Detecting the Mechanism behind the Transition from Fixed Two-Dimensional Patterned Sika Deer (Cervus nippon) Dermal Papilla Cells to Three-Dimensional Pattern

Abstract
:1. Introduction
2. Results
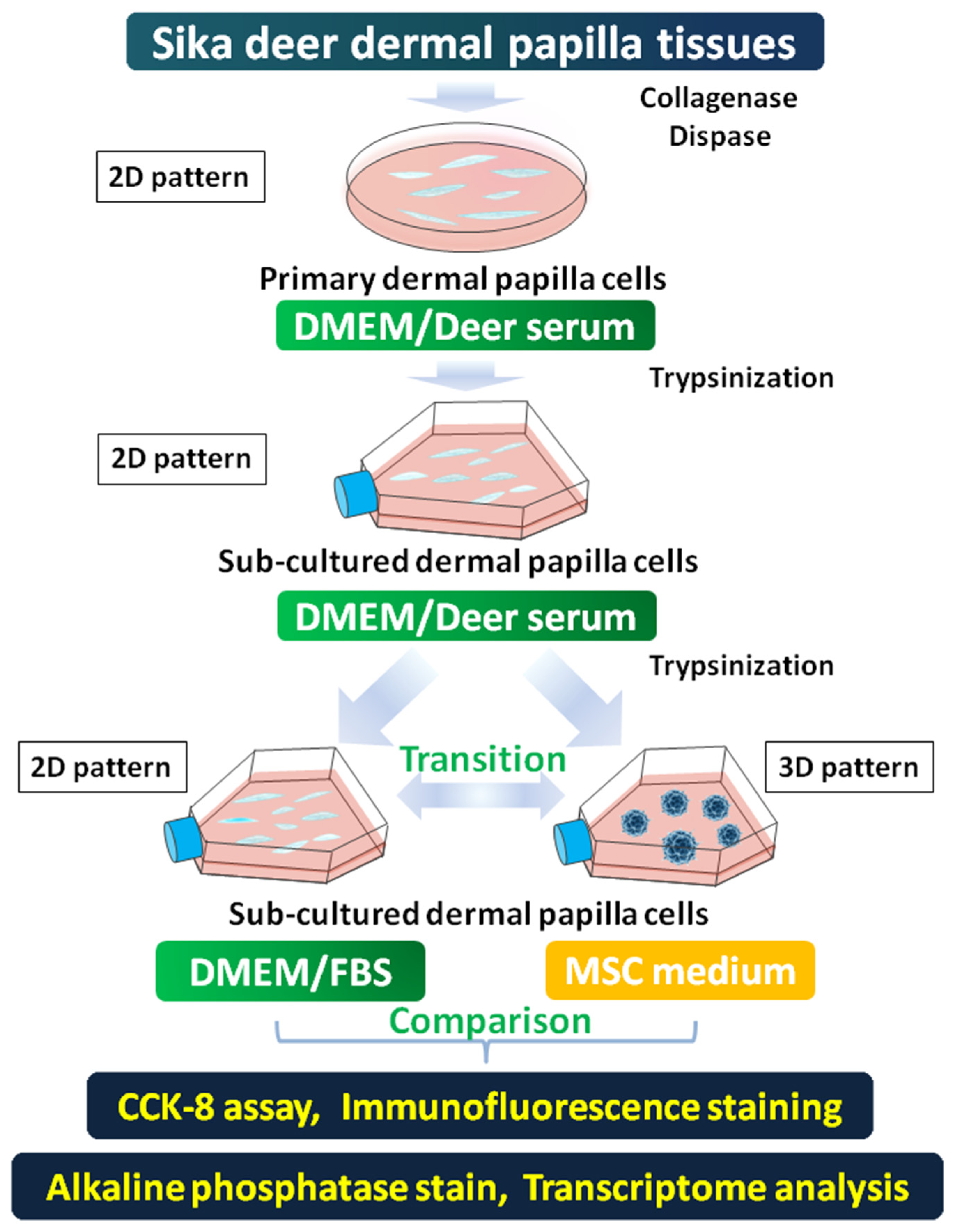
2.1. Sika Deer Dermal Papilla Cell Culture
2.2. Restoration of 3D-Pattern DP Cells from Fixed 2D Patterned DP Cells
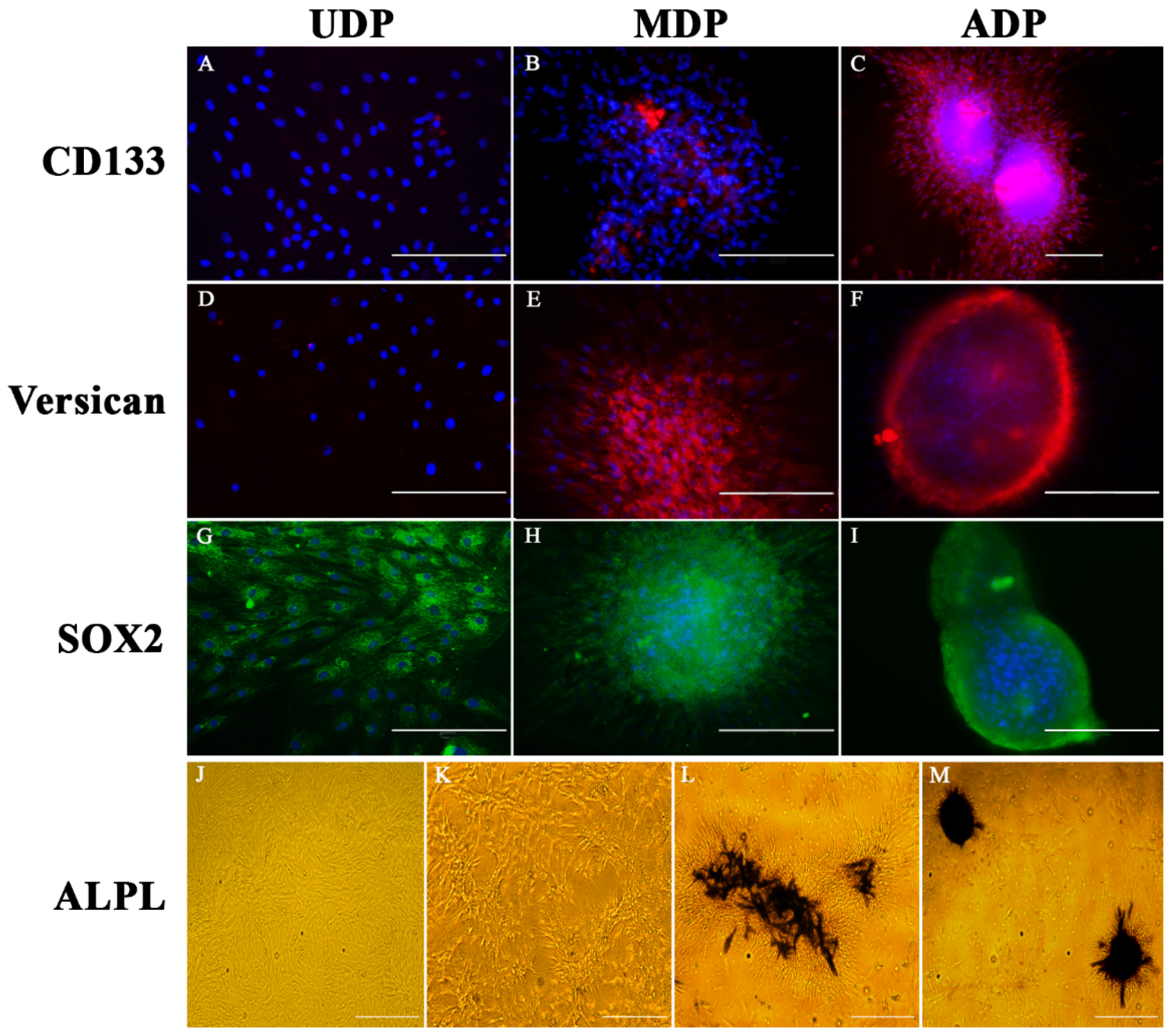
2.3. Identification of Hair Inducing Activity through Specific Expression of CD133, SOX2, Versican, and ALPL on DP Cells
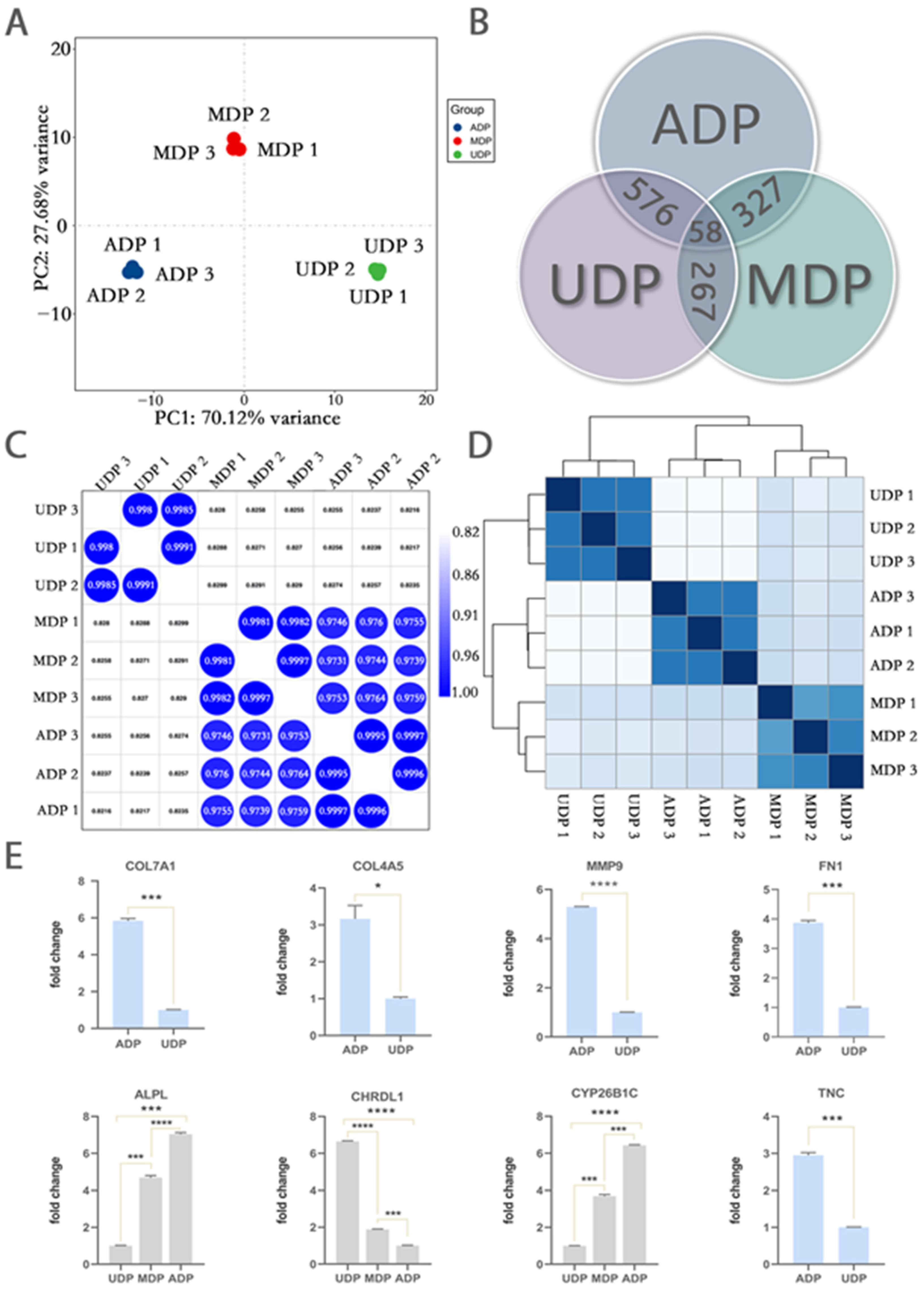
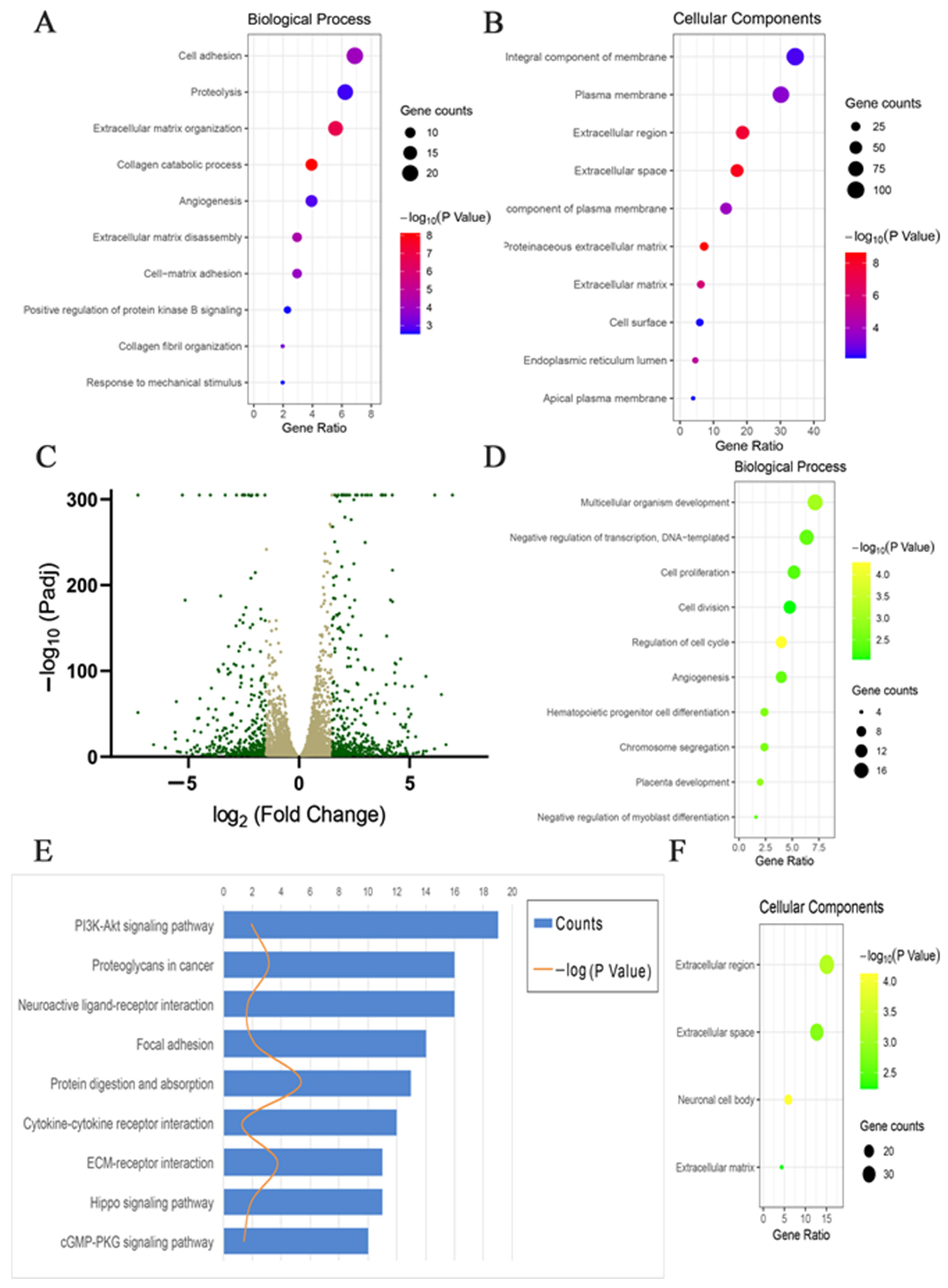
2.4. Transcriptome Analysis Revealed Important Genes Contributing to Restored 3D Pattern
2.5. Real-Time PCR Analysis Verified Transcriptome Results
3. Discussion
4. Materials & Methods
4.1. Animals
4.2. Dermal Papilla Isolation
4.3. Establishment of DP Cell Lines
4.4. Cell Counting Kit-8 Assay
4.5. Immunofluorescence Staining
4.6. Alkaline Phosphatase Stain
4.7. RNA Isolation and Real-Time PCR
4.8. Transcriptome Analysis
4.9. Bioinformatics Analysis on Gene Expression Pattern and Differentially Expressed Genes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandes, K.J.L.; McKenzie, I.A.; Mill, P.; Smith, K.M.; Akhavan, M.; Barnabé-Heider, F.; Biernaskie, J.; Junek, A.; Kobayashi, N.R.; Toma, J.G.; et al. A dermal niche for multipotent adult skin-derived precursor cells. Nat. Cell Biol. 2004, 6, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Ebling, F.J. Differentiation and Growth of Cells of the Skin. Differ. Growth Cells Vertebr. Tissues 1974, 14, 129–168. [Google Scholar] [CrossRef]
- Jahoda, C.A.B.; Oliver, R.F.; Reynolds, A.J.; Forrester, J.C.; Gillespie, J.W.; Cserhalmi-Friedman, P.B.; Christiano, A.M.; Horne, K.A. Transpecies hair growth induction by human hair follicle dermal papillae. J. Exp. Dermatol. 2011, 10, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R.F. The induction of hair follicle formation in the adult hooded rat by vibrissa dermal papillae. J. Embryol. Exp. Morphol. 1970, 23, 219–236. [Google Scholar] [PubMed]
- Pisansarakit, P.; Moore, G.P. Induction of hair follicles in mouse skin by rat vibrissa dermal papillae. J. Embryol. Exp. Morphol. 1986, 94, 113–119. [Google Scholar] [PubMed]
- Legue, E. Hair follicle renewal: Organization of stem cells in the matrix and the role of stereotyped lineages and behaviors. Development 2005, 132, 4143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, S.E. Molecular Mechanisms Regulating Hair Follicle Development. J. Investig. Dermatol. 2002, 118, 216–225. [Google Scholar] [CrossRef]
- Ibrahim, L.; Wright, E.A. A quantitative study of hair growth using mouse and rat vibrissal follicles. I. Dermal papilla volume determines hair volume. J. Embryol. Exp. Morphol. 1982, 72, 209–224. [Google Scholar]
- Higgins, C.A.; Chen, J.C.; Cerise, J.E.; Jahoda, C.A.B.; Christiano, A.M. Microenvironmental Reprogramming by Three-Dimensional Culture Enables Dermal Papilla Cells to Induce de Novo Human Hair-Follicle Growth. Proc. Natl. Acad. Sci. USA 2013, 110, 19679–19688. [Google Scholar] [CrossRef] [Green Version]
