
Involvement of Chaperone Sigma1R in the Anxiolytic Effect of Fabomotizole

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
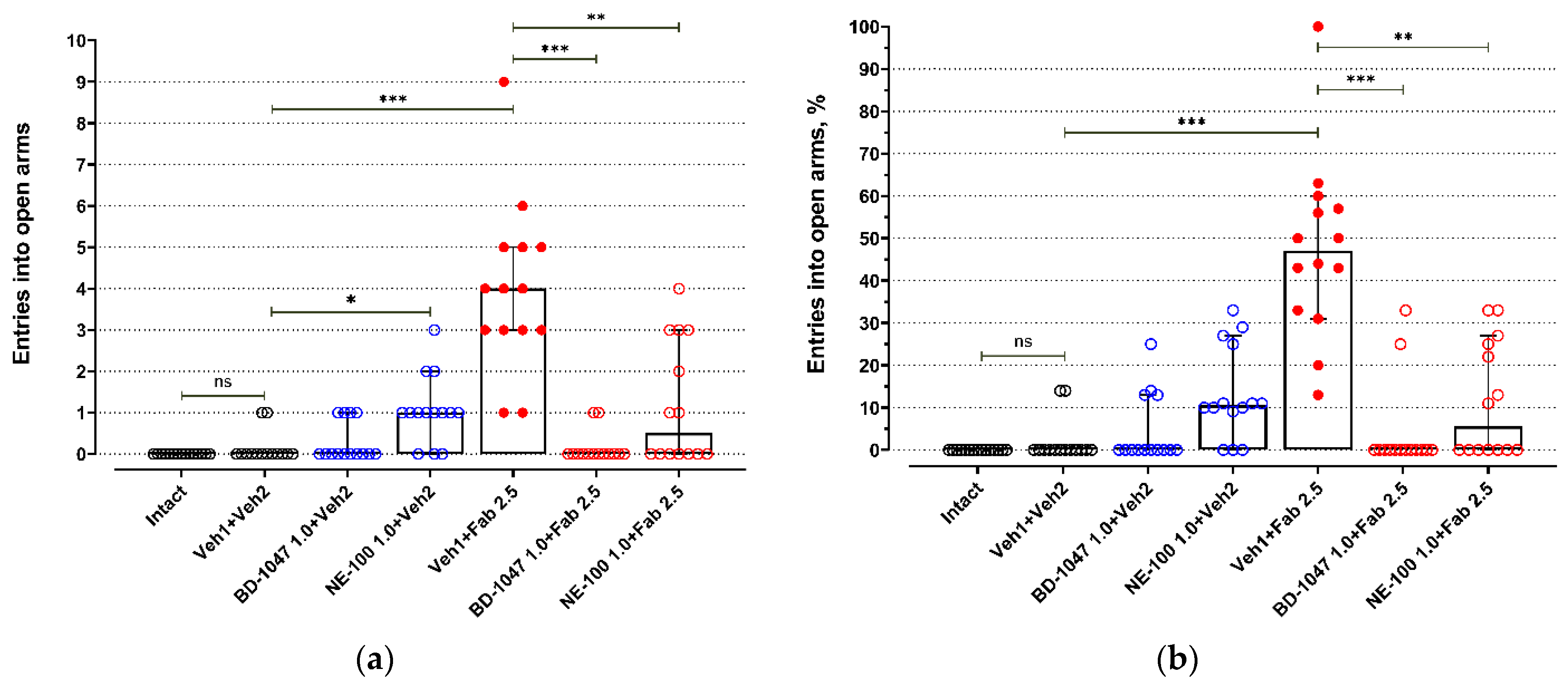
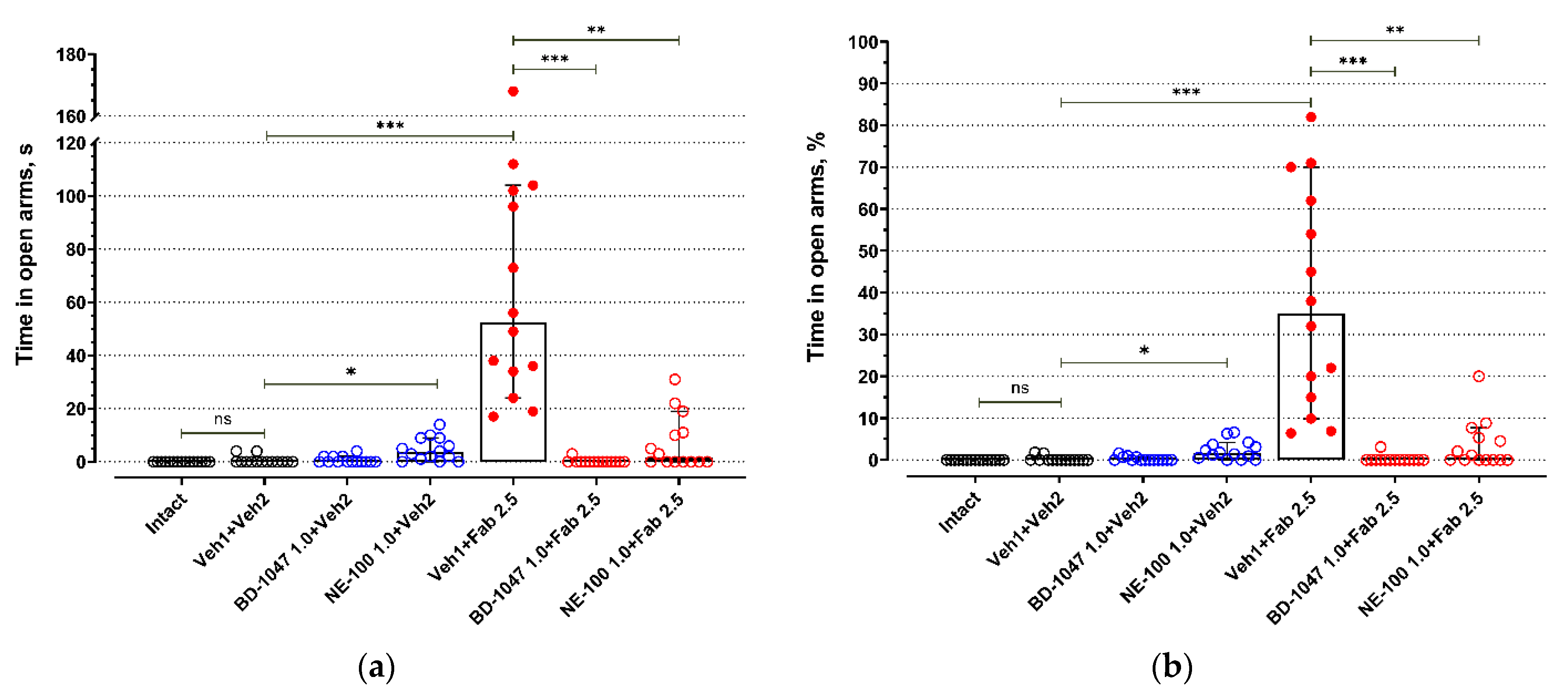
2.1. Elevated Plus Maze Test
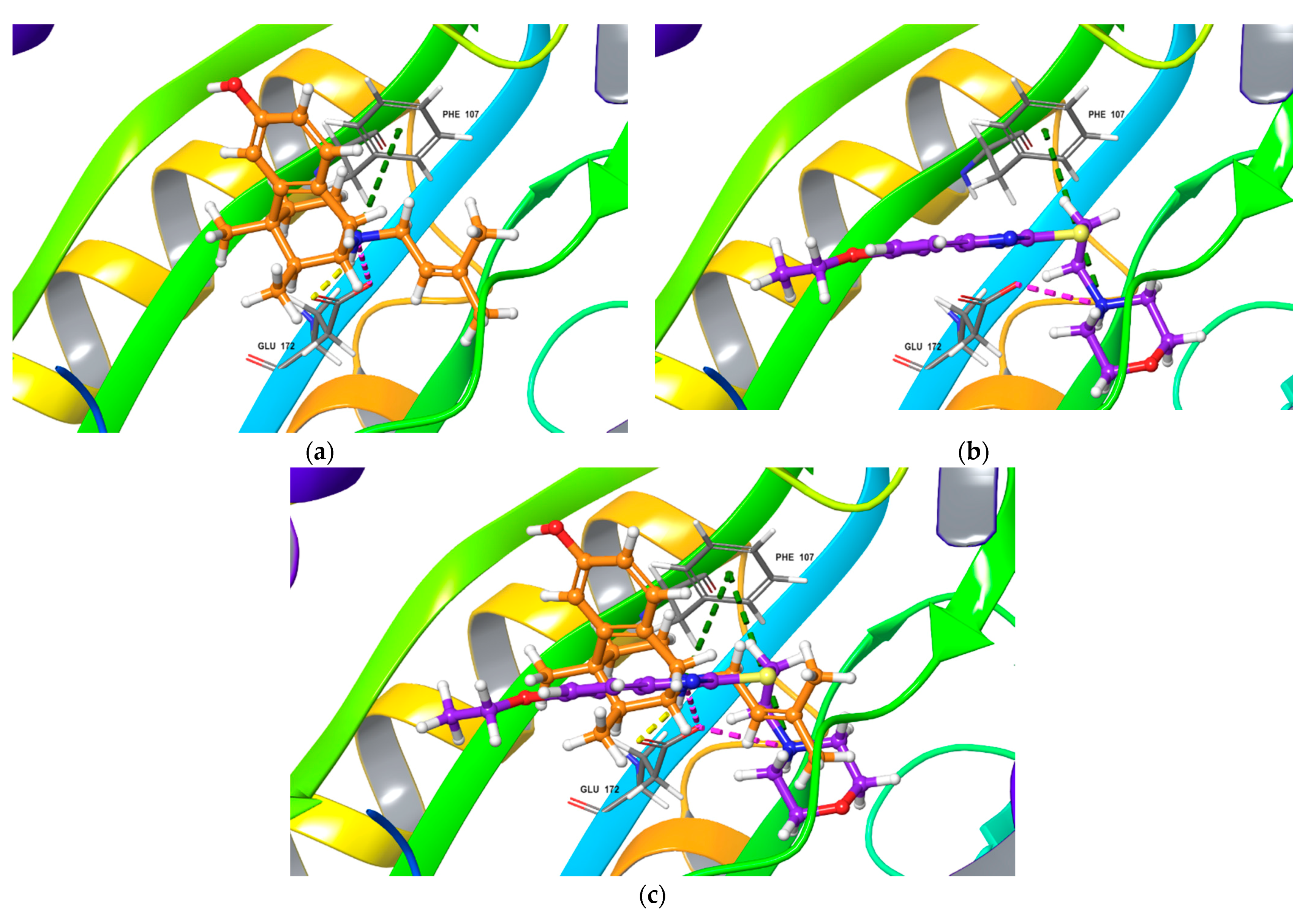
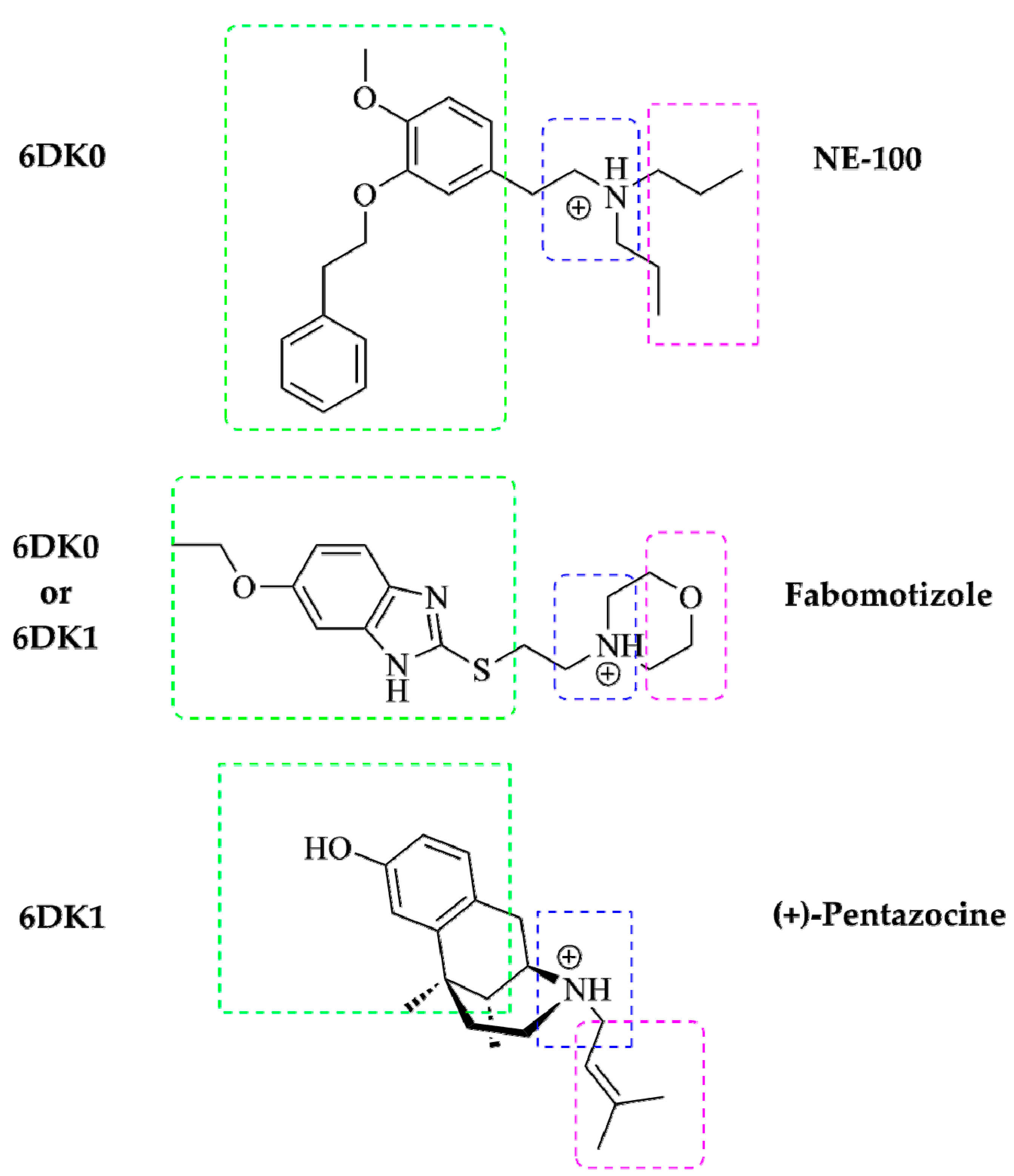
2.2. In Silico Docking Study

3. Discussion
- Compounds with antagonistic activity to Sigma1R prevent the development of anxiolytic-like action of fabomotizole;
- Fabomotizole interaction mode with Sigma1R is similar to that observed for known Sigma1R ligands. Fabomotizole engages Glu172 residue to form an electrostatic interaction with substitute amino groups;
- Fabomotizole is located in the 6DK1 binding site of agonist (+)-pentazocine, completely reproducing the mode of its interaction with key amino acids; ΔGbind values calculated for fabomotizole and (+)-pentazocine are similar (the difference is only −3.85 kcal/mol).
