
Expression and Roles of Individual HIF Prolyl 4-Hydroxylase Isoenzymes in the Regulation of the Hypoxia Response Pathway along the Murine Gastrointestinal Epithelium

 , ,
, ,
Abstract
:1. Introduction
2. Results
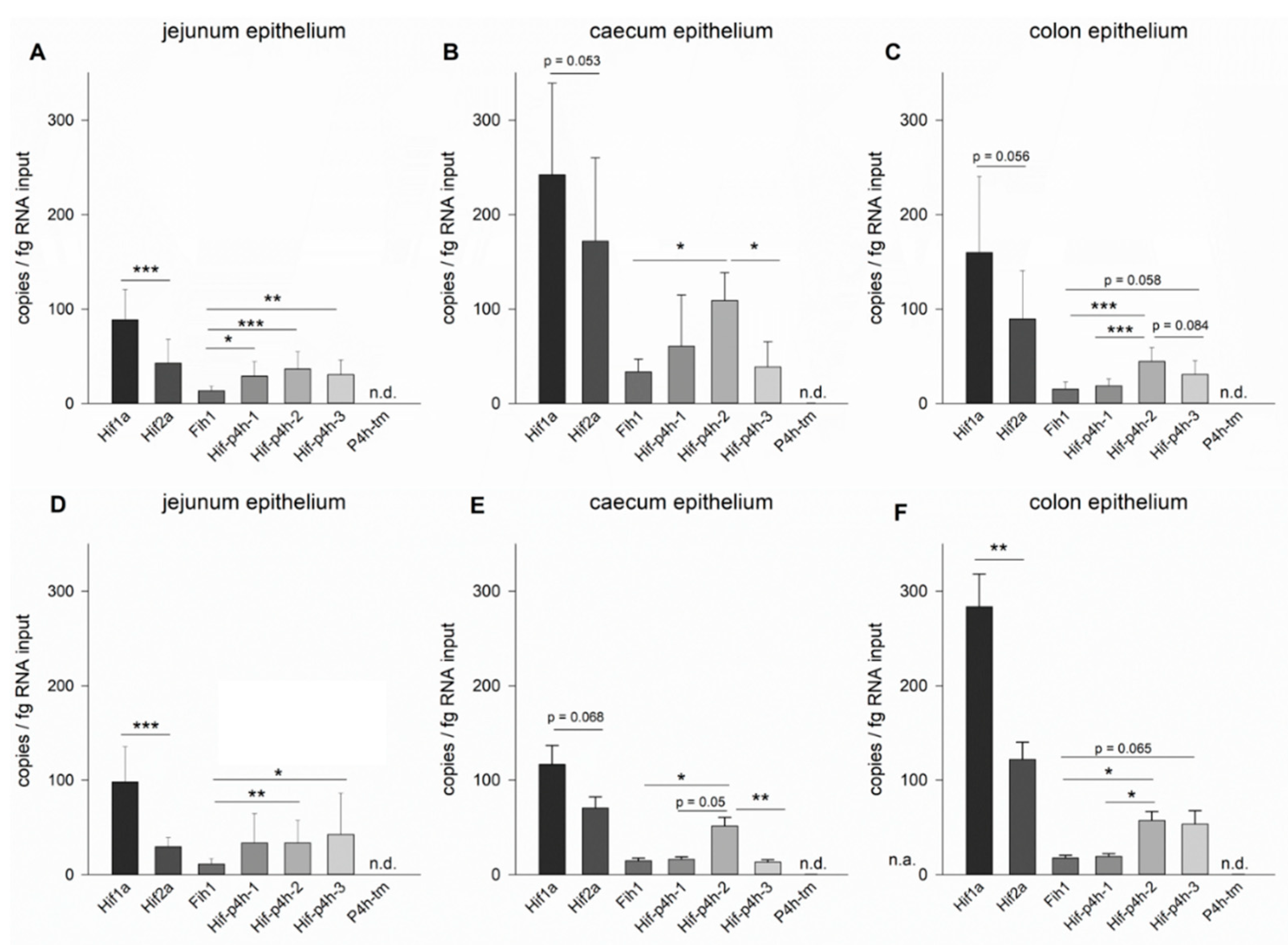
2.1. Expression Profiles of the HIF Pathway Components in Murine Jejunum, Caecum and Colon Epithelium
2.1.1. Jejunum Epithelium
2.1.2. Caecum Epithelium
2.1.3. Colon Epithelium
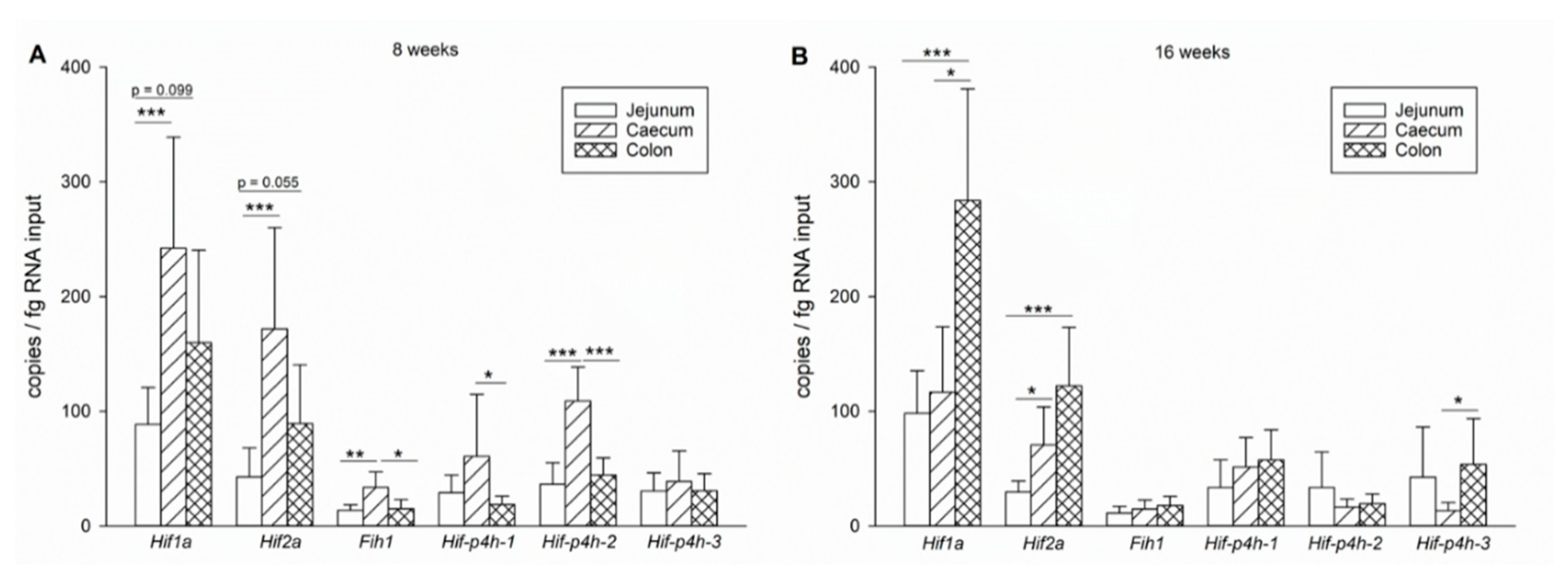
2.1.4. Expression Level Differences between Small and Large Intestines
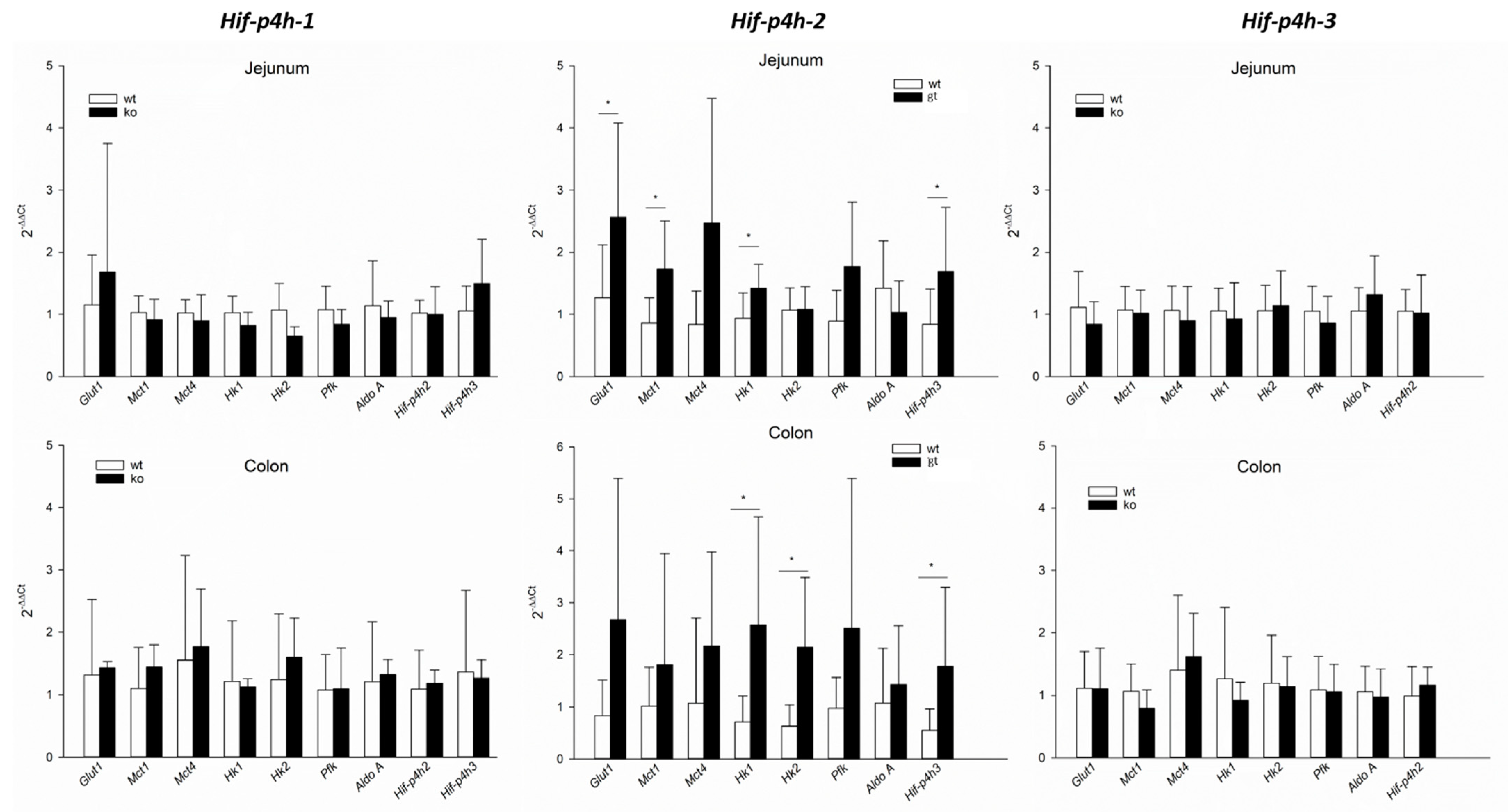
2.2. HIF-P4H-2 Plays a Major Role in the Regulation of Hypoxic Signalling in Murine Jejunum Epithelium
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. mRNA Extraction and PCR Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | 5′–3′ Sequence | 3′–5′ Sequence |
|---|---|---|
| Aldoa | AGTCCACCGGAAGCATTGC | CAGCCCCTGGGTAGTTGTC |
| Actb | AGAGGGAAATCGTGCGTGAC | CAATAGTGATGACCTGGCCGT |
| Glut1 | TCAAACATGGAACCACCGCTA | AAGAGGCCGACAGAGAAGGAA |
| Hif-p4h-2 | GCGTCCCAGTCTTTATTTAGATA | CTGGGCAACTACAGGATAAAC |
| Hif-p4h-3 | GCTGGGCAAATACTATGTCAAG | CTTATTCAGGTAGTAGATACAGGTGATACA |
| Hk1 | GAGTCTGAGGTCTACGACACC | CCCACGGGTAATTTCTTGTCC |
| Hk2 | GTGTGCTCCGAGTAAGGGTG | CAGGCATTCGGCAATGTGG |
| Hprt1 | TCAGTCAACGGGGGACATAAA | GGGGCTGTACTGCTTAACCAG |
| Mct1 | TGTTAGTCGGAGCCTTCATTTC | CACTGGTCGTTGCACTGAATA |
| Mct4 | TCACGGGTTTCTCCTACGC | GCCAAAGCGGTTCACACAC |
| Pfk | TGCAGCCTACAATCTGCTCC | GTCAAGTGTGCGTAGTTCTGA |
| Ppia | GAGCTGTTTGCAGACAAAGTTC | CCCTGGCACATGAATCCTGG |
4.3. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Semenza, G.L. Hypoxia-inducible factors in physiology and medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myllyharju, J.; Koivunen, P. Hypoxia-inducible factor prolyl 4-hydroxylases: Common and specific roles. Biol. Chem. 2013, 394, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G.; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Oxygen sensing, hypoxia-inducible factors, and disease pathophysiology. Annu. Rev. Pathol. 2014, 9, 47–71. [Google Scholar] [CrossRef]
