
Dysregulation of miR-15a-5p, miR-497a-5p and miR-511-5p Is Associated with Modulation of BDNF and FKBP5 in Brain Areas of PTSD-Related Susceptible and Resilient Mice

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Selection of miRNAs
2.2. Expression of miR-15a-5p, let-7d-5p, miR-497a-5p, miR-511-5p, and Their Potential Targets, BDNF and FKBP5
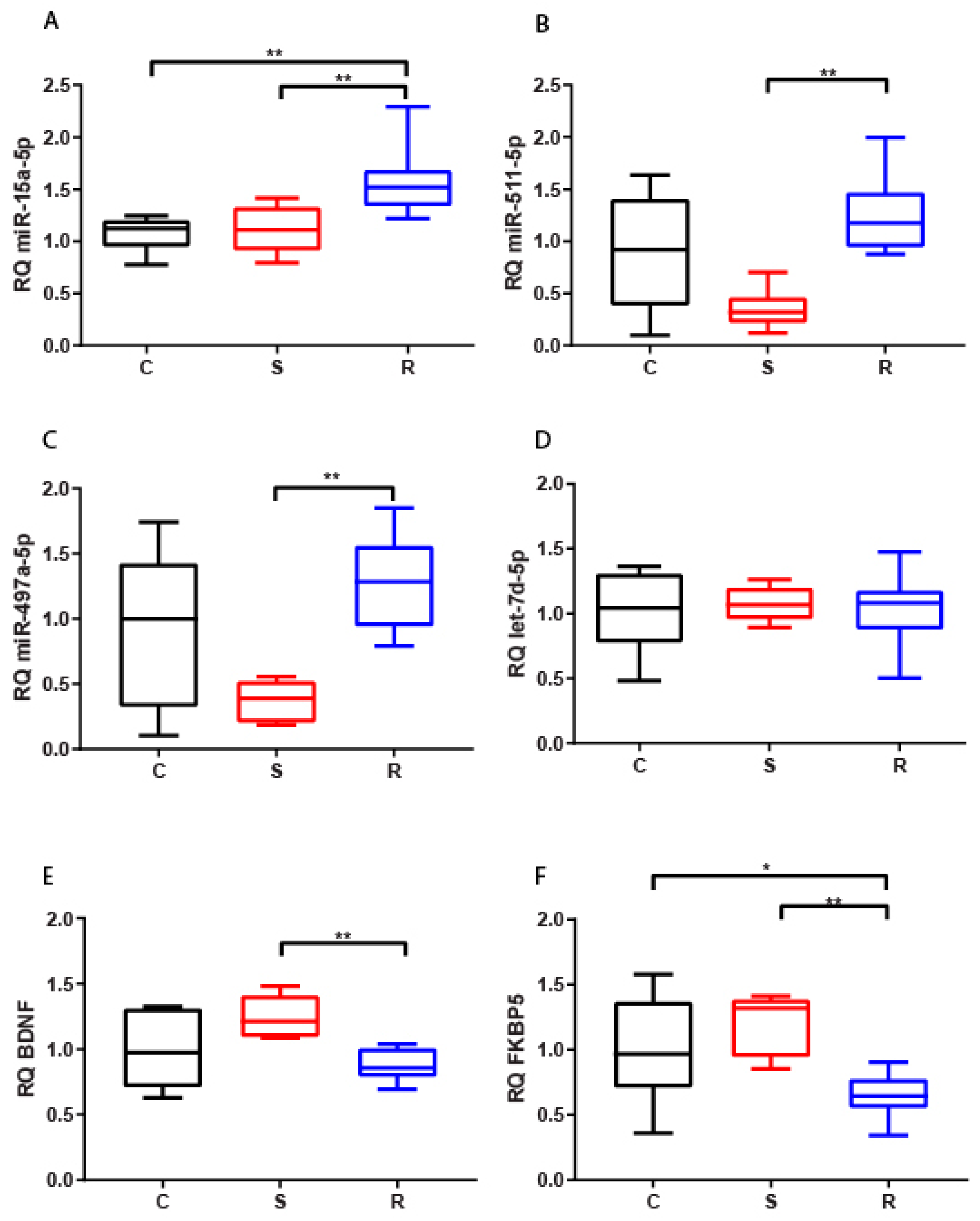
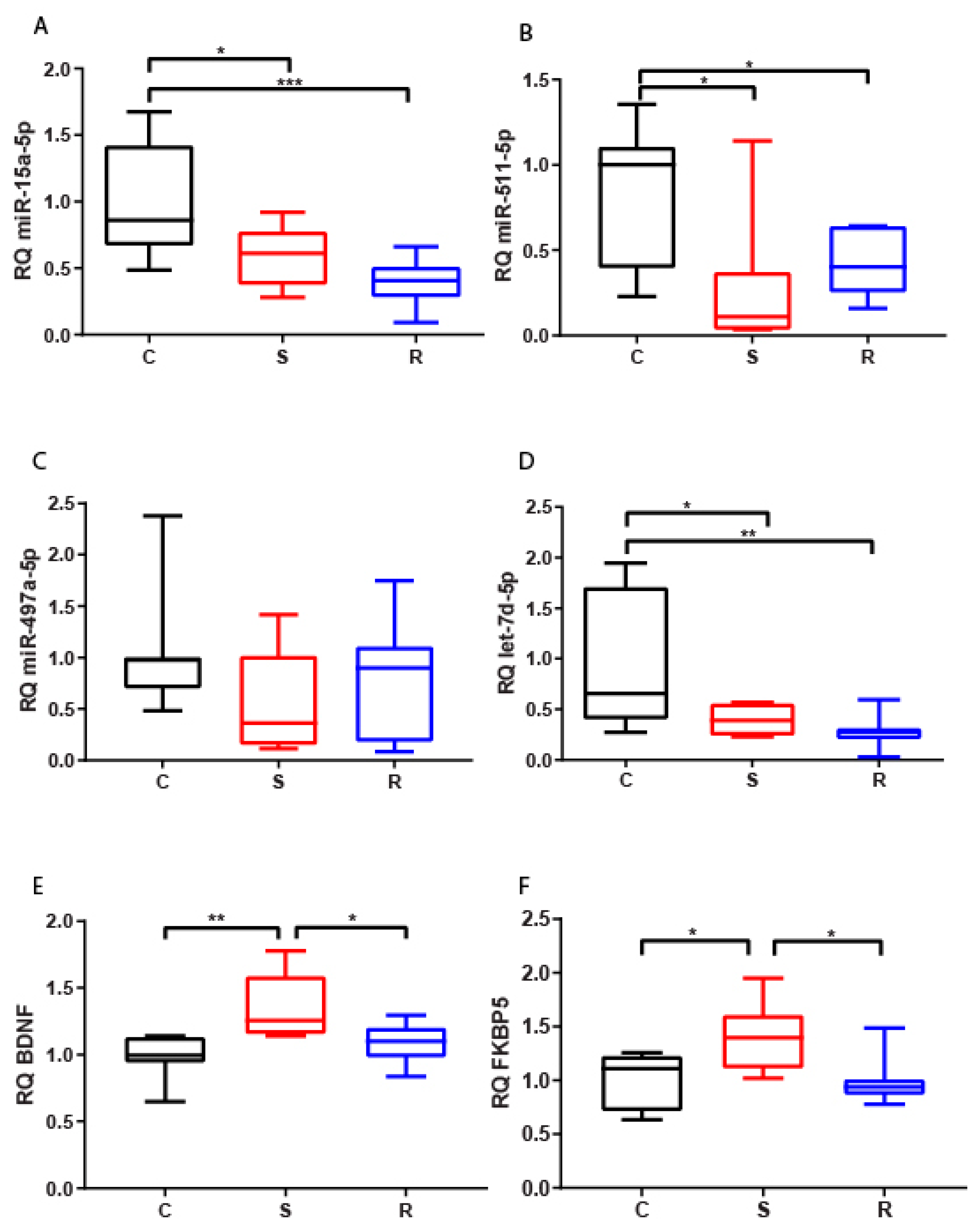
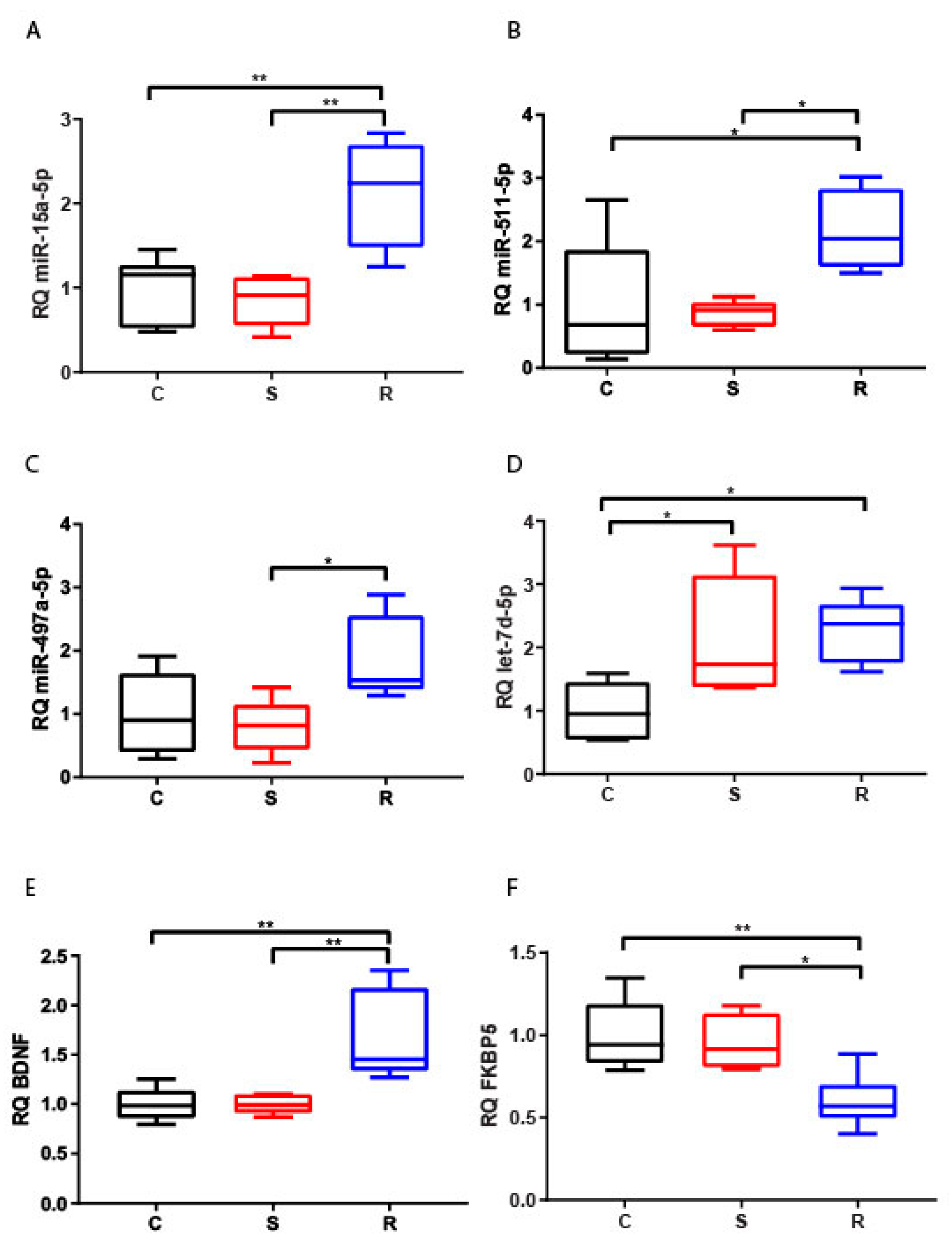
2.2.1. Hippocampus
2.2.2. Medial Prefrontal Cortex
2.2.3. Hypothalamus
2.2.4. Evaluation of Synergic Effect of miRNAs on FKBP5 and BDNF Expression
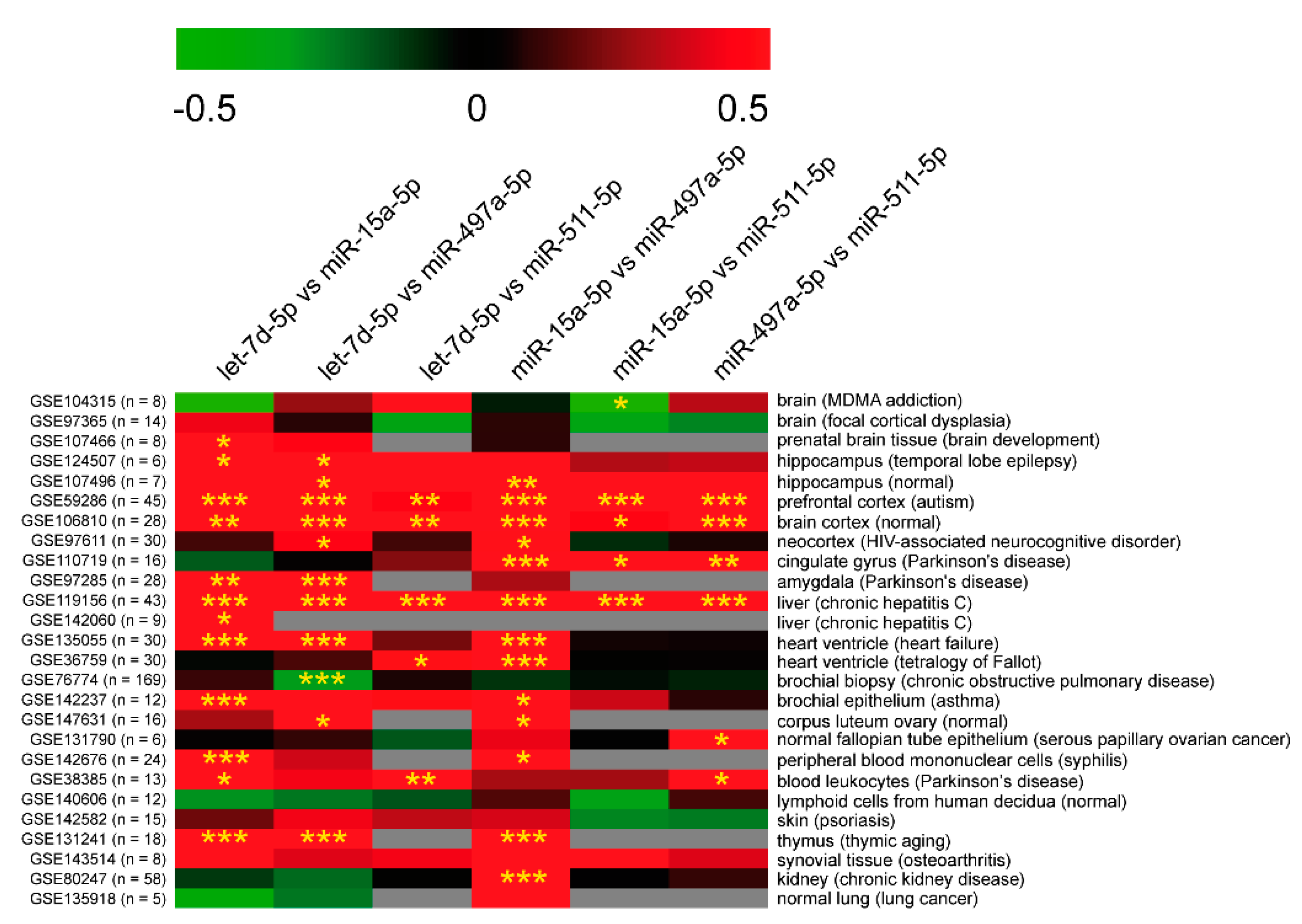
2.2.5. Potential Co-Regulation of PTSD-Related miRNAs
2.2.6. Correlation Analysis Between RNA Expression and Behavioral Scores
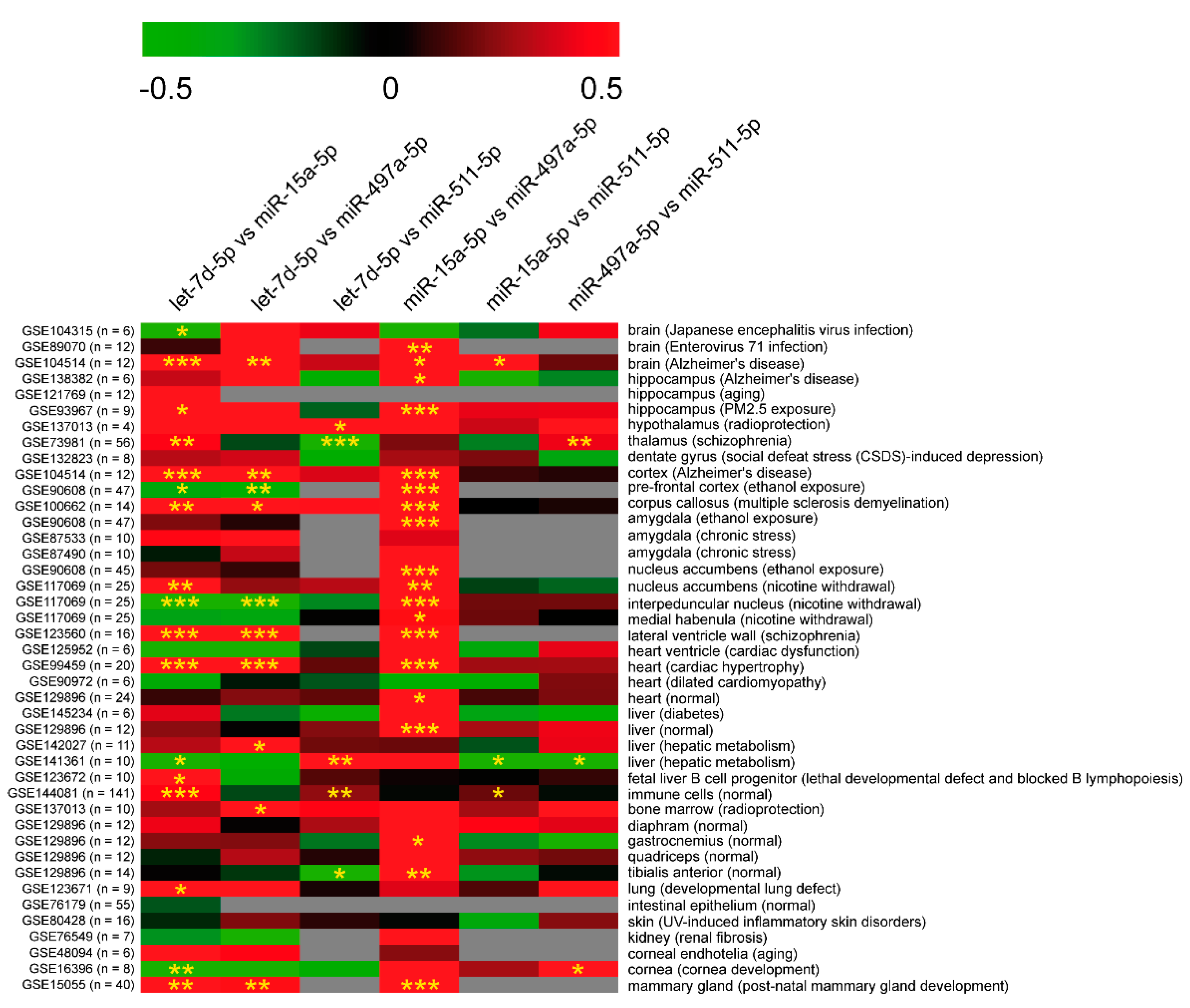
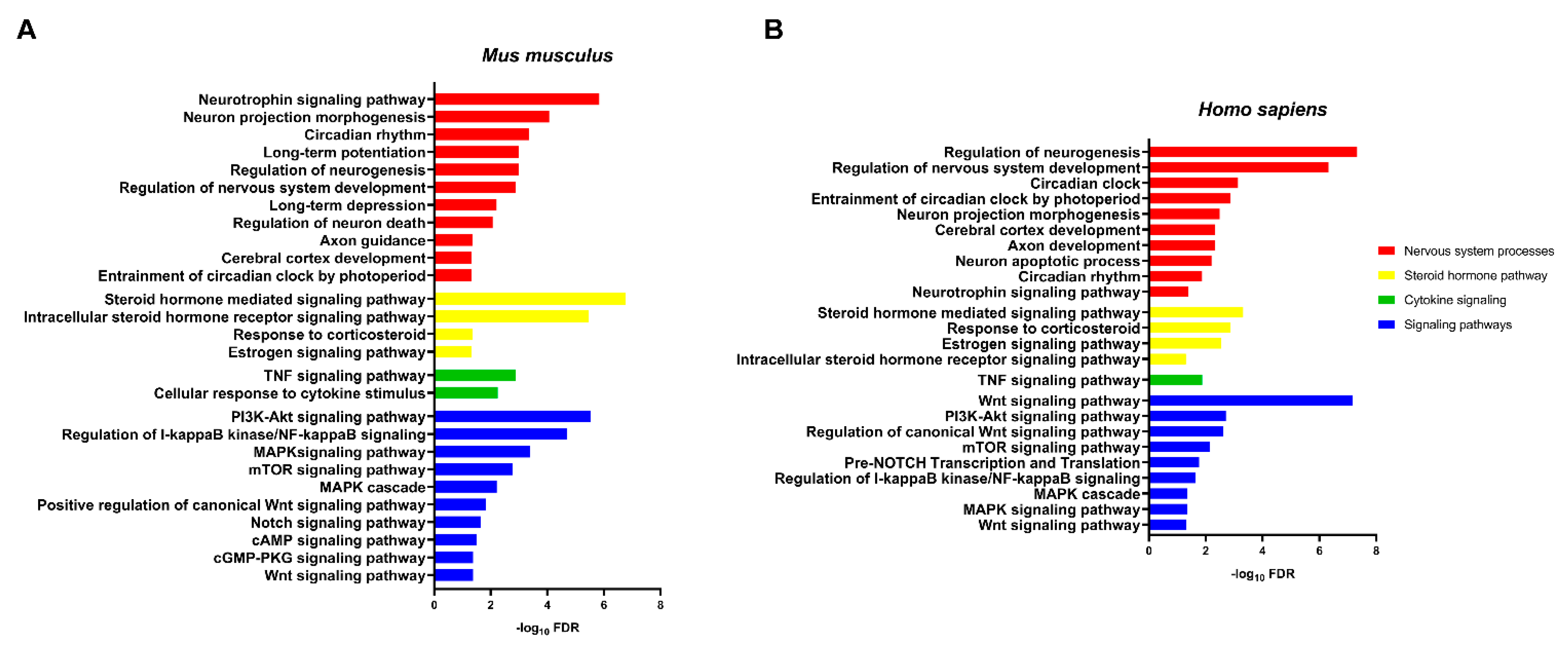
2.2.7. Dysregulation of miRNAs Affects PTSD-Related Pathways
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Arousal-Based Individual Screening (AIS), ASR Sessions, Z-Normalization Score, and Behavioral Paradigms
4.3. Selection of miRNAs
4.4. Tissue Collection
4.5. Total RNA Extraction
4.6. cDNA Synthesis and RT-qPCR
4.7. TaqMan microRNA Assays
4.8. Computational Identification of miRNA Regulators
4.9. Gene Ontology and Pathway Enrichment Analyses
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Juruena, M.F.; Eror, F.; Cleare, A.J.; Young, A.H. The Role of Early Life Stress in HPA Axis and Anxiety. Adv. Exp. Med. Biol. 2020, 1191, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Jeanneteau, F.D.; Lambert, W.M.; Ismaili, N.; Bath, K.G.; Lee, F.S.; Garabedian, M.J.; Chao, M.V. BDNF and glucocorticoids regulate cortico-trophin-releasing hormone (CRH) homeostasis in the hypothalamus. Proc. Natl. Acad. Sci. USA 2012, 109, 1305–1310. [Google Scholar] [CrossRef] [Green Version]
- Vukojevic, V.; Kolassa, I.-T.; Fastenrath, M.; Gschwind, L.; Spalek, K.; Milnik, A.; Heck, A.; Vogler, C.; Wilker, S.; Demougin, P.; et al. Epigenetic Modification of the Glucocorticoid Receptor Gene Is Linked to Traumatic Memory and Post-Traumatic Stress Disorder Risk in Genocide Survivors. J. Neurosci. 2014, 34, 10274–10284. [Google Scholar] [CrossRef]
- White, S.; Acierno, R.; Ruggiero, K.J.; Koenen, K.C.; Kilpatrick, D.G.; Galea, S.; Gelernter, J.; Williamson, V.; McMichael, O.; Vladimirov, V.I.; et al. Association of CRHR1 variants and posttraumatic stress symptoms in hurricane exposed adults. J. Anxiety Disord. 2013, 27, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Andero, R.; Ressler, K.J. Fear extinction and BDNF: Translating animal models of PTSD to the clinic. Genes Brain Behav. 2012, 11, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Règue, M.; Poilbout, C.; Martin, V.; Franc, B.; Lanfumey, L.; Mongeau, R. Increased 5-HT2C receptor editing predisposes to PTSD-like behaviors and alters BDNF and cytokines signaling. Transl. Psychiatry 2019, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Girgenti, M.J. Molecular and cellular studies of PTSD: Postmortem transcriptome analysis and novel therapeutic targets. J. Neurosci. Res. 2019, 97, 292–299. [Google Scholar] [CrossRef]
- Young, D.A.; Inslicht, S.S.; Metzler, T.J.; Neylan, T.C.; Ross, J.A. The effects of early trauma and the FKBP5 gene on PTSD and the HPA axis in a clinical sample of Gulf War veterans. Psychiatry Res. 2018, 270, 961–966. [Google Scholar] [CrossRef]