- Rufaut, N.W.; Nixon, A.J.; Goldthorpe, N.T.; Wallace, O.A.M.; Pearson, A.J.; Sinclair, R.D. An In Vitro Model for the Mor-phogenesis of Hair Follicle Dermal Papillae. J. Investig. Dermatol. 2013, 133, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osada, A.; Iwabuchi, T.; Kishimoto, J.; Hamazaki, T.S.; Okochi, H. Long-Term Culture of Mouse Vibrissal Dermal Papilla Cells and De Novo Hair Follicle Induction. Tissue Eng. 2007, 13, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Jahoda CA, B.; Oliver, R.F. Vibrissa dermal papilla cell aggregative behaviour in vivo and in vitro. J. Embryol. Exp. Morphol. 1984, 79, 211–224. [Google Scholar] [PubMed]
- Ohyama, M.; Zheng, Y.; Paus, R.; Stenn, K.S. The mesenchymal component of hair follicle neogenesis: Background, methods and molecular characterization. Exp. Dermatol. 2010, 19, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-C.; Cotsarelis, G. Review of hair follicle dermal cells. J. Dermatol. Sci. 2010, 57, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Horne, K.A.; Jahoda, C.A.; Oliver, R.F. Whisker growth induced by implantation of cultured vibrissa dermal papilla cells in the adult rat. J. Embryol. Exp. Morphol. 1986, 97, 111–124. [Google Scholar]
- Kishimoto, J.; Ehama, R.; Wu, L.; Jiang, S.; Jiang, N.; Burgeson, R.E. Selective Activation of the Versican Promoter by Epithelial- Mesenchymal Interactions during Hair Follicle Development. Proc. Natl. Acad. Sci. USA 1999, 96, 7336–7341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, W.C.; Goodman, L.V.; George, C.; Morgan, D.L.; Ledbetter, S.; Yuspa, S.H.; Lichti, U. Reconstitution of Hair Follicle Development In Vivo: Determination of Follicle Formation, Hair Growth, and Hair Quality by Dermal Cells. J. Investig. Dermatol. 1993, 100, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendl, M.; Polak, L.; Fuchs, E. BMP signaling in dermal papilla cells is required for their hair follicle-inductive properties. Genes Dev. 2008, 22, 543–557. [Google Scholar] [CrossRef] [Green Version]
- Kang, B.M.; Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Sphere Formation Increases the Ability of Cultured Human Dermal Papilla Cells to Induce Hair Follicles from Mouse Epidermal Cells in a Reconstitution Assay. J. Investig. Dermatol. 2012, 132, 237–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohyama, M.; Kobayashi, T.; Sasaki, T.; Shimizu, A.; Amagai, M. Restoration of the intrinsic properties of human dermal papilla in vitro. J. Cell Sci. 2012, 125, 4114–4125. [Google Scholar] [CrossRef] [Green Version]
- Bratka-Robia, C.B.; Mitteregger, G.; Aichinger, A.; Egerbacher, M.; Helmreich, M.; Bamberg, E. Primary cell culture and morphological characterization of canine dermal papilla cells and dermal fibroblasts. Vet. Dermatol. 2002, 13, 1–6. [Google Scholar] [CrossRef]