- McNeill, L.A.; Hewitson, K.S.; Claridge, T.D.; Seibel, J.F.; Horsfall, L.E.; Schofield, C.J. Hypoxia-inducible factor asparaginyl hydroxylase (FIH-1) catalyses hydroxylation at the beta-carbon of asparagine-803. Biochem. J. 2002, 367, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Singhal, R.; Shah, Y.M. Oxygen battle in the gut: Hypoxia and hypoxia-inducible factors in metabolic and inflammatory responses in the intestine. J. Biol. Chem. 2020, 295, 10493–10505. [Google Scholar] [CrossRef]
- Taniguchi, C.M.; Miao, Y.R.; Diep, A.N.; Wu, C.; Rankin, E.B.; Atwood, T.F.; Xing, L.; Giaccia, A.J. PHD inhibition mitigates and protects against radiation-induced gastrointestinal toxicity via HIF2. Sci. Transl. Med. 2014, 6, 236ra64. [Google Scholar] [CrossRef] [Green Version]
- Olcina, M.M.; Giaccia, A.J. Reducing radiation-induced gastrointestinal toxicity—The role of the PHD/HIF axis. J. Clin. Investig. 2016, 126, 3708–3715. [Google Scholar] [CrossRef] [Green Version]
- Sanghani, N.S.; Haase, V.H. Hypoxia-Inducible Factor Activators in Renal Anemia: Current Clinical Experience. Adv. Chronic Kidney Dis. 2019, 26, 253–266. [Google Scholar] [CrossRef]
- Dhillon, S. Roxadustat: First Global Approval. Drugs 2019, 79, 563–572. [Google Scholar] [CrossRef]
- Koivunen, P.; Tiainen, P.; Hyvärinen, J.; Williams, K.E.; Sormunen, R.; Klaus, S.J.; Kivirikko, K.I.; Myllyharju, J. An endoplasmic reticulum transmembrane prolyl 4-hydroxylase is induced by hypoxia and acts on hypoxia-inducible factor alpha. J. Biol. Chem. 2007, 282, 30544–30552. [Google Scholar] [CrossRef] [Green Version]
- Aro, E.; Khatri, R.; Gerard-O’Riley, R.; Mangiavini, L.; Myllyharju, J.; Schipani, E. Hypoxia-inducible factor-1 (HIF-1) but not HIF-2 is essential for hypoxic induction of collagen prolyl 4-hydroxylases in primary newborn mouse epiphyseal growth plate chondrocytes. J. Biol. Chem. 2012, 287, 37134–37144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oehme, F.; Ellinghaus, P.; Kolkhof, P.; Smith, T.J.; Ramakrishnan, S.; Hütter, J.; Schramm, M.; Flamme, I. Overexpression of PH-4, a novel putative proline 4-hydroxylase, modulates activity of hypoxia-inducible transcription factors. Biochem. Biophys. Res. Commun. 2002, 296, 343–349. [Google Scholar] [CrossRef]