- Binder, E.B. The role of FKBP5, a co-chaperone of the glucocorticoid receptor in the pathogenesis and therapy of affective and anxiety disorders. Psychoneuroendocrinology 2009, 34, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, C.F.; Phifer, J.; Bradley, B.; Ressler, K.J. Risk and resilience: Genetic and environmental influences on development of the stress response. Depress. Anxiety 2009, 26, 984–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, J.; Wagner, K.V.; Liebl, C.; Scharf, S.H.; Wang, X.-D.; Wolf, M.; Hausch, F.; Rein, T.; Schmidt, U.; Touma, C.; et al. The involvement of FK506-binding protein 51 (FKBP5) in the behavioral and neuroendocrine effects of chronic social defeat stress. Neuropharmacology 2012, 62, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Scharf, S.H.; Liebl, C.; Binder, E.B.; Schmidt, M.V.; Müller, M.B. Expression and Regulation of the Fkbp5 Gene in the Adult Mouse Brain. PLoS ONE 2011, 6, e16883. [Google Scholar] [CrossRef] [Green Version]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 21, 403–416. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, J.C.; Dharia, S.; Blair, L.J.; Brady, S.; Johnson, A.G.; Peters, M.; Cheung-Flynn, J.; Cox, M.B.; de Erausquin, G.; Weeber, E.J.; et al. A new anti-depressive strategy for the elderly: Ablation of FKBP5/FKBP51. PLoS ONE 2011, 6, e24840. [Google Scholar] [CrossRef]
- Sabbagh, J.J.; O’Leary, J.C.; Blair, L.J.; Klengel, T.; Nordhues, B.A.; Fontaine, S.N.; Binder, E.B.; Dickey, C.A. Age-associated epigenetic upregulation of the FKBP5 gene selectively impairs stress resiliency. PLoS ONE 2014, 9, e107241. [Google Scholar] [CrossRef]
- Touma, C.; Gassen, N.C.; Herrmann, L.; Cheung-Flynn, J.; Bull, D.R.; Ionescu, I.A.; Heinzmann, J.-M.; Knapman, A.; Siebertz, A.; Depping, A.-M.; et al. FK506 binding protein 5 shapes stress re-sponsiveness: Modulation of neuroendocrine reactivity and coping behavior. Biol. Psychiatry 2011, 70, 928–936. [Google Scholar] [CrossRef]
- Criado-Marrero, M.; Silva, R.J.M.; Velazquez, B.; Hernández, A.; Colon, M.; Cruz, E.; Soler-Cedeño, O.; Porter, J.T. Dynamic expression of FKBP5 in the medial prefrontal cortex regulates resiliency to conditioned fear. Learn. Mem. 2017, 24, 145–152. [Google Scholar] [CrossRef]
- Young, K.A.; Thompson, P.M.; Cruz, D.A.; Williamson, D.E.; Selemon, L.D. BA11 FKBP5 expression levels correlate with dendritic spine density in postmortem PTSD and controls. Neurobiol. Stress 2015, 2, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Minichiello, L. TrkB signalling pathways in LTP and learning. Nat. Rev. Neurosci. 2009, 10, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef]
- Notaras, M.; Buuse, M.V.D. Neurobiology of BDNF in fear memory, sensitivity to stress, and stress-related disorders. Mol. Psychiatry 2020, 25, 2251–2274. [Google Scholar] [CrossRef]
- Niknazar, S.; Nahavandi, A.; Peyvandi, A.A.; Peyvandi, H.; Akhtari, A.S.; Karimi, M. Comparison of the Adulthood Chronic Stress Effect on Hippocampal BDNF Signaling in Male and Female Rats. Mol. Neurobiol. 2016, 53, 4026–4033. [Google Scholar] [CrossRef]
- Kozlovsky, N.; Matar, M.A.; Kaplan, Z.; Kotler, M.; Zohar, J.; Cohen, H. Long-term down-regulation of BDNF mRNA in rat hippo-campal CA1 subregion correlates with PTSD-like behavioural stress response. Int. J. Neuropsychopharmacol. 2007, 10, 741–758. [Google Scholar] [CrossRef] [Green Version]
- Cunha, C.; Angelucci, A.; D’Antoni, A.; Dobrossy, M.D.; Dunnett, S.B.; Berardi, N.; Brambilla, R. Brain-derived neurotrophic factor (BDNF) overexpression in the forebrain results in learning and memory impairments. Neurobiol. Dis. 2009, 33, 358–368. [Google Scholar] [CrossRef]
- Naert, G.; Ixart, G.; Maurice, T.; Tapia-Arancibia, L.; Givalois, L. Brain-derived neurotrophic factor and hypothalam-ic-pituitary-adrenal axis adaptation processes in a depressive-like state induced by chronic restraint stress. Mol. Cell Neurosci. 2011, 46, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, J.M.; Lumley, L.A.; Medina, W.; Robison, C.L.; Chang, W.E.; Alagappan, A.; Bah, M.J.; Dawood, M.Y.; Shah, J.D.; Mark, B.; et al. Acute social defeat reduces neurotrophin expression in brain cortical and subcortical areas in mice. Brain Res. 2004, 1025, 10–20. [Google Scholar] [CrossRef]
- Tsankova, N.M.; Kumar, A.; Nestler, E.J. Histone Modifications at Gene Promoter Regions in Rat Hippocampus after Acute and Chronic Electroconvulsive Seizures. J. Neurosci. 2004, 24, 5603–5610. [Google Scholar] [CrossRef] [Green Version]
- Lubin, F.D.; Roth, T.L.; Sweatt, J.D. Epigenetic Regulation of bdnf Gene Transcription in the Consolidation of Fear Memory. J. Neurosci. 2008, 28, 10576–10586. [Google Scholar] [CrossRef] [PubMed]