- Fujisawa, A.; Kobayashi, T.; Amagai, M.; Iwasaki, T.; Ohyama, M. Molecular biological and morphological charaterization for canine dermal papilla cells facilitated the evaluation of condition for in vitro expansion. World Congr. Hair Res. 2010, 19, 604–605. [Google Scholar]
- Thornton, M.J.; Kato, S.; Hibberts, N.A.; Brinklow, B.R.; Loudon, A.; Randall, V.A. Ability to culture dermal pa-pilla cells from red deer (cervus elaphus) hair follicles with differing hormonal responses in vivo offers a new model for studying the control of hair follicle biology. J. Exp. Zool. 1996, 275, 452. [Google Scholar] [CrossRef]
- Inamatsu, M.; Matsuzaki, T.; Iwanari, H.; Yoshizato, K. Establishment of Rat Dermal Papilla Cell Lines that Sustain the Potency to Induce Hair Follicles from Afollicular Skin. J. Investig. Dermatol. 1998, 111, 767–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, J.; Burgeson, R.E.; Morgan, B.A. Wnt signaling maintains the hair inducing activity of the dermal papilla. Genes Dev. 2000, 14, 1181–1185. [Google Scholar] [PubMed]
- Ouji, Y.; Nakamura-Uchiyama, F.; Yoshikawa, M. Canonical Wnts, specifically Wnt- 10b, show ability to maintain dermal pa-pilla cells. Biochem. Biophys. Res. Commun. 2013, 438, 493–499. [Google Scholar] [CrossRef]
- Qiao, J.; Zawadzka, A.; Philips, E.; Turetsky, A.; Batchelor, S.; Peacock, J.; Durrant, S.; Garlick, D.; Kemp, P.; Teumer, J. Hair follicle neogenesis induced by cultured human scalp dermal papilla cells. Regen. Med. 2009, 4, 667–676. [Google Scholar] [CrossRef]
- Driskell, R.R.; Juneja, V.R.; Connelly, J.T.; Kretzschmar, K.; Tan, D.W.-M.; Watt, F.M. Clonal Growth of Dermal Papilla Cells in Hydrogels Reveals Intrinsic Differences between Sox2-Positive and -Negative Cells In Vitro and In Vivo. J. Investig. Dermatol. 2012, 132, 1084–1093. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Hamazaki, T.S.; Ohnuma, K.; Tamaki, K.; Asashima, M.; Okochi, H. Isolation of Murine Hair-Inducing Cells Using the Cell Surface Marker Prominin-1/CD133. J. Investig. Dermatol. 2007, 127, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Arnold, K.; Sarkar, A.; Yram, M.A.; Polo, J.M.; Bronson, R.; Sengupta, S.; Seandel, M.; Geijsen, N.; Hochedlinger, K. Sox2 + Adult Stem and Progenitor Cells Are Important for Tissue Regeneration and Survival of Mice. Cell Stem Cell 2011, 9, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Rendl, M.; Lewis, L.; Fuchs, E. Molecular Dissection of Mesenchymal-Epithelial Interactions in the Hair Follicle. PLoS Biol. 2005, 3, e331. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Giangreco, A.; Jensen, K.B.; Mulder, K.W.; Watt, F.M. Sox2-positive dermal papilla cells specify hair follicle type in mammalian epidermis. Development 2009, 136, 2815–2823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handjiski, B.; Eichmüller, S.; Hofmann, U.; Czarnetzki, B.; Paus, R. Alkaline phosphatase activity and localization during the murine hair cycle. Br. J. Dermatol. 1994, 131, 303–310. [Google Scholar] [CrossRef]
- Iida, M.; Ihara, S.; Matsuzaki, T. Hair cycle-dependent changes of alkaline phosphatase activity in the mesenchyme and epi-thelium in mouse vibrissal follicles. Dev. Growth Differ. 2010, 49, 185–195. [Google Scholar] [CrossRef]