4. Materials and Methods
4.1. In Vivo Study
4.1.1. Chemicals
4.1.2. Experimental Animals
4.1.3. Ethical Approval
4.1.4. In Vivo Experimental Design
4.1.5. Elevated Plus Maze Test
4.1.6. Statistical Analysis
4.2. Molecular Docking
4.2.1. Set of Compounds and RCSB Protein Data Bank Codes
4.2.2. Ligand Preparation
4.2.3. Protein Preparation
4.2.4. Docking Protocols and Calculations
4.2.5. Docking Study Design
- Docking (Glide XP + MM_GBSA) of fabomotizole into the structures of Sigma1R bound to (+)-pentazocine (PDB ID: 6DK1) and bound to NE-100 (PDB ID: 6DK0) with comparison of calculated parameters;
- Calculation of the binding energies (ΔGbind) for the best binding poses of fabomotizole and reference ligands, using Glide XP + MM_GBSA approach; calculation of ΔΔGbind values as a difference between ΔGbind/ref and ΔGbind/fab—one of the major characteristics for evaluation of similarity of fabomotizole to one of the tested references Sigma1R ligands;
- Comparisons of the pattern of interactions with the amino acids of the Sigma1R binding site and location inside of Sigma1R binding site and binding mode between fabomotizole and reference ligands (analysis of hydrogen bond interactions, π-π stacking interaction, π-cation, and salt bridge formations).
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-IBP | N-(1-benzylpiperidin-4-yl)-4-iodobenzamide |
| 5-HT | 5-hydroxytryptamine; serotonin |
| ASIC | Acid-sensing ion channel |
| BD-1047 | N′-[2-(3,4-dichlorophenyl)ethyl]-N,N,N′-trimethylethane-1,2-diamine |
| BD-1063 | 1-[2-(3,4-dichlorophenyl)ethyl]-4-methylpiperazine |
| BDNF | Brain-derived neurotrophic factor; human BDNF gene |
| BiP | Endoplasmic reticulum chaperone BiP |
| BMY-14802 | 1-(4-fluorophenyl)-4-[4-(5-fluoropyrimidin-2-yl)piperazin-1-yl]butan-1-ol |
| CB1R | Cannabinoid receptor 1 |
| Cnr1 | Rodent cannabinoid receptor 1 gene |
| D1R | D(1A) dopamine receptor |
| D2R | D(2) dopamine receptor |
| DAT | Sodium-dependent dopamine transporter |
| ΔGbind | The binding free energy |
| DHEA | Dehydroepiandrosterone |
| DHEAS | Dehydroepiandrosterone sulfate |
| EPM | Elevated plus maze test |
| ER | Endoplasmic reticulum |
| FAB | Fabomotizole; 4-[2-[(6-ethoxy-1H-benzimidazol-2-yl)sulfanyl]ethyl]morpholine dihydrochloride |
| GABA | gamma-aminobutyric acid type; 4-aminobutanoic acid |
| GluN | Glutamate receptor ionotropic, NMDA |
| HINT1 | Histidine triad nucleotide-binding protein 1 |
| IRE1 | Serine/threonine-protein kinase/endoribonuclease IRE1 |
| JO-1784 | (E)-N-(cyclopropylmethyl)-N-methyl-3,6-diphenylhex-5-en-3-amine hydrochloride |
| MAM | Mitochondria-associated membranes |
| MAO-A | Amine oxidase [flavin-containing] A |
| MT3 receptor | Melatonin receptor, type 3 |
| NE-100 | N-[2-[4-methoxy-3-(2-phenylethoxy)phenyl]ethyl]-N-propylpropan-1-amine |
| nH | Hill coefficient |
| NMDA | (2R)-2-(methylamino)butanedioic acid |
| NQO2 | NRH:quinone oxidoreductase 2; ribosyldihydronicotinamide dehydrogenase [quinone] |
| NPY | Pro-neuropeptide Y |
| NRH | dihydronicotinamide riboside; |
| 1-[(2R,3R,4S,5R)-3,4-dihydroxy-5-(hydroxymethyl)oxolan-2-yl]-4H-pyridine-3-carboxamide | |
| OF | Open field test |
| PTZ | (+)-pentazocine; (1S,9S,13S)-1,13-dimethyl-10-(3-methylbut-2-enyl)-10-azatricyclo[7.3.1.02,7]trideca-2(7),3,5-trien-4-ol |
| PCP | phencyclidine; 1-(1-phenylcyclohexyl)piperidine |
| PD144418 | 3-(4-methylphenyl)-5-(1-propyl-3,6-dihydro-2H-pyridin-5-yl)-1,2-oxazole |
| PDB | Protein Data Bank |
| PRE-084 | 2-morpholin-4-ylethyl 1-phenylcyclohexane-1-carboxylate |
| PREGS | Pregnenolone sulfate |
| PROG | Progesterone |
| PTSD | Post-traumatic stress disorder |
| RCSB | Research Collaboratory for Structural Bioinformatics |
| Sigma1R | Sigma nonopioid intracellular receptor 1; chaperon Sigma1R |
| Sigmar1 | Rodent Sigma1R gene |
| (+)-SKF 10.047 | (1S,9S,13S)-1,13-dimethyl-10-prop-2-enyl-10-azatricyclo[7.3.1.02,7]trideca-2(7),3,5-trien-4-ol |
| RMSD | Root-mean-square deviation |
| SNRI | Serotonin-noradrenaline reuptake inhibitor |
| SSRI | Selective serotonin reuptake inhibitor |
| UPR | Unfolded protein response |
| XBP1 | X-box-binding protein 1 |
References
- Craske, M.G.; Stein, M.B.; Eley, T.C.; Milad, M.R.; Holmes, A.; Rapee, R.; Wittchen, H.-U. Anxiety disorders. Nat. Rev. Dis. Prim. 2017, 3, 17024. [Google Scholar] [CrossRef]
- Penninx, B.W.J.H.; Pine, D.S.; Holmes, E.A.; Reif, A. Anxiety disorders. Lancet 2021, 397, 880. [Google Scholar] [CrossRef]
- Baxter, A.J.; Scott, K.M.; Vos, T.; Whiteford, H.A. Global prevalence of anxiety disorders: A systematic review and meta-regression. Psychol. Med. 2013, 43, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Petukhova, M.; Sampson, N.A.; Zaslavsky, A.M.; Wittchen, H.-U. Twelve-month and lifetime prevalence and lifetime morbid risk of anxiety and mood disorders in the United States. Int. J. Methods Psychiatr. Res. 2012, 21, 169–184. [Google Scholar] [CrossRef]
- Guo, X.; Meng, Z.; Huang, G.; Fan, J.; Zhou, W.; Ling, W.; Jiang, J.; Long, J.; Su, L. Meta-analysis of the prevalence of anxiety disorders in mainland China from 2000 to 2015. Sci. Rep. 2016, 6, 28033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirneshan, E.; Bailey, J.; Relyea, G.; Franklin, B.E.; Solomon, D.K.; Brown, L.M. Incremental direct medical expenditures associated with anxiety disorders for the U.S. adult population: Evidence from the Medical Expenditure Panel Survey. J. Anxiety Disord. 2013, 27, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Konnopka, A.; König, H. Economic Burden of Anxiety Disorders: A Systematic Review and Meta-Analysis. PharmacoEconomics 2020, 38, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xiao, W.-M.; Chen, Y.-K.; Qu, J.-F.; Liu, Y.-L.; Fang, X.-W.; Weng, H.-Y.; Luo, G.-P. Anxiety in Patients with Acute Ischemic Stroke: Risk Factors and Effects on Functional Status. Front. Psychiatry 2019, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Berge, L.I.; Skogen, J.C.; Sulo, G.; Igland, J.; Wilhelmsen, I.; Vollset, S.E.; Tell, G.S.; Knudsen, A.K. Health anxiety and risk of ischaemic heart disease: A prospective cohort study linking the Hordaland Health Study (HUSK) with the Cardiovascular Diseases in Norway (CVDNOR) project. BMJ Open 2016, 6, e012914. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-C.; Hu, Y.-W.; Hu, L.-Y.; Hung, M.-H.; Su, T.-P.; Huang, M.-W.; Tsai, C.-F.; Ou, S.-M.; Yen, S.-H.; Tzeng, C.-H.; et al. The Risk of Cancer in Patients with Generalized Anxiety Disorder: A Nationwide Population-Based Study. PLoS ONE 2013, 8, e57399. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; Deschênes, S.S.; Schmitz, N. Investigating the longitudinal association between diabetes and anxiety: A systematic review and meta-analysis. Diabet. Med. 2018, 35, 677–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donoghue, J.; Lader, M. Usage of benzodiazepines: A review. Int. J. Psychiatry Clin. Pract. 2010, 14, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.D.; Landon, B.E. Patterns in Outpatient Benzodiazepine Prescribing in the United States. JAMA Netw. Open 2019, 2, e187399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dell’Osso, B.; Lader, M. Do Benzodiazepines Still Deserve a Major Role in The Treatment of Psychiatric Disorders? A Critical Reappraisal. Eur. Psychiatry 2013, 28, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Seredenin, S.B.; Voronina, T.; Neznamov, G.G.; Blednov, I.; Badyshtov, B.; Viglinskaia, I.V.; Kozlovskaia, M.M.; Kolotilinskaia, N.V.; Iarkova, M.; Savel’Ev, V.L.; et al. The pharmacogenetic concept of the anxio-selective effect. Ann. Russ. Acad. Med. Sci. 1998, 11, 3–9. [Google Scholar]
- Soyka, M. Treatment of Benzodiazepine Dependence. N. Engl. J. Med. 2017, 376, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.; Duffy, L.; Ades, A.; Amos, R.; Araya, R.; Brabyn, S.; Button, K.S.; Churchill, R.; Derrick, C.; Dowrick, C.; et al. The clinical effectiveness of sertraline in primary care and the role of depression severity and duration (PANDA): A pragmatic, double-blind, placebo-controlled randomised trial. Lancet Psychiatry 2019, 6, 903–914. [Google Scholar] [CrossRef] [Green Version]
- El Hage, W.; Leman, S.; Camus, V.; Belzung, C. Mechanisms of antidepressant resistance. Front. Pharmacol. 2013, 4, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griebel, G.; Holmes, A. 50 years of hurdles and hope in anxiolytic drug discovery. Nat. Rev. Drug Discov. 2013, 12, 667–687. [Google Scholar] [CrossRef] [Green Version]
- Grillon, C.; Ernst, M. A way forward for anxiolytic drug development: Testing candidate anxiolytics with anxiety-potentiated startle in healthy humans. Neurosci. Biobehav. Rev. 2020, 119, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Bandelow, B. Current and Novel Psychopharmacological Drugs for Anxiety Disorders. Adv. Exp. Med. Biol. 2020, 1191, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Sartori, S.B.; Singewald, N. Novel pharmacological targets in drug development for the treatment of anxiety and anxiety-related disorders. Pharmacol. Ther. 2019, 204, 107402. [Google Scholar] [CrossRef] [PubMed]
- Sakata, M.; Ishiwata, K. Imaging of Sigma1 Receptors in the Human Brain Using PET and [11C]SA4503. Central Nerv. Syst. Agents Med. Chem. 2009, 9, 190–196. [Google Scholar] [CrossRef]
- Alonso, G.; Phan, V.-L.; Guillemain, I.; Saunier, M.; Legrand, A.; Anoal, M.; Maurice, T. Immunocytochemical localization of the sigma1 receptor in the adult rat central nervous system. Neuroscience 2000, 97, 155–170. [Google Scholar] [CrossRef]
- Guitart, X.; Codony, X.; Monroy, X. Sigma receptors: Biology and therapeutic potential. Psychopharmacology 2004, 174, 301–319. [Google Scholar] [CrossRef] [PubMed]
- James, M.L.; Shen, B.; Zavaleta, C.L.; Nielsen, C.H.; Mesangeau, C.; Vuppala, P.K.; Chan, C.; Avery, B.A.; Fishback, J.A.; Matsumoto, R.R.; et al. New Positron Emission Tomography (PET) Radioligand for Imaging σ-1 Receptors in Living Subjects. J. Med. Chem. 2012, 55, 8272–8282. [Google Scholar] [CrossRef]
- Bhuiyan, S.; Tagashira, H.; Fukunaga, K. Crucial interactions between selective serotonin uptake inhibitors and sigma-1 receptor in heart failure. J. Pharmacol. Sci. 2013, 121, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.; Bai, P.; Chen, Z.; Neelamegam, R.; Placzek, M.S.; Wang, H.; Fiedler, S.A.; Yang, J.; Yuan, G.; Qu, X.; et al. Novel radioligands for imaging sigma-1 receptor in brain using positron emission tomography (PET). Acta Pharm. Sin. B 2019, 9, 1204–1215. [Google Scholar] [CrossRef] [PubMed]
- Chevallier, N.; Keller, E.; Maurice, T. Behavioural phenotyping of knockout mice for the sigma-1 (σ1) chaperone protein revealed gender-related anxiety, depressive-like and memory alterations. J. Psychopharmacol. 2010, 25, 960–975. [Google Scholar] [CrossRef]
- Ji, L.-L.; Peng, J.-B.; Fu, C.-H.; Tong, L.; Wang, Z.-Y. Sigma-1 receptor activation ameliorates anxiety-like behavior through NR2A-CREB-BDNF signaling pathway in a rat model submitted to single-prolonged stress. Mol. Med. Rep. 2017, 16, 4987–4993. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.-L.; Peng, J.-B.; Fu, C.-H.; Cao, D.; Li, D.; Tong, L.; Wang, Z.-Y. Activation of Sigma-1 receptor ameliorates anxiety-like behavior and cognitive impairments in a rat model of post-traumatic stress disorder. Behav. Brain Res. 2016, 311, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Ossa, F.; Schnell, J.R.; Ortega-Roldan, J.L. A Review of the Human Sigma-1 Receptor Structure. Chem. Biol. Pteridines Folates 2017, 964, 15–29. [Google Scholar] [CrossRef]
- Kruse, A. Structural Insights into Sigma1 Function. Organotypic Models Drug Dev. 2016, 244, 13–25. [Google Scholar] [CrossRef]