- Fuchikami, M.; Yamamoto, S.; Morinobu, S.; Takei, S.; Yamawaki, S. Epigenetic regulation of BDNF gene in response to stress. Psychiatry Investig. 2010, 7, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Bredy, T.W.; Wu, H.; Crego, C.; Zellhoefer, J.; Sun, Y.E.; Barad, M. Histone modifications around individual BDNF gene promoters in prefrontal cortex are associated with extinction of conditioned fear. Learn. Mem. 2007, 14, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Chiaruttini, C.; Vicario, A.; Li, Z.; Baj, G.; Braiuca, P.; Wu, Y.; Lee, F.S.; Gardossi, L.; Baraban, J.M.; Tongiorgi, E. Dendritic trafficking of BDNF mRNA is mediated by translin and blocked by the G196A (Val66Met) mutation. Proc. Natl. Acad. Sci. USA 2009, 106, 16481–16486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, J.R.; Mutesa, L.; Uddin, M. Traumatic Stress Epigenetics. Curr. Behav. Neurosci. Rep. 2018, 5, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Rosenblat, J.D.; Brietzke, E.; Pan, Z.; Lee, Y.; Cao, B.; Zuckerman, H.; Kalantarova, A.; McIntyre, R.S. Stress, epigenetics and depression: A systematic review. Neurosci. Biobehav. Rev. 2019, 102, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Ramo-Fernández, L.; Boeck, C.; Koenig, A.M.; Schury, K.; Binder, E.B.; Gündel, H.; Fegert, J.M.; Karabatsiakis, A.; Kolassa, I.-T. The effects of childhood maltreatment on epigenetic regulation of stress-response associated genes: An intergenerational approach. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeshurun, S.; Hannan, A.J. Transgenerational epigenetic influences of paternal environmental exposures on brain function and predisposition to psychiatric disorders. Mol. Psychiatry 2019, 24, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Rajman, M.; Schratt, G. MicroRNAs in neural development: From master regulators to fine-tuners. Development 2017, 144, 2310–2322. [Google Scholar] [CrossRef] [Green Version]
- Follert, P.; Cremer, H.; Béclin, C. MicroRNAs in brain development and function: A matter of flexibility and stability. Front. Mol. Neurosci. 2014, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Olejniczak, M.; Kotowska-Zimmer, A.; Krzyzosiak, W. Stress-induced changes in miRNA biogenesis and functioning. Cell. Mol. Life Sci. 2017, 75, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Tan, E.K.; Zeng, L. microRNAs and Neurodegenerative Diseases. Adv. Exp. Med. Biol. 2015, 888, 85–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.S.; Smith, A.K.; Nievergelt, C.M.; Uddin, M. Neuroepigenetics of Post-Traumatic Stress Disorder. Prog. Molecular Biol. Transl. Sci. 2018, 158, 227–253. [Google Scholar] [CrossRef]
- Zannas, A.S.; Provençal, N.; Binder, E.B. Epigenetics of Posttraumatic Stress Disorder: Current Evidence, Challenges, and Future Directions. Biol. Psychiatry 2015, 78, 327–335. [Google Scholar] [CrossRef]
- Dickson, D.A.; Paulus, J.K.; Mensah, V.; Lem, J.; Saavedra-Rodriguez, L.; Gentry, A.; Pagidas, K.; Feig, L.A. Reduced levels of miRNAs 449 and 34 in sperm of mice and men exposed to early life stress. Transl. Psychiatry 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Toth, M. Mechanisms of Non-Genetic Inheritance and Psychiatric Disorders. Neuropsychopharmacology 2014, 40, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Dias, B.G.; Goodman, J.V.; Ahluwalia, R.; Easton, A.E.; Andero, R.; Ressler, K.J. Amygdala-Dependent Fear Memory Consolidation via miR-34a and Notch Signaling. Neuron 2014, 83, 906–918. [Google Scholar] [CrossRef] [Green Version]
- Griggs, E.M.; Young, E.J.; Rumbaugh, G.; Miller, C.A. MicroRNA-182 Regulates Amygdala-Dependent Memory Formation. J. Neurosci. 2013, 33, 1734–1740. [Google Scholar] [CrossRef] [Green Version]
- Vetere, G.; Barbato, C.; Pezzola, S.; Frisone, P.; Aceti, M.; Ciotti, M.; Cogoni, C.; Ammassari-Teule, M.; Ruberti, F. Selective inhibition of miR-92 in hippocampal neurons alters contextual fear memory. Hippocampus 2014, 24, 1458–1465. [Google Scholar] [CrossRef]
