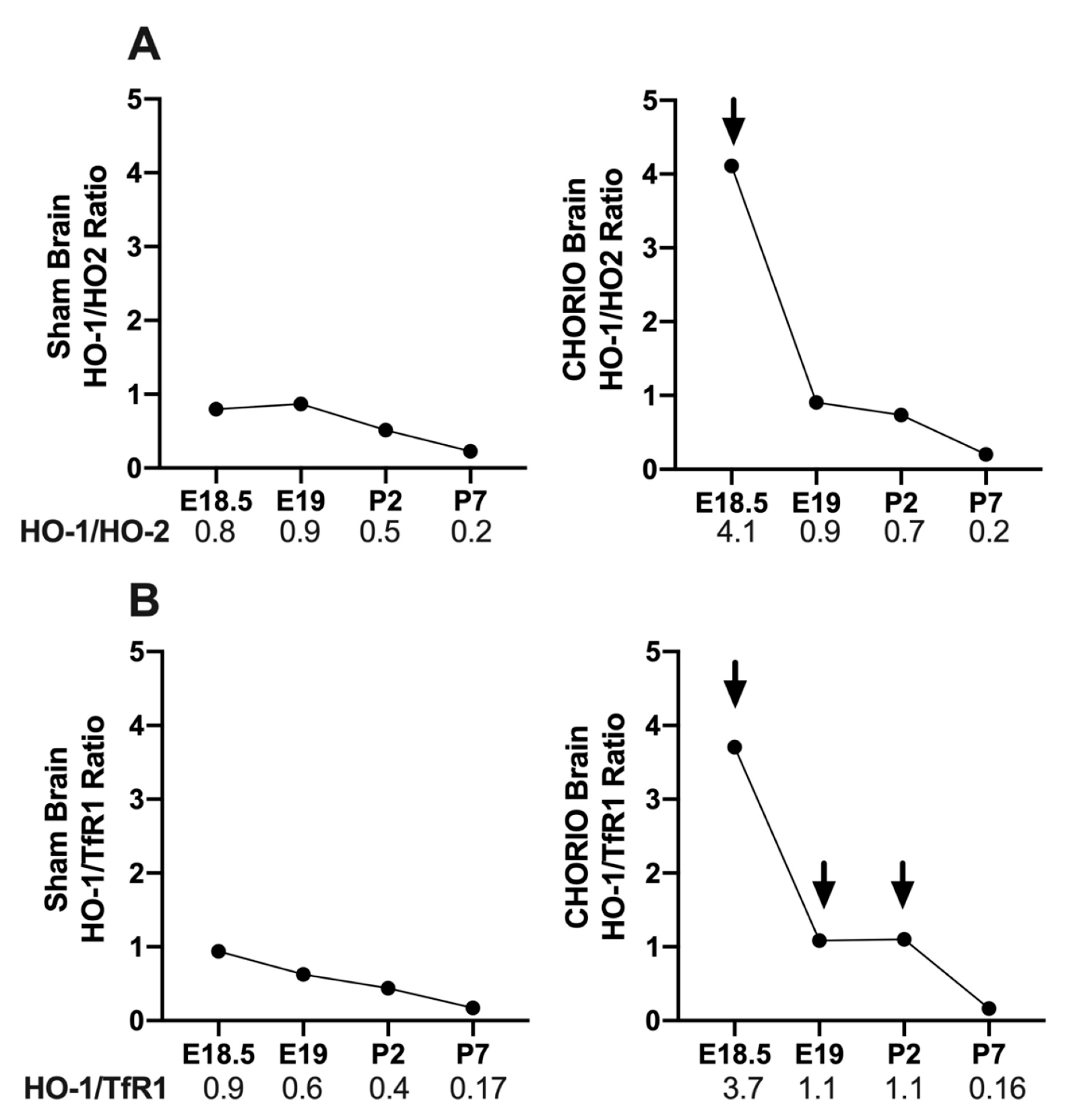
2.3.1. CHORIO-Induced Changes in Brain HO-1/HO-2 and HO-1/TfR1 Ratios
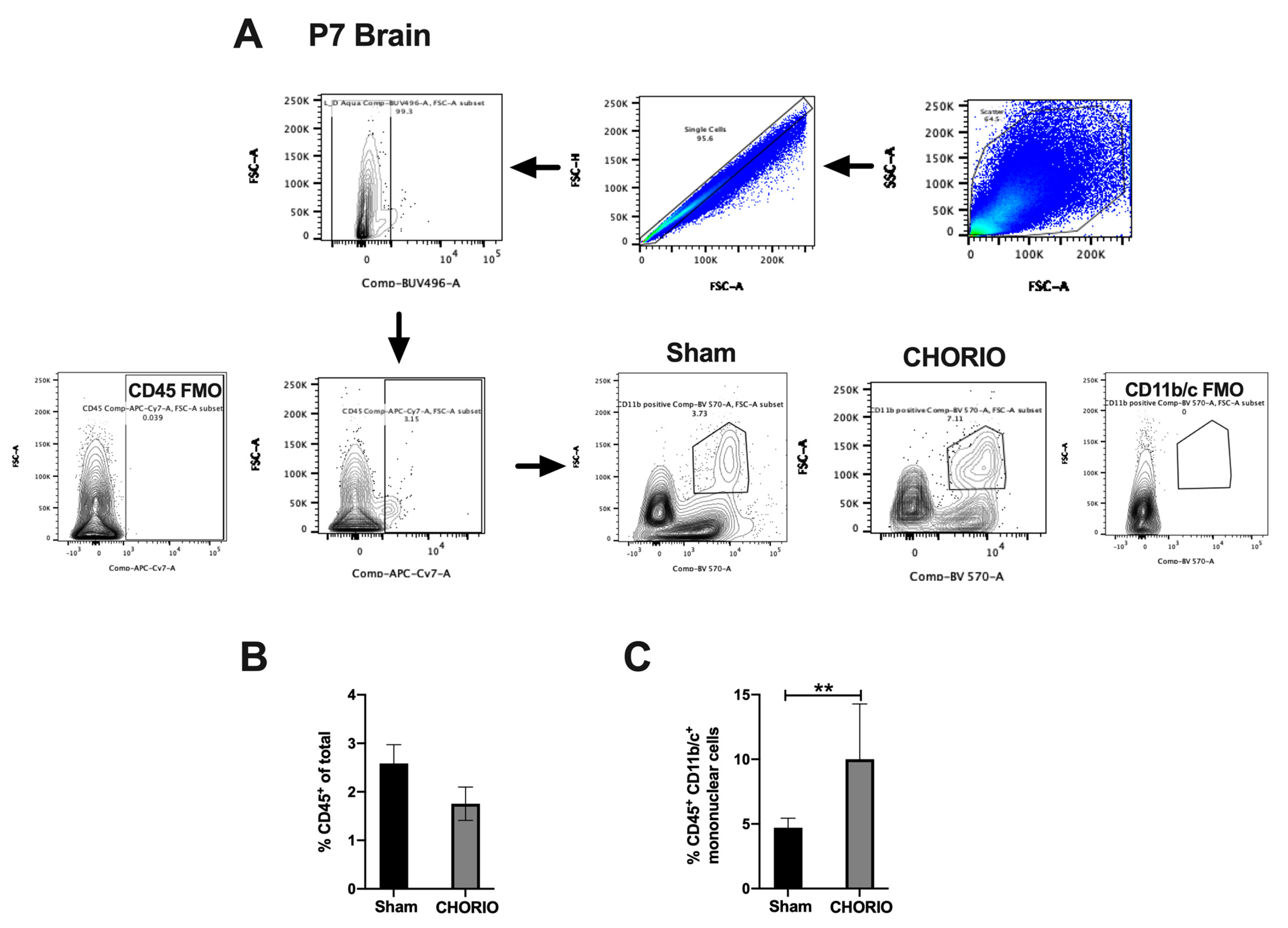
2.3.2. CHORIO-Induced Increase in Mononuclear Cells in Brain at P7
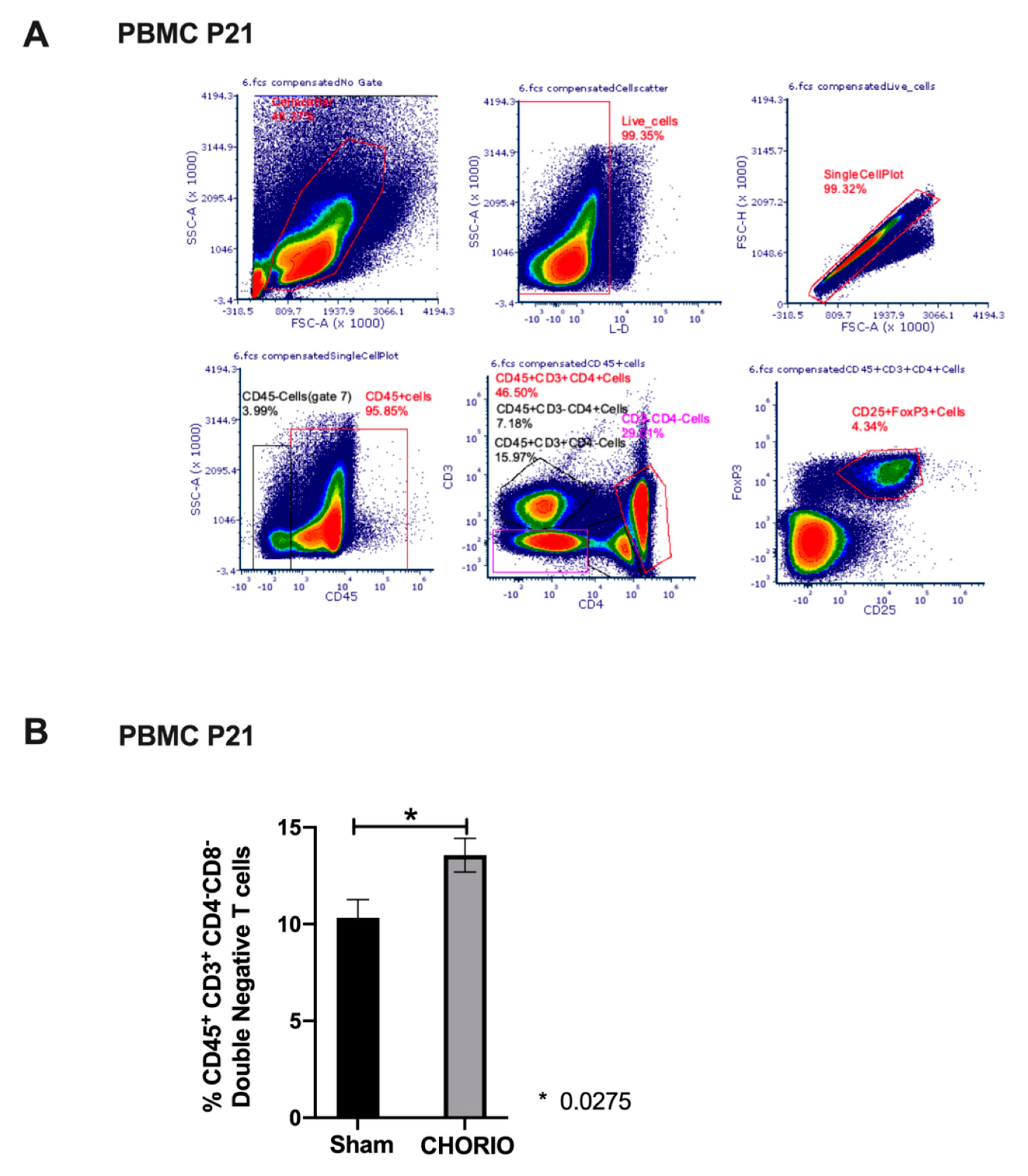
2.4. Peripheral Blood T Cell Composition at P21 after CHORIO
3. Discussions
4. Conclusions
5. Materials and Methods
5.1. Institutional Animal Care and Use Committee (IACUC) Approval
5.2. Induction of Chorioamnionitis
5.3. Tissue Collection
5.4. Extraction of RNA and Quantitative Real Time Polymerase Chain Reaction (RT-qPCR)
5.4.1. Isolation of Single Cells (Brain)
5.4.2. Isolation of Single Cells (Blood)
5.5. Multiparameter Flow Cytometry
5.6. Identification of Immune Cells and Phenotyping
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, L.W.; Lin, Y.C.; Wang, S.T.; Yeh, T.F.; Huang, C.C. Hypoxic/ischemic and infectious events have cumulative effects on the risk of cerebral palsy in very-low-birth-weight preterm infants. Neonatology 2014, 106, 209–215. [Google Scholar] [CrossRef]
- Pappas, A.; Kendrick, D.E.; Shankaran, S.; Stoll, B.J.; Bell, E.F.; Laptook, A.R.; Walsh, M.C.; Das, A.; Hale, E.C.; Newman, N.S.; et al. Chorioamnionitis and early childhood outcomes among extremely low-gestational-age neonates. JAMA Pediatr. 2014, 168, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Leviton, A.; Allred, E.N.; Kuban, K.C.; Hecht, J.L.; Onderdonk, A.B.; O’Shea, T.M.; Paneth, N. Microbiologic and histologic characteristics of the extremely preterm infant’s placenta predict white matter damage and later cerebral palsy. the ELGAN study. Pediatr. Res. 2010, 67, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute chorioamnionitis and funisitis: Definition, pathologic features, and clinical significance. Am. J. Obstet. Gynecol. 