- Schmidt, H.R.; Zheng, S.; Gurpinar, E.; Koehl, A.; Manglik, A.K.A.; Kruse, A.C. Crystal structure of the human σ1 receptor. Nature 2016, 532, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.R.; Betz, R.M.; Dror, R.O.; Kruse, A.C. Structural basis for σ1 receptor ligand recognition. Nat. Struct. Mol. Biol. 2018, 25, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Alon, A.; Schmidt, H.; Zheng, S.; Kruse, A.C. Structural Perspectives on Sigma-1 Receptor Function. Chem. Biol. Pteridines Folates 2017, 964, 5–13. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.-P. Cholesterol at the Endoplasmic Reticulum: Roles of the Sigma-1 Receptor Chaperone and Implications thereof in Human Diseases. Prokaryotic Cytoskelet. 2010, 51, 381–398. [Google Scholar] [CrossRef] [Green Version]
- Delprat, B.; Crouzier, L.; Su, T.-P.; Maurice, T. At the Crossing of ER Stress and MAMs: A Key Role of Sigma-1 Receptor? Adv. Exp. Med. Biol. 2020, 1131, 699–718. [Google Scholar] [CrossRef]
- Su, T.-P.; Su, T.-C.; Nakamura, Y.; Tsai, S.-Y. The Sigma-1 Receptor as a Pluripotent Modulator in Living Systems. Trends Pharmacol. Sci. 2016, 37, 262–278. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Torres, M.; Sha, H.; Halbrook, C.J.; Bergh, F.V.D.; Reinert, R.B.; Yamada, T.; Wang, S.; Luo, Y.; Hunter, A.H.; et al. Endoplasmic reticulum–associated degradation regulates mitochondrial dynamics in brown adipocytes. Science 2020, 368, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.-P. Intracellular Dynamics of σ-1 Receptors (σ1 Binding Sites) in NG108-15 Cells. J. Pharmacol. Exp. Ther. 2003, 306, 726–733. [Google Scholar] [CrossRef]
- Hayashi, T.; Su, T.-P. σ-1 Receptors (σ1 Binding Sites) Form Raft-Like Microdomains and Target Lipid Droplets on the Endoplasmic Reticulum: Roles in Endoplasmic Reticulum Lipid Compartmentalization and Export. J. Pharmacol. Exp. Ther. 2003, 306, 718–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Fujimoto, M. Detergent-Resistant Microdomains Determine the Localization of σ-1 Receptors to the Endoplasmic Reticulum-Mitochondria Junction. Mol. Pharmacol. 2010, 77, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Zhemkov, V.; Ditlev, J.; Lee, W.-R.; Wilson, M.; Liou, J.; Rosen, M.K.; Bezprozvanny, I. The role of sigma 1 receptor in organization of endoplasmic reticulum signaling microdomains. eLife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Muñoz, M.; Sánchez-Blázquez, P.; Herrero-Labrador, R.; Martínez-Murillo, R.; Merlos, M.; Vela, J.M.; Garzón, J. The σ1 Receptor Engages the Redox-Regulated HINT1 Protein to Bring Opioid Analgesia under NMDA Receptor Negative Control. Antioxid. Redox Signal. 2015, 22, 799–818. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Muñoz, M.; Sánchez-Blázquez, P.; Callado, L.F.; Meana, J.J.; Garzón-Niño, J. Schizophrenia and depression, two poles of endocannabinoid system deregulation. Transl. Psychiatry 2017, 7, 1291. [Google Scholar] [CrossRef] [Green Version]
- Carnally, S.M.; Johannessen, M.; Henderson, R.M.; Jackson, M.B.; Edwardson, J.M. Demonstration of a Direct Interaction between σ-1 Receptors and Acid-Sensing Ion Channels. Biophys. J. 2010, 98, 1182–1191. [Google Scholar] [CrossRef] [Green Version]
- Herrera, Y.; Katnik, C.; Rodriguez, J.D.; Hall, A.A.; Willing, A.; Pennypacker, K.R.; Cuevas, J. σ-1 Receptor Modulation of Acid-Sensing Ion Channel a (ASIC1a) and ASIC1a-Induced Ca2+ Influx in Rat Cortical Neurons. J. Pharmacol. Exp. Ther. 2008, 327, 491–502. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Rizzo, S.J.S.; Neal, S.J.; Lin, Q.; Jow, F.; Arias, R.L.; Rosenzweig-Lipson, S.; Dunlop, J.; Beyer, C.E. Acid sensing ion channel (ASIC) inhibitors exhibit anxiolytic-like activity in preclinical pharmacological models. Psychopharmacology 2009, 203, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.-H. Stress and the dopaminergic reward system. Exp. Mol. Med. 2020, 52, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Nikolaus, S.; Mamlins, E.; Hautzel, H.; Müller, H.-W. Acute anxiety disorder, major depressive disorder, bipolar disorder and schizophrenia are related to different patterns of nigrostriatal and mesolimbic dopamine dysfunction. Rev. Neurosci. 2019, 30, 381–426. [Google Scholar] [CrossRef]
- Kamei, H.; Noda, Y.; Kameyama, T.; Nabeshima, T. Role of (+)-SKF-10,047-sensitive sub-population of σ1 receptors in amelioration of conditioned fear stress in rats: Association with mesolimbic dopaminergic systems. Eur. J. Pharmacol. 1997, 319, 165–172. [Google Scholar] [CrossRef]
- Navarro, G.; Moreno, E.; Aymerich, M.; Marcellino, D.; McCormick, P.J.; Mallol, J.; Cortes, A.; Casado, V.; Canela, E.I.; Ortiz, J.; et al. Direct involvement of -1 receptors in the dopamine D1 receptor-mediated effects of cocaine. Proc. Natl. Acad. Sci. USA 2010, 107, 18676–18681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, G.; Moreno, E.; Bonaventura, J.; Brugarolas, M.; Farré, D.; Aguinaga, D.; Mallol, J.; Cortés, A.; Casadó, V.; Lluís, C.; et al. Cocaine Inhibits Dopamine D2 Receptor Signaling via Sigma-1-D2 Receptor Heteromers. PLoS ONE 2013, 8, e61245. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.C.; Yano, H.; Hiranita, T.; Chin, F.T.; McCurdy, C.R.; Su, T.-P.; Amara, S.; Katz, J.L. The sigma-1 receptor modulates dopamine transporter conformation and cocaine binding and may thereby potentiate cocaine self-administration in rats. J. Biol. Chem. 2017, 292, 11250–11261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedoce, A.D.G.; Ferreira, F.; Bota, R.G.; Bonet-Costa, V.; Sun, P.Y.; Davies, K.J.A. The role of oxidative stress in anxiety disorder: Cause or consequence? Free. Radic. Res. 2018, 52, 737–750. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T. Conversion of psychological stress into cellular stress response: Roles of the sigma-1 receptor in the process. Psychiatry Clin. Neurosci. 2015, 69, 179–191. [Google Scholar] [CrossRef]
- Goguadze, N.; Zhuravliova, E.; Morin, D.; Mikeladze, D.; Maurice, T. Sigma-1 Receptor Agonists Induce Oxidative Stress in Mitochondria and Enhance Complex I Activity in Physiological Condition but Protect Against Pathological Oxidative Stress. Neurotox. Res. 2019, 35, 1–18. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, C.-S. Sigma-1 receptor activation ameliorates LPS-induced NO production and ROS formation through the Nrf2/HO-1 signaling pathway in cultured astrocytes. Neurosci. Lett. 2019, 711, 134387. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.-P. Sigma-1 Receptor Chaperones at the ER- Mitochondrion Interface Regulate Ca2+ Signaling and Cell Survival. Cell 2007, 131, 596–610. [Google Scholar] [CrossRef] [Green Version]