- Jovasevic, V.; Corcoran, A.K.; Leaderbrand, K.; Yamawaki, N.; Guedea, A.L.; Chen, H.J.; Shepherd, G.M.G.; Radulovic, J. GABAergic mechanisms regulated by miR-33 encode state-dependent fear. Nat. Neurosci. 2015, 18, 1265–1271. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.Y.; Phang, R.Z.; Hsu, P.H.; Wang, W.H.; Huang, H.T.; Liu, I.Y. In vivo knockdown of hippocampal miR-132 expression impairs memory acquisition of trace fear conditioning. Hippocampus 2013, 23, 625–633. [Google Scholar] [CrossRef]
- Chen, R.J.; Kelly, G.; Sengupta, A.; Heydendael, W.; Nicholas, B.; Beltrami, S.; Luz, S.; Peixoto, L.; Abel, T.; Bhatnagar, S. MicroRNAs as biomarkers of resilience or vul-nerability to stress. Neuroscience 2015, 305, 36–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillivan, S.E.; Jamieson, S.; de Nijs, L.; Jones, M.; Snijders, C.; Klengel, T.; Joseph, N.F.; Krauskoph, J.; Kleinjans, J.; Vinkers, C.; et al. MicroRNA regulation of persistent stress-enhanced memory. Mol. Psychiatry 2020, 25, 965–976. [Google Scholar] [CrossRef]
- Torrisi, S.A.; Lavanco, G.; Maurel, O.M.; Gulisano, W.; Laudani, S.; Geraci, F.; Grasso, M.; Barbagallo, C.; Caraci, F.; Bucolo, C.; et al. A novel arousal-based individual screening reveals susceptibility and resilience to PTSD-like phenotypes in mice. Neurobiol. Stress 2021, 14, 100286. [Google Scholar] [CrossRef] [PubMed]
- Hager, T.; Jansen, R.F.; Pieneman, A.W.; Manivannan, S.N.; Golani, I.; Van Der Sluis, S.; Smit, A.B.; Verhage, M.; Stiedl, O. Display of individuality in avoidance behavior and risk assessment of inbred mice. Front. Behav. Neurosci. 2014, 8, 314. [Google Scholar] [CrossRef] [Green Version]
- Dirven, B.C.J.; Homberg, J.R.; Kozicz, T.; Henckens, M. Epigenetic programming of the neuroendocrine stress response by adult life stress. J. Mol. Endocrinol. 2017, 59, 11–31. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, R.M.; Gururajan, A.; Dinan, T.G.; Kenny, P.J.; Cryan, J.F. All Roads Lead to the miRNome: miRNAs Have a Central Role in the Molecular Pathophysiology of Psychiatric Disorders. Trends Pharmacol. Sci. 2016, 37, 1029–1044. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, W.; Xu, J.; Guo, Y.; Yan, J.; Meng, L.; Jiang, C.; Lu, S. Interpreting the MicroRNA-15/107 family: Interaction identification by combining network based and experiment supported approach. BMC Med. Genet. 2019, 20, 96. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.J.; Murrough, J.W.; Han, M.-H.; Charney, D.S.; Nestler, E.J. Neurobiology of resilience. Nat. Neurosci. 2012, 15, 1475–1484. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Pan, J.; Liu, W.; Li, Y.; Li, F.; Liu, M. MicroRNA-15a-5p serves as a potential biomarker and regulates the viability and apoptosis of hippocampus neuron in children with temporal lobe epilepsy. Diagn. Pathol. 2020, 15, 46–47. [Google Scholar] [CrossRef]
- Volk, N.; Pape, J.C.; Engel, M.; Zannas, A.S.; Cattane, N.; Cattaneo, A.; Binder, E.B.; Chen, A. Amygdalar MicroRNA-15a Is Essential for Coping with Chronic Stress. Cell Rep. 2016, 17, 1882–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, D.; Sabbagh, J.J.; Blair, L.J.; Darling, A.L.; Wen, X.; Dickey, C.A. MicroRNA-511 Binds to FKBP5 mRNA, Which Encodes a Chaperone Protein, and Regulates Neuronal Differentiation. J. Biol. Chem. 2016, 291, 17897–17906. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, R.; Liu, Y.; Wang, W.; Liu, D.; Jiang, H.; Pan, F. Short- and long-term alterations of FKBP5-GR and specific microRNAs in the prefrontal cortex and hippocampus of male rats induced by adolescent stress contribute to depression susceptibility. Psychoneuroendocrinology 2019, 101, 204–215. [Google Scholar] [CrossRef]
- Yoshino, Y.; Roy, B.; Dwivedi, Y. Differential and unique patterns of synaptic miRNA expression in dorsolateral prefrontal cortex of depressed subjects. Neuropsychopharmacology 2021, 46, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Smalheiser, N.R.; Lugli, G.; Rizavi, H.S.; Torvik, V.I.; Turecki, G.; Dwivedi, Y. MicroRNA Expression Is Down-Regulated and Reorganized in Prefrontal Cortex of Depressed Suicide Subjects. PLoS ONE 2012, 7, e33201. [Google Scholar] [CrossRef] [PubMed]