2015, 213, S29–S52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Kim, J.S. Chronic inflammation of the placenta: Definition, classification, pathogenesis, and clinical significance. Am. J. Obstet. Gynecol. 2015, 213, S53–S69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, E.; Romero, R.; Yeo, L.; Diaz-Primera, R.; Marin-Concha, J.; Para, R.; Lopez, A.M.; Pacora, P.; Gomez-Lopez, N.; Yoon, B.H.; et al. The fetal inflammatory response syndrome: The origins of a concept, pathophysiology, diagnosis, and obstetrical implications. Semin. Fetal Neonatal Med. 2020, 25, 101146. [Google Scholar] [CrossRef]
- Schipper, H.M.; Song, W.; Tavitian, A.; Cressatti, M. The sinister face of heme oxygenase-1 in brain aging and disease. Prog. Neurobiol. 2019, 172, 40–70. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Omata, Y.; Sakamoto, H.; Higashimoto, Y.; Hara, T.; Sagara, Y.; Noguchi, M. Characterization of rat heme oxygenase-3 gene. Implication of processed pseudogenes derived from heme oxygenase-2 gene. Gene 2004, 336, 241–250. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef]
- Ozen, M.; Zhao, H.; Lewis, D.B.; Wong, R.J.; Stevenson, D.K. Heme oxygenase and the immune system in normal and pathological pregnancies. Front. Pharmacol. 2015, 6, 84. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Ozen, M.; Wong, R.J.; Stevenson, D.K. Heme oxygenase-1 in pregnancy and cancer: Similarities in cellular invasion, cytoprotection, angiogenesis, and immunomodulation. Front. Pharmacol. 2014, 5, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozzelino, R.; Jeney, V.; Soares, M.P. Mechanisms of cell protection by heme oxygenase-1. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 323–354. [Google Scholar] [CrossRef] [Green Version]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttner, D.M.; Dennery, P.A. Reversal of HO-1 related cytoprotection with increased expression is due to reactive iron. FASEB J. 1999, 13, 1800–1809. