
Lumican Inhibits Osteoclastogenesis and Bone Resorption by Suppressing Akt Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
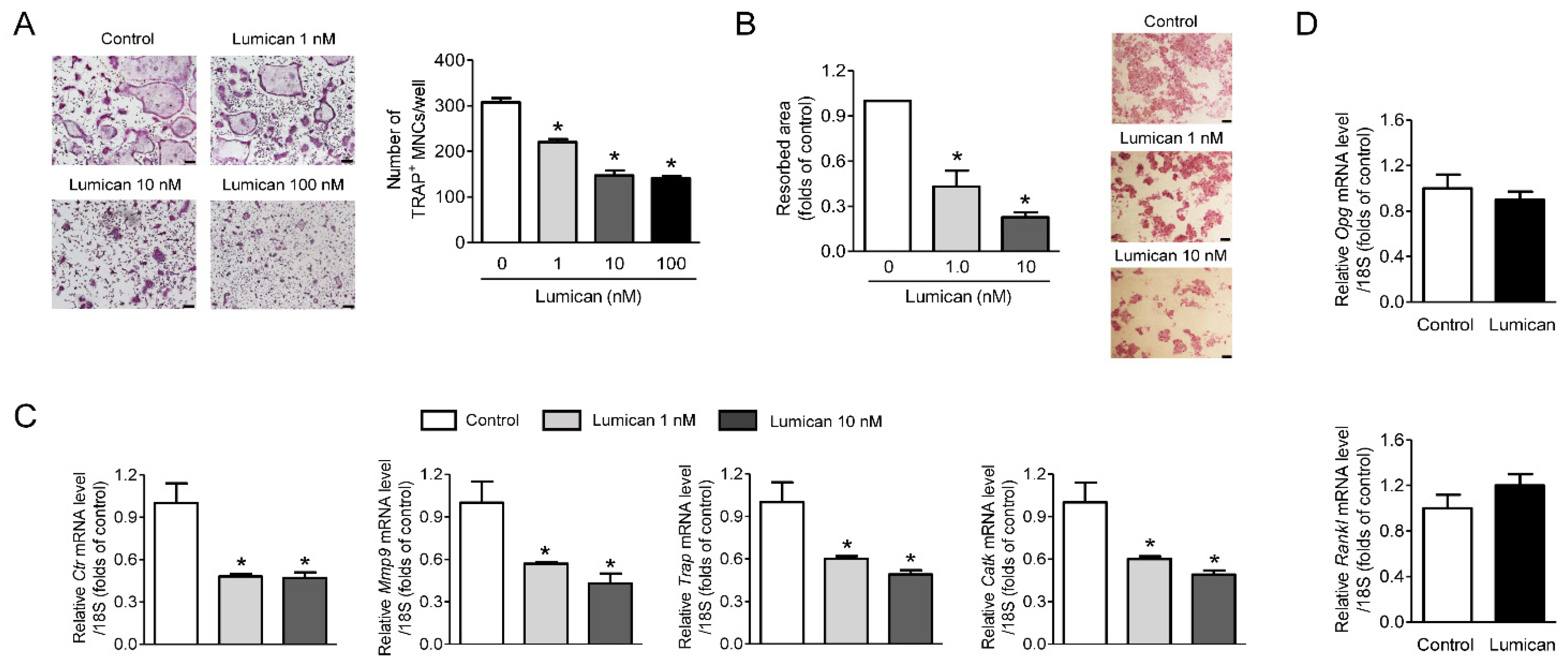
2.1. Inhibition of In Vitro Bone Resorption and Osteoclastogenesis by Lumican Treatment
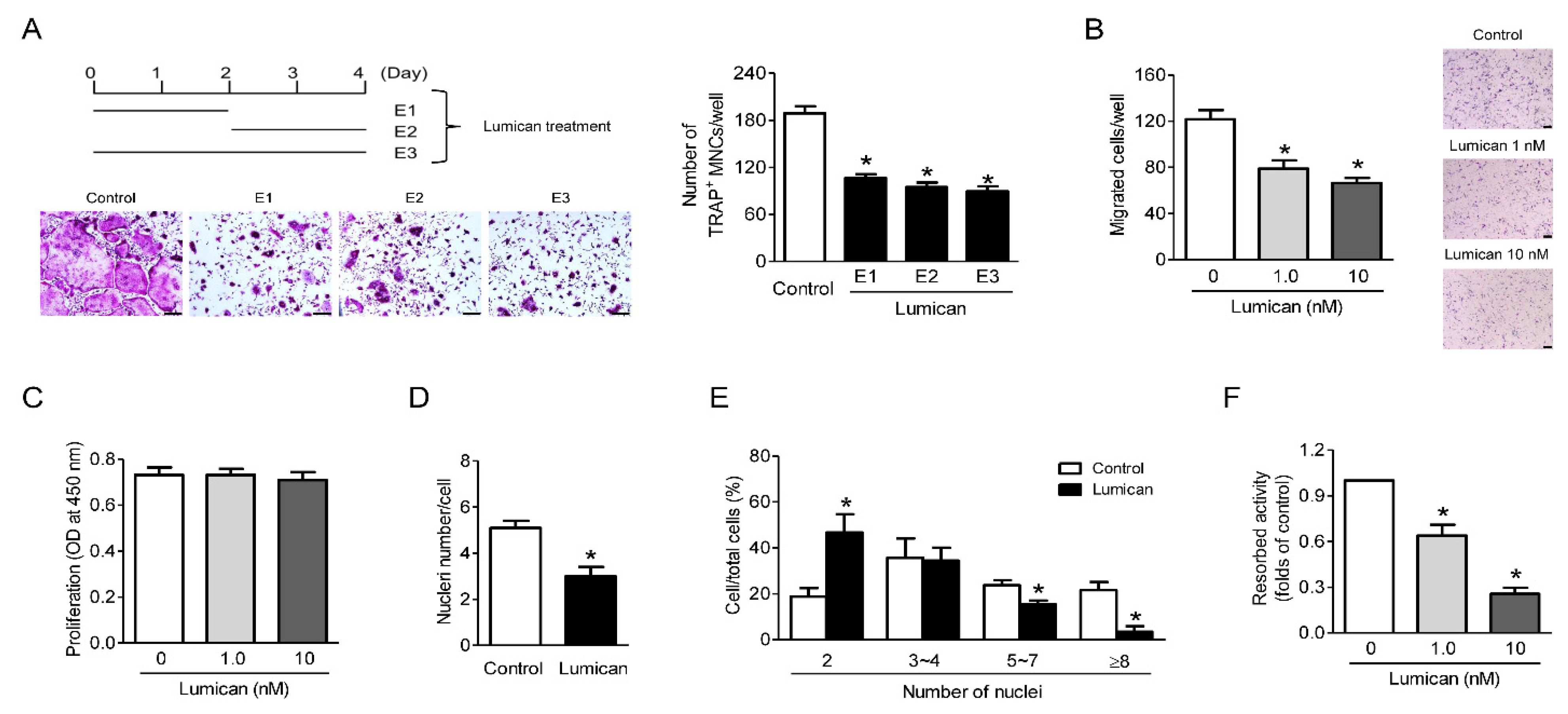
2.2. Lumican Decreases Bone Resorptive Activity as Well as Migration and Fusion of Preosteoclasts
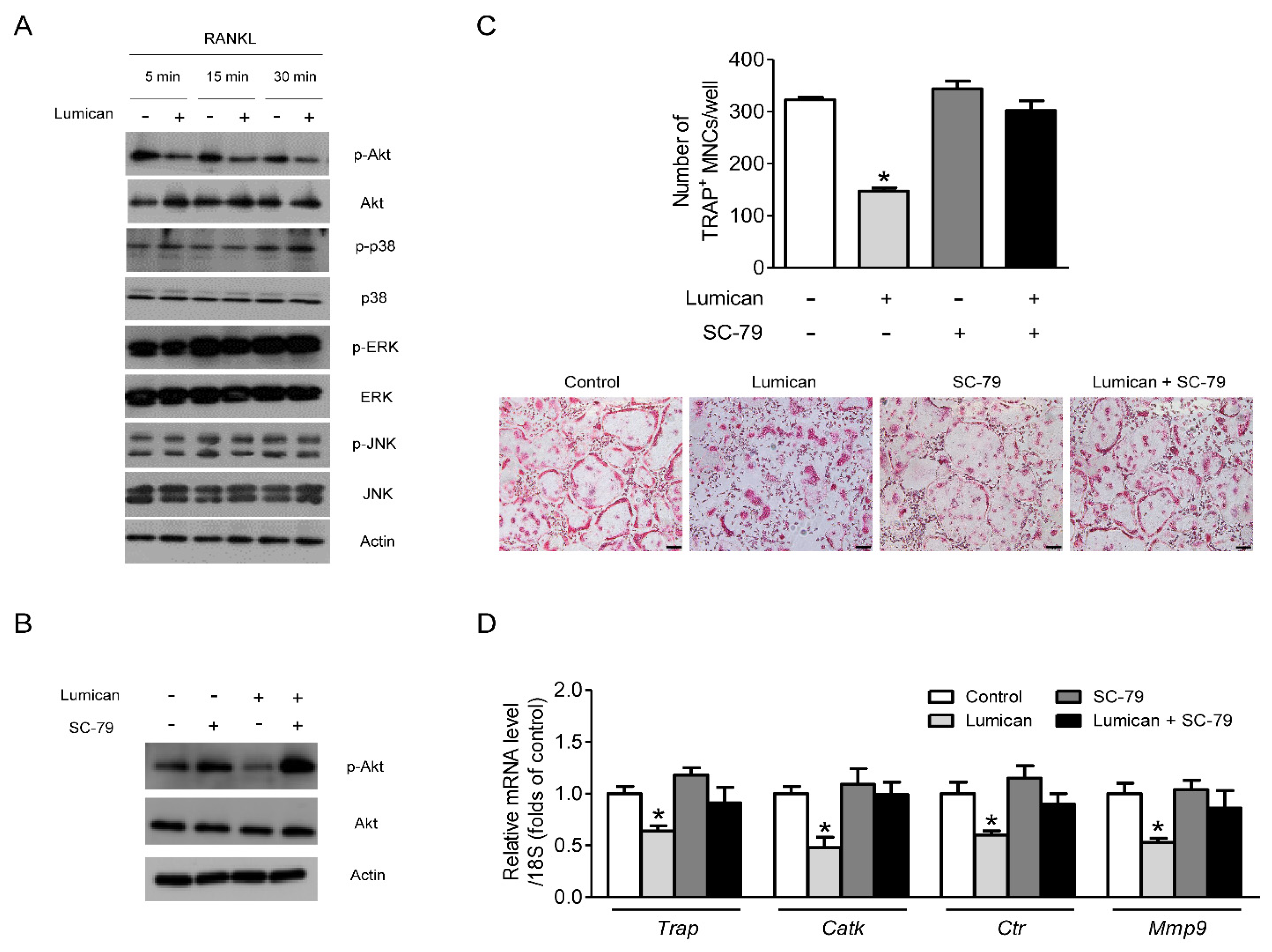
2.3. Akt Signaling Mediates the Effects of Lumican on Osteoclastogenesis
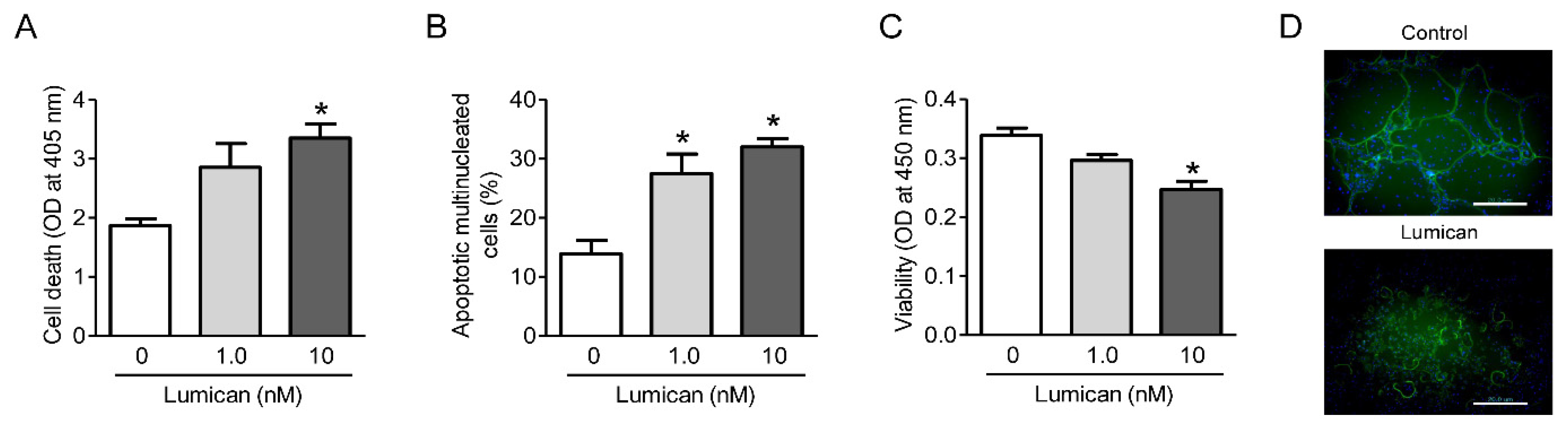
2.4. Lumican Increases Apoptosis of Osteoclasts and Decreases Actin Ring Formation
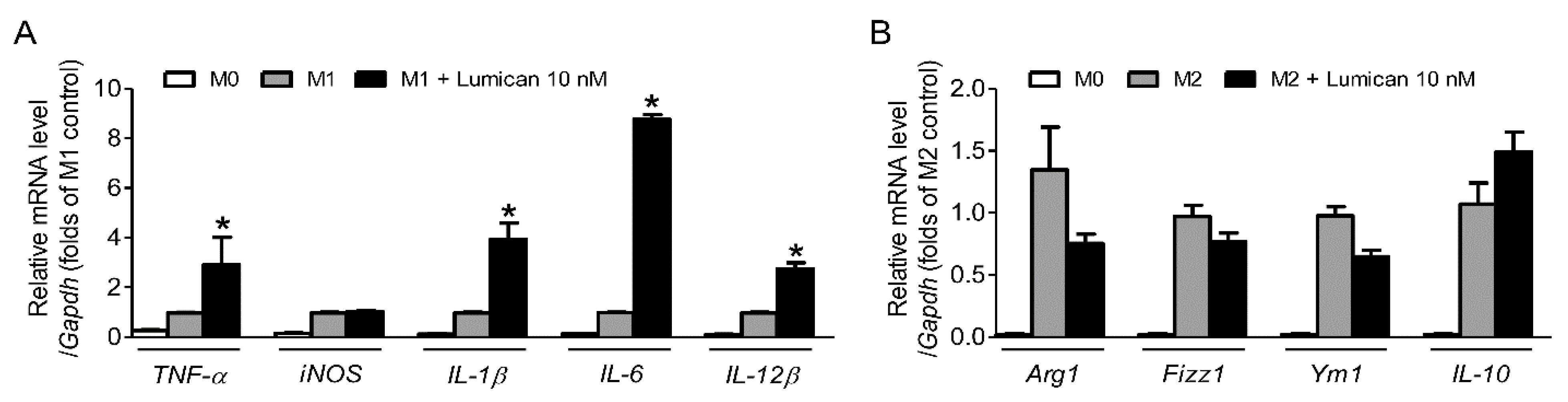
2.5. Lumican Stimulates the Polarization of M0 Macrophages to M1 Macrophages
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. TRAP Stain
4.3. In Vitro Resorption Assay
4.4. Quantitative Reverse-Transcription PCR (qRT-PCR)
4.5. Migration Assay
4.6. Proliferation Assay
4.7. Western Blot Analysis
4.8. Apoptosis Assays
4.9. Viability Assay
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schaefer, L.; Iozzo, R.V. Biological functions of the small leucine-rich proteoglycans: From genetics to signal transduction. J. Biol. Chem. 2008, 283, 21305–21309. [Google Scholar] [CrossRef] [Green Version]
- Iozzo, R.V. Matrix proteoglycans: From molecular design to cellular function. Annu. Rev. Biochem. 1998, 67, 609–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwan, P.A.; Scott, P.G.; Bishop, P.N.; Bella, J. Structural correlations in the family of small leucine-rich repeat proteins and proteoglycans. J. Struct. Biol. 2006, 155, 294–305. [Google Scholar] [CrossRef]