- Banigan, M.G.; Kao, P.F.; Kozubek, J.A.; Winslow, A.R.; Medina, J.; Costa, J.; Schmitt, A.; Schneider, A.; Cabral, H.; Cagsal-Getkin, O.; et al. Differential Expression of Exosomal microRNAs in Prefrontal Cortices of Schizophrenia and Bipolar Disorder Patients. PLoS ONE 2013, 8, e48814. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Pandey, A.; Shukla, A.; Talwelkar, S.S.; Kumar, A.; Pant, A.B.; Parmar, D. miR-497 and miR-302b Regulate Ethanol-induced Neuronal Cell Death through BCL2 Protein and Cyclin D2*. J. Biol. Chem. 2011, 286, 37347–37357. [Google Scholar] [CrossRef] [Green Version]
- Maffioletti, E.; Cattaneo, A.; Rosso, G.; Maina, G.; Maj, C.; Gennarelli, M.; Tardito, D.; Bocchio-Chiavetto, L. Peripheral whole blood microRNA alterations in major depression and bipolar disorder. J. Affect. Disord. 2016, 200, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Bahi, A.; Dreyer, J.-L. Lentiviral-mediated let-7d microRNA overexpression induced anxiolytic- and anti-depressant-like behaviors and impaired dopamine D3 receptor expression. Eur. Neuropsychopharmacol. 2018, 28, 1394–1404. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.Y.; Qing, D.; Wang, B.; Zeng, Q.-Y.; Chen, Y.-C.; Jin, Y.; Zeng, C.-C.; Bao, R. Comparison of microRNA expression in hippocampus and the marginal division (MrD) of the neostriatum in rats. J. Biomed. Sci. 2013, 20, 9. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekar, V.; Dreyer, J.-L. Regulation of MiR-124, Let-7d, and MiR-181a in the Accumbens Affects the Expression, Extinction, and Reinstatement of Cocaine-Induced Conditioned Place Preference. Neuropsychopharmacology 2011, 36, 1149–1164. [Google Scholar] [CrossRef]
- Kfoury, N.; Kapatos, G. Identification of neuronal target genes for CCAAT/Enhancer Binding Proteins. Mol. Cell. Neurosci. 2009, 40, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Calella, A.M.; Nerlov, C.; Lopez, R.G.; Sciarretta, C.; Halbach, O.V.B.; Bereshchenko, O.; Minichiello, L. Neurotrophin/Trk receptor signaling mediates C/EBPalpha, -beta and NeuroD recruitment to immediate-early gene promoters in neuronal cells and re-quires C/EBPs to induce immediate-early gene transcription. Neural Dev. 2007, 2, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.M.; Gibbons, H.M.; Oldfield, R.L.; Bergin, P.M.; Mee, E.W.; Faull, R.L.M.; Dragunow, M. The transcription factor PU.1 is critical for viability and function of human brain microglia. Glia 2013, 61, 929–942. [Google Scholar] [CrossRef]
- Rustenhoven, J.; Smith, A.M.; Smyth, L.C.; Jansson, D.; Scotter, E.L.; Swanson, M.E.V.; Aalderink, M.; Coppieters, N.; Narayan, P.; Handley, R.; et al. PU.1 regulates Alzheimer’s disease-associated genes in primary human microglia. Mol. Neurodegener. 2018, 13, 1–16. [Google Scholar] [CrossRef]
- Zhang, J.; Chang, L.; Pu, Y.; Hashimoto, K. Abnormal expression of colony stimulating factor 1 receptor (CSF1R) and transcription factor PU.1 (SPI1) in the spleen from patients with major psychiatric disorders: A role of brain–spleen axis. J. Affect. Disord. 2020, 272, 110–115. [Google Scholar] [CrossRef]
- Yu, S.; Yu, M.; He, X.; Wen, L.; Bu, Z.; Feng, J. KCNQ1OT1 promotes autophagy by regulating miR-200a/FOXO3/ATG7 pathway in cerebral ischemic stroke. Aging Cell 2019, 18, e12940. [Google Scholar] [CrossRef] [PubMed]
- Li, M.M.; Liu, X.-H.; Zhao, Y.-C.; Ma, X.-Y.; Zhou, Y.-C.; Zhao, Y.-X.; Liu, X.-Y. Long noncoding RNA KCNQ1OT1 promotes apoptosis in neu-roblastoma cells by regulating miR-296-5p/Bax axis. FEBS J. 2020, 287, 561–577. [Google Scholar] [CrossRef]
- Wang, H.J.; Tang, X.L.; Huang, G.; Li, Y.B.; Pan, R.H.; Zhan, J.; Wu, Y.-K.; Liang, J.-F.; Bai, X.-X.; Cai, J. Long Non-Coding KCNQ1OT1 Promotes Oxygen-Glucose-Deprivation/Reoxygenation-Induced Neurons Injury Through Regulating MIR-153-3p/FOXO3 Axis. J. Stroke Cerebrovasc. Dis. 2020, 29, 105126. [Google Scholar] [CrossRef]