- Reynolds, A.J.; Jahoda, C.A. Hair matrix germinative epidermal cells confer follicle-inducing capabilities on dermal sheath and high passage papilla cells. Development 1996, 122, 3085–3094. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, N.; Tondreau, T.; Delforge, A.; Dejeneffe, M.; Massy, M.; Libertalis, M. Human marrow mesenchy-mal stem cell culture: Serum-free medium allows better expansion than classical α-MEM medium. Eur. J. Hae-Matol. 2006, 76, 309–316. [Google Scholar] [CrossRef]
- Messenger, A.; Jennifer, H.; Bleehen, S. The in vitro properties of dermal papilla cell lines established from human hair follicles. Br. J. Dermatol. 1986, 114, 425–430. [Google Scholar] [CrossRef]
- Jahoda, C.; Oliver, R. The growth of vibrissa dermal papilla cells in vitro. Br. J. Dermatol. 1981, 105, 623–627. [Google Scholar] [CrossRef]
- Almond-Roesler, B.; Schön, M.; Schön, M.P.; Blume-Peytavi, U.; Sommer, C.; Löster, K.; Orfanos, C.E. Cultured dermal papilla cells of the rat vibrissa follicle. Proliferative activity, adhesion properties and reorganization of the extracellular matrix in vitro. Arch. Dermatol. Res. 1997, 289, 698–704. [Google Scholar] [CrossRef]
- Wessells, N.K.; Roessner, K.D. Nonproliferation in dermal condensations of mouse vibrissae and pelage hairs. Dev. Biol. 1965, 12, 419–433. [Google Scholar] [CrossRef]
- Piérard, G.E.; De La Brassinne, M. Modulation of Dermal Cell Activity During Hair Growth in the Rat. J. Cutan. Pathol. 1975, 2, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Jahoda, C.A.; Reynolds, A.J.; Oliver, R.F. Induction of Hair Growth in Ear Wounds by Cultured Dermal Papilla Cells. J. Investig. Dermatol. 1993, 101, 584–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, T.H.; Lee, C.Y.; Chiu, H.C.; Hsu, C.J.; Lin, S.J. Self-assembly of dermal papilla cells into inductive spheroidal mi-crotissues on poly(ethylene-co-vinyl alcohol) membranes for hair follicle regeneration. Biomaterials 2008, 29, 3521–3530. [Google Scholar] [CrossRef]
- Hamano, T.; Teramoto, A.; Iizuka, E.; Abe, K. Effects of polyelectrolyte complex (PEC) on human periodontal ligament fi-broblast (HPLF) function. I. Three-dimensional structure of HPLF cultured on PEC. J. Biomed. Mater. Res. Part B Appl. Biomater. 1998, 41, 257–269. [Google Scholar] [CrossRef]
- Koide, N.; Sakaguchi, K.; Koide, Y.; Asano, K.; Kawaguchi, M.; Matsushima, H.; Takenami, T.; Shinji, T.; Mori, M.; Tsuji, T. Formation of multicellular spheroids composed of adult rat hepatocytes in dishes with positively charged surfaces and under other nonadherent environments. Exp. Cell Res. 1990, 186, 227–235. [Google Scholar] [CrossRef]
- Ryan, P.L.; Foty, R.A.; Kohn, J.; Steinberg, M.S. Tissue spreading on implantable substrates is a competitiveoutcome of cell–cell vs. cell–substratum adhesivity. Proc. Natl. Acad. Sci. USA 2001, 98, 4323–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, T.-H.; Tu, H.-R.; Chan, C.-C.; Huang, Y.-C.; Yen, M.-H.; Cheng, N.-C.; Chiu, H.-C.; Lin, S.-J. The enhancement of dermal papilla cell aggregation by extracellular matrix proteins through effects on cell–substratum adhesivity and cell motility. Biomaterials 2009, 30, 5031–5040. [Google Scholar] [CrossRef]