- Merline, R.; Schaefer, R.M.; Schaefer, L. The matricellular functions of small leucine-rich proteoglycans (SLRPs). J. Cell Commun. Signal. 2009, 3, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Zappia, J.; Joiret, M.; Sanchez, C.; Lambert, C.; Geris, L.; Muller, M.; Henrotin, Y. From Translation to Protein Degradation as Mechanisms for Regulating Biological Functions: A Review on the SLRP Family in Skeletal Tissues. Biomolecules 2020, 10, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikitovic, D.; Aggelidakis, J.; Young, M.F.; Iozzo, R.V.; Karamanos, N.K.; Tzanakakis, G.N. The biology of small leucine-rich proteoglycans in bone pathophysiology. J. Biol. Chem. 2012, 287, 33926–33933. [Google Scholar] [CrossRef] [Green Version]
- Young, M.F.; Bi, Y.; Ameye, L.; Xu, T.; Wadhwa, S.; Heegaard, A.; Kilts, T.; Chen, X.D. Small leucine-rich proteoglycans in the aging skeleton. J. Musculoskelet. Neuronal Interact. 2006, 6, 364–365. [Google Scholar]
- Xu, T.; Bianco, P.; Fisher, L.W.; Longenecker, G.; Smith, E.; Goldstein, S.; Bonadio, J.; Boskey, A.; Heegaard, A.M.; Sommer, B.; et al. Targeted disruption of the biglycan gene leads to an osteoporosis-like phenotype in mice. Nat. Genet. 1998, 20, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Nielsen, K.L.; Kilts, T.M.; Yoon, A.M.A.K.; Wimer, H.F.; Greenfield, E.M.; Heegaard, A.M.; Young, M.F. Biglycan deficiency increases osteoclast differentiation and activity due to defective osteoblasts. Bone 2006, 38, 778–786. [Google Scholar] [CrossRef]