- Green, C.R.; Corsi-Travali, S.; Neumeister, A. The Role of BDNF-TrkB Signaling in the Pathogenesis of PTSD. J. Depress. Anxiety 2013, 29, 105126. [Google Scholar]
- Mahan, A.L.; Ressler, K.J. Fear conditioning, synaptic plasticity and the amygdala: Implications for posttraumatic stress disorder. Trends Neurosci. 2012, 35, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Rasmusson, A.M.; Pineles, S.L. Neurotransmitter, Peptide, and Steroid Hormone Abnormalities in PTSD: Biological Endopheno-types Relevant to Treatment. Curr Psychiatry Rep. 2018, 20, 52. [Google Scholar] [CrossRef]
- Aliev, G.; Beeraka, N.M.; Nikolenko, V.N.; Svistunov, A.A.; Rozhnova, T.; Kostyuk, S.; Cherkesov, I.; Gavryushova, L.V.; Chekhonatsky, A.A.; Mikhaleva, L.M.; et al. Neurophysiology and Psychopathology Underlying PTSD and Recent Insights into the PTSD Therapies—A Comprehensive Review. J. Clin. Med. 2020, 9, 2951. [Google Scholar] [CrossRef] [PubMed]
- Fifield, K.; Hebert, M.; Angel, R.; Adamec, R.; Blundell, J. Inhibition of mTOR kinase via rapamycin blocks persistent predator stress-induced hyperarousal. Behav. Brain Res. 2013, 256, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Knox, D.; Della Valle, R.; Mohammadmirzaei, N.; Shultz, B.; Biddle, M.; Farkash, A.; Chamness, M.; Moulton, E. PI3K-Akt Signaling in the Basolateral Amygdala Facilitates Traumatic Stress Enhancements in Fear Memory. Int. J. Neuropsychopharmacol. 2021, 24, 229–238. [Google Scholar] [CrossRef]
- Hauger, R.L.; Olivares-Reyes, J.A.; Dautzenberg, F.M.; Lohr, J.B.; Braun, S.; Oakley, R.H. Molecular and cell signaling targets for PTSD pathophysiology and pharmacotherapy. Neuropharmacology 2012, 62, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Yehuda, R.; Golier, J.A.; Kaufman, S. Circadian Rhythm of Salivary Cortisol in Holocaust Survivors with and without PTSD. Am. J. Psychiatry 2005, 162, 998–1000. [Google Scholar] [CrossRef]
- Gao, J.; Wang, H.; Liu, Y.; Li, Y.Y.; Chen, C.; Liu, L.M.; Wu, Y.-M.; Li, S.; Yang, C. Glutamate and GABA imbalance promotes neuronal apoptosis in hip-pocampus after stress. Med. Sci. Monit. 2014, 20, 499–512. [Google Scholar]
- Glover, E.M.; Jovanovic, T.; Norrholm, S.D. Estrogen and Extinction of Fear Memories:Implications for Posttraumatic Stress Disorder Treatment. Biol. Psychiatry 2015, 78, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Liufu, Z.; Zhao, Y.; Guo, L.; Miao, G.; Xiao, J.; Lyu, Y.; Chen, Y.; Shi, S.; Tang, T.; Wu, C.-I. Redundant and incoherent regulations of multiple phenotypes suggest microRNAs’ role in stability control. Genome Res. 2017, 27, 1665–1673. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.; Zhou, Y.; Hu, Z.; Lou, J.; Song, W.; Li, J.; Liang, X.; Chen, C.; Wang, S.; Yang, B.; et al. 24-hour-restraint stress induces long-term depressive-like phenotypes in mice. Sci. Rep. 2016, 6, 32935. [Google Scholar] [CrossRef] [Green Version]
- Mozhui, K.; Karlsson, R.-M.; Kash, T.L.; Ihne, J.; Norcross, M.; Patel, S.; Farrell, M.R.; Hill, E.E.; Graybeal, C.; Martin, K.P.; et al. Strain differences in stress re-sponsivity are associated with divergent amygdala gene expression and glutamate-mediated neuronal excitability. J. Neurosci. 2010, 14, 5357–5367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbitsky, A.; Dopfel, D.; Zhang, N. Rodent models of post-traumatic stress disorder: Behavioral assessment. Transl. Psychiatry 2020, 10, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Ellenbroek, B.; Youn, J. Rodent models in neuroscience research: Is it a rat race? Dis. Model. Mech. 2016, 9, 1079–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alekseeva, I.V.; Abramova, A.Y.; Kozlov, A.Y.; Koplik, E.V.; Pertsov, A.S.; Lyadov, D.A.; Nikenina, E.V. State of Stress-Marker Organs in Rats after a Single Exposure to Long-Term Stress and Treatment with Lipopolysaccharide. Bull. Exp. Biol. Med. 2019, 167, 624–627. [Google Scholar] [CrossRef] [PubMed]