- Appelhoff, R.J.; Tian, Y.-M.; Raval, R.R.; Turley, H.; Harris, A.L.; Pugh, C.W.; Ratcliffe, P.J.; Gleadle, J.M. Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. J. Biol. Chem. 2004, 279, 38458–38465. [Google Scholar] [CrossRef] [Green Version]
- Willam, C.; Maxwell, P.H.; Nichols, L.; Lygate, C.; Tian, Y.M.; Bernhardt, W.; Wiesener, M.; Ratcliffe, P.J.; Eckardt, K.-U.; Pugh, C.W. HIF prolyl hydroxylases in the rat; organ distribution and changes in expression following hypoxia and coronary artery ligation. J. Mol. Cell. Cardiol. 2006, 41, 68–77. [Google Scholar] [CrossRef]
- Lieb, M.E.; Menzies, K.; Moschella, M.C.; Ni, R.; Taubman, M.B. Mammalian EGLN genes have distinct patterns of mRNA expression and regulation. Biochem. Cell Biol. 2002, 80, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, H.-S.; Fong, G.-H.; Xi, Q.-L.; Wu, G.-H.; Bai, C.-G.; Ling, Z.-Q.; Fan, L.; Xu, Y.-M.; Qin, Y.-Q.; et al. PHD3 Stabilizes the Tight Junction Protein Occludin and Protects Intestinal Epithelial Barrier Function. J. Biol. Chem. 2015, 290, 20580–20589. [Google Scholar] [CrossRef] [Green Version]
- Kennel, K.B.; Burmeister, J.; Schneider, M.; Taylor, C.T. The PHD1 oxygen sensor in health and disease. J. Physiol. 2018, 596, 3899–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soilleux, E.J.; Turley, H.; Tian, Y.M.; Pugh, C.W.; Gatter, K.C.; Harris, A.L. Use of novel monoclonal antibodies to determine the expression and distribution of the hypoxia regulatory factors PHD-1, PHD-2, PHD-3 and FIH in normal and neoplastic human tissues. Histopathology 2005, 47, 602–610. [Google Scholar] [CrossRef]