- Nuka, S.; Zhou, W.; Henry, S.P.; Gendron, C.M.; Schultz, J.B.; Shinomura, T.; Johnson, J.; Wang, Y.; Keene, D.R.; Ramirez-Solis, R.; et al. Phenotypic characterization of epiphycan-deficient and epiphycan/biglycan double-deficient mice. Osteoarthr. Cartil. 2010, 18, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Nikitovic, D.; Katonis, P.; Tsatsakis, A.; Karamanos, N.K.; Tzanakakis, G.N. Lumican, a small leucine-rich proteoglycan. IUBMB Life 2008, 60, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Nikitovic, D.; Berdiaki, A.; Zafiropoulos, A.; Katonis, P.; Tsatsakis, A.; Karamanos, N.K.; Tzanakakis, G.N. Lumican expression is positively correlated with the differentiation and negatively with the growth of human osteosarcoma cells. FEBS J. 2008, 275, 350–361. [Google Scholar] [CrossRef]
- D’Onofrio, M.F.; Brezillon, S.; Baranek, T.; Perreau, C.; Roughley, P.J.; Maquart, F.X.; Wegrowski, Y. Identification of beta1 integrin as mediator of melanoma cell adhesion to lumican. Biochem. Biophys. Res. Commun. 2008, 365, 266–272. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, S.J.; Kim, D.A.; Lee, S.H.; Koh, J.M.; Kim, B.J. Muscle-Derived Lumican Stimulates Bone Formation via Integrin α2β1 and the Downstream ERK Signal. Front. Cell Dev. Biol. 2020, 8, 565826. [Google Scholar] [CrossRef]
- Boyce, B.F. Advances in osteoclast biology reveal potential new drug targets and new roles for osteoclasts. J. Bone Miner. Res. 2013, 28, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Yim, M. The Role of Toll-Like Receptors in Osteoclastogenesis. J. Bone Metab. 