- Neelamegham, S.; Munn, L.L.; Zygourakis, K. A model for the kinetics of homotypic cellular aggregation under static conditions-ScienceDirect. Biophys. J. 1997, 72, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Hsiao, W.; Young, T.; Yu, H.; Jee, S.S.J.L. Behavior of Human Melanocytes on Chitosan: From Monolayer to Three-Dimensional Spheroids. Ph.D. Thesis, National Taiwan University, Taipei, China, 2006. [Google Scholar]
- Biggs, L.C.; Mäkelä, O.J.; Myllymäki, S.-M.; Das Roy, R.; Närhi, K.; Pispa, J.; Mustonen, T.; Mikkola, M.L. Hair follicle dermal condensation forms via Fgf20 primed cell cycle exit, cell motility, and aggregation. eLife 2018, 7. [Google Scholar] [CrossRef]
- Hashimoto, K.S. Ultrastructural study on Differentiation and Function of Hair. In Iology and Disease of the Hair Tokyo; Kobori, T., Montagna, W., Eds.; University of Tokyo Press: Tokyo, Japan, 1976; pp. 23–56. [Google Scholar]
- Yamanishi, H.; Soma, T.; Yoshida, Y.; Ishimatsu, Y. Method for Promoting Hair Growth or Hair Regeneration by Maintaining or Increasing Expression of Cell-Adhesion Factor. U.S. Patent No 8,334,136, 18 December 2012. [Google Scholar]
- Jacobson, C.M. A comparative study of the mechanisms by which X-irradiation and genetic mutation cause loss of vibrissae in embryo mice. J. Embryol. Exp. Morphol. 1966, 16, 369–379. [Google Scholar]
- Newgreen, D.; Thiery, J.-P. Fibronectin in early avian embryos: Synthesis and distribution along the migration pathways of neural crest cells. Cell Tissue Res. 1980, 211, 269–291. [Google Scholar] [CrossRef] [PubMed]
- Bronner-Fraser, M. An antibody to a receptor for fibronectin and laminin perturbs cranial neural crest development in vivo. Dev. Biol. 1986, 117, 528–536. [Google Scholar] [CrossRef]
- Mauger, A.; Demarchez, M.; Herbage, D.; Grimaud, J.-A.; Druguet, M.; Hartmann, D.; Sengel, P. Immunofluorescent localization of collagen types I and III, and of fibronectin during feather morphogenesis in the chick embryo. Dev. Biol. 1982, 94, 93–105. [Google Scholar] [CrossRef]
- Holbrook, K.A.; Smith, L.T.; Kaplan, E.D.; Minami, S.A.; Hebert, G.P.; Underwood, R.A. Expression of Morphogens During Human Follicle Development In Vivo and a Model for Studying Follicle Morphogenesis In Vitro. J. Investig. Dermatol. 1993, 101, S39–S49. [Google Scholar] [CrossRef]
- Jiang, T.-X.; Chuong, C.-M. Mechanism of skin morphogenesis. I. Analyses with antibodies to adhesion molecules tenascin, N-CAM, and integrin. Dev. Biol. 1992, 150, 82–98. [Google Scholar] [CrossRef]
- Couchman, J.R.; Gibson, W.T. Expression of basement membrane components through morphological changes in the hair growth cycle. Dev. Biol. 1985, 108, 290–298. [Google Scholar] [CrossRef]