- Leinonen, H.; Koivisto, H.; Lipponen, H.-R.; Matilainen, A.; Salo, A.M.; Dimova, E.Y.; Hämäläinen, E.; Stavén, S.; Miettinen, P.; Myllyharju, J.; et al. Null mutation in P4h-tm leads to decreased fear and anxiety and increased social behavior in mice. Neuropharmacology 2019, 153, 63–72. [Google Scholar] [CrossRef]
- Byts, N.; Sharma, S.; Laurila, J.; Paudel, P.; Miinalainen, I.; Ronkainen, V.-P.; Hinttala, R.; Törnquist, K.; Koivunen, P.; Myllyharju, J. Transmembrane Prolyl 4-Hydroxylase is a Novel Regulator of Calcium Signaling in Astrocytes. eNeuro 2021, 8. [Google Scholar] [CrossRef]
- Ward, J.B.J.; Keely, S.J.; Keely, S.J. Oxygen in the regulation of intestinal epithelial transport. J. Physiol. 2014, 592, 2473–2489. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.; Keely, S.; Karhausen, J.; Gerich, M.E.; Furuta, G.T.; Colgan, S.P. Mucosal protection by hypoxia-inducible factor prolyl hydroxylase inhibition. Gastroenterology 2008, 134, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, A.C.; Gleadle, J.M.; McNeill, L.A.; Hewitson, K.S.; O’Rourke, J.; Mole, D.R.; Mukherji, M.; Metzen, E.; Wilson, M.I.; Dhanda, A.; et al. C. elegans EGL-9 and Mammalian Homologs Define a Family of Dioxygenases that Regulate HIF by Prolyl Hydroxylation. Cell 2001, 107, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Van Welden, S.; Selfridge, A.C.; Hindryckx, P. Intestinal hypoxia and hypoxia-induced signalling as therapeutic targets for IBD. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 596–611. [Google Scholar] [CrossRef]
- Tambuwala, M.M.; Cummins, E.P.; Lenihan, C.R.; Kiss, J.; Stauch, M.; Scholz, C.C.; Fraisl, P.; Lasitschka, F.; Mollenhauer, M.; Saunders, S.P.; et al. Loss of prolyl hydroxylase-1 protects against colitis through reduced epithelial cell apoptosis and increased barrier function. Gastroenterology 2010, 139, 2093–2101. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.; Rowan, C.; Strowitzki, M.J.; Fagundes, R.R.; Faber, K.N.; Güntsch, A.; Halligan, D.N.; Kugler, J.; Jones, F.; Lee, C.T.; et al. Mucosal inflammation downregulates PHD1 expression promoting a barrier-protective HIF-1α response in ulcerative colitis patients. FASEB J. 2020, 34, 3732–3742. [Google Scholar] [CrossRef] [Green Version]