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.C.; Chiang, S.K.; Chen, S.E.; Yu, Y.L.; Chou, R.H.; Chang, W.C. Heme oxygenase-1 mediates BAY 11–7085 induced ferroptosis. Cancer Lett. 2018, 416, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.Y.; Park, E.; Lee, S.J.; Chung, S.W. Heme oxygenase-1 accelerates erastin-induced ferroptotic cell death. Oncotarget 2015, 6, 24393–24403. [Google Scholar] [CrossRef] [Green Version]
- Andrews, M.; Leiva, E.; Arredondo-Olguin, M. Short repeats in the heme oxygenase 1 gene promoter is associated with increased levels of inflammation, ferritin and higher risk of type-2 diabetes mellitus. J. Trace Elem. Med. Biol. 2016, 37, 25–30. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef]
- Feng, H.; Schorpp, K.; Jin, J.; Yozwiak, C.E.; Hoffstrom, B.G.; Decker, A.M.; Rajbhandari, P.; Stokes, M.E.; Bender, H.G.; Csuka, J.M.; et al. Transferrin Receptor Is a Specific Ferroptosis Marker. Cell Rep. 2020, 30, 3411–3423.e7. [Google Scholar] [CrossRef] [Green Version]
- Jabara, H.H.; Boyden, S.E.; Chou, J.; Ramesh, N.; Massaad, M.J.; Benson, H.; Bainter, W.; Fraulino, D.; Rahimov, F.; Sieff, C.; et al. A missense mutation in TFRC, encoding transferrin receptor 1, causes combined immunodeficiency. Nat. Genet. 2016, 48, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, K.; Kalish, F.; Zhao, H.; Wong, R.J.; Stevenson, D.K. Heme oxygenase-1 deficiency promotes severity of sepsis in a non-surgical preterm mouse model. Pediatr. Res. 2018, 84, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Schulz, S.; Chisholm, K.M.; Zhao, H.; Kalish, F.; Yang, Y.; Wong, R.J.; Stevenson, D.K. Heme oxygenase-1 confers protection and alters T-cell populations in a mouse model of neonatal intestinal inflammation. Pediatr. Res. 2015, 77, 