- Mori, T.; Hayashi, T.; Hayashi, E.; Su, T.-P. Sigma-1 Receptor Chaperone at the ER-Mitochondrion Interface Mediates the Mitochondrion-ER-Nucleus Signaling for Cellular Survival. PLoS ONE 2013, 8, e76941. [Google Scholar] [CrossRef] [Green Version]
- Saito, A.; Cai, L.; Matsuhisa, K.; Ohtake, Y.; Kaneko, M.; Kanemoto, S.; Asada, R.; Imaizumi, K. Neuronal activity-dependent local activation of dendritic unfolded protein response promotes expression of brain-derived neurotrophic factor in cell soma. J. Neurochem. 2017, 144, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syunyakov, T.S.; Neznamov, G.G. Evaluation of the therapeutic efficacy and safety of the selective anxiolytic afobazole in generalized anxiety disorder and adjustment disorders: Results of a multicenter randomized comparative study of diazepam. Ter. Arkhiv 2016, 88, 73–86. [Google Scholar] [CrossRef]
- Voronin, M.V.; Kadnikov, I.A. Contribution of Sigma-1 receptor to cytoprotective effect of afobazole. Pharmacol. Res. Perspect. 2016, 4, e00273. [Google Scholar] [CrossRef] [Green Version]
- Katnik, C.; Garcia, A.; Behensky, A.A.; Yasny, I.E.; Shuster, A.M.; Seredenin, S.B.; Petrov, A.V.; Cuevas, J. Activation of σ1 and σ2 receptors by afobazole increases glial cell survival and prevents glial cell activation and nitrosative stress after ischemic stroke. J. Neurochem. 2016, 139, 497–509. [Google Scholar] [CrossRef]
- Katnik, C.; Garcia, A.; Behensky, A.; Yasny, I.; Shuster, A.; Seredenin, S.; Petrov, A.; Seifu, S.; McAleer, J.; Willing, A.; et al. Treatment with afobazole at delayed time points following ischemic stroke improves long-term functional and histological outcomes. Neurobiol. Dis. 2014, 62, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Behensky, A.A.; Yasny, I.E.; Shuster, A.M.; Seredenin, S.B.; Petrov, A.V.; Cuevas, J. Afobazole Activation of σ-1 Receptors Modulates Neuronal Responses to Amyloid-β25–35. J. Pharmacol. Exp. Ther. 2013, 347, 468–477. [Google Scholar] [CrossRef]
- Behensky, A.A.; Yasny, I.E.; Shuster, A.M.; Seredenin, S.B.; Petrov, A.V.; Cuevas, J. Stimulation of Sigma Receptors with Afobazole Blocks Activation of Microglia and Reduces Toxicity Caused by Amyloid-β25–35. J. Pharmacol. Exp. Ther. 2013, 347, 458–467. [Google Scholar] [CrossRef]
- Cuevas, J.; Rodriguez, A.; Behensky, A.; Katnik, C. Afobazole Modulates Microglial Function via Activation of Both σ-1 and σ-2 Receptors. J. Pharmacol. Exp. Ther. 2011, 339, 161–172. [Google Scholar] [CrossRef]
- Cuevas, J.; Behensky, A.; Deng, W.; Katnik, C. Afobazole Modulates Neuronal Response to Ischemia and Acidosis via Activation of σ-1 Receptors. J. Pharmacol. Exp. Ther. 2011, 339, 152–160. [Google Scholar] [CrossRef]
- Voronin, M.V.; Kadnikov, I.A.; Voronkov, D.N.; Seredenin, S.B. Chaperone Sigma1R mediates the neuroprotective action of afobazole in the 6-OHDA model of Parkinson’s disease. Sci. Rep. 2019, 9, 17020. [Google Scholar] [CrossRef]
- Voronin, M.V.; Kadnikov, I.A.; Seredenin, S.B. Afobazole Restores the Dopamine Level in a 6-Hydroxydopamine Model of Parkinson’s Disease. Neurochem. J. 2019, 13, 49–56. [Google Scholar] [CrossRef]
- Kadnikov, I.A.; Verbovaya, E.R.; Voronkov, D.N.; Voronin, M.V.; Seredenin, S.B. Deferred Administration of Afobazole Induces Sigma1R-Dependent Restoration of Striatal Dopamine Content in a Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 7620. [Google Scholar] [CrossRef] [PubMed]
- Seredenin, S.B.; Molodavkin, G.M.; Voronin, M.V.; Voronina, T. Antidepressant properties of afobazole in Porsolt and Nomura tests. Eksperimental'naia Klin. Farmakol. 2009, 72, 19–21. [Google Scholar]
- Seredenin, S.B.; Garibova, T.; Kuznetsova, A.L.; Voronin, M.V.; Iarkova, M.; Voronina, T. Afobazole decreases motor side effects induced by haloperidol. Eksperimental'naia Klin. Farmakol. 2009, 72, 15–18. [Google Scholar]
- Seredenin, S.B.; Voronin, M.V. Neuroreceptor mechanisms of the afobazole effect. Eksperimental'naia Klin. Farmakol. 2009, 72, 3–11. [Google Scholar]
- Ryaskina, E.V.; Voronin, M.V.; Seredenin, S.B. Interaction of 2-mercaptobenzimidazole derivatives with sigma-1 receptors. Pharm. Chem. J. 2012, 46, 334–336. [Google Scholar] [CrossRef]
- Abramova, E.V.; Voronin, M.V.; Seredenin, S.B. Interaction of Afobazole with Sigma-1 Receptors in the Mouse Brain. Pharm. Chem. J. 2015, 49, 7–9. [Google Scholar] [CrossRef]
- Iarkova, M. Analysis of the binding capacity of the benzodiazepine site of gabaa receptor in mice C57BL/6 and BALB/C pretreated with anxiolytics. Eksperimental'naia Klin. Farmakol. 2011, 74, 3–7. [Google Scholar]
- Seredenin, S.B.; Antipova, T.A.; Voronin, M.V.; Kurchashova, S.Y.; Kuimov, A.N. Interaction of Afobazole with σ1-Receptors. Bull. Exp. Biol. Med. 2009, 148, 42–44. [Google Scholar] [CrossRef]
- Ryskamp, D.A.; Zhemkov, V.; Bezprozvanny, I. Mutational Analysis of Sigma-1 Receptor’s Role in Synaptic Stability. Front. Neurosci. 2019, 13, 1012. [Google Scholar] [CrossRef] [Green Version]
- Mavlyutov, T.A.; Yang, H.; Epstein, M.L.; Ruoho, A.E.; Yang, J.; Guo, L.-W. APEX2-enhanced electron microscopy distinguishes sigma-1 receptor localization in the nucleoplasmic reticulum. Oncotarget 2017, 8, 51317–51330. [Google Scholar] [CrossRef]
- Yamamoto, H.; Miura, R.; Yamamoto, T.; Shinohara, K.; Watanabe, M.; Okuyama, S.; Nakazato, A.; Nukada, T. Amino acid residues in the transmembrane domain of the type 1 sigma receptor critical for ligand binding. FEBS Lett. 1999, 445, 19–22. [Google Scholar] [CrossRef] [Green Version]