- Kaplan, E.D.; Holbrook, K.A. Dynamic expression patterns of tenascin, proteoglycans, and cell adhesion molecules during human hair follicle morphogenesis. Dev. Dyn. 1994, 199, 141–155. [Google Scholar] [CrossRef]
- Schalkwijk, J.; Steijlen, P.M.; Van Vlijmen-Willems, I.M.; Oosterling, B.; Mackie, E.J.; Verstraeten, A.A. Tenascin expression in human dermis is related to epidermal proliferation. Am. J. Pathol. 1991, 139, 1143–1150. [Google Scholar]
- Chuong, C.M.; Chen, H.M.; Jiang, T.X.; Chia, J. Adhesion Molecules in Skin Development: Morphogenesis of Feather and Hair. Ann. N. Y. Acad. Sci. 1991, 642, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Shames, R.B.; Jennings, A.G.; Sawyer, R.H. The initial expression and patterned appearance of tenascin in scutate scales is absent from the dermis of the scaleless (sc/sc) chicken. Dev. Biol. 1991, 147, 174–186. [Google Scholar] [CrossRef]
- Lightner, V.A.; Gumkowski, F.; Bigner, D.D.; Erickson, H.P. Tenascin/hexabrachion in human skin: Biochemical identification and localization by light and electron microscopy. J. Cell Biol. 1989, 108, 2483–2493. [Google Scholar] [CrossRef] [PubMed]
- Werb, Z. ECM and Cell Surface Proteolysis: Regulating Cellular Ecology. Cell 1997, 91, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Elliott, K.; Messenger, A.G.; Stephenson, T.J. Differences in Hair Follicle Dermal Papilla Volume are Due to Extracellular Matrix Volume and Cell Number: Implications for the Control of Hair Follicle Size and Androgen Responses. J. Investig. Dermatol. 1999, 113, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Botchkarev, V.A.; Kishimoto, J. Molecular Control of Epithelial–Mesenchymal Interactions During Hair Follicle Cycling. J. Investig. Dermatol. Symp. Proc. 2003, 8, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricard-Blum, S.; Ruggiero, F.; Van Der Rest, M. The Collagen Superfamily. Top. Curr. Chem. 2005, 247, 35–84. [Google Scholar] [CrossRef]
- Jahoda, C.A.; Mauger, A.; Bard, S.; Sengel, P. Changes in fibronectin, laminin and type IV collagen distribution relate to basement membrane restructuring during the rat vibrissa follicle hair growth cycle. J. Anat. 1992, 181, 47–60. [Google Scholar]
- Dublet, B.; Rest, M.V.D.; Ricardblum, S. Unconventional Collagens: Types VI, VII, VIII, IX, X, XII, XIV, XVI and XIX; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Gordon, M.K.; Hahn, R.A. Collagens. Cell Tissue Res. 2010, 339, 247. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, E.; Roberts, U.; Corbo, T.; Jacenko, O. Congenic Mice Confirm That Collagen X Is Required for Proper Hematopoietic Development. PLoS ONE 2010, 5, e9518. [Google Scholar] [CrossRef]
- Plumb, D.A.D.; Mironov, A.; Ferrara, L.; Poulsom, R.; Kadler, K.E.; Thornton, D.J.; Briggs, M.D.; Boot-Handford, R.P. Collagen XXVII Is Developmentally Regulated and Forms Thin Fibrillar Structures Distinct from Those of Classical Vertebrate Fibrillar ColI-agens. J. Biol. Chem. 2007, 282, 12791–12795. [Google Scholar] [CrossRef] [Green Version]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [Green Version]