640–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Wong, R.J.; Nguyen, X.; Kalish, F.; Mizobuchi, M.; Vreman, H.J.; Stevenson, D.K.; Contag, C.H. Expression and regulation of heme oxygenase isozymes in the developing mouse cortex. Pediatr. Res. 2006, 60, 518–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, J.R.; Denson, J.L.; Joste, N.E.; Robinson, S.; Jantzie, L.L. Combined in utero hypoxia-ischemia and lipopolysaccharide administration in rats induces chorioamnionitis and a fetal inflammatory response syndrome. Placenta 2015, 36, 1378–1384. [Google Scholar] [CrossRef]
- Jantzie, L.L.; Miller, R.H.; Robinson, S. Erythropoietin signaling promotes oligodendrocyte development following prenatal systemic hypoxic-ischemic brain injury. Pediatr. Res. 2013, 74, 658–667. [Google Scholar] [CrossRef] [Green Version]
- Jantzie, L.L.; Corbett, C.J.; Berglass, J.; Firl, D.J.; Flores, J.; Mannix, R.; Robinson, S. Complex pattern of interaction between in utero hypoxia-ischemia and intra-amniotic inflammation disrupts brain development and motor function. J. Neuroinflamm. 2014, 11, 131. [Google Scholar] [CrossRef] [Green Version]
- Jantzie, L.L.; Winer, J.L.; Maxwell, J.R.; Chan, L.A.; Robinson, S. Modeling Encephalopathy of Prematurity Using Prenatal Hypoxia-ischemia with Intra-amniotic Lipopolysaccharide in Rats. J. Vis. Exp. 2015, 105, 53196. [Google Scholar] [CrossRef] [Green Version]
- Yellowhair, T.R.; Noor, S.; Mares, B.; Jose, C.; Newville, J.C.; Maxwell, J.R.; Northington, F.J.; Milligan, E.D.; Robinson, S.; Jantzie, L.L. Chorioamnionitis in Rats Precipitates Extended Postnatal Inflammatory Lymphocyte Hyperreactivity. Dev. Neurosci. 2019, 1–11. [Google Scholar] [CrossRef]
- Jennifer, B.; Berg, V.; Modak, M.; Puck, A.; Seyerl-Jiresch, M.; Künig, S.; Zlabinger, G.J.; Steinberger, P.; Chou, J.; Geha, R.S.; et al. Transferrin receptor 1 is a cellular receptor for human heme-albumin. Commun. Biol. 2020, 3, 621. [Google Scholar] [CrossRef]