- Van Welden, S.; de Vos, M.; Wielockx, B.; Tavernier, S.J.; Dullaers, M.; Neyt, S.; Descamps, B.; Devisscher, L.; Devriese, S.; van den Bossche, L.; et al. Haematopoietic prolyl hydroxylase-1 deficiency promotes M2 macrophage polarization and is both necessary and sufficient to protect against experimental colitis. J. Pathol. 2017, 241, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.; Taylor, C.T. Hypoxia-sensitive pathways in intestinal inflammation. J. Physiol. 2017, 596, 2985–2989. [Google Scholar] [CrossRef] [Green Version]
- Hyvärinen, J.; Hassinen, I.E.; Sormunen, R.; Mäki, J.M.; Kivirikko, K.I.; Koivunen, P.; Myllyharju, J. Hearts of hypoxia-inducible factor prolyl 4-hydroxylase-2 hypomorphic mice show protection against acute ischemia-reperfusion injury. J. Biol. Chem. 2010, 285, 13646–13657. [Google Scholar] [CrossRef] [Green Version]
- Ullah, K.; Rosendahl, A.-H.; Izzi, V.; Bergmann, U.; Pihlajaniemi, T.; Mäki, J.M.; Myllyharju, J. Hypoxia-inducible factor prolyl-4-hydroxylase-1 is a convergent point in the reciprocal negative regulation of NF-κB and p53 signaling pathways. Sci. Rep. 2017, 7, 17220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Kelly, C.J.; Colgan, S.P. Physiologic hypoxia and oxygen homeostasis in the healthy intestine. A Review in the Theme: Cellular Responses to Hypoxia. Am. J. Physiol. Cell Physiol. 2015, 309, C350–C360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.; Shankar, R.A.; Chzhan, M.; Samouilov, A.; Kuppusamy, P.; Zweier, J.L. Noninvasive measurement of anatomic structure and intraluminal oxygenation in the gastrointestinal tract of living mice with spatial and spectral EPR imaging. Proc. Natl. Acad. Sci. USA 1999, 96, 4586–4591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, S.; Roy, S.; Maurer, M.; Ratan, R.R.; Sen, C.K. Oxygen-sensitive reset of hypoxia-inducible factor transactivation response: Prolyl hydroxylases tune the biological normoxic set point. Free Radic. Biol. Med. 2006, 40, 2147–2154. [Google Scholar] [CrossRef] [Green Version]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Dengler, F.; Kraetzig, A.; Gäbel, G. Butyrate Protects Porcine Colon Epithelium from Hypoxia-Induced Damage on a Functional Level. Nutrients 2021, 13, 305. [Google Scholar] [CrossRef]