2020, 27, 227–235. [Google Scholar] [CrossRef]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nagase, Y.; Hirose, J.; Tokuyama, N.; Yasui, T.; Kadono, Y.; Ueki, K.; Kadowaki, T.; Nakamura, K.; Tanaka, S. Regulation of bone resorption and sealing zone formation in osteoclasts occurs through protein kinase B-mediated microtubule stabilization. J. Bone Miner. Res. 2013, 28, 1191–1202. [Google Scholar] [CrossRef]
- Sugatani, T.; Hruska, K.A. Akt1/Akt2 and mammalian target of rapamycin/Bim play critical roles in osteoclast differentiation and survival, respectively, whereas Akt is dispensable for cell survival in isolated osteoclast precursors. J. Biol. Chem. 2005, 280, 3583–3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Wan, Y. Molecular determinants for the polarization of macrophage and osteoclast. Semin. Immunopathol. 2019, 41, 551–563. [Google Scholar] [CrossRef]
- Kim, B.J.; Koh, J.M. Coupling factors involved in preserving bone balance. Cell. Mol. Life Sci. 2019, 76, 1243–1253. [Google Scholar] [CrossRef]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef] [Green Version]
- Reyes, C.; Hitz, M.; Prieto-Alhambra, D.; Abrahamsen, B. Risks and Benefits of Bisphosphonate Therapies. J. Cell. Biochem. 2016, 117, 20–28. [Google Scholar] [CrossRef]
- Min, Y.K. Update on denosumab treatment in postmenopausal women with osteoporosis. Endocrinol. Metab. 2015, 30, 19–26. [Google Scholar] [CrossRef]