- Seth, P.; Ganapathy, M.; Conway, S.J.; Bridges, C.D.; Smith, S.B.; Casellas, P.; Ganapathy, V. Expression pattern of the type 1 sigma receptor in the brain and identity of critical anionic amino acid residues in the ligand-binding domain of the receptor. Biochim. Biophys. Acta (BBA) Bioenerg. 2001, 1540, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Boström, J.; Greenwood, J.R.; Gottfries, J. Assessing the performance of OMEGA with respect to retrieving bioactive conformations. J. Mol. Graph. Model. 2003, 21, 449–462. [Google Scholar] [CrossRef]
- Velázquez-Libera, J.L.; Rossino, G.; Navarro-Retamal, C.; Collina, S.; Caballero, J. Docking, Interaction Fingerprint, and Three-Dimensional Quantitative Structure–Activity Relationship (3D-QSAR) of Sigma1 Receptor Ligands, Analogs of the Neuroprotective Agent RC-33. Front. Chem. 2019, 7, 496. [Google Scholar] [CrossRef] [Green Version]
- Glennon, R. Pharmacophore Identification for Sigma-1 (σ1) Receptor Binding: Application of the “Deconstruction–Reconstruction–Elaboration” Approach. Mini-Rev. Med. Chem. 2005, 5, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 1985, 14, 149–167. [Google Scholar] [CrossRef]
- File, S. Factors controlling measures of anxiety and responses to novelty in the mouse. Behav. Brain Res. 2001, 125, 151–157. [Google Scholar] [CrossRef]
- Varty, G.; Morgan, C.; Cohen-Williams, M.; Coffin, V.L.; Carey, G.J. The Gerbil Elevated Plus-Maze I Behavioral Characterization and Pharmacological Validation. Neuropsychopharmacology 2002, 27, 357–370. [Google Scholar] [CrossRef]
- LaLonde, R.; Strazielle, C. Relations between open-field, elevated plus-maze, and emergence tests in C57BL/6J and BALB/c mice injected with GABA- and 5HT-anxiolytic agents. Fundam. Clin. Pharmacol. 2009, 24, 365–376. [Google Scholar] [CrossRef]
- Holoubek, G. Specific modulation of sigma binding sites by the anxiolytic drug opipramol. J. Neural Transm. 2003, 110, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Emamghoreishi, M.; Shahpari, M.; Keshavarz, M. Interaction of sigma-1 receptor modulators with seizure development in pentylenetetrazole-induced kindled mice. Epilepsy Res. 2019, 154, 74–76. [Google Scholar] [CrossRef]
- Müller, W.; Siebert, B.; Holoubek, G.; Gentsch, C. Neuropharmacology of the Anxiolytic Drug Opripramol, a Sigma Site Ligand. Pharmacopsychiatry 2004, 37, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Maren, S.; Holmes, A. Stress and Fear Extinction. Neuropsychopharmacology 2016, 41, 58–79. [Google Scholar] [CrossRef] [Green Version]
- Kamei, H.; Kameyama, T.; Nabeshima, T. SKF-10,047 reverses stress-induced motor suppression: Interaction with dopaminergic system. Eur. J. Pharmacol. 1994, 260, 39–46. [Google Scholar] [CrossRef]
- Kamei, H.; Kameyama, T.; Nabeshima, T. (+)-SKF-10,047 and dextromethorphan ameliorate conditioned fear stress through the activation of phenytoin-regulated σ1 sites. Eur. J. Pharmacol. 1996, 299, 21–28. [Google Scholar] [CrossRef]
- Noda, Y.; Kamei, H.; Nagai, T.; Nishida, M.; Nabeshima, T. Neurosteroids Ameliorate Conditioned Fear Stress an Association with Sigma1 Receptors. Neuropsychopharmacology 2000, 23, 276–284. [Google Scholar] [CrossRef]
- Urani, A.; Romieu, P.; Roman, F.J.; Yamada, K.; Noda, Y.; Kamei, H.; Tran, H.M.; Nagai, T.; Nabeshima, T.; Maurice, T. Enhanced antidepressant efficacy of σ1 receptor agonists in rats after chronic intracerebroventricular infusion of β-amyloid-(1–40) protein. Eur. J. Pharmacol. 2004, 486, 151–161. [Google Scholar] [CrossRef]
- Kamei, H.; Kameyama, T.; Nabeshima, T. (+)-SKF-10,047 and dextromethorphan ameliorate conditioned fear stress via dopaminergic systems linked to phenytoin-regulated σ1 sites. Eur. J. Pharmacol. 1996, 309, 149–158. [Google Scholar] [CrossRef]
- Gue, M.; Junien, J.L.; Del Rio, C.; Bueno, L. Neuropeptide Y and sigma ligand (JO 1784) suppress stress-induced colonic motor disturbances in rats through sigma and cholecystokinin receptors. J. Pharmacol. Exp. Ther. 1992, 261, 850–855. [Google Scholar] [PubMed]
- Rennekamp, A.J.; Huang, X.-P.; Wang, Y.; Patel, S.; Lorello, P.J.; Cade, L.; Gonzales, A.P.W.; Yeh, J.-R.J.; Caldarone, P.J.L.B.J.; Roth, X.-P.H.B.L.; et al. σ1 receptor ligands control a switch between passive and active threat responses. Nature 2016, 12, 552–558. [Google Scholar] [CrossRef]
- Seredin, S.B.; Melkumian, D.S.; Val’Dman, E.; Iarkova, M.; Seredina, T.C.; Voronin, M.V.; Lapitskaia, A.S. Effects of afobazole on the BDNF content in brain structures of inbred mice with different phenotypes of emotional stress reaction. Eksperimental'naia Klin. Farmakol. 2006, 69, 3–6. [Google Scholar]
- Egashira, N.; Harada, S.; Okuno, R.; Matsushita, M.; Nishimura, R.; Mishima, K.; Iwasaki, K.; Orito, K.; Fujiwara, M. Involvement of the sigma1 receptor in inhibiting activity of fluvoxamine on marble-burying behavior: Comparison with paroxetine. Eur. J. Pharmacol. 2007, 563, 149–154. [Google Scholar] [CrossRef] [PubMed]
- de Brouwer, G.; Fick, A.; Harvey, B.H.; Wolmarans, D.W. A critical inquiry into marble-burying as a preclinical screening paradigm of relevance for anxiety and obsessive–compulsive disorder: Mapping the way forward. Cogn. Affect. Behav. Neurosci. 2019, 19, 1–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voronin, M.V.; Vakhitova, Y.V.; Seredenin, S.B. Chaperone Sigma1R and Antidepressant Effect. Int. J. Mol. Sci. 2020, 21, 7088. [Google Scholar] [CrossRef]
- Nevels, R.M.; Gontkovsky, S.T.; Williams, B.E. Paroxetine—The Antidepressant from Hell? Probably Not, But Caution Required. Psychopharmacol. Bull. 2016, 46, 77–104. [Google Scholar]
- Ishima, T.; Fujita, Y.; Hashimoto, K. Interaction of new antidepressants with sigma-1 receptor chaperones and their potentiation of neurite outgrowth in PC12 cells. Eur. J. Pharmacol. 2014, 727, 167–173. [Google Scholar] [CrossRef]
- Narita, N.; Hashimoto, K.; Tomitaka, S.-I.; Minabe, Y. Interactions of selective serotonin reuptake inhibitors with subtypes of σ receptors in rat brain. Eur. J. Pharmacol. 1996, 307, 117–119. [Google Scholar] [CrossRef]
- Mizuno, T.; Yotsuyanagi, S.; Nagasaka, Y.; Namiki, M. Dehydroepiandrosterone Alleviates Copulatory Disorder Induced by Social Stress in Male Rats. J. Sex. Med. 2006, 3, 612–618. [Google Scholar] [CrossRef]