- Ruel, J.; Ruane, D.; Mehandru, S.; Gower-Rousseau, C.; Colombel, J.-F. IBD across the age spectrum: Is it the same disease? Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 88–98. [Google Scholar] [CrossRef]
- Laitala, A.; Aro, E.; Walkinshaw, G.; Mäki, J.M.; Rossi, M.; Heikkilä, M.; Savolainen, E.-R.; Arend, M.; Kivirikko, K.I.; Koivunen, P.; et al. Transmembrane prolyl 4-hydroxylase is a fourth prolyl 4-hydroxylase regulating EPO production and erythropoiesis. Blood 2012, 120, 3336–3344. [Google Scholar] [CrossRef] [Green Version]
- van Welden, S.; Laukens, D.; Ferdinande, L.; de Vos, M.; Hindryckx, P. Differential expression of prolyl hydroxylase 1 in patients with ulcerative colitis versus patients with Crohn’s disease/infectious colitis and healthy controls. J. Inflamm. (Lond.) 2013, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.P.; Seeballuck, F.; Keely, S.J.; Mangan, N.E.; Callanan, J.J.; Fallon, P.G.; Taylor, C.T. The hydroxylase inhibitor dimethyloxalylglycine is protective in a murine model of colitis. Gastroenterology 2008, 134, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Metzen, E.; Berchner-Pfannschmidt, U.; Stengel, P.; Marxsen, J.H.; Stolze, I.; Klinger, M.; Huang, W.Q.; Wotzlaw, C.; Hellwig-Bürgel, T.; Jelkmann, W.; et al. Intracellular localisation of human HIF-1 alpha hydroxylases: Implications for oxygen sensing. J. Cell Sci. 2003, 116, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.P.; Berra, E.; Comerford, K.M.; Ginouves, A.; Fitzgerald, K.T.; Seeballuck, F.; Godson, C.; Nielsen, J.E.; Moynagh, P.; Pouyssegur, J.; et al. Prolyl hydroxylase-1 negatively regulates IkappaB kinase-beta, giving insight into hypoxia-induced NFkappaB activity. Proc. Natl. Acad. Sci. USA 2006, 103, 18154–18159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockman, M.E.; Lippl, K.; Tian, Y.