- Seomun, Y.; Joo, C.-K. Lumican induces human corneal epithelial cell migration and integrin expression via ERK 1/2 signaling. Biochem. Biophys. Res. Commun. 2008, 372, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Brézillon, S.; Pietraszek, K.; Maquart, F.-X.; Wegrowski, Y. Lumican effects in the control of tumour progression and their links with metalloproteinases and integrins. FEBS J. 2013, 280, 2369–2381. [Google Scholar] [CrossRef]
- Duong, L.T.; Lakkakorpi, P.; Nakamura, I.; Rodan, G.A. Integrins and signaling in osteoclast function. Matrix Biol. J. Int. Soc. Matrix Biol. 2000, 19, 97–105. [Google Scholar] [CrossRef]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A.; et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, N. Signaling Pathways in Osteoclast Differentiation. Chonnam. Med.J. 2016, 52, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Moon, J.B.; Kim, J.H.; Kim, K.; Youn, B.U.; Ko, A.; Lee, S.Y.; Kim, N. Akt induces osteoclast differentiation through regulating the GSK3β/NFATc1 signaling cascade. J. Immunol. 2012, 188, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Golden, L.H.; Insogna, K.L. The expanding role of PI3-kinase in bone. Bone 2004, 34, 3–12. [Google Scholar] [CrossRef]
- Lee, K.; Seo, I.; Choi, M.H.; Jeong, D. Roles of Mitogen-Activated Protein Kinases in Osteoclast Biology. Int. J. Mol. Sci. 2018, 19, 4. [Google Scholar] [CrossRef] [Green Version]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef]
- Park, S.J.; Lee, J.Y.; Lee, S.H.; Koh, J.M.; Kim, B.J. SLIT2 inhibits osteoclastogenesis and bone resorption by suppression of Cdc42 activity. Biochem. Biophys. Res. Commun. 