- Abramova, A.Y.; Pertsov, S.S.; Alekseeva, I.V.; Nikenina, E.V.; Kozlov, A.Y.; Chekhlov, V.V.; Chukhnina, M.E. Blood Cytokine Concentration in Rats during Antigenic Treatment after a Single Long-Term Stress Exposure. Bull. Exp. Biol. Med. 2020, 168, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Spijker, S. Dissection of Rodent Brain Region. In Neuroproteomics; Humana Press: Totowa, NJ, USA, 2011; pp. 13–26. [Google Scholar]
- Leggio, G.M.; Camillieri, G.; Platania, C.B.M.; Castorina, A.; Marrazzo, G.; Torrisi, S.A.; Nona, C.N.; D’Agata, V.; Nobrega, J.; Stark, H.; et al. Dopamine D3 Receptor Is Necessary for Ethanol Consumption: An Approach with Buspirone. Neuropsychopharmacology 2014, 39, 2017–2028. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Area | Target | miR-15a-5p | miR-511-5p | miR-497a-5p | let-7d-5p |
|---|---|---|---|---|---|
| HIP | BDNF | −0.09 (0.65) | −0.04 (0.82) | −0.04 (0.80) | −0.18 (0.36) |
| FKBP5 | −0.40 (0.03) | −0.45 (0.02) | −0.43 (0.02) | −0.07 (0.69) | |
| HT | BDNF | 0.66 (0.005) | 0.73 (0.01) | 0.53 (0.03) | 0.62 (0.01) |
| FKBP5 | −0.69 (0.002) | −0.60 (0.01) | −0.62 (0.022) | −0.38 (0.20) | |
| mPFC | BDNF | −0.10 (0.60) | −0.38 (0.10) | −0.01 (0.94) | −0.32 (0.09) |
| FKBP5 | 0.03 (0.85) | −0.20 (0.29) | 0.06 (0.76) | −0.16 (0.39) |
| Brain Area | Target | miRNA Combination | R | R2 | p-Value |
|---|---|---|---|---|---|
| HIP | FKBP5 | let-7d-5p, miR-15a-5p, miR-497a-5p, miR-511-5p | 0.657 | 0.431 | 0.024 |
| miR-15a-5p, miR-497a-5p, miR-511-5p | 0.652 | 0.425 | 0.01 | ||
| miR-497a-5p, miR-511-5p | 0.641 | 0.411 | 0.004 | ||
| BDNF | let-7d-5p, miR-15a-5p, miR-497a-5p, miR-511-5p | 0.191 | 0.037 | 0.936 | |
| let-7d-5p, miR-15a-5p, miR-511-5p | 0.191 | 0.037 | 0.841 | ||
| let-7d-5p, miR-15a-5p | 0.186 | 0.035 | 0.656 | ||
| HT | FKBP5 | let-7d-5p, miR-15a-5p, miR-497a-5p, miR-511-5p | 0.749 | 0.56 | 0.044 |
| miR-15a-5p, miR-497a-5p, miR-511-5p | 0.741 | 0.549 | 0.019 | ||
| miR-15a-5p, miR-511-5p | 0.719 | 0.517 | 0.009 | ||
| mPFC | FKBP5 | let-7d-5p, miR-15a-5p, miR-497a-5p, miR-511-5p | 0.429 | 0.184 | 0.322 |
| let-7d-5p, miR-15a-5p, miR-511-5p | 0.42 | 0.177 | 0.206 | ||
| let-7d-5p, miR-15a-5p | 0.323 | 0.105 | 0.251 | ||
| BDNF | let-7d-5p, miR-15a-5p, miR-497a-5p, miR-511-5p | 0.597 | 0.357 | 0.038 | |
| let-7d-5p, miR-15a-5p, miR-511-5p | 0.587 | 0.345 | 0.019 | ||
| let-7d-5p, miR-511-5p | 0.487 | 0.237 | 0.039 |
| miRNAs | HIP | HT | mPFC | ||||||
|---|---|---|---|---|---|---|---|---|---|
| miR-15a-5p | let-7d-5p | miR-511-5p | miR-15a-5p | let-7d-5p | miR-511-5p | miR-15a-5p | let-7d-5p | miR-511-5p | |
| let-7d-5p | 0.41 (0.03) | 0.55 (0.02) | 0.46 (0.01) | ||||||
| miR-511-5p | 0.38 (0.04) | 0.39 (0.04) | 0.70 (0.002) | 0.63 (0.008) | 0.32 (0.09) | 0.18 (0.36) | |||
| miR-497a-5p | 0.25 (0.19) | 0.35 (0.06) | 0.64 (0.0003) | 0.39 (0.12) | 0.38 (0.13) | 0.62 (0.01) | 0.57 (0.001) | 0.54 (0.003) | 0.14 (0.46) |
| miRNAs | miR-15a | let-7d | miR-497a |
|---|---|---|---|
| let-7d | Cebpa, Spi1 | ||
| miR-497a | Bcl11b, Brd4, Cbfb, Cebpb, Ctcf, Ep300, Erg, Ets1, Hdac2, Ikzf1, Max, Med1, Myc, Nelfe, Nr1d1, Nr3c1, Otx2, Pou5f1, Pparg, Rad21, Rela, Smarca4, Spi1, Suz12, Tfap4, Zfp281, Zfp384 | Spi1 | |
| miR-511 | Batf, Btaf1, Cebpa, Cebpb, Mafb, Nr3c1, Rela, Spi1, Tcf12 | Cebpa, Spi1 | Cebpb, Fli1, Nr3c1, Rela, Spi1, Stat5a |
| miR-15a-5p | let-7d-5p | miR-497a-5p | |
|---|---|---|---|
| let-7d-5p | Gm20605, Kcnq1ot1, Ptprv, Gm4117 | ||
| miR-497a-5p | Gm20605, Ptprv, ENSMUSG00000093535, Gm11579, Rab10os, Kcnq1ot1, Gm4117 | Gm20605, Kcnq1ot1, Ptprv, Gm4117, Mecomos | |
| miR-511-5p | Kcnq1ot1, ENSMUSG00000093535, Gm4117, Gm11579, ENSMUSG00000085287, Rab10os | Kcnq1ot1, Gm26856, Gm4117, Zfp950 | Kcnq1ot1, ENSMUSG00000093535, Gm4117, Gm11579, Rab10os |
| miRNAs/mRNAs | HIP | |||
|---|---|---|---|---|
| Arousal | Avoidance | Social Memory | PTSD-Like | |
| miR-15a-5p | −0.44 (0.02) | −0.33 (0.08) | −0.12 (0.56) | −0.31 (0.11) |
| let-7d-5p | −0.08 (0.68) | 0.26 (0.20) | 0.30 (0.13) | 0.16 (0.42) |