- Moriguchi, S.; Yamamoto, Y.; Ikuno, T.; Fukunaga, K. Sigma-1 receptor stimulation by dehydroepiandrosterone ameliorates cognitive impairment through activation of CaM kinase II, protein kinase C and extracellular signal-regulated kinase in olfactory bulbectomized mice. J. Neurochem. 2011, 117, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, S.; Shinoda, Y.; Yamamoto, Y.; Sasaki, Y.; Miyajima, K.; Tagashira, H.; Fukunaga, K. Stimulation of the Sigma-1 Receptor by DHEA Enhances Synaptic Efficacy and Neurogenesis in the Hippocampal Dentate Gyrus of Olfactory Bulbectomized Mice. PLoS ONE 2013, 8, e60863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, J.F.; Beltrán, D.; Cavas, M. Effects of (+) SKF 10,047, a sigma-1 receptor agonist, on anxiety, tested in two laboratory models in mice. Psicothema 2012, 24, 427–430. [Google Scholar] [PubMed]
- McCann, D.J.; Rabin, R.A.; Rens-Domiano, S.; Winter, J. Phencyclidine/SKF-10,047 binding sites: Evaluation of function. Pharmacol. Biochem. Behav. 1989, 32, 87–94. [Google Scholar] [CrossRef]
- Johnson, K.M.; Jones, S.M. Neuropharmacology of Phencyclidine: Basic Mechanisms and Therapeutic Potential. Annu. Rev. Pharmacol. Toxicol. 1990, 30, 707–750. [Google Scholar] [CrossRef]
- Turgeon, S.M.; Kim, D.; Pritchard, M.; Salgado, S.; Thaler, A. The effects of phencyclidine (PCP) on anxiety-like behavior in the elevated plus maze and the light-dark exploration test are age dependent, sexually dimorphic, and task dependent. Pharmacol. Biochem. Behav. 2011, 100, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Urani, A.; Roman, F.J.; Phan, V.L.; Su, T.P.; Maurice, T. The antidepressant-like effect induced by sigma(1)-receptor agonists and neuroactive steroids in mice submitted to the forced swimming test. J. Pharmacol. Exp. Ther. 2001, 298, 1269–1279. [Google Scholar]
- Maurice, T. Bi-phasic dose response in the preclinical and clinical developments of sigma-1 receptor ligands for the treatment of neurodegenerative disorders. Expert Opin. Drug Discov. 2021, 16, 373–389. [Google Scholar] [CrossRef]
- Ennaceur, A.; Chazot, P.L. Preclinical animal anxiety research—Flaws and prejudices. Pharmacol. Res. Perspect. 2016, 4, e00223. [Google Scholar] [CrossRef] [Green Version]
- Griebel, G.; Belzung, C.; Perrault, G.; Sanger, D.J. Differences in anxiety-related behaviours and in sensitivity to diazepam in inbred and outbred strains of mice. Psychopharmacology 2000, 148, 164–170. [Google Scholar] [CrossRef]
- GOST 33215-2014. Guidelines for Accommodation and Care of Animals. Environment, Housing and Management. Interstate Counsil for Standartization, Metrology and Certification 2016. Available online: http://protect.gost.ru/document.aspx?control=7&id=202494 (accessed on 21 May 2021).
- National Research Council (U.S.); Committee for the Update of the Guide for the Care and Use of Laboratory Animals; Institute for Laboratory Animal Research (U.S.); National Academies Press (U.S.). Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011; 220p. [Google Scholar]
- Berardi, F. A multireceptorial binding reinvestigation on an extended class of σ ligands: N-[ω-(indan-1-yl and tetralin-1-yl)alkyl] derivatives of 3,3-dimethylpiperidine reveal high affinities towards σ1 and EBP sites. Bioorganic Med. Chem. 2001, 9, 1325–1335. [Google Scholar] [CrossRef]
- Harder, E.; Damm, W.; Maple, J.R.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L.; et al. OPLS3: A Force Field Providing Broad Coverage of Drug-like Small Molecules and Proteins. J. Chem. Theory Comput. 2016, 12, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Shelley, J.C.; Cholleti, A.; Frye, L.L.; Greenwood, J.R.; Timlin, M.R.; Uchimaya, M. Epik: A software program for pK a prediction and protonation state generation for drug-like molecules. J. Comput. Mol. Des. 2007, 21, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.P.; Pincus, D.L.; Rapp, C.S.; Day, T.J.F.; Honig, B.; Shaw, D.E.; Friesner, R.A. A hierarchical approach to all-atom protein loop prediction. Proteins Struct. Funct. Bioinform. 2004, 55, 351–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, M.P.; Friesner, R.A.; Xiang, Z.; Honig, B. On the Role of the Crystal Environment in Determining Protein Side-chain Conformations. J. Mol. Biol. 2002, 320, 597–608. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein−Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 2. Enrichment Factors in Database Screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB ID | Ligand | Glide XP + MM_GBSA Mode | RMSD, Å | |||
|---|---|---|---|---|---|---|
| Residues in H-Bond Interaction | Other Interactions | ΔGbind, kcal/mol | ΔΔGbind, kcal/mol | |||
| 6DK0 | NE-100 | - | salt bridge: Glu172 π-cation: Phe107 | −74.76 | −13.49 | 1.032 |
| fabomotizole | Glu172, Trp164 | π-cation: Phe107 salt bridge: Glu172 CH-π: Tyr103 | −61.27 | - | ||
| 6DK1 | (+)-pentazocine | Glu172 | π -cation: Phe107 salt bridge: Glu172 | −60.50 | −3.85 | 0.301 |
| fabomotizole | Glu172 | π -cation: Phe107 salt bridge: Glu172 | −56.65 | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voronin, M.V.; Vakhitova, Y.V.; Tsypysheva, I.P.; Tsypyshev, D.O.; Rybina, I.V.; Kurbanov, R.D.; Abramova, E.V.; Seredenin, S.B. Involvement of Chaperone Sigma1R in the Anxiolytic Effect of Fabomotizole. Int. J. Mol. Sci. 2021, 22, 5455. https://doi.org/10.3390/ijms22115455
Voronin MV, Vakhitova YV, Tsypysheva IP, Tsypyshev DO, Rybina IV, Kurbanov RD, Abramova EV, Seredenin SB. Involvement of Chaperone Sigma1R in the Anxiolytic Effect of Fabomotizole. International Journal of Molecular Sciences. 2021; 22(11):5455. https://doi.org/10.3390/ijms22115455
Chicago/Turabian StyleVoronin, Mikhail V., Yulia V. Vakhitova, Inna P. Tsypysheva, Dmitry O. Tsypyshev, Inna V. Rybina, Rustam D. Kurbanov, Elena V. Abramova, and Sergei B. Seredenin. 2021. "Involvement of Chaperone Sigma1R in the Anxiolytic Effect of Fabomotizole" International Journal of Molecular Sciences 22, no. 11: 5455. https://doi.org/10.3390/ijms22115455