-M.; Pegg, H.B.; Figg, W.D.; Abboud, M.I.; Heilig, R.; Fischer, R.; Myllyharju, J.; Schofield, C.J.; et al. Lack of activity of recombinant HIF prolyl hydroxylases (PHDs) on reported non-HIF substrates. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Mähler Convenor, M.; Berard, M.; Feinstein, R.; Gallagher, A.; Illgen-Wilcke, B.; Pritchett-Corning, K.; Raspa, M. FELASA recommendations for the health monitoring of mouse, rat, hamster, guinea pig and rabbit colonies in breeding and experimental units. Lab. Anim. 2014, 48, 178–192. [Google Scholar] [CrossRef] [PubMed]




| Gene Name | Order No. |
|---|---|
| Hif1a | Mm00468869_m1 |
| Hif2a | Mm01236112_m1 |
| Fih1 | Mm01198376_m1 |
| Hif-p4h-1 | Mm00519067_m1 |
| Hif-p4h-2 | Mm00459770_m1 |
| Hif-p4h-3 | Mm00472200_m1 |
| Hif-p4h-tm | Mm00512331_m1 |
| Sglt1 | Mm00451203_m1 |
| Vim | Mm01333430_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dengler, F.; Sova, S.; Salo, A.M.; Mäki, J.M.; Koivunen, P.; Myllyharju, J. Expression and Roles of Individual HIF Prolyl 4-Hydroxylase Isoenzymes in the Regulation of the Hypoxia Response Pathway along the Murine Gastrointestinal Epithelium. Int. J. Mol. Sci. 2021, 22, 4038. https://doi.org/10.3390/ijms22084038
Dengler F, Sova S, Salo AM, Mäki JM, Koivunen P, Myllyharju J. Expression and Roles of Individual HIF Prolyl 4-Hydroxylase Isoenzymes in the Regulation of the Hypoxia Response Pathway along the Murine Gastrointestinal Epithelium. International Journal of Molecular Sciences. 2021; 22(8):4038. https://doi.org/10.3390/ijms22084038
Chicago/Turabian StyleDengler, Franziska, Sofia Sova, Antti M. Salo, Joni M. Mäki, Peppi Koivunen, and Johanna Myllyharju. 2021. "Expression and Roles of Individual HIF Prolyl 4-Hydroxylase Isoenzymes in the Regulation of the Hypoxia Response Pathway along the Murine Gastrointestinal Epithelium" International Journal of Molecular Sciences 22, no. 8: 4038. https://doi.org/10.3390/ijms22084038