- Lin, C.Y.; Chang, Y.C.; Wang, S.T.; Lee, T.Y.; Lin, C.F.; Huang, C.C. Altered inflammatory responses in preterm children with cerebral palsy. Ann. Neurol. 2010, 68, 204–212. [Google Scholar] [CrossRef]
- Kuban, K.C.; Joseph, R.; O’Shea, T.M.; Heeren, T.; Fichorova, R.N.; Douglass, L.; Jara, H.; Frazier, J.A.; Hirtz, D.; Rollins, J.V.; et al. Circulating Inflammatory-Associated Proteins in the First Month of Life and Cognitive Impairment at Age 10 Years in Children Born Extremely Preterm. J. Pediatr. 2017, 180, 116–123.e1. [Google Scholar] [CrossRef] [Green Version]
- Volpe, J.J. The encephalopathy of prematurity—Brain injury and impaired brain development inextricably intertwined. Semin. Pediatr. Neurol. 2009, 16, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Silvestroff, L.; Franco, P.G.; Pasquini, J.M. Neural and oligodendrocyte progenitor cells: Transferrin effects on cell proliferation. ASN Neuro 2013, 5, e00107. [Google Scholar] [CrossRef] [PubMed]
- Moos, T.; Rosengren Nielsen, T.; Skjorringe, T.; Morgan, E.H. Iron trafficking inside the brain. J. Neurochem. 2007, 103, 1730–1740. [Google Scholar] [CrossRef]
- Cheli, V.T.; Correale, J.; Paez, P.M.; Pasquini, J.M. Iron Metabolism in Oligodendrocytes and Astrocytes, Implications for Myelination and Remyelination. ASN Neuro 2020, 12, 1759091420962681. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.F.; Claus, C.P.; Vreman, H.J.; Wong, R.J.; Noble-Haeusslein, L.J. Heme regulation in traumatic brain injury: Relevance to the adult and developing brain. J. Cereb. Blood Flow Metab. 2005, 25, 1401–1417. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, M.; Ferriero, D.M.; Sharp, F.R. Developmental expression of heme oxygenase-1 (HSP32) in rat brain: An immunocytochemical study. Brain Res. Dev. Brain Res. 1998, 105, 181–194. [Google Scholar] [CrossRef]