- Myllyharju, J.; Kivirikko, K.I. Collagens, modifying enzymes and their mutations in humans, flies and worms. Trends Genet. 2004, 20, 33–43. [Google Scholar]
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzym. Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, T.K. Physiology and pathophysiology of matrix metalloproteases. Amino Acids 2011, 41, 271–291. [Google Scholar]
- Han, S.; Makareeva, E.; Kuznetsova, N.V.; DeRidder, A.M.; Sutter, M.B.; Losert, W.; Phillips, C.L.; Visse, R.; Nagase, H.; Leikin, S. Molecular Mechanism of Type I Collagen Homotrimer Resistance to Mammalian Collagenases. J. Biol. Chem. 2010, 285, 22276–22281. [Google Scholar] [CrossRef] [Green Version]
- Abu-Abed, S.; Maclean, G.; Fraulob, V.; Chambon, P.; Dollé, P. Differential expression of the retinoic acid-metabolizing en-zymes CYP26A1 and CYP26B1 during murine organogenesis. Mech. Dev. 2020, 110, 173–177. [Google Scholar] [CrossRef]
- Topletz, A.R.; Thatcher, J.E.; Zelter, A.; Lutz, J.D.; Tay, S.; Nelson, W.L.; Isoherranen, N. Comparison of the function and expression of CYP26A1 and CYP26B1, the two retinoic acid hydroxylases. Biochem. Pharmacol. 2012, 83, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Okano, J.; Levy, C.; Lichti, U.; Sun, H.-W.; Yuspa, S.H.; Sakai, Y.; Morasso, M.I. Cutaneous Retinoic Acid Levels Determine Hair Follicle Development and Downgrowth. J. Biol. Chem. 2012, 287, 39304–39315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foitzik, K.; Spexard, T.; Nakamura, M.; Halsner, U.; Paus, R. Towards dissecting the pathogenesis of retinoid-induced hair loss: All-trans retinoic acid induces premature hair follicle regression (catagen) by upregulation of transforming growth fac-tor-beta2 in the dermal papilla. J. Investig. Dermatol. 2005, 124, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Everts, H.B. Endogenous retinoids in the hair follicle and sebaceous gland. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2012, 1821, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Kwack, M.H.; Jang, Y.J.; Won, G.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Overexpression of alkaline phosphatase improves the hair-inductive capacity of cultured human dermal papilla spheres. J. Dermatol. Sci. 2019, 95, 126–129. [Google Scholar] [CrossRef] [Green Version]
- Higgins, C.A.; Richardson, G.D.; Ferdinando, D.; Westgate, G.E.; Jahoda, C.A. Modelling the hair follicle dermal papilla using spheroid cell cultures. Exp. Dermatol. 2010, 19, 546–548. [Google Scholar] [CrossRef] [PubMed]
- Coffinier, C.; Tran, U.; Larraín, J.; De Robertis, E.; De Robertis, E. Neuralin-1 is a novel Chordin-related molecule expressed in the mouse neural plate. Mech. Dev. 2001, 100, 119–122. [Google Scholar] [CrossRef]
- Nakayama, N.; Han, C.-Y.E.; Scully, S.; Nishinakamura, R.; He, C.; Zeni, L.; Yamane, H.; Chang, D.; Yu, D.; Yokota, T.; et al. A Novel Chordin-like Protein Inhibitor for Bone Morphogenetic Proteins Expressed Preferentially in Mesenchymal Cell Lineages. Dev. Biol. 2001, 232, 372–387. [Google Scholar] [CrossRef] [Green Version]