2019, 514, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Lee, Y.S.; Lee, S.Y.; Baek, W.Y.; Choi, Y.J.; Moon, S.A.; Lee, S.H.; Kim, J.E.; Chang, E.J.; Kim, E.Y.; et al. Osteoclast-secreted SLIT3 coordinates bone resorption and formation. J. Clin. Investig. 2018, 128, 1429–1441. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.J.; Lee, J.Y.; Park, S.J.; Lee, S.H.; Kim, S.J.; Yoo, H.J.; Rivera De Pena, S.I.; McGee-Lawrence, M.; Isales, C.M.; Koh, J.M.; et al. Elevated ceramides 18:0 and 24:1 with aging are associated with hip fracture risk through increased bone resorption. Aging 2019, 11, 9388–9404. [Google Scholar] [CrossRef]
- Kim, D.A.; Park, S.J.; Lee, J.Y.; Kim, J.H.; Lee, S.; Lee, E.; Jang, I.Y.; Jung, H.W.; Park, J.H.; Kim, B.J. Effect of CCL11 on In Vitro Myogenesis and Its Clinical Relevance for Sarcopenia in Older Adults. Endocrinol. Metab. 2021. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-Y.; Kim, D.-A.; Kim, E.-Y.; Chang, E.-J.; Park, S.-J.; Kim, B.-J. Lumican Inhibits Osteoclastogenesis and Bone Resorption by Suppressing Akt Activity. Int. J. Mol. Sci. 2021, 22, 4717. https://doi.org/10.3390/ijms22094717
Lee J-Y, Kim D-A, Kim E-Y, Chang E-J, Park S-J, Kim B-J. Lumican Inhibits Osteoclastogenesis and Bone Resorption by Suppressing Akt Activity. International Journal of Molecular Sciences. 2021; 22(9):4717. https://doi.org/10.3390/ijms22094717
Chicago/Turabian StyleLee, Jin-Young, Da-Ae Kim, Eun-Young Kim, Eun-Ju Chang, So-Jeong Park, and Beom-Jun Kim. 2021. "Lumican Inhibits Osteoclastogenesis and Bone Resorption by Suppressing Akt Activity" International Journal of Molecular Sciences 22, no. 9: 4717. https://doi.org/10.3390/ijms22094717