- Sun, Y.; Rotenberg, M.O.; Maines, M.D. Developmental expression of heme oxygenase isozymes in rat brain. Two HO-2 mRNAs are detected. J. Biol. Chem. 1990, 265, 8212–8217. [Google Scholar] [CrossRef]
- van Tilborg, E.; de Theije, C.G.M.; van Hal, M.; Wagenaar, N.; de Vries, L.S.; Benders, M.J.; Rowitch, D.H.; Nijboer, C.H. Origin and dynamics of oligodendrocytes in the developing brain: Implications for perinatal white matter injury. Glia 2018, 66, 221–238. [Google Scholar] [CrossRef]
- Guardia Clausi, M.; Pasquini, L.A.; Soto, E.F.; Pasquini, J.M. Apotransferrin-induced recovery after hypoxic/ischaemic injury on myelination. ASN Neuro 2010, 2, e00048. [Google Scholar]
- Guardia Clausi, M.; Paez, P.M.; Campagnoni, A.T.; Pasquini, L.A.; Pasquini, J.M. Intranasal administration of aTf protects and repairs the neonatal white matter after a cerebral hypoxic-ischemic event. Glia 2012, 60, 1540–1554. [Google Scholar] [CrossRef] [PubMed]
- Benjamins, J.A.; Morell, P. Proteins of myelin and their metabolism. Neurochem. Res. 1978, 3, 137–174. [Google Scholar] [CrossRef] [PubMed]
- Rueda, C.M.; Wells, C.B.; Gisslen, T.; Jobe, A.H.; Kallapur, S.G.; Chougnet, C.A. Effect of chorioamnionitis on regulatory T cells in moderate/late preterm neonates. Hum. Immunol. 2015, 76, 65–73. [Google Scholar] [CrossRef] [Green Version]
- D’Acquisto, F.; Crompton, T. CD3+CD4-CD8-(double negative) T cells: Saviours or villains of the immune response? Biochem. Pharmacol. 2011, 82, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Meng, H.; Zhao, H.; Cao, X.; Hao, J.; Zhang, H.; Liu, Y.; Zhu, M.S.; Fan, L.; Weng, L.; Qian, L.; et al. Double-negative T cells remarkably promote neuroinflammation after ischemic stroke. Proc. Natl. Acad. Sci. USA 2019, 116, 5558–5563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yellowhair, T.R.; Noor, S.; Maxwell, J.R.; Anstine, C.V.; Oppong, A.Y.; Robinson, S.; Milligan, E.D.; Jantzie, L.L. Preclinical chorioamnionitis dysregulates CXCL1/CXCR2 signaling throughout the placental-fetal-brain axis. Exp. Neurol. 