- Balemans, W.; Hul, W.V. Extracellular Regulation of BMP Signaling in Vertebrates: A Cocktail of Modulators. Dev. Biol. 2002, 250, 231–250. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Pachter, L. Streaming fragment assignment for real-time analysis of sequencing experiments. Nat. Methods 2013, 10, 71–73. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. gplots: Various R Programming Tools for Plotting Data. 2020. Available online: https://CRAN.R-project.org/package=gplots (accessed on 15 March 2020).
- Frank, E.; Harrell, E., Jr. Hmisc: Harrell Miscellaneous. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 20 March 2020).
- Taiyun, W.V.S. Corrplot: Visualization of a Correlation Matrix. 2017. Available online: https://CRAN.R-project.org/package=corrplot (accessed on 15 March 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Love, M.I.H.W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nature Protoc. 2009, 4, 44–57. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Gene Symbol | Log2FoldChange ADP vs. MDP | Log2FoldChange MDP vs. UDP |
|---|---|---|---|---|
| Celaphus_00014740 | Alkaline phosphatase, tissue-nonspecific isozyme | ALPL | 2.1 | 4.3 |
| Celaphus_00011342 | Liprin-alpha-2 | Ppfia2 | 2.2 | 2.8 |
| Celaphus_00005746 | Cytochrome P450 26B1 | CYP26B1 | 2.2 | 2.1 |
| Celaphus_00011343 | Liprin-alpha-4 (Fragment) | Ppfia4 | 2.7 | 2.1 |
| Celaphus_00008669 | Disintegrin and metalloproteinase domain-containing protein 8 | ADAM8 | 3.7 | 2.1 |
| Celaphus_00001834 | SLAM family member 8 | SLAMF8 | 2.13 | 2.12 |
| Celaphus_00005340 | Kelch-like protein 29 | KLHL29 | −4.2 | −2.4 |
| Celaphus_00012937 | Solute carrier organic anion transporter family member 2A1 | Slco2a1 | −2.2 | −2.5 |
| Celaphus_00009791 | Chordin-like protein 1 | CHRDL1 | −2.7 | −2.6 |
| Celaphus_00013117 | Insulin-like growth factor II | IGF2 | −5.8 | −3.1 |
| Celaphus_00001878 | Platelet endothelial aggregation receptor 1 | PEAR1 | −3.8 | −3.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, G.; Sun, H.; Wei, H.; Qin, T.; Yang, Y.; Xu, X.; Zhao, S. Detecting the Mechanism behind the Transition from Fixed Two-Dimensional Patterned Sika Deer (Cervus nippon) Dermal Papilla Cells to Three-Dimensional Pattern. Int. J. Mol. Sci. 2021, 22, 4715. https://doi.org/10.3390/ijms22094715
Wei G, Sun H, Wei H, Qin T, Yang Y, Xu X, Zhao S. Detecting the Mechanism behind the Transition from Fixed Two-Dimensional Patterned Sika Deer (Cervus nippon) Dermal Papilla Cells to Three-Dimensional Pattern. International Journal of Molecular Sciences. 2021; 22(9):4715. https://doi.org/10.3390/ijms22094715
Chicago/Turabian StyleWei, Guanning, Hongmei Sun, Haijun Wei, Tao Qin, Yifeng Yang, Xiaohong Xu, and Shoujing Zhao. 2021. "Detecting the Mechanism behind the Transition from Fixed Two-Dimensional Patterned Sika Deer (Cervus nippon) Dermal Papilla Cells to Three-Dimensional Pattern" International Journal of Molecular Sciences 22, no. 9: 4715. https://doi.org/10.3390/ijms22094715