| miR-511-5p | −0.44 (0.02) | −0.14 (0.49) | −0.10 (0.61) | −0.34 (0.08) |
| miR-497a-5p | −0.22 (0.28) | −0.12 (0.56) | −0.10 (0.61) | −0.24 (0.22) |
| FKBP5 | 0.39 (0.04) | −0.04 (0.84) | −0.09 (0.65) | 0.11 (0.61) |
| BDNF | 0.47 (0.01) | −0.02 (0.89) | −0.10 (0.62) | 0.27 (0.18) |
| HT | ||||
| Arousal | Avoidance | Social Memory | PTSD-like | |
| miR-15a-5p | −0.26 (0.31) | −0.27 (0.30) | −0.11 (0.66) | −0.26 (0.31) |
| let-7d-5p | −0.12 (0.64) | −0.20 (0.45) | 0.09 (0.72) | −0.07 (0.79) |
| miR-511-5p | −0.13 (0.62) | −0.63 (0.007) | −0.31 (0.23) | −0.39 (0.12) |
| miR-497a-5p | −0.61 (0.01) | −0.57 (0.01) | −0.37 (0.15) | −0.67 (0.004) |
| FKBP5 | −0.02 (0.92) | 0.22 (0.39) | 0.27 (0.29) | −0.17 (0.51) |
| BDNF | −0.25 (0.33) | −0.54 (0.02) | −0.37 (0.15) | −0.46 (0.07) |
| mPFC | ||||
| Arousal | Avoidance | Social Memory | PTSD-like | |
| miR-15a-5p | 0.09 (0.65) | −0.10 (0.60) | 0.10 (0.61) | 0.04 (0.83) |
| let-7d-5p | 0.11 (0.58) | −0.13 (0.53) | 0.18 (0.38) | 0.13 (0.52) |
| miR-511-5p | −0.09 (0.67) | −0.37 (0.07) | −0.09 (0.64) | −0.19 (0.34) |
| miR-497a-5p | −0.02 (0.92) | −0.27 (0.19) | −0.15 (0.45) | −0.15 (0.45) |
| FKBP5 | 0.27 (0.18) | 0.16 (0.41) | 0.18 (0.37) | 0.30 (0.12) |
| BDNF | −0.27 (0.19) | 0.16 (0.43) | 0.28 (0.16) | 0.02 (0.89) |
| Scores | miRNAs/mRNAs | HIPPOCAMPUS | HYPOTHALAMUS | mPFC | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| R | R2 | p-value | R | R2 | p-Value | R | R2 | p-Value | ||
| Arousal score | miRNA + mRNAs | 0.72 | 0.519 | 0.032 | 0.563 | 0.317 | 0.791 | 0.385 | 0.149 | 0.88 |
| MiRNA | 0.442 | 0.196 | 0.335 | 0.981 | 0.962 | 0.00022 | 0.199 | 0.04 | 0.937 | |
| mRNAs | 0.523 | 0.273 | 0.03 | 0.274 | 0.075 | 0.761 | 0.103 | 0.011 | 0.885 | |
| Avoidance-like score | miRNA + mRNAs | 0.529 | 0.28 | 0.402 | 0.728 | 0.53 | 0.449 | 0.503 | 0.253 | 0.632 |
| miRNA | 0.349 | 0.122 | 0.606 | 0.711 | 0.506 | 0.304 | 0.488 | 0.238 | 0.247 | |
| mRNAs | 0.133 | 0.018 | 0.822 | 0.688 | 0.473 | 0.106 | 0.038 | 0.001 | 0.983 | |
| Social memory score | miRNA + mRNAs | 0.691 | 0.477 | 0.058 | 0.884 | 0.782 | 0.094 | 0.541 | 0.293 | 0.526 |
| miRNA | 0.512 | 0.263 | 0.172 | 0.884 | 0.781 | 0.035 | 0.207 | 0.043 | 0.928 | |
| mRNAs | 0.241 | 0.058 | 0.518 | 0.725 | 0.525 | 0.074 | 0.254 | 0.065 | 0.464 | |
| PTSD-like score | miRNA + mRNAs | 0.699 | 0.488 | 0.05 | 0.69 | 0.476 | 0.541 | 0.531 | 0.282 | 0.554 |
| miRNA | 0.502 | 0.252 | 0.194 | 0.915 | 0.838 | 0.015 | 0.33 | 0.109 | 0.681 | |
| MRNAs | 0.347 | 0.121 | 0.244 | 0.541 | 0.293 | 0.298 | 0.162 | 0.026 | 0.737 | |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| FKBP5 | 5′-TGAGGGCACCAGTAACAATGG-3′ | 5′-CAACATCCCTTTGTAGTGGACAT-3′ |
| BDNF | 5′-GTTCGAGAGGTCTGACGACG-3′ | 5′-AGTCCGCGTCCTTATGGTTT-3′ |
| GAPDH | 5′-AGGTCGGTGTGAACGGATTTG-3′ | 5′-TGTAGACCATGTAGTTGAGGTCA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurel, O.M.; Torrisi, S.A.; Barbagallo, C.; Purrello, M.; Salomone, S.; Drago, F.; Ragusa, M.; Leggio, G.M. Dysregulation of miR-15a-5p, miR-497a-5p and miR-511-5p Is Associated with Modulation of BDNF and FKBP5 in Brain Areas of PTSD-Related Susceptible and Resilient Mice. Int. J. Mol. Sci. 2021, 22, 5157. https://doi.org/10.3390/ijms22105157
Maurel OM, Torrisi SA, Barbagallo C, Purrello M, Salomone S, Drago F, Ragusa M, Leggio GM. Dysregulation of miR-15a-5p, miR-497a-5p and miR-511-5p Is Associated with Modulation of BDNF and FKBP5 in Brain Areas of PTSD-Related Susceptible and Resilient Mice. International Journal of Molecular Sciences. 2021; 22(10):5157. https://doi.org/10.3390/ijms22105157
Chicago/Turabian StyleMaurel, Oriana Maria, Sebastiano Alfio Torrisi, Cristina Barbagallo, Michele Purrello, Salvatore Salomone, Filippo Drago, Marco Ragusa, and Gian Marco Leggio. 2021. "Dysregulation of miR-15a-5p, miR-497a-5p and miR-511-5p Is Associated with Modulation of BDNF and FKBP5 in Brain Areas of PTSD-Related Susceptible and Resilient Mice" International Journal of Molecular Sciences 22, no. 10: 5157. https://doi.org/10.3390/ijms22105157