2018, 301, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Yellowhair, T.R.; Newville, J.C.; Noor, S.; Maxwell, J.R.; Milligan, E.D.; Robinson, S.; Jantzie, L.L. CXCR2 Blockade Mitigates Neural Cell. Injury Following Preclinical Chorioamnionitis. Front. Physiol. 2019, 10, 324. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer Sequence |
|---|---|
| HO-1 (Inducible HO) | (F)5′CCTTCCCGAACATCGACAGCC3′ (R)5′GCAGCTCCTCAAACAGCTCAA3′ |
| HO-2 (Constitutive HO) | (F)5′GGAGGGGGTAGATGAGTCAGA3′ (R)5′TCGGTCATGTGCTTCCTTGGT3′ |
| TfR1 (Transferrin R) | (F)5′AGTTGAACAAAGTGGCACGAGCAG3′ (R)5′AGCAGTTGGCTGTTGTACCTCTCA3′ |
| 18 S | (F)5′TCCCTAGTGATCCCCGAGAAGT3′ (R)5′CCCTTAATGGCAGTGATAGCGA3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozen, M.; Kitase, Y.; Vasan, V.; Burkhardt, C.; Ramachandra, S.; Robinson, S.; Jantzie, L.L. Chorioamnionitis Precipitates Perinatal Alterations of Heme-Oxygenase-1 (HO-1) Homeostasis in the Developing Rat Brain. Int. J. Mol. Sci. 2021, 22, 5773. https://doi.org/10.3390/ijms22115773
Ozen M, Kitase Y, Vasan V, Burkhardt C, Ramachandra S, Robinson S, Jantzie LL. Chorioamnionitis Precipitates Perinatal Alterations of Heme-Oxygenase-1 (HO-1) Homeostasis in the Developing Rat Brain. International Journal of Molecular Sciences. 2021; 22(11):5773. https://doi.org/10.3390/ijms22115773
Chicago/Turabian StyleOzen, Maide, Yuma Kitase, Vikram Vasan, Christopher Burkhardt, Sindhu Ramachandra, Shenandoah Robinson, and Lauren L. Jantzie. 2021. "Chorioamnionitis Precipitates Perinatal Alterations of Heme-Oxygenase-1 (HO-1) Homeostasis in the Developing Rat Brain" International Journal of Molecular Sciences 22, no. 11: 5773. https://doi.org/10.3390/ijms22115773